Metabolomic Profiling of Wildtype and Transgenic Giardia lamblia Strains by 1H HR-MAS NMR Spectroscopy



Abstract

1. Introduction
2. Results and Discussion
2.1. Relative Amounts of Identified Metabolites
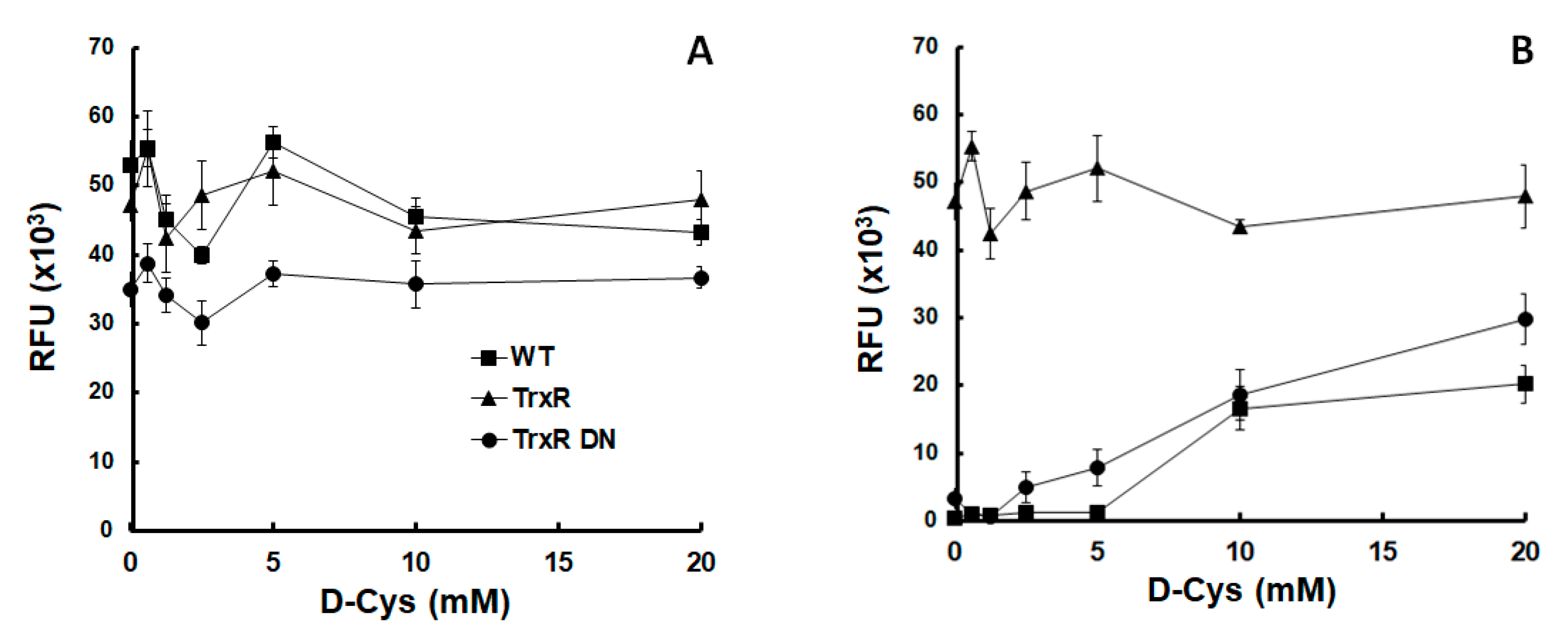
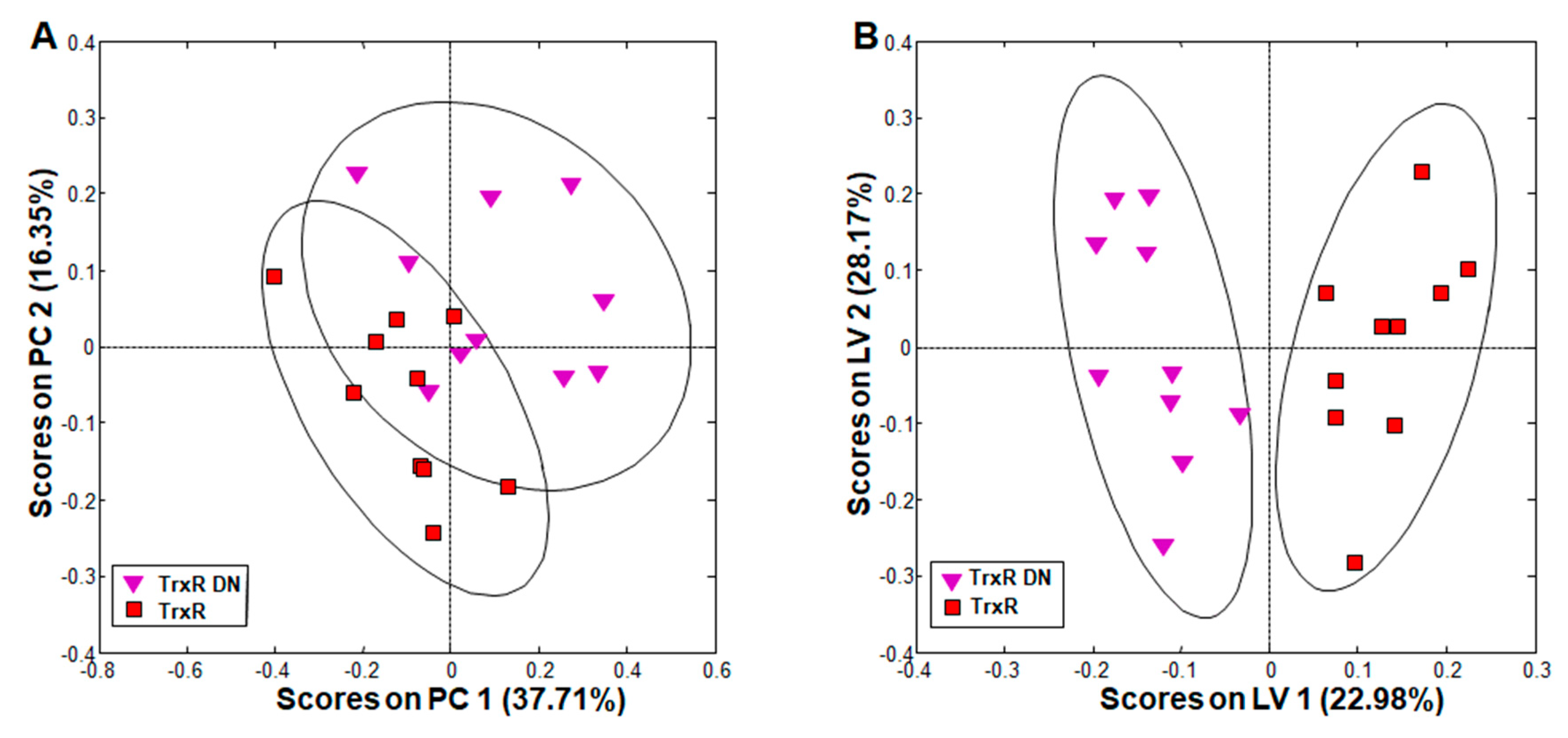
2.2. Phenotype and Metabolite Patterns Related to Thioredoxin Reductase Overexpression
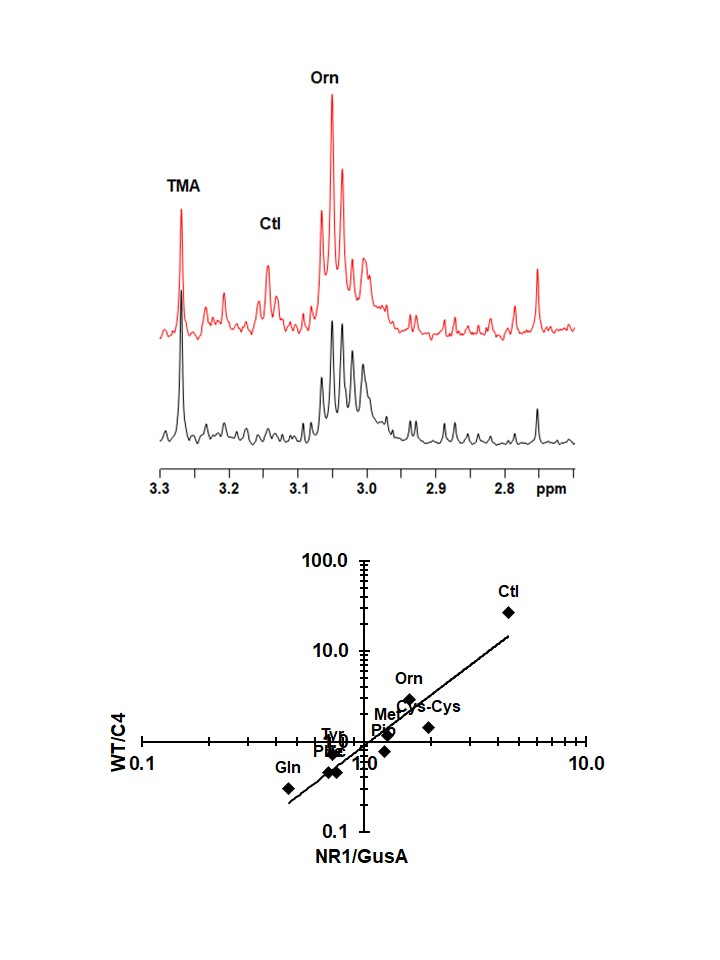
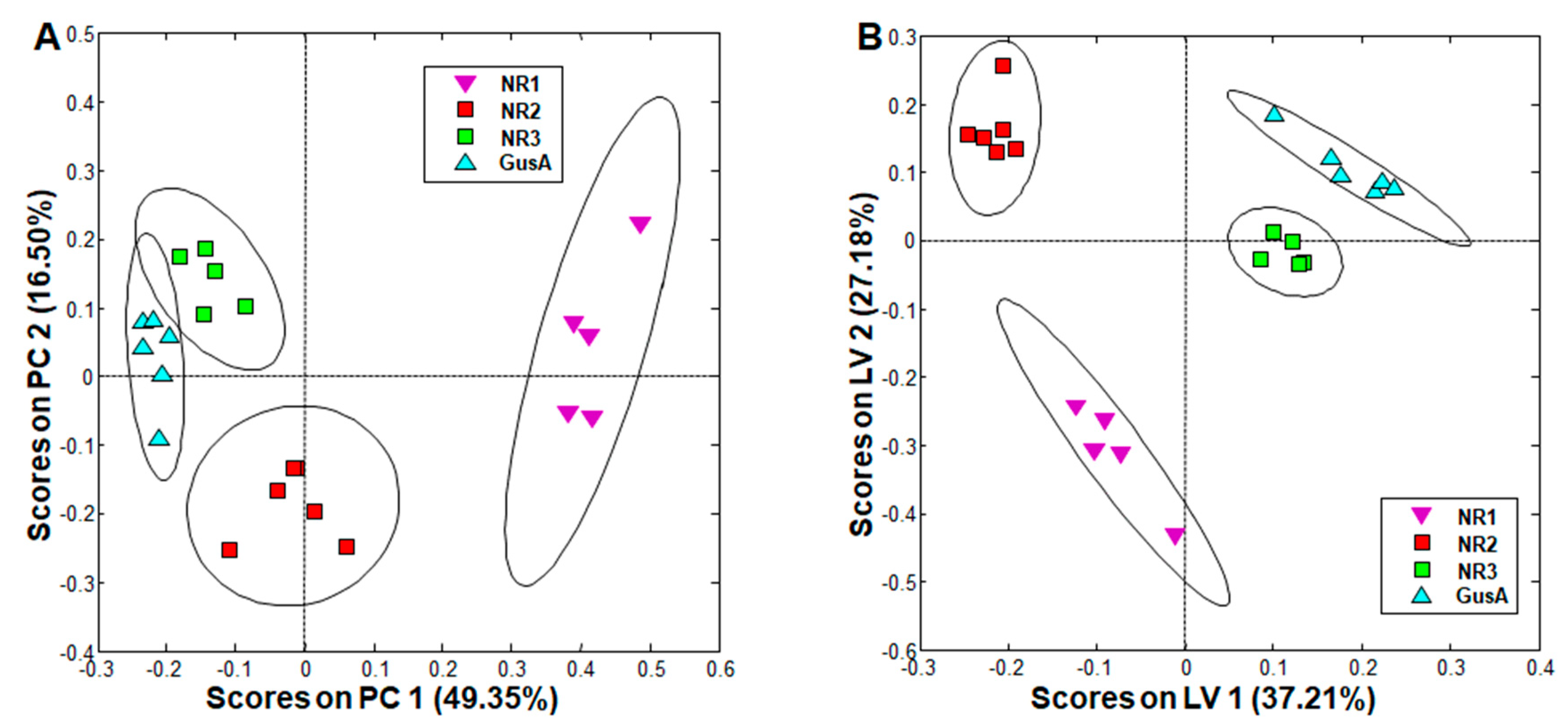
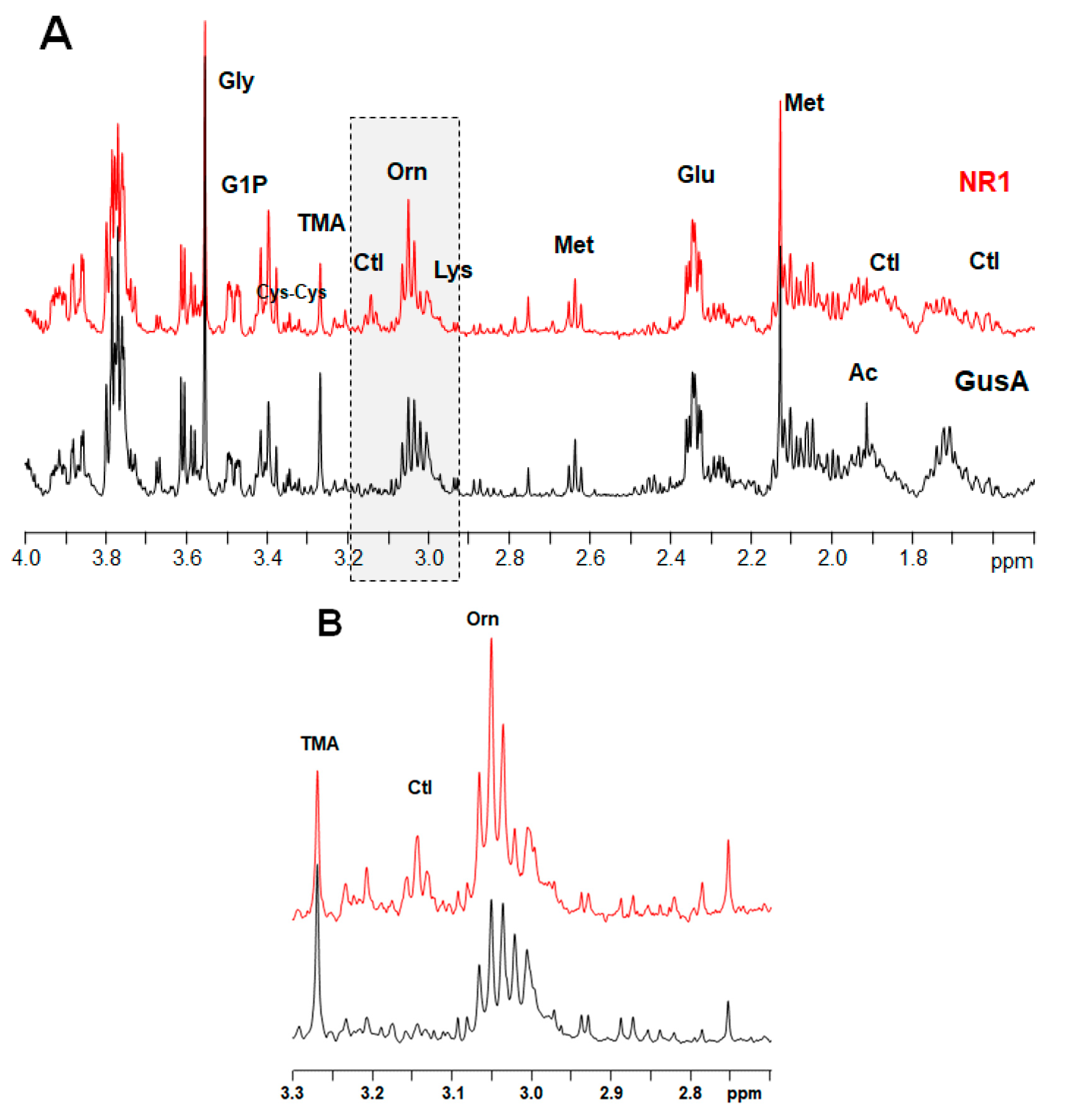
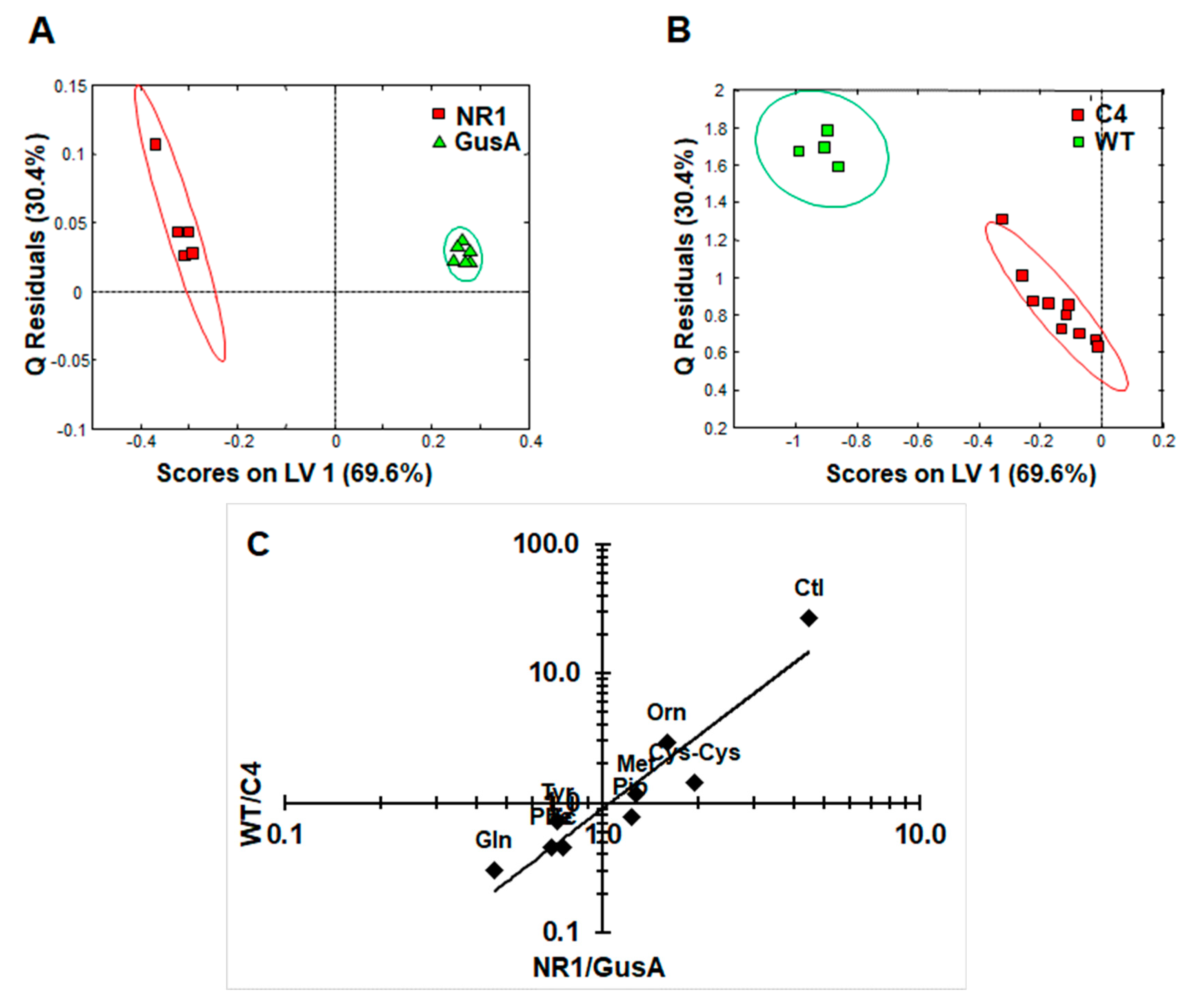
