Metabolomic Signature of Amino Acids, Biogenic Amines and Lipids in Blood Serum of Patients with Severe Osteoarthritis

 , ,
, ,
Abstract
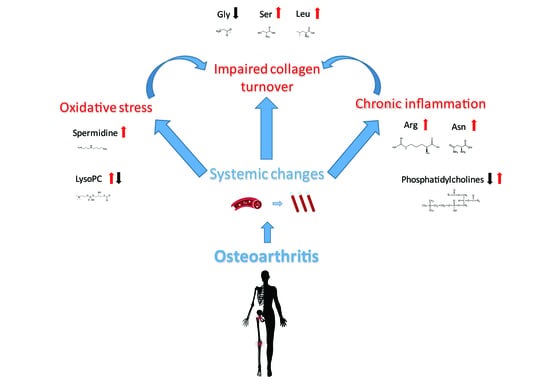
1. Introduction
2. Results
3. Discussion
3.1. Amino Acids and Biogenic Amines
3.2. Lipidomic Analysis
4. Materials and Methods
4.1. Study Participants
4.2. Study Protocol
4.3. Biochemical Analysis
4.4. Targeted Metabolite Assessment
4.5. Radiological Measurement of Osteoarthritis Severity
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tootsi, K.; Märtson, A.; Kals, J.; Paapstel, K.; Zilmer, M. Metabolic factors and oxidative stress in osteoarthritis: A case–control study. Scand. J. Clin. Lab. Investig. 2017, 77, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Pegg, A.E. Mammalian polyamine metabolism and function. IUBMB Life 2009, 61, 880–894. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Functional Amino Acids in Growth, Reproduction, and Health12. Adv. Nutr. 2010, 1, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Likhodii, S.; Zhang, Y.; Aref-Eshghi, E.; Harper, P.E.; Randell, E.; Green, R.; Martin, G.; Furey, A.; Sun, G.; et al. Classification of osteoarthritis phenotypes by metabolomics analysis. BMJ Open 2014, 4, e006286. [Google Scholar] [CrossRef] [PubMed]
- Zhai, G.; Wang-Sattler, R.; Hart, D.J.; Arden, N.K.; Hakim, A.J.; Illig, T.; Spector, T.D. Serum branched-chain amino acid to histidine ratio: A novel metabolomic biomarker of knee osteoarthritis. Ann. Rheum. Dis. 2010, 69, 1227–1231. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, G.; Likhodii, S.; Liu, M.; Aref-Eshghi, E.; Harper, P.; Martin, G.; Furey, A.; Green, R.; Randell, E.; et al. Metabolomic analysis of human plasma reveals that arginine is depleted in knee osteoarthritis patients. Osteoarthr. Cartil. 2016, 24, 827–834. [Google Scholar] [CrossRef]
- Chiurchiù, V.; Leuti, A.; Maccarrone, M. Bioactive Lipids and Chronic Inflammation: Managing the Fire Within. Front. Immunol. 2018, 9, 38. [Google Scholar] [CrossRef]
- Tootsi, K.; Kals, J.; Zilmer, M.; Paapstel, K.; Ottas, A.; Märtson, A. Medium- and long-chain acylcarnitines are associated with osteoarthritis severity and arterial stiffness in end-stage osteoarthritis patients: A case-control study. Int. J. Rheum. Dis. 2018, 21, 1211–1218. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, G.; Aitken, D.; Likhodii, S.; Liu, M.; Martin, G.; Furey, A.; Randell, E.; Rahman, P.; Jones, G.; et al. Lysophosphatidylcholines to phosphatidylcholines ratio predicts advanced knee osteoarthritis. Rheumatology 2016, 55, 1566–1574. [Google Scholar] [CrossRef]
- Pousinis, P.; Gowler, P.R.W.; Burston, J.J.; Ortori, C.A.; Chapman, V.; Barrett, D.A. Lipidomic identification of plasma lipids associated with pain behaviour and pathology in a mouse model of osteoarthritis. Metabolomics 2020, 16, 32. [Google Scholar] [CrossRef] [PubMed]
- Kosinska, M.K.; Liebisch, G.; Lochnit, G.; Wilhelm, J.; Klein, H.; Kaesser, U.; Lasczkowski, G.; Rickert, M.; Schmitz, G.; Steinmeyer, J. A Lipidomic Study of Phospholipid Classes and Species in Human Synovial Fluid. Arthritis Rheum. 2013, 65, 2323–2333. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-H.; Chen, L.; Hsieh, M.-S.; Chang, C.-P.; Chou, D.-T.; Tsai, S.-H. Evidence for a protective role for adiponectin in osteoarthritis. Biochim. Biophys. Acta Mol. Basis Dis. 2006, 1762, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Maturo, M.G.; Soligo, M.; Gibson, G.; Manni, L.; Nardini, C. The greater inflammatory pathway—High clinical potential by innovative predictive, preventive, and personalized medical approach. EPMA J. 2019, 11, 1–16. [Google Scholar] [CrossRef]
- Qian, S.; Golubnitschaja, O.; Zhan, X. Chronic inflammation: Key player and biomarker-set to predict and prevent cancer development and progression based on individualized patient profiles. EPMA J. 2019, 10, 365–381. [Google Scholar] [CrossRef]
- Morris, S.M. Arginine Metabolism: Boundaries of Our Knowledge. J. Nutr. 2007, 137, 1602S–1609S. [Google Scholar] [CrossRef]
- Sharma, J.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Bezerra, M.M.; Brain, S.D.; Greenacre, S.; Jerônimo, S.M.B.; De Melo, L.B.; Keeble, J.; Rocha, F.A.C. Reactive nitrogen species scavenging, rather than nitric oxide inhibition, protects from articular cartilage damage in rat zymosan-induced arthritis. Br. J. Pharmacol. 2003, 141, 172–182. [Google Scholar] [CrossRef]
- Shi, H.P.; Wang, S.M.; Zhang, G.X.; Zhang, Y.J.; Barbul, A. Supplemental l-arginine enhances wound healing following trauma/hemorrhagic shock. Wound Repair Regen. 2007, 15, 66–70. [Google Scholar] [CrossRef]
- Chen, R.; Han, S.; Liu, X.; Wang, K.; Zhou, Y.; Yang, C.; Zhang, X. Perturbations in amino acids and metabolic pathways in osteoarthritis patients determined by targeted metabolomics analysis. J. Chromatogr. B 2018, 1085, 54–62. [Google Scholar] [CrossRef]
- Pascale, V.; Pascale, W.; Lavanga, V.; Sansone, V.; Ferrario, P.; Colonna, V.D.G. L-arginine, asymmetric dimethylarginine, and symmetric dimethylarginine in plasma and synovial fluid of patients with knee osteoarthritis. Med Sci. Monit. 2013, 19, 1057–1062. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, F.; Okayasu, H.; Tashiro, M.; Hashimoto, K.; Yokote, Y.; Akahane, K.; Hongo, S.; Sakagami, H. Effect of lignins and their precursors on nitric oxide, citrulline and asparagine production by mouse macrophage-like Raw 264.7 cells. Anticancer Res. 2003, 22, 2719–2724. [Google Scholar]
- Li, P.; Yin, J.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Melendez-Hevia, E.; Lugo, P.D.P.; Cornish-Bowden, A.; Cárdenas, M.L.; Paz-Lugo, P. A weak link in metabolism: The metabolic capacity for glycine biosynthesis does not satisfy the need for collagen synthesis. J. Biosci. 2009, 34, 853–872. [Google Scholar] [CrossRef] [PubMed]
- Skamarauskas, J.T.; Oakley, F.; Smith, F.E.; Bawn, C.; Dunn, M.; Vidler, D.S.; Clemence, M.; Blain, P.G.; Taylor, R.; Gamcsik, M.P.; et al. Noninvasive in vivo magnetic resonance measures of glutathione synthesis in human and rat liver as an oxidative stress biomarker. Hepatology 2014, 59, 2321–2330. [Google Scholar] [CrossRef]
- Issa, R.I.; Griffin, T.M. Pathobiology of obesity and osteoarthritis: Integrating biomechanics and inflammation. Pathobiol. Aging Age Relat. Dis. 2012, 2, 671. [Google Scholar] [CrossRef]
- Lugo, P.D.P.; Lupiáñez, J.A.; Meléndez-Hevia, E. High glycine concentration increases collagen synthesis by articular chondrocytes in vitro: Acute glycine deficiency could be an important cause of osteoarthritis. Amino Acids 2018, 50, 1357–1365. [Google Scholar] [CrossRef]
- Zhai, G. Alteration of Metabolic Pathways in Osteoarthritis. Metabolites 2019, 9, 11. [Google Scholar] [CrossRef]
- Maher, A.D.; Coles, C.; White, J.D.; Bateman, J.F.; Fuller, E.S.; Burkhardt, D.; Little, C.B.; Cake, M.; Read, R.; McDonagh, M.B.; et al. 1H NMR Spectroscopy of Serum Reveals Unique Metabolic Fingerprints Associated with Subtypes of Surgically Induced Osteoarthritis in Sheep. J. Proteome Res. 2012, 11, 4261–4268. [Google Scholar] [CrossRef]
- Lamers, R.; Van Nesselrooij, J.; Kraus, V.; Jordan, J.; Renner, J.; Dragomir, A.; Luta, G.; Van Der Greef, J.; DeGroot, J. Identification of an urinary metabolite profile associated with osteoarthritis. Osteoarthr. Cartil. 2005, 13, 762–768. [Google Scholar] [CrossRef]
- Rider, J.E.; Hacker, A.; Mackintosh, C.A.; Pegg, A.E.; Woster, P.M.; Casero, R.A., Jr. Spermine and spermidine mediate protection against oxidative damage caused by hydrogen peroxide. Amino Acids 2007, 33, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Brazill, J.M.; Liu, S.; Bello, C.; Zhu, Y.; Morimoto, M.; Cascio, L.; Pauly, R.; Diaz-Perez, Z.; Malicdan, M.C.; et al. Spermine synthase deficiency causes lysosomal dysfunction and oxidative stress in models of Snyder-Robinson syndrome. Nat. Commun. 2017, 8, 1257. [Google Scholar] [CrossRef] [PubMed]
- Krüger, A.; Vowinckel, J.; Mülleder, M.; Grote, P.; Capuano, F.; Bluemlein, K.; Ralser, M. Tpo1-mediated spermine and spermidine export controls cell cycle delay and times antioxidant protein expression during the oxidative stress response. EMBO Rep. 2013, 14, 1113–1119. [Google Scholar] [CrossRef] [PubMed]
- Igari, T.; Shimamura, T. Serotonin Metabolism and its Enzymic Activities in Joint Diseases. Clin. Orthop. Relat. Res. 1979, 232–249. [Google Scholar] [CrossRef]
- Lavoie, B.; Lian, J.B.; Mawe, G.M. Regulation of Bone Metabolism by Serotonin. Retinal Degener. Dis. 2017, 1033, 35–46. [Google Scholar] [CrossRef]
- Kosinska, M.K.; Liebisch, G.; Lochnit, G.; Wilhelm, J.; Klein, H.; Kaesser, U.; Lasczkowski, G.; Rickert, M.; Schmitz, G.; Steinmeyer, J. Sphingolipids in Human Synovial Fluid - A Lipidomic Study. PLoS ONE 2014, 9, e91769. [Google Scholar] [CrossRef]
- Castro-Perez, J.M.; Kamphorst, J.; DeGroot, J.; Lafeber, F.; Goshawk, J.; Yu, K.; Shockcor, J.P.; Vreeken, R.J.; Hankemeier, T. Comprehensive LC-MS E lipidomic analysis using a shotgun approach and its application to biomarker detection and identification in osteoarthritis patients. J. Proteome Res. 2010, 9, 2377–2389. [Google Scholar] [CrossRef]
- Farooqui, A.A.; Horrocks, L.A.; Farooqui, T. Glycerophospholipids in brain: Their metabolism, incorporation into membranes, functions, and involvement in neurological disorders. Chem. Phys. Lipids 2000, 106, 1–29. [Google Scholar] [CrossRef]
- White, D.A. The phospholipid composition of mammalian tissues. In Form Function Phospholipids; Ansell, G.B., Hawthorne, J.N., Dawson, R.M.C., Eds.; Elsevier: London, UK, 1973. [Google Scholar]
- Schmidt, T.A.; Gastelum, N.S.; Nguyen, Q.T.; Schumacher, B.L.; Sah, R.L. Boundary lubrication of articular cartilage: Role of synovial fluid constituents. Arthritis Rheum. 2007, 56, 882–891. [Google Scholar] [CrossRef]
- Courties, A.; Gualillo, O.; Berenbaum, F.; Sellam, J. Metabolic stress-induced joint inflammation and osteoarthritis. Osteoarthr. Cartil. 2015, 23, 1955–1965. [Google Scholar] [CrossRef]
- Sun, A.R.; Friis, T.; Sekar, S.; Crawford, R.; Xiao, Y.; Prasadam, I. Is Synovial Macrophage Activation the Inflammatory Link Between Obesity and Osteoarthritis? Curr. Rheumatol. Rep. 2016, 18, 57. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, J.P.; Major, A.S. How Oxidized Low-Density Lipoprotein Activates Inflammatory Responses. Crit. Rev. Immunol. 2018, 38, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Zhai, G.; Pelletier, J.-P.; Liu, M.; Randell, E.W.; Rahman, P.; Martel-Pelletier, J. Serum lysophosphatidylcholines to phosphatidylcholines ratio is associated with symptomatic responders to symptomatic drugs in knee osteoarthritis patients. Arthritis Res. 2019, 21, 224–225. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, S.; Futerman, A. The metabolism and function of sphingolipids and glycosphingolipids. Cell. Mol. Life Sci. 2007, 64, 2270–2284. [Google Scholar] [CrossRef] [PubMed]
- Altman, R.; Alarcon, G.; Appelrouth, D.; Bloch, D.; Borenstein, D.; Brandt, K.; Brown, C.; Cooke, T.D.; Daniel, W.; Feldman, D.; et al. The American College of Rheumatology criteria for the classification and reporting of osteoarthritis of the hip. Arthritis Rheum. 1991, 34, 505–514. [Google Scholar] [CrossRef]
- Altman, R.; Asch, E.; Bloch, D.; Bole, G.; Borenstein, D.; Brandt, K.; Christy, W.; Cooke, T.D.; Greenwald, R.; Hochberg, M.; et al. Development of criteria for the classification and reporting of osteoarthritis: Classification of osteoarthritis of the knee. Arthritis Rheum. 1986, 29, 1039–1049. [Google Scholar] [CrossRef]
- Kellgren, J.H.; Lawrence, J.S. Radiological Assessment of Osteo-Arthrosis. Ann. Rheum. Dis. 1957, 16, 494–502. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Variable | Osteoarthritis (n = 70) | Controls (n = 82) | p-Value |
|---|---|---|---|
| Age (years) | 62 ± 7 | 61 ± 8 | 0.173 |
| Male/Female (n) | 36/34 | 38/44 | 0.871 |
| BMI (kg/m2) | 27.8 ± 3.2 | 26.0 ± 3.5 | 0.001 |
| hs-CRP (mg/L) | 1.9 ± 1.1 | 1.5 ± 1.2 | 0.014 |
| WBC (109/L) | 6.5 ± 1.4 | 5.7 ± 1.9 | 0.001 |
| LDL-cholesterol (mmol/L) | 4.0 ± 0.9 | 3.9 ± 1.1 | 0.804 |
| Triglycerides (mmol/L) | 1.7 ± 0.8 | 1.4 ± 1.1 | 0.001 |
| HDL-cholesterol (mmol/L) | 1.5 ± 0.5 | 1.7 ± 0.5 | 0.001 |
| Total cholesterol (mmol/L) | 5.8 ± 1.1 | 5.7 ± 1.3 | 0.539 |
| Fasting glucose (mmol/L) | 5.7 ± 0.7 | 5.7 ± 0.5 | 0.509 |
| Urea (mmol/L) | 6.1 ± 2.1 | 5.2 ± 1.2 | 0.014 |
| Variable | Osteoarthritis (n = 70) | Controls (n = 82) | p-Value |
|---|---|---|---|
| Alanine | 430.9 ± 95.9 | 403.8 ± 89.5 | 0.172 |
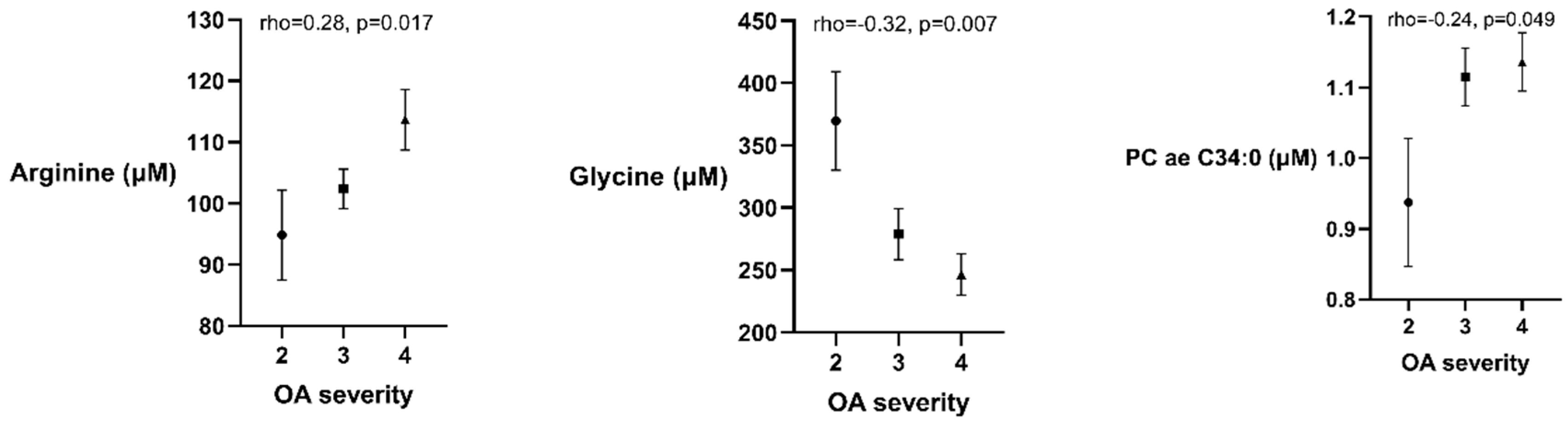
| Arginine | 105.2 ± 22.4 | 96.3 ± 23.5 | 0.020 * |
| Asparagine | 48.4 ± 8.3 | 44.1 ± 9.7 | <0.001 |
| Aspartate | 28.0 ± 8.8 | 29.2 ± 10.8 | 0.692 |
| Citrulline | 36.8 ± 10.1 | 36.2 ± 9.7 | 0.302 |
| Glutamine | 703.9 ± 128.4 | 679.8 ± 131.5 | 0.130 |
| Glutamate | 50.7 ± 21.0 | 48.4 ± 20.6 | 0.872 |
| Glycine | 280.8 ± 116.2 | 289.8 ± 114.6 | 0.794 |
| Histidine | 106.3 ± 16.7 | 101.8 ± 16.1 | 0.185 |
| Isoleucine | 86.9 ± 25.5 | 79.9 ± 21.7 | 0.207 |
| Leucine | 180.9 ± 50.0 | 156.7 ± 39.5 | 0.008 |
| Lysine | 233.9 ± 42.2 | 224.4 ± 44.8 | 0.479 |
| Methionine | 24.3 ± 6.1 | 22.7 ± 5.7 | 0.137 |
| Ornithine | 92.8 ± 20.4 | 92.1 ± 23.2 | 0.892 |
| Phenylalanine | 74.1 ± 12.9 | 68.3 ± 11.8 | 0.016 |
| Proline | 162.3 ± 32.9 | 167.3 ± 48.2 | 0.352 |
| Serine | 137.4 ± 29.1 | 121.8 ± 27.6 | 0.001 |
| Threonine | 116.5 ± 21.8 | 118.0 ± 25.2 | 0.845 |
| Tryptophan | 68.6 ± 15.6 | 64.6 ± 14.7 | 0.220 |
| Tyrosine | 72.1 ± 16.8 | 67.3 ± 17.1 | 0.234 |
| Valine | 265.2 ± 60.3 | 254 ± 51.5 | 0.694 |
| ADMA | 0.618 ± 0.156 | 0.559 ± 0.133 | 0.037 |
| Serotonin | 0.576 ± 0.515 | 0.685 ± 0.280 | 0.004 |
| Spermidine | 0.188 ± 0.051 | 0.161 ± 0.046 | 0.001 |
| Taurine | 122.2 ± 32.0 | 124.2 ± 29.4 | 0.825 |
| Spermine | 0.158 ± 0.014 | 0.159 ± 0.016 | 0.677 |
| Spermine-spermidine ratio | 0.898 ± 0.227 | 1.060 ± 0.279 | <0.001 |
| Metabolite | Control | Std | Osteoarthritis | Std | p-Value |
|---|---|---|---|---|---|
| lysoPC a C14:0 | 5.256 | ±0.873 | 4.748 | ±0.563 | <0.001 |
| lysoPC a C20:4 | 8.689 | ±2.414 | 9.843 | ±2.589 | 0.026 |
| PC aa C30:0 | 4.143 | ±1.165 | 3.384 | ±0.77 | <0.001 |
| PC aa C32:2 | 3.069 | ±1.052 | 2.141 | ±0.794 | <0.001 |
| PC aa C32:3 | 0.424 | ±0.097 | 0.374 | ±0.085 | 0.010 |
| PC aa C34:3 | 12.75 | ±3.573 | 10.64 | ±2.79 | 0.010 |
| PC aa C34:4 | 1.367 | ±0.481 | 1.107 | ±0.379 | <0.001 |
| PC aa C38:6 | 76.57 | ±24.64 | 91.61 | ±25.77 | 0.010 |
| PC aa C40:6 | 27.41 | ±9.457 | 33.12 | ±9.85 | 0.010 |
| PC ae C30:0 | 0.430 | ±0.103 | 0.368 | ±0.075 | <0.001 |
| PC ae C34:2 | 9.942 | ±3.11 | 8.207 | ±1.757 | 0.010 |
| PC ae C34:3 | 6.139 | ±2.02 | 4.768 | ±1.202 | <0.001 |
| SM C20:2 | 0.118 | ±0.051 | 0.143 | ±0.049 | 0.032 |
| (a) Regression Model with Glycine as the Dependent Variable in the Osteoarthritis Patients Group. | ||||
| Variable | B | Std Error | Beta | p-Value |
| Constant | 482.1 | 173.1 | 0.007 | |
| OA severity | −43.2 | 20.6 | −0.25 | 0.040 |
| Gender | −64.7 | 28.1 | −0.28 | 0.025 |
| Age | 0.5 | 1.8 | 0.03 | 0.777 |
| BMI | 3.0 | 4.3 | 0.08 | 0.483 |
| Glucose | −25–3 | 20.7 | −0.15 | 0.227 |
| (b) Regression Model with Arginine as the Dependent Variable in the Osteoarthritis Patients Group. | ||||
| Variable | B | Std Error | Beta | p-Value |
| Constant | 140.1 | 34.9 | 0.001 | |
| OA severity | 9.12 | 4.1 | 0.28 | 0.031 |
| Gender | 5.77 | 5.6 | 0.13 | 0.310 |
| Age | −0.31 | 0.37 | −0.10 | 0.392 |
| BMI | −0.24 | 0.86 | −0.033 | 0.782 |
| Glucose | −7.11 | 4.15 | −0.21 | 0.092 |
| (c) Regression Model with PC ae 34:0 as the Dependent Variable in the Osteoarthritis Patients Group. | ||||
| Variable | B | Std Error | Beta | p-Value |
| Constant | 0.440 | 0.34 | 0.196 | |
| OA severity | 0.07 | 0.04 | 0.19 | 0.097 |
| Gender | 0.03 | 0.06 | 0.06 | 0.593 |
| Age | −0.01 | 0.01 | 0.01 | 0.689 |
| BMI | −0.01 | 0.01 | −0.07 | 0.499 |
| Cholesterol | 0.115 | 0.026 | 0.50 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tootsi, K.; Vilba, K.; Märtson, A.; Kals, J.; Paapstel, K.; Zilmer, M. Metabolomic Signature of Amino Acids, Biogenic Amines and Lipids in Blood Serum of Patients with Severe Osteoarthritis. Metabolites 2020, 10, 323. https://doi.org/10.3390/metabo10080323
Tootsi K, Vilba K, Märtson A, Kals J, Paapstel K, Zilmer M. Metabolomic Signature of Amino Acids, Biogenic Amines and Lipids in Blood Serum of Patients with Severe Osteoarthritis. Metabolites. 2020; 10(8):323. https://doi.org/10.3390/metabo10080323
Chicago/Turabian StyleTootsi, Kaspar, Kadri Vilba, Aare Märtson, Jaak Kals, Kaido Paapstel, and Mihkel Zilmer. 2020. "Metabolomic Signature of Amino Acids, Biogenic Amines and Lipids in Blood Serum of Patients with Severe Osteoarthritis" Metabolites 10, no. 8: 323. https://doi.org/10.3390/metabo10080323
APA StyleTootsi, K., Vilba, K., Märtson, A., Kals, J., Paapstel, K., & Zilmer, M. (2020). Metabolomic Signature of Amino Acids, Biogenic Amines and Lipids in Blood Serum of Patients with Severe Osteoarthritis. Metabolites, 10(8), 323. https://doi.org/10.3390/metabo10080323