
Identification of α-Glucosidase Inhibitors from Leaf Extract of Pepper (Capsicum spp.) through Metabolomic Analysis

 ,
,  and
and 
Abstract
:
1. Introduction
2. Results and Discussion
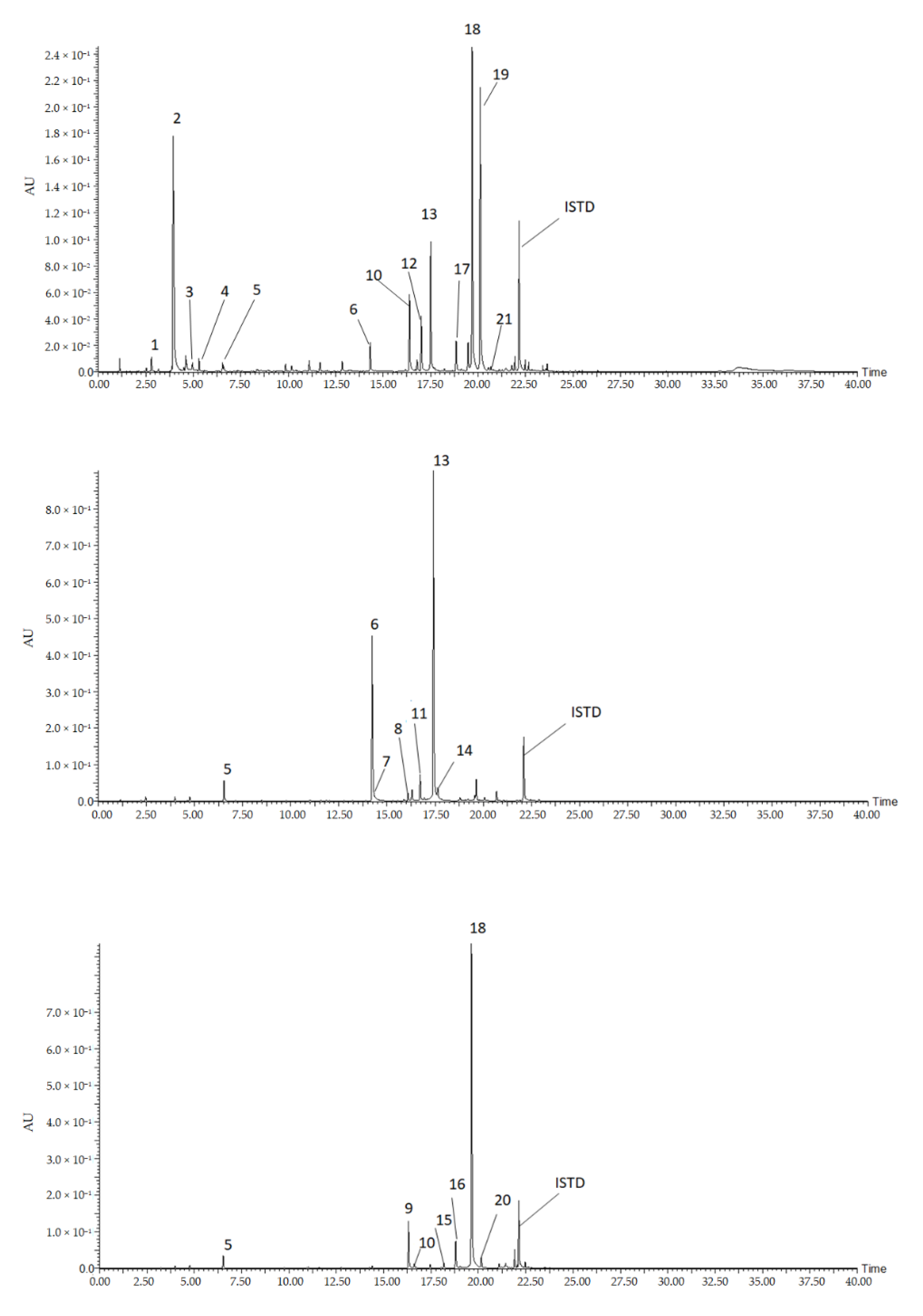
2.1. Identification of Polyphenols from Pepper Leaves Extracts
2.1.1. Hydroxycinnamic Acid Conjugates
2.1.2. Flavones
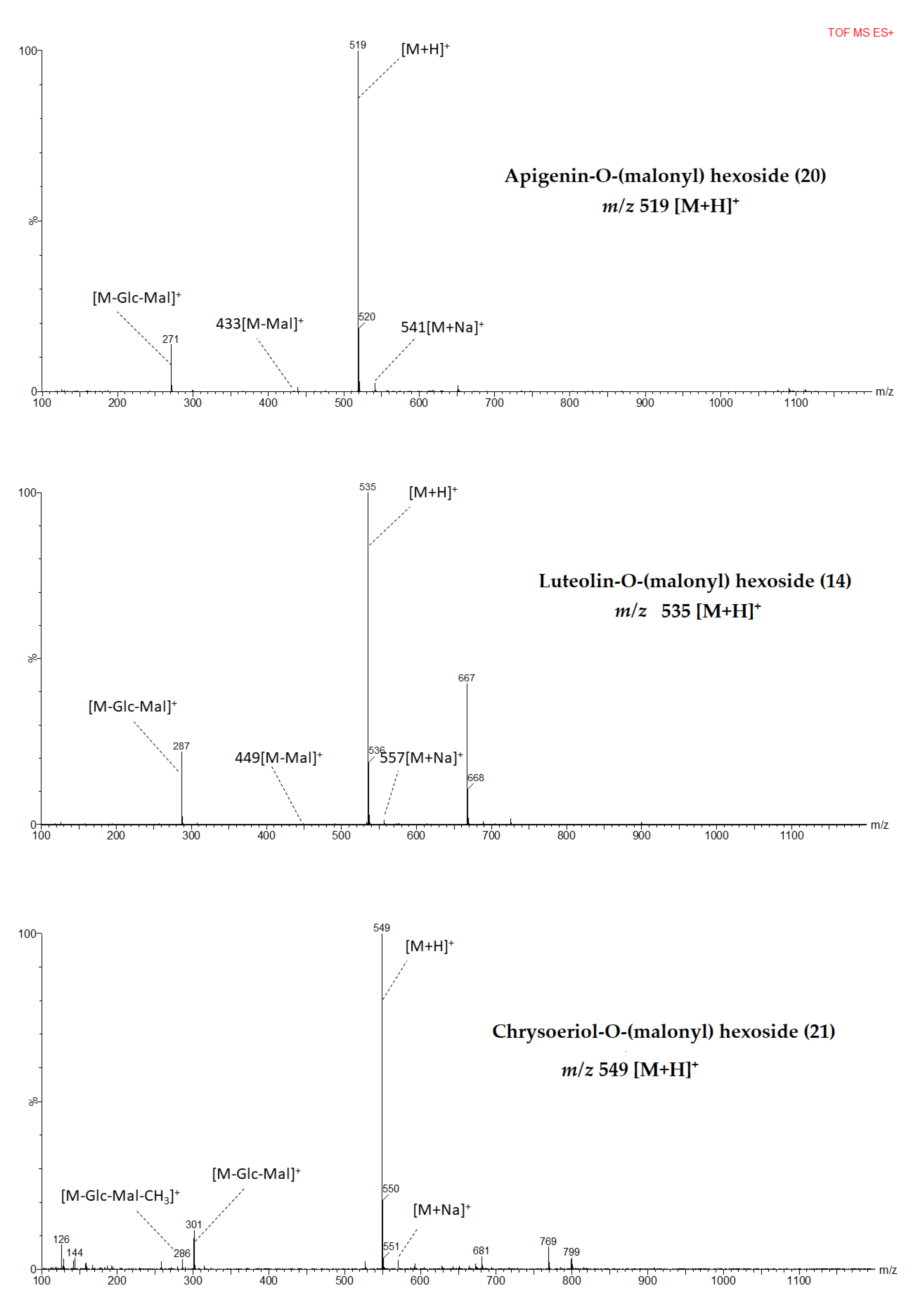
Identification of Luteolin Derivatives
Identification of Apigenin Derivatives
Identification of Chrysoeriol Derivatives
2.2. Flavones Decorations/Modifications
2.3. Relationship between AGI Activity and Metabolite Profile of Pepper Leaves
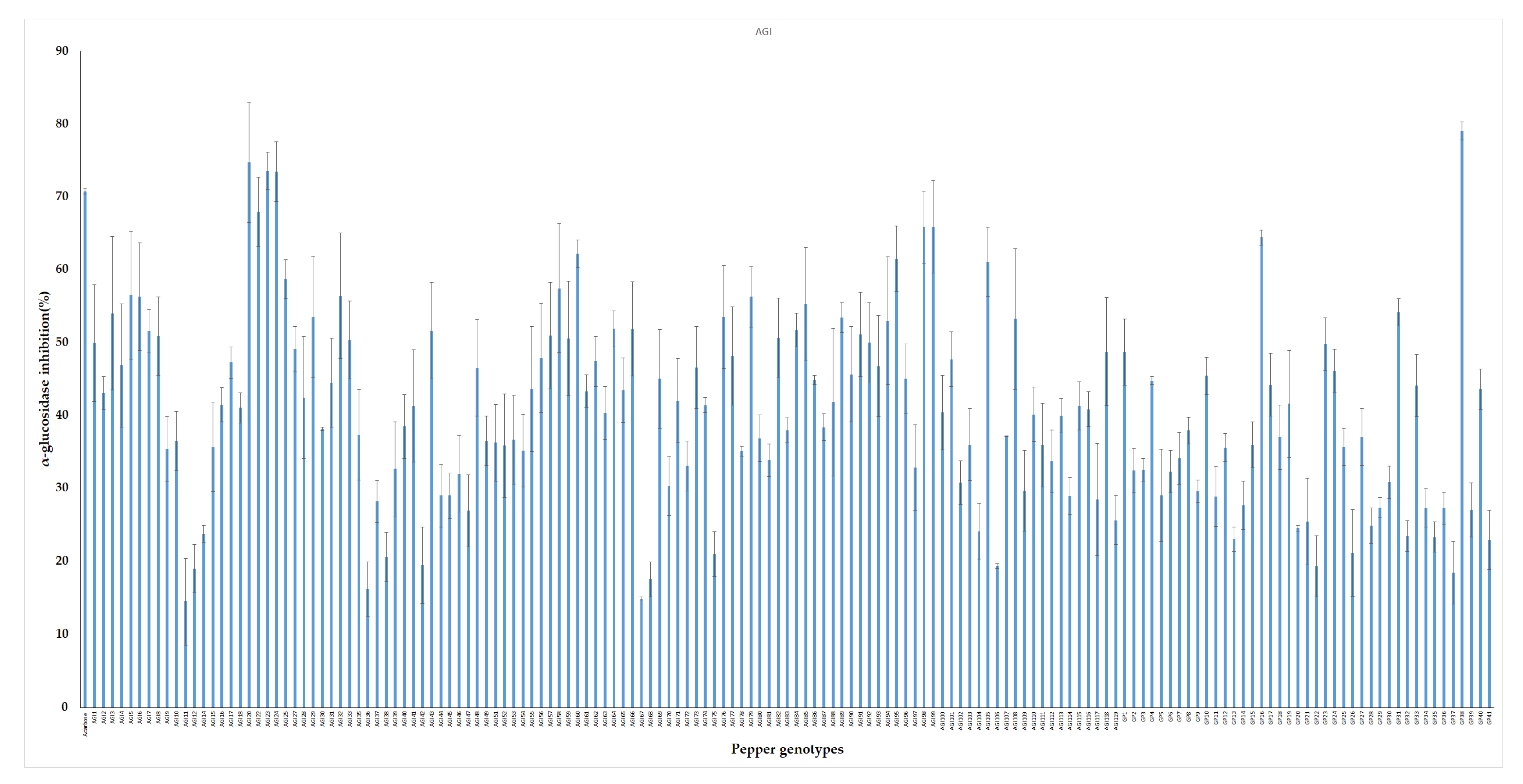
2.3.1. Evaluation of AGI Activity of the Pepper Leaf Extracts
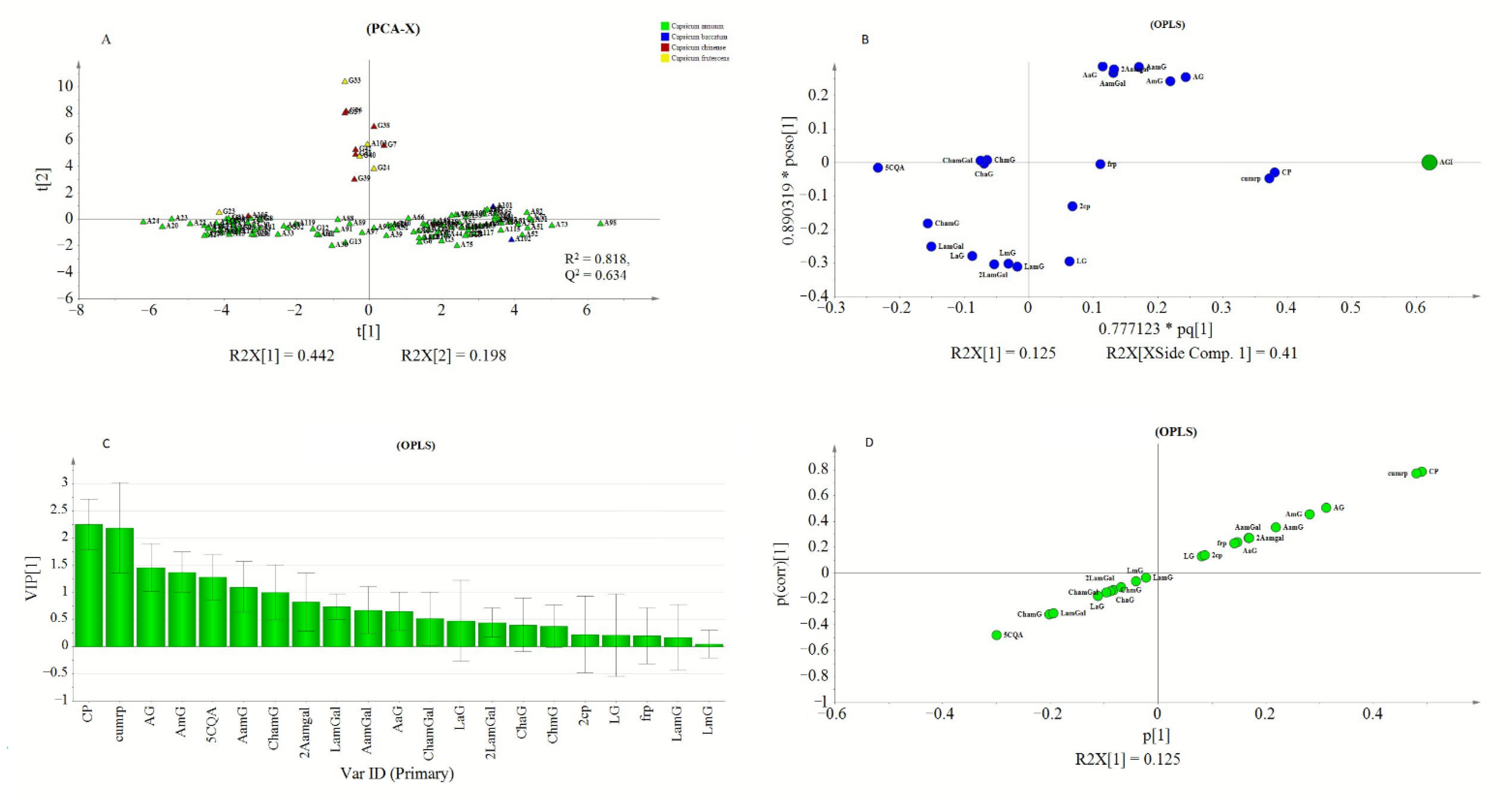
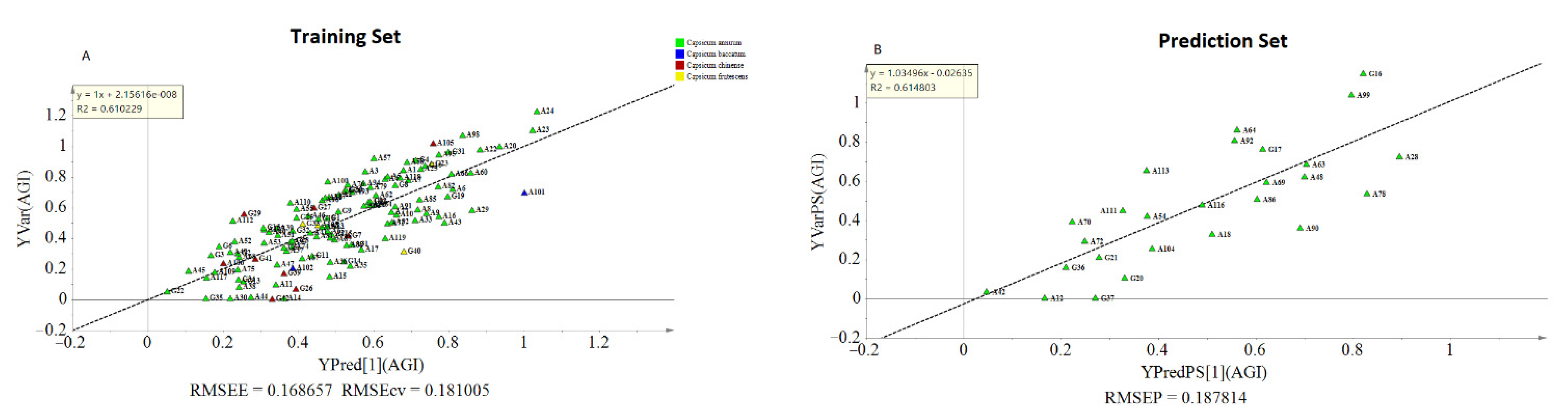
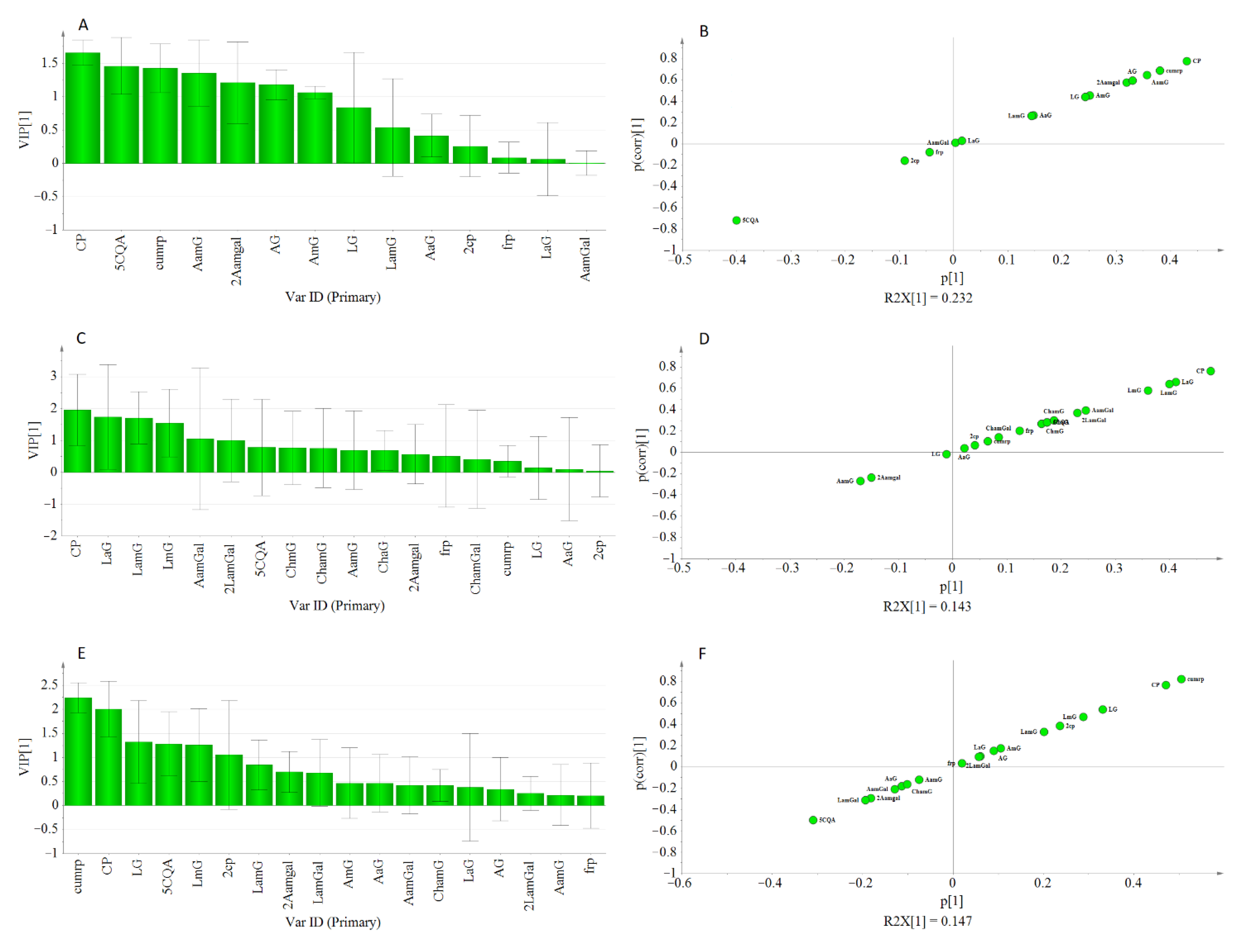
2.3.2. Multivariate Analysis
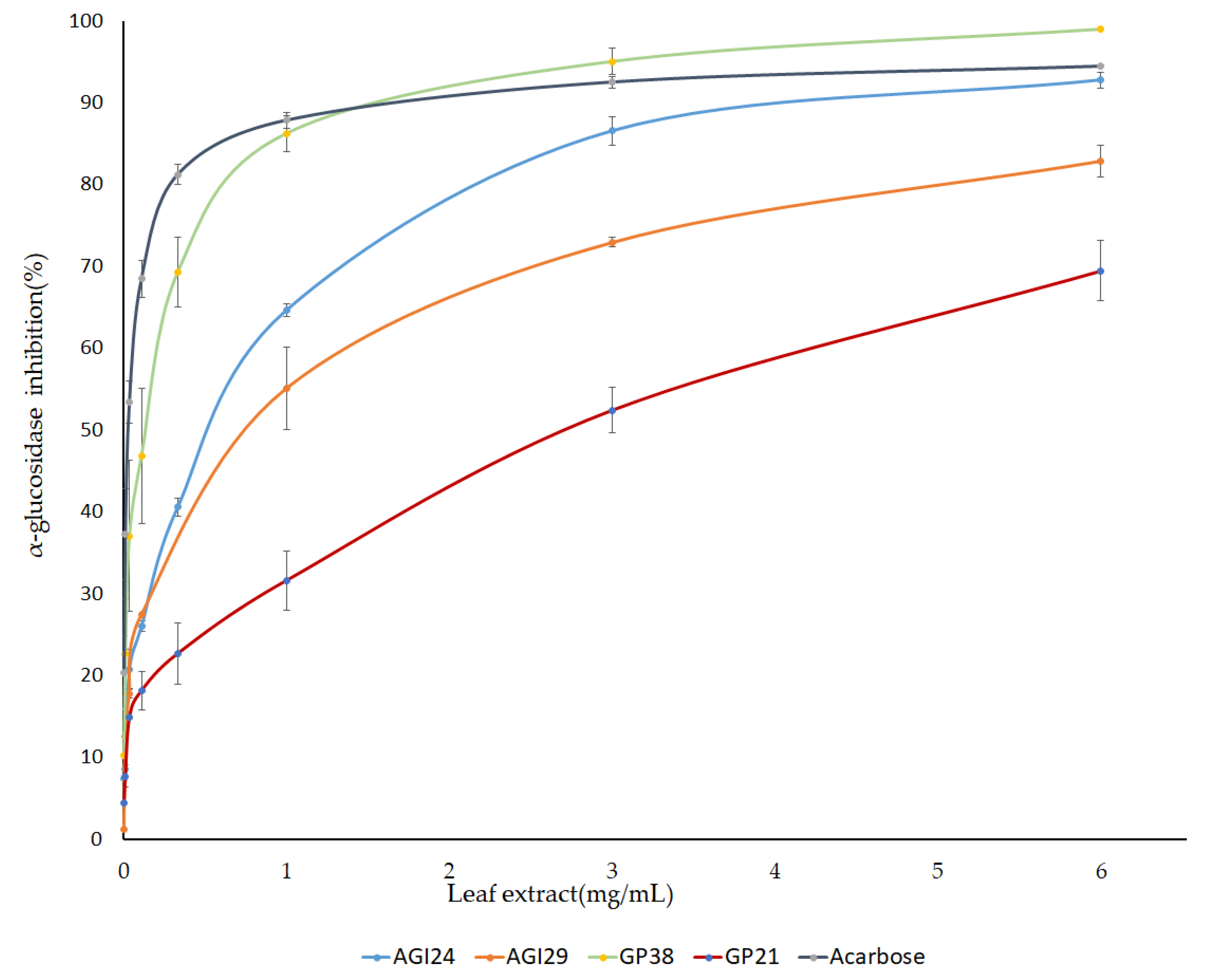
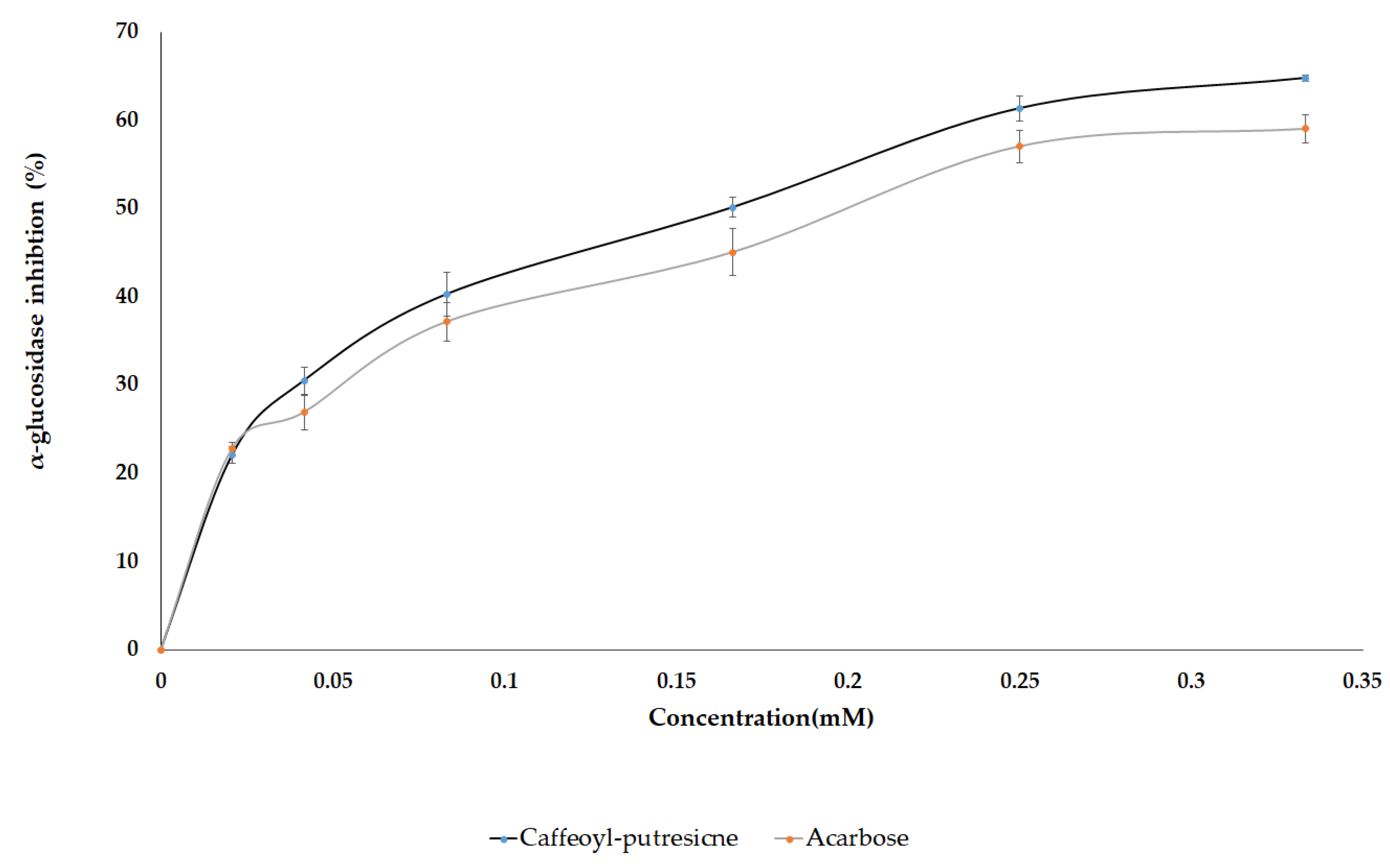
2.4. AGI Activity from Pure Standard Compounds
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Pepper Plant Materials
3.3. Preparation of Leaf Extracts
3.4. Solid-Phase Extraction (SPE)
3.5. Identification of Phenolic Compounds Using UPLC-DAD-QToF-MS
3.6. Yeast α-Glucosidase Inhibition (AGI) Assay
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 5CQA | 5 caffeoylquinic acid |
| 2cp | N-cis Caffeoyl putrescine |
| CP | N-tans Caffeoyl putrescine |
| cumrp | Coumaroyl putrescine |
| frp | Feruloyl putrescine |
| AaG | Apigenin 7-O-(2”-O-apiosyl) glucoside |
| AamGal | Apigenin-O-(apiosyl malonyl) hexoside |
| 2AamGal | Apigenin-O-(apiosyl malonyl) hexoside |
| AamG | Apigenin 7-O-(2”-O-apiosyl-6”-O-malonyl) glucoside |
| AmG | Apigenin-O-(malonyl) hexoside |
| AG | Apigenin 7-O-glucoside |
| ChmG | Chrysoeriol-O-(malonyl) hexoside |
| ChaG | Chrysoeriol-O-(apiosyl) hexoside |
| ChaGal | Chrysoeriol-O-(apiosyl malonyl) hexoside |
| ChamG | Chrysoeriol-O-(apiosyl malonyl) hexoside |
| LaG | Luteolin 7-O-(2”-O-apiosyl) glucoside |
| LamGal | Luteolin-O-(apiosyl malonyl) hexoside |
| 2LamGal | Luteolin-O-(apiosyl malonyl) hexoside |
| LamG | Luteolin 7-O-(2”-O-apiosyl-6”-O-malonyl) glucoside |
| LmG | Luteolin-O-(malonyl) hexoside |
| LG | Luteolin 7-O-glucoside |
| AU | Absorbance unit |
References
- Reinehr, T. Type 1 diabetes mellitus in children and adolescents. World J. Diabetes 2013, 4, 270–281. [Google Scholar] [CrossRef]
- Lankatillake, C.; Huynh, T.; Dias, D.A. Understanding glycaemic control and current approaches for screening antidiabetic natural products from evidence-based medicinal plants. Plant Methods 2019, 15, 1–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J.; Kai, G.; Yamamoto, K.; Chen, X. Advance in Dietary Polyphenols as α-Glucosidases Inhibitors: A Review on Structure-Activity Relationship Aspect. Crit. Rev. Food Sci. Nutr. 2013, 53, 818–836. [Google Scholar] [CrossRef] [PubMed]
- Hakamata, W.; Kurihara, M.; Okuda, H.; Nishio, T.; Oku, T. Design and Screening Strategies for α-Glucosidase Inhibitors Based on Enzymological Information. Curr. Top. Med. Chem. 2009, 9, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Pyner, A.; Nyambe-Silavwe, H.; Williamson, G. Inhibition of human and rat sucrase and maltase activities to assess antiglycemic potential: Optimization of the assay using acarbose and polyphenols. J. Agric. Food Chem. 2017, 65, 8643–8651. [Google Scholar] [CrossRef] [PubMed]