2.3. Metabolite Patterns Related to Nitroreductase Overexpression
2.4. Comparison of the NR1 Pattern with the Situation in Nitro Drug-Resistant Trophozoites
3. Conclusions
4. Materials and Methods
4.1. Biochemicals
4.2. Growth of Thioredoxin Reductase Overexpressors
4.3. Preparation of Cell Samples for HR-MAS NMR
4.4. HR-MAS NMR Spectroscopy and Processing of Spectra
4.5. Bioinformatics and Statistical Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cernikova, L.; Faso, C.; Hehl, A.B. Five facts about Giardia lamblia. PLoS Pathog. 2018, 14, e1007250. [Google Scholar] [CrossRef]
- Hemphill, A.; Müller, N.; Müller, J. Comparative pathobiology of the intestinal protozoan parasites Giardia lamblia, Entamoeba histolytica, and Cryptosporidium parvum. Pathogens 2019, 8, 116. [Google Scholar] [CrossRef] [PubMed]
- Morrison, H.G.; McArthur, A.G.; Gillin, F.D.; Aley, S.B.; Adam, R.D.; Olsen, G.J.; Best, A.A.; Cande, W.Z.; Chen, F.; Cipriano, M.J.; et al. Genomic minimalism in the early diverging intestinal parasite Giardia lamblia. Science 2007, 317, 1921–1926. [Google Scholar] [CrossRef] [PubMed]
- Nixon, J.E.; Wang, A.; Field, J.; Morrison, H.G.; McArthur, A.G.; Sogin, M.L.; Loftus, B.J.; Samuelson, J. Evidence for lateral transfer of genes encoding ferredoxins, nitroreductases, NADH oxidase, and alcohol dehydrogenase 3 from anaerobic prokaryotes to Giardia lamblia and Entamoeba histolytica. Eukaryot. Cell 2002, 1, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Ma’ayeh, S.Y.; Knorr, L.; Svard, S.G. Transcriptional profiling of Giardia intestinalis in response to oxidative stress. Int. J. Parasitol. 2015, 45, 925–938. [Google Scholar] [CrossRef]
- Brown, D.M.; Upcroft, J.A.; Edwards, M.R.; Upcroft, P. Anaerobic bacterial metabolism in the ancient eukaryote Giardia duodenalis. Int. J. Parasitol. 1998, 28, 149–164. [Google Scholar] [CrossRef]
- Schofield, P.J.; Costello, M.; Edwards, M.R.; O’Sullivan, W.J. The arginine dihydrolase pathway is present in Giardia intestinalis. Int. J. Parasitol. 1990, 20, 697–699. [Google Scholar] [CrossRef]
- Edwards, M.R.; Schofield, P.J.; O’Sullivan, W.J.; Costello, M. Arginine metabolism during culture of Giardia intestinalis. Mol. Biochem. Parasitol. 1992, 53, 97–103. [Google Scholar] [CrossRef]
- Schofield, P.J.; Edwards, M.R.; Matthews, J.; Wilson, J.R. The pathway of arginine catabolism in Giardia intestinalis. Mol. Biochem. Parasitol. 1992, 51, 29–36. [Google Scholar] [CrossRef]
- Nash, T.E. Treatment of Giardia lamblia infections. Pediatr. Infect. Dis. J. 2001, 20, 193–195. [Google Scholar] [CrossRef]
- Leitsch, D. A review on metronidazole: An old warhorse in antimicrobial chemotherapy. Parasitology 2017, 146, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Hemphill, A. New approaches for the identification of drug targets in protozoan parasites. Int Rev. Cell Mol. