Effects of Cyclic High Ambient Temperature and Dietary Supplementation of Orotic Acid, a Pyrimidine Precursor, on Plasma and Muscle Metabolites in Broiler Chickens

 ,
,  ,
,
Abstract
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Determination of MDA Concentration
4.3. Determination of Free Amino Acid Concentrations
4.4. Sample Preparation for GC-MS/MS Analysis
4.5. GC-MS/MS Analysis and Data Processing
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ensminger, M.E.; Oldfield, J.E.; Heineman, W.W. Feeds and Nutrition, 2nd ed.; Ensminger Pub Co: Prentice Hall: Upper Saddle River, NJ, USA, 1990; pp. 8–110. [Google Scholar]
- Sahin, K.; Sahin, N.; Kucuk, O.; Hayirili, A.; Prasad, A.S. Role of dietary zinc in heat stressed poultry: A review. Poult. Sci. 2009, 88, 2176–2183. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.U.; Naz, S.; Nikousefat, Z.; Selvaggi, M.; Laudadio, V.; Tufarelli, V. Effect of ascorbic acid in heat-stressed poultry. Worlds Poult. Sci. J. 2012, 68, 477–489. [Google Scholar] [CrossRef]
- Yalçin, S.; Settar, P.; Ozkan, S.; Cahaner, A. Comparative evaluation of three commercial broiler stocks in hot versus temperate climates. Poult. Sci. 1997, 76, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Yunis, R.; Cahaner, A. The effects of naked neck (Na) and frizzle (F) genes on growth and meat yields of broilers and their interactions with ambient temperatures and potential growth rate. Poult. Sci. 1999, 78, 1347–1352. [Google Scholar] [CrossRef]
- Whitehead, C.C.; Keller, T. An update on ascorbic acid in poultry. Worlds Poult. Sci. J. 2003, 59, 161–184. [Google Scholar] [CrossRef]
- Zeferino, C.P.; Komiyama, C.M.; Pelícia, V.C.; Fascina, V.B.; Aoyagi, M.M.; Coutinho, L.L.; Sartori, J.R.; Moura, A.S.A.M.T. Carcass and meat quality traits of chickens fed diets concurrently supplemented with vitamins C and E under constant heat stress. Animal 2016, 10, 163–171. [Google Scholar] [CrossRef]
- Zhu, X.; Ruusunen, M.; Gusella, M.; Zhou, G.; Puolanne, E. High post-mortem temperature combined with rapid glycolysis induces phosphorylase denaturation and produces pale and exudative characteristics in broiler pectoralis major muscles. Meat Sci. 2011, 89, 181–188. [Google Scholar] [CrossRef]
- Wang, R.H.; Liang, R.R.; Lin, H.; Zhu, L.X.; Zhang, Y.M.; Mao, Y.W.; Dong, P.C.; Niu, L.B.; Zhang, M.H.; Luo, X.X. Effect of acute heat stress and slaughter processing on poultry meat quality and postmortem carbohydrate metabolism. Poult. Sci. 2017, 96, 738–746. [Google Scholar] [CrossRef]
- Sosnicki, A.A.; Greaser, M.L.; Pieterzak, M.; Pospiech, E.; Sante, V. PSE-like syndrome in breast muscle of domestic turkeys: A review. J. Muscle Foods 1998, 9, 13–23. [Google Scholar] [CrossRef]
- Hashizawa, Y.; Kubota, M.; Kadowaki, N.; Fujimura, S. Effect of dietary vitamin E on broiler meat qualities, color, water-holding capacity and shear force value, under heat stress conditions. Anim. Sci. J. 2013, 84, 732–736. [Google Scholar] [CrossRef]
- Petracci, M.; Betti, M.; Cavani, C. Color variation and characterization of broiler breast meat during processing in Italy. Poult. Sci. 2004, 83, 2085–2092. [Google Scholar] [CrossRef]
- Inoue, H.; Simamoto, S.; Takahashi, H.; Kawashima, Y.; Sato, W.; Ijiri, D.; Ohtsuka, A. Effects of astaxanthin-rich dried cell powder from paracoccus carotinifaciens on carotenoid composition and lipid peroxidation in skeletal muscle of broiler chickens under thermo-neutral or realistic high temperature conditions. Anim Sci. J. 2019, 90, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Al-Sagan, A.A.; Khalil, S.; Hussein, E.O.S.; Attia, Y.A. Effects of fennel seed powder supplementation on growth performance, carcass characteristics, meat quality, and economic efficiency of broilers under thermoneutral and chronic heat stress conditions. Animals (Basel) 2020, 10, 206. [Google Scholar] [CrossRef]
- Tomonaga, S.; Okuyama, H.; Tachibana, T.; Makino, R. Effects of high ambient temperature on plasma metabolomic profiles in chicks. Anim. Sci. J. 2018, 89, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Tabiri, H.Y.; Sato, K.; Takahashi, K.; Toyomizu, M.; Akiba, Y. Effects of heat stress and dietary tryptophan on performance and plasma amino acid concentrations of broiler chickens. Asian Austral. J. Anim. 2002, 15, 247–253. [Google Scholar] [CrossRef]
- Del Vesco, A.P.; Gasparino, E.; Grieser, D.O.; Zancanela, V.; Voltolini, D.M.; Khatlab, A.S.; Guimarães, S.E.; Soares, M.A.; Oliveira Neto, A.R. Effects of methionine supplementation on the expression of protein deposition-related genes in acute heat stress-exposed broilers. PLoS ONE 2015, 25, e0115821. [Google Scholar] [CrossRef] [PubMed]
- Löffler, M.; Carrey, E.A.; Zameitat, E. Orotic acid, more than just an intermediate of pyrimidine de novo synthesis. J Genet. Genom. 2015, 42, 207–219. [Google Scholar] [CrossRef]
- Freeman, B.A.; Crapo, J.D. Biology of disease: Free radicals and tissue injury. Lab. Investig. 1982, 47, 412–426. [Google Scholar]
- Liu, D.; Wen, J.; Liu, J.; Li, L. The roles of free radicals in amyotrophic lateral sclerosis: Reactive oxygen species and elevated oxidation of protein, DNA, and membrane phospholipids. FASEB J. 1999, 13, 2318–2328. [Google Scholar] [CrossRef]
- Grashorn, M.A. Functionality of poultry meat. J. Appl. Poult. Res. 2007, 16, 99–106. [Google Scholar] [CrossRef]
- Dai, S.F.; Gao, F.; Zhang, W.H.; Song, S.X.; Xu, X.L.; Zhou, G.H. Effects of dietary glutamine and gamma-aminobutyric acid on performance, carcass characteristics and serum parameters in broilers under circular heat stress. Anim. Feed Sci. Technol. 2011, 168, 51–60. [Google Scholar] [CrossRef]
- Wang, B.; Wu, G.; Zhou, Z.; Dai, Z.; Sun, Y.; Ji, Y.; Li, W.; Wang, W.; Liu, C.; Han, F.; et al. Glutamine and intestinal barrier function. Amino Acids 2015, 47, 2143–2154. [Google Scholar] [CrossRef] [PubMed]
- Nanto-Hara, F.; Kikusato, M.; Ohwada, S.; Toyomizu, M. Heat stress directly affects intestinal integrity in broiler chickens. J. Poult. Sci. 2019. [Google Scholar] [CrossRef]
- Wu, G. Intestinal mucosal amino acid catabolism. Nutr. J. 1998, 128, 1249–1252. [Google Scholar] [CrossRef] [PubMed]
- Ippolito, D.L.; Lewis, J.A.; Yu, C.; Leon, L.R.; Stallings, J.D. Alteration in circulating metabolites during and after heat stress in the conscious rat: Potential biomarkers of exposure and organ-specific injury. BMC Physiol. 2014, 14, 14. [Google Scholar] [CrossRef]
- Ames, B.N.; Cathcart, R.; Schwiers, E.; Hochstein, P. Uric acid provides an antioxidant defense in humans against oxidant- and radical-caused aging and cancer: A hypothesis. Proc. Natl. Acad. Sci. USA 1981, 78, 6858–6862. [Google Scholar] [CrossRef]
- Sevanian, A.; Davies, K.J.; Hochstein, P. Serum urate as an antioxidant for ascorbic acid. Am. J. Clin. Nutr. 1991, 54, 1129S–1134S. [Google Scholar] [CrossRef]
- Yunianto, V.D.; Hayashi, K.; Kaneda, S.; Ohtsuka, A.; Tomita, Y. Effect of environmental temperature on muscle protein turnover and heat production in tube-fed broiler chickens. Br. J. Nutr. 1997, 77, 897–909. [Google Scholar] [CrossRef]
- Dzeja, P.P.; Zeleznikar, R.J.; Goldberg, N.D. Adenylate kinase: Kinetic behavior in intact cells indicates it is integral to multiple cellular processes. Mol. Cell. Biochem. 1998, 184, 169–182. [Google Scholar] [CrossRef]
- Beis, I.; Newsholme, E.A. The contents of adenine nucleotides, phosphagens and some glycolytic intermediates in resting muscles from vertebrates and invertebrates. Biochem. J. 1975, 152, 23–32. [Google Scholar] [CrossRef]
- Miller, J.W.; Nadeau, M.R.; Smith, J.; Smith, D.; Selhub, J. Folate-deficiency-induced homocysteinaemia in rats: Disruption of S-adenosylmethionine’s co-ordinate regulation of homocysteine metabolism. Biochem. J. 1994, 298, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Onderci, M.; Sahin, N.; Gursu, M.F.; Kucuk, O. Dietary vitamin C and folic acid Supplementation ameliorates the detrimental effects of heat stress in Japanese quail. J. Nutr. 2003, 133, 1882–1886. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Onderci, M.; Sahin, N.; Gursu, M.F.; Khachik, F.; Kucuk, O. Effects of lycopene supplementation on antioxidant status, oxidative stress, performance and carcass characteristics in heat-stressed Japanese quail. J. Therm. Biol. 2006, 31, 307–312. [Google Scholar] [CrossRef]
- O’Neill, L.M.; Galvina, K.; Morrisseya, P.A.; Buckleyb, D.J. Inhibition of lipid oxidation in chicken by carnosine and dietary α-tocopherol supplementation and its determination by derivative spectrophotometry. Meat Sci. 1998, 50, 479–488. [Google Scholar] [CrossRef]
- Yang, P.; Hao, Y.; Feng, J.; Lin, H.; Feng, Y.; Wu, X.; Yang, X.; Gu, X. The expression of carnosine and its effect on the antioxidant capacity of longissimus dorsi muscle in finishing pigs exposed to constant heat stress. Asian-Australas. J. Anim. Sci. 2014, 27, 1763–1772. [Google Scholar] [CrossRef]
- Prachayasittikul, S.; Wongsawatkul, O.; Worachartcheewan, A.; Ruchirawat, S.; Prachayasittikul, V. Vasorelaxation and superoxide scavenging activities of orotic acid. Int. J. Pharmacol. 2010, 6, 413–418. [Google Scholar]
- Chadha, R.; Mahal, H.S.; Mukherjee, T.; Kapoor, S. Evidence for a possible role of 3-hydroxyanthranilic acid as an antioxidant. J. Phys. Org. Chem. 2008, 22, 349–354. [Google Scholar] [CrossRef]
- Lu, X.; Li, N.; Qiao, X.; Qiu, Z.; Liu, P. Composition analysis and antioxidant properties of black garlic extract. J. Food Drug Anal. 2017, 25, 340–349. [Google Scholar] [CrossRef]
- Simon, G.; Drori, J.B.; Cohen, M.M. Mechanism of conversion of aspartate into glutamate in cerebral-cortex slices. Biochem. J. 1967, 102, 153–162. [Google Scholar] [CrossRef]
- Hopkins, G.F.; Dixon, M. On glutathione. II. A thermostable oxidation-reduction system. J. Biol. Chem. 1992, 54, 527–563. [Google Scholar]
- Hunter, G.; Eagles, B.A. Glutathione: A critical study. J. Biol. Chem. 1927, 72, 147–166. [Google Scholar]
- Windmueller, H.G.; Spaeth, A.E. Stimulation of hepatic purine biosynthesis by orotic acid. J. Biol. Chem. 1965, 240, 4398–4405. [Google Scholar] [PubMed]
- Tomonaga, T.; Matsumoto, M.; Furuse, M. β-Alanine enhances brain and muscle carnosine levels in broiler chicks. J. Poult. Sci. 2012, 4, 308–312. [Google Scholar] [CrossRef]
- Cong, J.; Zhang, L.; Li, J.; Wang, S.; Gao, F.; Zhou, G. Effects of dietary supplementation with carnosine on meat quality and antioxidant capacity in broiler chickens. Br. Poult. Sci. 2017, 58, 69–75. [Google Scholar] [CrossRef]
- Yagi, K. A simple fluorometric assay for lipoperoxide in blood plasma. Biochem. Med. 1976, 15, 212–216. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Azuma, K.; Hirao, Y.; Hayakawa, Y.; Murahata, Y.; Osaki, T.; Tsuka, T.; Imagawa, T.; Okamoto, Y.; Ito, N. Application of pre-column labeling liquid chromatography for canine plasma-free amino acid analysis. Metabolites 2016, 6, 3. [Google Scholar] [CrossRef]
- Goto, T.; Mori, H.; Shiota, S.; Tomonaga, S. Metabolomics approach reveals the effects of breed and feed on the composition of chicken eggs. Metabolites 2019, 9, 224. [Google Scholar] [CrossRef]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K.; et al. Identifying metabolites by integrating metabolome databases with mass spectrometry cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef]
- Tsugawa, H.; Kanazawa, M.; Ogiwara, A.; Arita, M. MRMPROBS suite for metabolomics using large-scale MRM assays. Bioinformatics 2014, 30, 2379–2380. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 17 December 2019).
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, 486–494. [Google Scholar] [CrossRef] [PubMed]



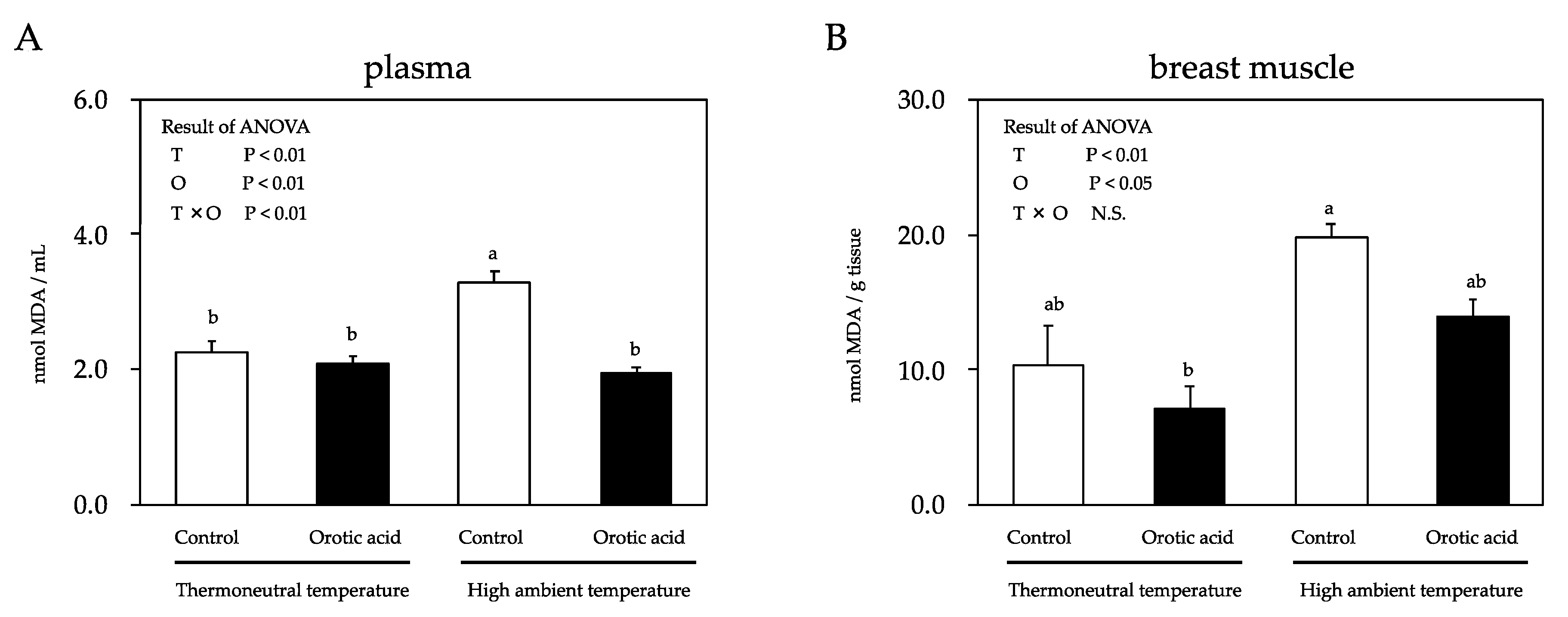{kind=link}
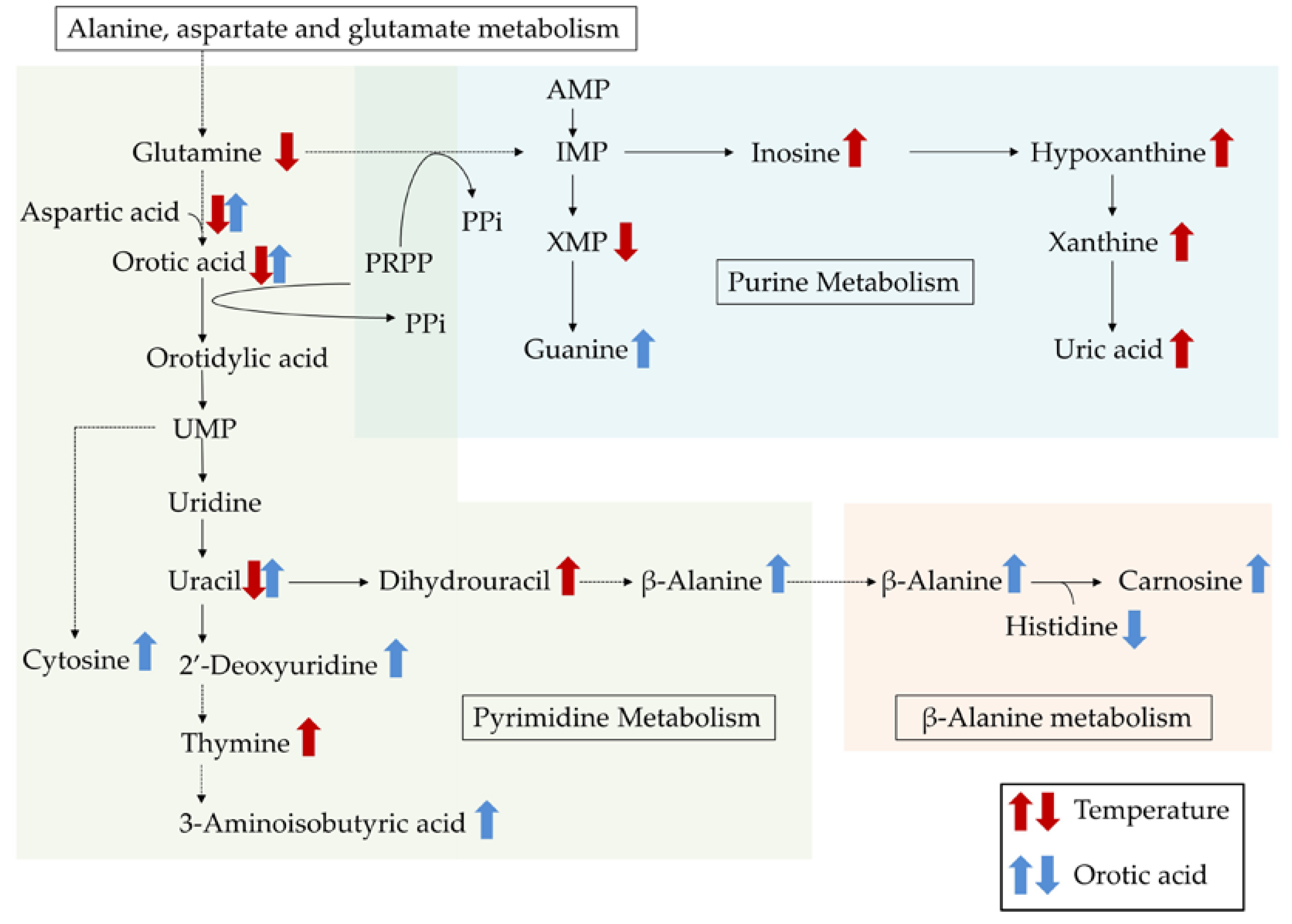{kind=link}
| Thermoneutral Temperature (25 ± 1 °C) | High Ambient Temperature (35 ± 1 °C for 8 h/day) | T | O | T × O | |||
|---|---|---|---|---|---|---|---|
| Control | Orotic Acid | Control | Orotic Acid | ||||
| Final body weight (g) | 1177.96 ± 64.68 | 1175.04 ± 70.06 | 1085.43 ± 33.39 | 1140.79 ± 40.13 | N.S. | N.S. | N.S. |
| Body weight gain (g) | 786.36 ± 64.27 | 777.49 ± 68.44 | 691.74 ± 36.24 | 746.31 ± 35.78 | N.S. | N.S. | N.S. |
| Feed intake (g) | 1216.59 ± 74.66 | 1268.61 ± 101.63 | 1022.19 ± 65.40 | 1118.18 ± 64.43 | <0.05 | N.S. | N.S. |
| Feed conversion ratio | 1.56 ± 0.04 | 1.64 ± 0.05 | 1.48 ± 0.06 | 1.50 ± 0.05 | N.S. | N.S. | N.S. |
| Body temperature (°C) | 40.18 ± 0.07 c | 40.68 ± 0.16 bc | 41.37 ± 0.20 a | 41.13 ± 0.15 ab | <0.001 | N.S. | <0.05 |
| Thermoneutral Temperature (25 ± 1 °C) | High Ambient Temperature (35 ± 1 °C for 8 h/day) | T | O | T × O | |||
|---|---|---|---|---|---|---|---|
| Control | Otrotic Acid | Control | Orotic Acid | ||||
| Breast muscle | 207.91 ± 11.01 ab | 222.19 ± 13.22 a | 182.65 ± 6.23 b | 188.28 ± 4.70 ab | <0.01 | N.S. | N.S. |
| Breast tender muscle | 45.83 ± 2.54 a | 45.98 ± 2.49 a | 40.25 ± 1.21 a | 41.52 ± 1.30 a | <0.05 | N.S. | N.S. |
| Leg muscles | 226.10 ± 16.26 a | 215.90 ± 13.47 a | 216.70 ± 6.56 a | 220.00 ± 8.56 a | N.S. | N.S. | N.S. |
| Liver | 22.20 ± 1.97 ab | 23.41 ± 1.72 a | 17.78 ± 0.65 b | 19.35 ± 1.31 ab | <0.05 | N.S. | N.S. |
| Heart | 6.31 ± 0.54 a | 6.44 ± 0.66 a | 3.83 ± 0.16 b | 4.38 ± 0.34 b | <0.001 | N.S. | N.S. |
| Abdominal fat tissue | 3.88 ± 1.25 b | 4.35 ± 1.00 b | 9.71 ± 1.55 a | 9.83 ± 1.24 a | <0.001 | N.S. | N.S. |
| Thermoneutral Temperature (25 ± 1 °C) | High Ambient Temperature (35 ± 1 °C for 8 h/day) | T | O | T × O | |||
|---|---|---|---|---|---|---|---|
| Control | Orotic Acid | Control | Orotic Acid | ||||
| Aspartic acid | 15.82 ± 1.63 | 19.18 ± 3.13 | 12.00 ± 0.70 | 15.72 ± 2.10 | N.S. | 0.04 | N.S. |
| Glutamic acid | 28.52 ± 6.09 | 36.27 ± 3.15 | 32.13 ± 1.92 | 38.20 ± 3.82 | N.S. | 0.02 | N.S. |
| Asparagine | 17.56 ± 2.33 | 12.83 ± 3.05 | 14.04 ± 1.60 | 13.33 ± 2.06 | N.S. | N.S. | N.S. |
| Serine | 131.48 ± 8.88 ab | 151.03 ± 11.88 a | 98.78 ± 10.14 b | 95.74 ± 6.92 b | <0.001 | N.S. | N.S. |
| Glutamine | 134.46 ± 8.69 a | 116.84 ± 13.86 a | 85.65 ± 10.75 b | 81.62 ± 8.81 b | <0.05 | N.S. | N.S. |
| Histidine | 124.47 ± 11.15 | 151.57 ± 11.35 | 155.82 ± 12.55 | 161.53 ± 17.13 | N.S. | N.S. | N.S. |
| Glycine | 65.28 ± 6.59 | 85.55 ± 9.03 | 73.16 ± 7.27 | 69.77 ± 6.17 | N.S. | N.S. | N.S. |
| Threonine | 11.04 ± 1.05 | 17.10 ± 3.01 | 12.36 ± 1.93 | 14.60 ± 3.79 | N.S. | N.S. | N.S. |
| Arginine | 57.92 ± 5.42 | 58.30 ± 10.47 | 73.64 ± 8.20 | 60.60 ± 7.94 | N.S. | N.S. | N.S. |
| Tyrosine | 30.25 ± 2.00 a | 22.66 ± 2.78 ab | 24.09 ± 4.25 ab | 15.67 ± 1.73 b | <0.05 | <0.01 | N.S. |
| Valine | 21.28 ± 2.46 | 24.30 ± 2.38 | 20.02 ± 2.05 | 18.24 ± 1.49 | N.S. | N.S. | N.S. |
| Methionine | 13.89 ± 1.11 | 13.49 ± 2.35 | 12.27 ± 1.49 | 12.00 ± 1.12 | N.S. | N.S. | N.S. |
| Tryptophan | 19.69 ± 0.99 | 19.19 ± 1.17 | 19.77 ± 1.39 | 18.31 ± 1.01 | N.S. | N.S. | N.S. |
| Phenylalanine | 32.98 ± 3.11 | 33.37 ± 4.31 | 31.49 ± 3.99 | 27.36 ± 1.36 | N.S. | N.S. | N.S. |
| Isoleucine | 44.95 ± 4.43 | 44.59 ± 5.31 | 40.63 ± 5.29 | 35.77 ± 1.26 | N.S. | N.S. | N.S. |
| Leucine | 12.57 ± 1.20 | 12.83 ± 1.47 | 11.59 ± 1.87 | 9.68 ± 0.88 | N.S. | N.S. | N.S. |
| Lysine | 59.65 ± 10.69 | 75.17 ± 13.33 | 83.31 ± 14.47 | 72.88 ± 12.45 | N.S. | N.S. | N.S. |
| Proline | 51.70 ± 5.46 | 55.55 ± 4.10 | 64.98 ± 7.26 | 64.56 ± 6.29 | N.S. | N.S. | N.S. |
| Thermoneutral Temperature (25 ± 1 °C) | High Ambient Temperature (35 ± 1 °C for 8 h/day) | T | O | T × O | |||
|---|---|---|---|---|---|---|---|
| Control | Orotic Acid | Control | Orotic Acid | ||||
| Metabolites affected by temperature | |||||||
| Nicotinic acid | 100 ± 40 ab | 76 ± 26 b | 248 ± 95 ab | 336 ± 94 a | <0.01 | N.S. | N.S. |
| Methionine | 100 ± 25 ab | 136 ± 27 a | 39 ± 7 b | 68 ± 17 ab | <0.01 | N.S. | N.S. |
| Galactosamine | 100 ± 25 a | 92 ± 28 a | 30 ± 10 ab | 20 ± 7 b | <0.01 | N.S. | N.S. |
| Uric acid | 100 ± 28 b | 148 ± 55 ab | 521 ± 161 ab | 621 ± 201 a | <0.01 | N.S. | N.S. |
| Xanthine | 100 ± 23 b | 269 ± 84 ab | 585 ± 176 a | 635 ± 112 a | <0.01 | N.S. | N.S. |
| Xanthosine monophosphate | 100 ± 18 | 103 ± 26 | 61 ± 15 | 48 ± 11 | <0.05 | N.S. | N.S. |
| Oleic acid | 100 ± 29 | 70 ± 22 | 36 ± 13 | 29 ± 9 | <0.05 | N.S. | N.S. |
| Thymine | 100 ± 21 | 94 ± 18 | 155 ± 24 | 199 ± 55 | <0.05 | N.S. | N.S. |
| Aspartic acid | 100 ± 30 | 149 ± 40 | 49 ± 11 | 62 ± 16 | <0.05 | N.S. | N.S. |
| Dihydrouracil | 100 ± 21 | 119 ± 27 | 193 ± 48 | 189 ± 42 | <0.05 | N.S. | N.S. |
| Ascorbic acid | 100 ± 16 | 103 ± 18 | 64 ± 11 | 72 ± 13 | <0.05 | N.S. | N.S. |
| Inosine | 100 ± 22 | 100 ± 18 | 191 ± 62 | 192 ± 49 | <0.05 | N.S. | N.S. |
| Ornithine | 100 ± 26 | 112 ± 22 | 62 ± 11 | 59 ± 16 | <0.05 | N.S. | N.S. |
| 3-Phenyllactic acid | 100 ± 17 | 114 ± 26 | 192 ± 65 | 257 ± 82 | <0.05 | N.S. | N.S. |
| Cysteine | 100 ± 21 | 124 ± 25 | 60 ± 12 | 79 ± 12 | <0.05 | N.S. | N.S. |
| Glutaric acid | 100 ± 25 | 135 ± 32 | 164 ± 44 | 258 ± 70 | <0.05 | N.S. | N.S. |
| 2-Hydroxyglutaric acid | 100 ± 23 | 105 ± 17 | 169 ± 47 | 165 ± 33 | <0.05 | N.S. | N.S. |
| Sucrose | 100 ± 45 | 152 ± 70 | 35 ± 7 | 48 ± 11 | <0.05 | N.S. | N.S. |
| Asparagine | 100 ± 23 | 108 ± 27 | 56 ± 10 | 72 ± 12 | <0.05 | N.S. | N.S. |
| Serine | 100 ± 16 | 110 ± 17 | 77 ± 15 | 68 ± 14 | <0.05 | N.S. | N.S. |
| Hypoxanthine | 100 ± 18 | 123 ± 22 | 145 ± 34 | 182 ± 30 | <0.05 | N.S. | N.S. |
| Metabolites affected by orotic acid | |||||||
| Niacinamide | 100 ± 17 a | 66 ± 12 ab | 95 ± 19 ab | 42 ± 10 b | N.S. | <0.01 | N.S. |
| β-Alanine | 100 ± 25 ab | 191 ± 33 a | 78 ± 18 b | 164 ± 38 ab | N.S. | <0.01 | N.S. |
| Uridine | 100 ± 16 | 167 ± 28 | 90 ± 19 | 171 ± 36 | N.S. | <0.05 | N.S. |
| Guanine | 100 ± 18 | 147 ± 21 | 110 ± 19 | 156 ± 22 | N.S. | <0.05 | N.S. |
| 3-Hydroxyanthranilic acid | 100 ± 27 b | 166 ± 46 ab | 150 ± 22 ab | 271 ± 65 a | N.S. | <0.05 | N.S. |
| Glycerol 3-phosphate | 100 ± 28 | 151 ± 44 | 66 ± 14 | 140 ± 17 | N.S. | <0.05 | N.S. |
| 2’-Deoxyuridine | 100 ± 15 | 152 ± 27 | 107 ± 30 | 171 ± 36 | N.S. | <0.05 | N.S. |
| Cytosine | 100 ± 23 ab | 115 ± 38 ab | 74 ± 13 b | 195 ± 42 a | N.S. | <0.05 | N.S. |
| 3-Aminoisobutyric acid | 100 ± 34 | 189 ± 61 | 100 ± 16 | 269 ± 94 | N.S. | <0.05 | N.S. |
| Phenylacetic acid | 100 ± 29 | 180 ± 55 | 59 ± 9 | 127 ± 35 | N.S. | <0.05 | N.S. |
| Metabolites affected by temperature and orotic acid | |||||||
| Uracil | 100 ± 18 c | 273 ± 55 ab | 173 ± 31 bc | 379 ± 67 a | <0.05 | <0.01 | N.S. |
| Orotic acid | 100 ± 45 ab | 221 ± 30 a | 28 ± 6 b | 160 ± 34 a | <0.05 | <0.01 | N.S. |
| Metabolism Name | p-Value |
|---|---|
| Metabolic pathways affected by high ambient temperature | |
| Purine metabolism | <0.01 |
| Ammonia recycling | <0.01 |
| Pyrimidine metabolism | <0.01 |
| Homocysteine degradation | <0.05 |
| Glutamate metabolism | <0.05 |
| Urea cycle | <0.05 |
| β-Alanine metabolism | <0.05 |
| Glycine and serine metabolism | <0.05 |
| Aspartate metabolism | <0.05 |
| Metabolic pathway affected by orotic acid | |
| Pyrimidine metabolism | <0.01 |
| β-Alanine metabolism | <0.01 |
| Malate–aspartate shuttle | <0.01 |
| Aspartate metabolism | <0.05 |
| Thermoneutral Temperature (25 ± 1 °C) | High Ambient Temperature (35 ± 1 °C for 8 h/day) | T | O | T × O | |||
|---|---|---|---|---|---|---|---|
| Control | Orotic Acid | Control | Orotic Acid | ||||
| Aspartic acid | 5.91 ± 0.78 | 6.99 ± 0.48 | 4.99 ± 0.32 | 5.93 ± 0.73 | N.S. | N.S. | N.S. |
| Glutamic acid | 18.74 ± 3.10 | 19.30 ± 2.69 | 21.29 ± 1.39 | 21.91 ± 1.48 | N.S. | N.S. | N.S. |
| Asparagine | 5.98 ± 0.56 | 4.91 ± 0.21 | 5.25 ± 0.25 | 5.41 ± 0.26 | N.S. | N.S. | N.S. |
| Serine | 21.37 ± 1.60 ab | 22.27 ± 1.59 a | 16.49 ± 1.41 b | 16.76 ± 1.45 b | <0.05 | N.S. | N.S. |
| Glutamine | 25.59 ± 1.47 a | 21.98 ± 2.18 ab | 17.00 ± 1.50 b | 18.84 ± 2.23 b | <0.05 | N.S. | N.S. |
| Histidine | 1.11 ± 0.32 | 0.57 ± 0.08 | 1.31 ± 0.27 | 0.79 ± 0.05 | N.S. | <0.05 | N.S. |
| Glycine | 33.19 ± 11.45 | 31.47 ± 4.48 | 37.68 ± 7.35 | 35.29 ± 4.71 | N.S. | N.S. | N.S. |
| Threonine | 12.37 ± 1.82 | 15.21 ± 1.66 | 14.92 ± 0.53 | 13.97 ± 0.91 | N.S. | N.S. | N.S. |
| Arginine | 9.77 ± 1.25 b | 11.83 ± 1.34 ab | 16.63 ± 2.13 ab | 13.82 ± 1.34 ab | <0.05 | N.S. | N.S. |
| Tyrosine | 5.65 ± 0.94 | 5.82 ± 0.47 | 6.74 ± 0.32 | 4.78 ± 0.24 | N.S. | N.S. | N.S. |
| Valine | 3.23 ± 0.35 | 4.19 ± 0.62 | 4.21 ± 0.28 | 3.82 ± 0.31 | N.S. | N.S. | N.S. |
| Methionine | 1.25 ± 0.27 b | 2.19 ± 0.42 a | 1.57 ± 0.13 b | 1.28 ± 0.25 b | <0.05 | <0.05 | <0.05 |
| Tryptophan | 5.03 ± 0.27 | 5.62 ± 0.93 | 4.98 ± 0.41 | 4.74 ± 0.64 | N.S. | N.S. | N.S. |
| Phenylalanine | 3.60 ± 0.20 | 3.89 ± 0.38 | 4.62 ± 0.29 | 4.24 ± 0.28 | <0.05 | N.S. | N.S. |
| Isoleucine | 2.66 ± 0.22 | 3.11 ± 0.50 | 2.76 ± 0.24 | 2.59 ± 0.26 | N.S. | N.S. | N.S. |
| Leucine | 4.03 ± 0.44 | 4.35 ± 0.59 | 4.02 ± 0.31 | 3.85 ± 0.34 | N.S. | N.S. | N.S. |
| Lysine | 6.30 ± 2.20 | 11.36 ± 1.56 | 11.44 ± 1.58 | 10.38 ± 1.01 | N.S. | N.S. | N.S. |
| Proline | 4.37 ± 0.53 | 4.40 ± 0.35 | 4.23 ± 0.27 | 4.71 ± 0.66 | N.S. | N.S. | N.S. |
| Carnosine | 364.01 ± 69.97 ab | 446.29 ± 41.85 a | 236.35 ± 26.80 b | 437.63 ± 34.67 ab | N.S. | <0.05 | N.S. |
| Ingredients (g/100 g) | |
|---|---|
| Corn meal | 57.90 |
| Soybean meal | 34.00 |
| Corn oil | 4.30 |
| CaCO3 | 0.66 |
| CaHPO4 | 2.00 |
| NaCl | 0.50 |
| DL-Methionine | 0.14 |
| Mineral and vitamin premix 1 | 0.50 |
| Calculated analysis | |
| Crude protein (%) | 20.00 |
| Metabolizable energy (Mcal/kg) | 3.10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimamoto, S.; Nakamura, K.; Tomonaga, S.; Furukawa, S.; Ohtsuka, A.; Ijiri, D. Effects of Cyclic High Ambient Temperature and Dietary Supplementation of Orotic Acid, a Pyrimidine Precursor, on Plasma and Muscle Metabolites in Broiler Chickens. Metabolites 2020, 10, 189. https://doi.org/10.3390/metabo10050189
Shimamoto S, Nakamura K, Tomonaga S, Furukawa S, Ohtsuka A, Ijiri D. Effects of Cyclic High Ambient Temperature and Dietary Supplementation of Orotic Acid, a Pyrimidine Precursor, on Plasma and Muscle Metabolites in Broiler Chickens. Metabolites. 2020; 10(5):189. https://doi.org/10.3390/metabo10050189
Chicago/Turabian StyleShimamoto, Saki, Kiriko Nakamura, Shozo Tomonaga, Satoru Furukawa, Akira Ohtsuka, and Daichi Ijiri. 2020. "Effects of Cyclic High Ambient Temperature and Dietary Supplementation of Orotic Acid, a Pyrimidine Precursor, on Plasma and Muscle Metabolites in Broiler Chickens" Metabolites 10, no. 5: 189. https://doi.org/10.3390/metabo10050189
APA StyleShimamoto, S., Nakamura, K., Tomonaga, S., Furukawa, S., Ohtsuka, A., & Ijiri, D. (2020). Effects of Cyclic High Ambient Temperature and Dietary Supplementation of Orotic Acid, a Pyrimidine Precursor, on Plasma and Muscle Metabolites in Broiler Chickens. Metabolites, 10(5), 189. https://doi.org/10.3390/metabo10050189