- Adefegha, S.A.; Oboh, G. Inhibition of key enzymes linked to type 2 diabetes and sodium nitroprusside-induced lipid peroxidation in rat pancreas by water extractable phytochemicals from some tropical spices. Pharm. Biol. 2012, 50, 857–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mumtaz, M.W.; Al-Zuaidy, M.H.; Hamid, A.A.; Danish, M.; Akhtar, M.T.; Mukhtar, H. Metabolite profiling and inhibitory properties of leaf extracts of Ficus benjamina towards α-glucosidase and α-amylase. Int. J. Food Prop. 2018, 21, 1560–1574. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Sun, S.; Yang, F.; Zhou, K. UHPLC/MS Identifying Potent α-glucosidase Inhibitors of Grape Pomace via Enzyme Immobilized Method. J. Food Sci. 2018, 83, 1131–1139. [Google Scholar] [CrossRef]
- Kongstad, K.T.; Özdemir, C.; Barzak, A.; Wubshet, S.G.; Staerk, D. Combined use of high-resolution α-glucosidase inhibition profiling and high-performance liquid chromatography-high-resolution mass spectrometry-solid-phase extraction-nuclear magnetic resonance spectroscopy for investigation of antidiabetic principles in crude plant extracts. J. Agric. Food Chem. 2015, 63, 2257–2263. [Google Scholar]
- Yuliana, N.D.; Khatib, A.; Choi, Y.H.; Verpoorte, R. Metabolomics for bioactivity assessment of natural products. Phyther. Res. 2011, 25, 157–169. [Google Scholar] [CrossRef]
- Wahyuni, Y.; Ballester, A.R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Metabolite biodiversity in pepper (Capsicum) fruits of thirty-two diverse accessions: Variation in health-related compounds and implications for breeding. Phytochemistry 2011, 72, 1358–1370. [Google Scholar] [CrossRef]
- Lemos, V.C.; Reimer, J.J.; Wormit, A. Color for life: Biosynthesis and distribution of phenolic compounds in pepper (Capsicum annuum). Agriculture 2019, 9, 81. [Google Scholar] [CrossRef] [Green Version]
- Wahyuni, Y.; Ballester, A.R.; Tikunov, Y.; de Vos, R.C.H.; Pelgrom, K.T.B.; Maharijaya, A.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Metabolomics and molecular marker analysis to explore pepper (Capsicum sp.) biodiversity. Metabolomics 2013, 9, 130–144. [Google Scholar] [CrossRef] [Green Version]
- Materska, M.; Konopacka, M.; Rogoliński, J.; Ślosarek, K. Antioxidant activity and protective effects against oxidative damage of human cells induced by X-radiation of phenolic glycosides isolated from pepper fruits Capsicum annuum L. Food Chem. 2015, 168, 546–553. [Google Scholar] [CrossRef]
- Materska, M.; Perucka, I. Antioxidant activity of the main phenolic compounds isolated from hot pepper fruit (Capsicum annuum L.). J. Agric. Food Chem. 2005, 53, 1750–1756. [Google Scholar] [CrossRef]
- de Sá Mendes, N.; Branco de Andrade Gonçalves, É.C. The role of bioactive components found in peppers. Trends Food Sci. Technol. 2020, 99, 229–243. [Google Scholar] [CrossRef]
- Kim, W.R.; Kim, E.O.; Kang, K.; Oidovsambuu, S.; Jung, S.H.; Kim, B.S.; Nho, C.W.; Um, B.H. Antioxidant activity of phenolics in leaves of three red pepper (Capsicum annuum) cultivars. J. Agric. Food Chem. 2014, 62, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Topuz, A.; Ozdemir, F. Assessment of carotenoids, capsaicinoids and ascorbic acid composition of some selected pepper cultivars (Capsicum annuum L.) grown in Turkey. J. Food Compos. Anal. 2007, 20, 596–602. [Google Scholar] [CrossRef]
- Tilahun, S.; Paramaguru, P.; Rajamani, K. Capsaicin and Ascorbic Acid Variability in Chilli and Paprika As Revealed By Hplc Analysis. J. Plant Breed. Genet. 2013, 1, 85–89. [Google Scholar]
- Wahyuni, Y.; Ballester, A.R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Secondary metabolites of Capsicum species and their importance in the human diet. J. Nat. Prod. 2013, 76, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Maharijaya, A.; Vosman, B.; Pelgrom, K.; Wahyuni, Y.; de Vos, R.C.H.; Voorrips, R.E. Genetic variation in phytochemicals in leaves of pepper (Capsicum) in relation to thrips resistance. Arthropod. Plant Interact. 2019, 13, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Jin, C.H. Inhibitory activity of flavonoids, chrysoeriol and luteolin-7-o-glucopyranoside, on soluble epoxide hydrolase from Capsicum chinense. Biomolecules 2020, 10, 180. [Google Scholar] [CrossRef] [Green Version]
- Park, M.S.; Zhu, Y.X.; Pae, H.O.; Park, S.H. In Vitro and In Vivo α-Glucosidase and α-Amylase Inhibitory Effects of the Water Extract of Leaves of Pepper (Capcicum nnuum L. Cultivar Dangjo) and the Active Constituent Luteolin 7-O-Glucoside. J. Food Biochem. 2016, 40, 696–703. [Google Scholar] [CrossRef]
- Kim, M.-S.; Jin, J.B.; Lee, J.H.; An, H.S.; Pan, C.-H.; Park, J.-S. Rapid separation of Capsicum annuum L. leaf extract using automated HPLC/SPE/HPLC coupling system (Sepbox system) and identification of α-glucosidase inhibitory active substances. J. Appl. Biol. Chem. 2021, 64, 25–32. [Google Scholar] [CrossRef]
- Park, S.; Jeong, W.Y.; Lee, J.H.; Kim, Y.H.; Jeong, S.W.; Kim, G.S.; Bae, D.W.; Lim, C.S.; Jin, J.S.; Lee, S.J.; et al. Determination of polyphenol levels variation in Capsicum annuum L. cv. Chelsea (yellow bell pepper) infected by anthracnose (Colletotrichum gloeosporioides) using liquid chromatography-tandem mass spectrometry. Food Chem. 2012, 130, 981–985. [Google Scholar] [CrossRef]
- Akimoto, N.; Ara, T.; Nakajima, D.; Suda, K.; Ikeda, C.; Takahashi, S.; Muneto, R.; Yamada, M.; Suzuki, H.; Shibata, D.; et al. Flavonoid Search: A system for comprehensive flavonoid annotation by mass spectrometry. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Fang, Y.; Huang, M.J.; Xiao, Y.; Liu, Y.; Ma, X.R.; Zhao, H. High flavonoid accompanied with high starch accumulation triggered by nutrient starvation in bioenergy crop duckweed (Landoltia punctata). BMC Genom. 2017, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhao, C.; Zhao, X.; Xia, Y.; Sun, X.; Xie, W.; Ye, Y.; Lu, X.; Xu, G. Deep Annotation of Hydroxycinnamic Acid Amides in Plants Based on Ultra-High-Performance Liquid Chromatography-High-Resolution Mass Spectrometry and Its in Silico Database. Anal. Chem. 2018, 90, 14321–14330. [Google Scholar] [CrossRef] [Green Version]
- Taofiq, O.; González-Paramás, A.M.; Barreiro, M.F.; Ferreira, I.C.F.R.; McPhee, D.J. Hydroxycinnamic acids and their derivatives: Cosmeceutical significance, challenges and future perspectives, a review. Molecules 2017, 22, 281. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Dunshea, F.R.; Suleria, H.A.R. LC-ESI-QTOF/MS characterization of phenolic compounds from medicinal plants (Hops and Juniper Berries) and their antioxidant activity. Foods 2020, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Gao, Y.; Chen, W.; Wang, W.; Gong, L.; Liu, X.; Luo, J. Spatiotemporal distribution of phenolamides and the genetics of natural variation of hydroxycinnamoyl spermidine in rice. Mol. Plant 2015, 8, 111–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wianowska, D.; Gil, M. Recent advances in extraction and analysis procedures of natural chlorogenic acids. Phytochem. Rev. 2019, 18, 273–302. [Google Scholar] [CrossRef] [Green Version]
- Mudrić, S.; Gašić, U.M.; Dramićanin, A.M.; Ćirić, I.; Milojković-Opsenica, D.M.; Popović-Đorđević, J.B.; Momirović, N.M.; Tešić, Ž.L. The polyphenolics and carbohydrates as indicators of botanical and geographical origin of Serbian autochthonous clones of red spice paprika. Food Chem. 2017, 217, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Asamenew, G.; Kim, H.W.; Lee, M.K.; Lee, S.H.; Lee, S.; Cha, Y.S.; Lee, S.H.; Yoo, S.M.; Kim, J.B. Comprehensive characterization of hydroxycinnamoyl derivatives in green and roasted coffee beans: A new group of methyl hydroxycinnamoyl quinate. Food Chem. X 2019, 2, 100033. [Google Scholar] [CrossRef]
- Martins, N.; Barros, L.; Santos-Buelga, C.; Silva, S.; Henriques, M.; Ferreira, I.C.F.R. Decoction, infusion and hydroalcoholic extract of cultivated thyme: Antioxidant and antibacterial activities, and phenolic characterisation. Food Chem. 2015, 167, 131–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, Y.K.; Jung, E.S.; Lee, H.A.; Choi, D.; Lee, C.H. Metabolomic Characterization of Hot Pepper (Capsicum annuum “cM334”) during Fruit Development. J. Agric. Food Chem. 2015, 63, 9452–9460. [Google Scholar] [CrossRef] [PubMed]
- Marín, A.; Ferreres, F.; Tomás-Barberán, F.A.; Gil, M.I. Characterization and quantitation of antioxidant constituents of sweet pepper (Capsicum annuum L.). J. Agric. Food Chem. 2004, 52, 3861–3869. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.Z.; Lu, S.; Harnly, J.M. Detection and quantification of glycosylated flavonoid malonates in celery, chinese celery, and celery seed by LC-DAD-ESI/MS. J. Agric. Food Chem. 2007, 55, 1321–1326. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Zhuang, L.; Song, D.; Lu, C.; Xu, X. Isolation, purification, and identification of the main phenolic compounds from leaves of celery (Apium graveolens L. var. dulce Mill./Pers.). J. Sep. Sci. 2017, 40, 472–479. [Google Scholar] [CrossRef]
- Freitas, J.R.L.; Vendramini, P.H.; Augusti, R.; Eberlin, M.N. Assessing the Spatial Distribution of Key Flavonoids in Mentha × piperita Leaves: An Application of Desorption Electrospray Ionization Mass Spectrometry Imaging (DESI-MSI). J. Braz. Chem. Soc. 2019, 30, 1437–1446. [Google Scholar] [CrossRef]
- Desta, K.T.; Kim, G.S.; El-Aty, A.M.A.; Raha, S.; Kim, M.B.; Jeong, J.H.; Warda, M.; Hacımüftüoğlu, A.; Shin, H.C.; Shim, J.H.; et al. Flavone polyphenols dominate in Thymus schimperi Ronniger: LC–ESI–MS/MS characterization and study of anti-proliferative effects of plant extract on AGS and HepG2 cancer cells. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1053, 1–8. [Google Scholar] [CrossRef]
- Akbari-Ahangar, A.; Delnavazi, M.R. Flavone glycosides from the aerial parts of Stachys lavandulifolia vahl. Pharm. Sci. 2020, 26, 198–202. [Google Scholar] [CrossRef]
- Sugawara, T.; Igarashi, K. Identification of Major Flavonoids in Petals of Edible Chrysanthemum Flowers and Their Suppressive Effect on Carbon Tetrachloride-Induced Liver Injury in Mice. Food Sci. Technol. Res. 2009, 15, 499–506. [Google Scholar] [CrossRef] [Green Version]
- Jeong, W.Y.; Jin, J.S.; Cho, Y.A.; Lee, J.H.; Park, S.; Jeong, S.W.; Kim, Y.H.; Lim, C.S.; Abd El-Aty, A.M.; Kim, G.S.; et al. Determination of polyphenols in three Capsicum annuum L. (bell pepper) varieties using high-performance liquid chromatographytandem mass spectrometry: Their contribution to overall antioxidant and anticancer activity. J. Sep. Sci. 2011, 34, 2967–2974. [Google Scholar] [CrossRef] [PubMed]
- Koley, T.K.; Khan, Z.; Oulkar, D.; Singh, B.K.; Maurya, A.; Singh, B.; Banerjee, K. High resolution LC-MS characterization of phenolic compounds and the evaluation of antioxidant properties of a tropical purple radish genotype. Arab. J. Chem. 2020, 13, 1355–1366. [Google Scholar] [CrossRef]
- Bannour, M.; Fellah, B.; Rocchetti, G.; Ashi-Smiti, S.; Lachenmeier, D.W.; Lucini, L.; Khadhri, A. Phenolic profiling and antioxidant capacity of Calligonum azel Maire, a Tunisian desert plant. Food Res. Int. 2017, 101, 148–154. [Google Scholar] [CrossRef]
- Rocchetti, G.; Miras-Moreno, M.B.; Zengin, G.; Senkardes, I.; Sadeer, N.B.; Mahomoodally, M.F.; Lucini, L. UHPLC-QTOF-MS phytochemical profiling and in vitro biological properties of Rhamnus petiolaris (Rhamnaceae). Ind. Crops Prod. 2019, 142, 111856. [Google Scholar] [CrossRef]
- Švehlíková, V.; Bennett, R.N.; Mellon, F.A.; Needs, P.W.; Piacente, S.; Kroon, P.A.; Bao, Y. Isolation, identification and stability of acylated derivatives of apigenin 7-O-glucoside from chamomile (Chamomilla recutita [L.] Rauschert). Phytochemistry 2004, 65, 2323–2332. [Google Scholar] [CrossRef]
- Slimestad, R.; Fossen, T.; Brede, C. Flavonoids and other phenolics in herbs commonly used in Norwegian commercial kitchens. Food Chem. 2020, 309, 125678. [Google Scholar] [CrossRef]
- Peng, Y.; Yan, Y.; Wan, P.; Dong, W.; Huang, K.; Ran, L.; Mi, J.; Lu, L.; Zeng, X.; Cao, Y. Effects of long-term intake of anthocyanins from Lycium ruthenicum Murray on the organism health and gut microbiota in vivo. Food Res. Int. 2020, 130, 108952. [Google Scholar] [CrossRef]
- Aldini, G.; Regazzoni, L.; Pedretti, A.; Carini, M.; Cho, S.M.; Park, K.M.; Yeum, K.J. An integrated high resolution mass spectrometric and informatics approach for the rapid identification of phenolics in plant extract. J. Chromatogr. A 2011, 1218, 2856–2864. [Google Scholar] [CrossRef]
- Rocchetti, G.; Lucini, L.; Ahmed, S.R.; Saber, F.R. In vitro cytotoxic activity of six Syzygium leaf extracts as related to their phenolic profiles: An untargeted UHPLC-QTOF-MS approach. Food Res. Int. 2019, 126, 108715. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, G.; Senizza, B.; Zengin, G.; Mahomodally, M.F.; Senkardes, I.; Lobine, D.; Lucini, L. Untargeted metabolomic profiling of three Crataegus species (hawthorn) and their in vitro biological activities. J. Sci. Food Agric. 2020, 100, 1998–2006. [Google Scholar] [CrossRef] [PubMed]
- Mara De Menezes Epifanio, N.; Rykiel Iglesias Cavalcanti, L.; Falcão Dos Santos, K.; Soares Coutinho Duarte, P.; Kachlicki, P.; Ozarowski, M.; Jorge Riger, C.; Siqueira De Almeida Chaves, D. Chemical characterization and in vivo antioxidant activity of parsley (Petroselinum crispum) aqueous extract. Food Funct. 2020, 11, 5346–5356. [Google Scholar] [CrossRef] [PubMed]
- Alseekh, S.; Perez de Souza, L.; Benina, M.; Fernie, A.R. The style and substance of plant flavonoid decoration; towards defining both structure and function. Phytochemistry 2020, 174, 112347. [Google Scholar] [CrossRef] [PubMed]
- Campos, L.; López-Gresa, M.P.; Fuertes, D.; Bellés, J.M.; Rodrigo, I.; Lisón, P. Tomato glycosyltransferase Twi1 plays a role in flavonoid glycosylation and defence against virus. BMC Plant Biol. 2019, 19, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Bontpart, T.; Cheynier, V.; Ageorges, A.; Terrier, N. BAHD or SCPL acyltransferase? What a dilemma for acylation in the world of plant phenolic compounds. New Phytol. 2015, 208, 695–707. [Google Scholar] [CrossRef]
- Rea, K.A.; Casaretto, J.A.; Al-Abdul-Wahid, M.S.; Sukumaran, A.; Geddes-McAlister, J.; Rothstein, S.J.; Akhtar, T.A. Biosynthesis of cannflavins A and B from Cannabis sativa L. Phytochemistry 2019, 164, 162–171. [Google Scholar] [CrossRef]
- Marín, L.; Gutiérrez-del-Río, I.; Yagüe, P.; Manteca, Á.; Villar, C.J.; Lombó, F. De novo biosynthesis of apigenin, luteolin, and eriodictyol in the actinomycete Streptomyces albus and production improvement by feeding and spore conditioning. Front. Microbiol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lam, P.Y.; Liu, H.; Lo, C. Completion of tricin biosynthesis pathway in rice: Cytochrome P450 75B4 is a unique chrysoeriol 5′-hydroxylase. Plant Physiol. 2015, 168, 1527–1536. [Google Scholar] [CrossRef] [Green Version]
- Muth, D.; Marsden-Edwards, E.; Kachlicki, P.; Stobiecki, M. Differentiation of isomeric malonylated flavonoid glyconjugates in plant extracts with UPLC-ESI/MS/MS. Phytochem. Anal. 2008, 19, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Abad-García, B.; Berrueta, L.A.; Garmón-Lobato, S.; Gallo, B.; Vicente, F. A general analytical strategy for the characterization of phenolic compounds in fruit juices by high-performance liquid chromatography with diode array detection coupled to electrospray ionization and triple quadrupole mass spectrometry. J. Chromatogr. A 2009, 1216, 5398–5415. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, X.; Mo, T.; Yan, Y.; Song, Y.; Zhao, Y.; Li, J.; Shi, S.; Liu, X.; Tu, P. Identification and functional application of a new malonyltransferase NbMaT1 towards diverse aromatic glycosides from Nicotiana benthamiana. RSC Adv. 2017, 7, 21028. [Google Scholar] [CrossRef] [Green Version]
- Kachlicki, P.; Piasecka, A.; Stobiecki, M.; Marczak, Ł. Structural characterization of flavonoid glycoconjugates and their derivatives with mass spectrometric techniques. Molecules 2016, 21, 1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Kim, H.W.; Lee, M.K.; Kim, Y.J.; Asamenew, G.; Cha, Y.S.; Kim, J.B. Phenolic profiling and quantitative determination of common sage (Salvia plebeia R. Br.) by UPLC-DAD-QTOF/MS. Eur. Food Res. Technol. 2018, 244, 1637–1646. [Google Scholar] [CrossRef] [Green Version]
- Park, D.; Barka, G.D.; Yang, E.-Y.; Cho, M.-C.; Yoon, J.B.; Lee, J. Identification of QTLs Controlling α-Glucosidase Genotyping-by-Sequencing Analysis. Genes 2020, 11, 1116. [Google Scholar] [CrossRef]
- KWON, Y.-I.; Apostolidis, E.; Shetty, K. Evaluation of pepper (Capsicum annuum) for management of diabetes and hypertension. J. Food Biochem. 2006, 31, 370–385. [Google Scholar] [CrossRef]
- Thanakosai, W.; Phuwapraisirisan, P. First identification of α-glucosidase inhibitors from okra (Abelmoschus esculentus) seeds. Nat. Prod. Commun. 2013, 8, 1085–1088. [Google Scholar] [CrossRef] [Green Version]
- Poovitha, S.; Parani, M. In vitro and in vivo α-amylase and α-glucosidase inhibiting activities of the protein extracts from two varieties of bitter gourd (Momordica charantia L.). BMC Complement. Altern. Med. 2016, 16, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Banihani, S.A. Radish (Raphanus sativus) and diabetes. Nutrients 2017, 9, 1014. [Google Scholar] [CrossRef] [Green Version]
- Vadivelan, R.; Gopala Krishnan, R.; Kannan, R. Antidiabetic potential of Asparagus racemosus Willd leaf extracts through inhibition of α-amylase and α-glucosidase. J. Tradit. Complement. Med. 2019, 9, 1–4. [Google Scholar] [CrossRef]
- Zhang, L.; Tu, Z.C.; Yuan, T.; Wang, H.; Xie, X.; Fu, Z.F. Antioxidants and α-glucosidase inhibitors from Ipomoea batatas leaves identified by bioassay-guided approach and structure-activity relationships. Food Chem. 2016, 208, 61–67. [Google Scholar] [CrossRef]
- Alhadrami, H.A.; Sayed, A.M.; El-Gendy, A.O.; Shamikh, Y.I.; Gaber, Y.; Bakeer, W.; Sheirf, N.H.; Attia, E.Z.; Shaban, G.M.; Khalifa, B.A.; et al. A metabolomic approach to target antimalarial metabolites in the Artemisia annua fungal endophytes. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Fujimura, Y.; Kawano, C.; Maeda-Murayama, A.; Nakamura, A.; Koike-Miki, A.; Yukihira, D.; Hayakawa, E.; Ishii, T.; Tachibana, H.; Wariishi, H.; et al. A Chemometrics-driven Strategy for the Bioactivity Evaluation of Complex Multicomponent Systems and the Effective Selection of Bioactivity-predictive Chemical Combinations. Sci. Rep. 2017, 7, 6–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimura, Y.; Kurihara, K.; Ida, M.; Kosaka, R.; Miura, D.; Wariishi, H.; Maeda-Yamamoto, M.; Nesumi, A.; Saito, T.; Kanda, T.; et al. Metabolomics-driven nutraceutical evaluation of diverse green tea cultivars. PLoS ONE 2011, 6, e23426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demarque, D.P.; Dusi, R.G.; de Sousa, F.D.M.; Grossi, S.M.; Silvério, M.R.S.; Lopes, N.P.; Espindola, L.S. Mass spectrometry-based metabolomics approach in the isolation of bioactive natural products. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raheem, D.J.; Tawfi, A.F.; Abdelmohsen, U.R. Application of metabolomics and molecular networking in investigating the chemical profile and antitrypanosomal activity of British bluebells (Hyacinthoides non-scripta). Sci. Rep. 2019, 9, 1–13. [Google Scholar]
- Wang, W.; Snooks, H.D.; Sang, S. The Chemistry and Health Benefits of Dietary Phenolamides. J. Agric. Food Chem. 2020, 68, 6248–6267. [Google Scholar] [CrossRef] [PubMed]
- Niwa, T.; Doi, U.; Osawa, T. Inhibitory activity of corn-derived bisamide compounds against α-glucosidase. J. Agric. Food Chem. 2003, 51, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Stewart, D.A.; Ye, X.M.; Yin, L.H.; Pathmasiri, W.W.; McRitchie, S.L.; Fennell, T.R.; Cheung, H.Y.; Sumner, S.J. A metabolomics approach to investigate kukoamine B—A potent natural product with anti-diabetic properties. Front. Pharmacol. 2019, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Alagesan, K.; Thennarasu, P.; Kumar, V.; Sankarnarayanan, S.; Balsamy, T. Identification of α-Glucosidase Inhibitors From Psidium guajava Leaves and Syzygium cumini Linn. Seeds. Int. J. Pharm. Sci. Res. 2012, 3, 316–322. [Google Scholar]
- Ahn-Jarvis, J.H.; Parihar, A.; Doseff, A.I. Dietary flavonoids for immunoregulation and cancer: Food design for targeting disease. Antioxidants 2019, 8, 202. [Google Scholar] [CrossRef] [Green Version]
- Day, A.J.; Cañada, F.J.; Díaz, J.C.; Kroon, P.A.; McLauchlan, R.; Faulds, C.B.; Plumb, G.W.; Morgan, M.R.A.; Williamson, G. Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of lactase phlorizin hydrolase. FEBS Lett. 2000, 468, 166–170. [Google Scholar] [CrossRef] [Green Version]
- Asamenew, G.; Kim, H.W.; Lee, M.K.; Lee, S.H.; Kim, Y.J.; Cha, Y.S.; Yoo, S.M.; Kim, J.B. Characterization of phenolic compounds from normal ginger (Zingiber officinale Rosc.) and black ginger (Kaempferia parviflora Wall.) using UPLC–DAD–QToF–MS. Eur. Food Res. Technol. 2019, 245, 653–665. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Cheng, X.; Wang, L.; Wang, S.; Ren, G. A determination of potential α-glucosidase inhibitors from azuki beans (Vigna angularis). Int. J. Mol. Sci. 2011, 12, 6445–6451. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compounds Name | RT (min) | UVλmax (nm) | MolecularWeight | Molecular Formula | Experimental(m/z) [M+H]+,[M+Na]+,[M+K]+ | Fragmentations (m/z) |
|---|---|---|---|---|---|---|---|
| 1 | N-cis caffeoyl putrescine | 2.82 | 230 | 250 | C13H18N2O3 | 251,273,ND | 163,135 |
| 2 | N-tans caffeoyl putrescine | 3.95 | 230,318 | 250 | C13H18N2O3 | 251,273,ND | 163,135 |
| 3 | Feruloyl putrescine | 4.97 | 227,230 | 264 | C14H20N2O3 | 265,277,ND | 177,145 |
| 4 | Coumaroyl putrescine | 5.33 | 230,292 | 234 | C13H18N2O2 | 235,257,ND | 147,119 |
| 5 a | 5 caffeoylquinci acid (5CQA) | 6.59 | 231,325 | 354 | C16H18O9 | 355,377,393 | 337,181,163,145,135 |
| 6 a | Luteolin 7-O-(2”-O-apiosyl)glucoside | 14.32 | 227,253,348 | 580 | C26H28O15 | 581,603,619 | 449,287 |
| 7 a | Luteolin 7-O-glucoside | 14.39 | 227,347 | 448 | C21H20O11 | 449,471,ND | 287 |
| 8 b | Luteolin-O-apiosyl malonyl). Hexoside (isomer of 13) | 16.18 | 227,348 | 666 | C21H20O11 | 667,689,705 | 535,287 |
| 9 a | Apigenin 7-O-(2”-O-apiosyl)glucoside | 16.38 | 228,266,337 | 564 | C26H28O14 | 565,587,603 | 433,271 |
| 10 a | Apigenin 7-O-glucoside | 16.69 | 227,266,336 | 432 | C21H20O10 | 433,455,ND | 271 |
| 11 b | Luteolin 7-O-(2”-O-apiosyl-6”-O-malonyl)glucosideLuteolin-O-(apiosyl malonyl) hexoside (isomer of 13) | 16.8 | 227,348 | 666 | C29H30O18 | 667,689,705 | 535,287 |
| 12 | Chrysoeriol-O-(apiosyl) hexoside | 17.01 | 227,347 | 594 | C27H30O15 | 595,617,633 | 463,301,286 |
| 13 | Luteolin 7-O-(2”-O-apiosyl-6”-O-malonyl)glucoside | 17.48 | 227,254,348 | 666 | C29H30O18 | 667,689,705 | 535,287 |
| 14 b | Luteolin-O-(malonyl) hexoside | 17.71 | 227.348 | 534 | C24H22O14 | 535,557,ND | 449,287 |
| 15 b | Apigenin-O-(apiosyl malonyl) hexoside(isomer of 18) | 18.23 | 227,336 | 650 | C29H30O17 | 651,673,689 | 519,271 |
| 16 b | Apigenin-O-(apiosyl malonyl) hexoside (isomer of 18) | 18.85 | 227,267,337 | 650 | C29H30O17 | 651,673,689 | 519,271 |
| 17 b | Chrysoeriol-O-(apiosyl malonyl) hexoside (isomer) | 19.5 | 227,347 | 680 | C30H32O18 | 681,703,719 | 549,301,286 |
| 18 | Apigenin 7-O-(2”-O-apiosyl-6”-O-malonyl)glucoside | 19.67 | 230,267,336 | 650 | C29H30O17 | 651,673,689 | 519,271 |
| 19 | Chrysoeriol-O-(apiosyl malonyl) hexoside (isomer) | 20.1 | 227,251,346 | 680 | C30H32O18 | 681,703,719 | 549,301,286 |
| 20 b | Apigenin-O-(malonyl) hexoside | 20.2 | 227,267,336 | 518 | C24H22O13 | 519,541,ND | 433,271 |
| 21 b | Chrysoeriol-O-(malonyl) hexoside | 20.62 | 231,347 | 548 | C25H24O14 | 549,571,ND | 301,286 |
| Pepper Extract a | ||||
|---|---|---|---|---|
| Genotypes | IC50 (mg/mL) | Standard Compounds b | Inhibition% (250 µM) | IC50 (µM) |
| A24 | 0.34 ± 0.04 | Caffeoyl-Putrescine | 61.36 ± 1.37 | 145 |
| A29 | 0.5 ± 0.035 | Acarbose | 57.06 ± 1.8 | 197 |
| G38 | 0.122± 0.004 | Apigenin-7-O glucoside | 30.68 ± 5.35 | >300 |
| G21 | 2.5 ± 0.22 | Luteolin-7-O glucoside | 37.65 ± 3.08 | >300 |
| ACARBOSE | 0.19 ± 0.034 | Apiin | 13.25 ± 0.3 d | ND |
| Luteolin-7-O-(2” apiosyl)glucoside | 29.9 ± 4.02 d | ND | ||
| Luteolin | 97.84 ± 0.56 c | 7.6 | ||
| Apigenin | 56.14 ± 2.37 c | 81 | ||
| OPLS Model | SS | DF | MS | F | p | SD |
|---|---|---|---|---|---|---|
| Total | 125 | 125 | 1 | 1 | ||
| Regression | 67.5143 | 4 | 16.8786 | 35.5272 | 1.32 × 10−19 | 4.10835 |
| Residual | 57.4857 | 121 | 0.475089 | 0.689267 |
| Model | Type | Components | N | R2X (cum) | R2Y (cum) | Q2 (cum) |
|---|---|---|---|---|---|---|
| M1 | PCA-X | 4 | 126 | 0.818 | - | 0.634 |
| M2 | OPLS | 1 + 1 + 0 | 126 | 0.535 | 0.61 | 0.54 |
| M3 | OPLS-Class (1) | 1 + 2 + 0 | 39 | 0.578 | 0.856 | 0.775 |
| M4 | OPLS-Class (2) | 1 + 3 + 0 | 11 | 0.753 | 0.945 | 0.565 |
| M5 | OPLS-Class (3) | 1 + 2 + 0 | 76 | 0.544 | 0.726 | 0.605 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assefa, S.T.; Yang, E.-Y.; Asamenew, G.; Kim, H.-W.; Cho, M.-C.; Lee, J. Identification of α-Glucosidase Inhibitors from Leaf Extract of Pepper (Capsicum spp.) through Metabolomic Analysis. Metabolites 2021, 11, 649. https://doi.org/10.3390/metabo11100649
Assefa ST, Yang E-Y, Asamenew G, Kim H-W, Cho M-C, Lee J. Identification of α-Glucosidase Inhibitors from Leaf Extract of Pepper (Capsicum spp.) through Metabolomic Analysis. Metabolites. 2021; 11(10):649. https://doi.org/10.3390/metabo11100649
Chicago/Turabian StyleAssefa, Samuel Tilahun, Eun-Young Yang, Gelila Asamenew, Heon-Woong Kim, Myeong-Cheoul Cho, and Jundae Lee. 2021. "Identification of α-Glucosidase Inhibitors from Leaf Extract of Pepper (Capsicum spp.) through Metabolomic Analysis" Metabolites 11, no. 10: 649. https://doi.org/10.3390/metabo11100649
APA StyleAssefa, S. T., Yang, E.-Y., Asamenew, G., Kim, H.-W., Cho, M.-C., & Lee, J. (2021). Identification of α-Glucosidase Inhibitors from Leaf Extract of Pepper (Capsicum spp.) through Metabolomic Analysis. Metabolites, 11(10), 649. https://doi.org/10.3390/metabo11100649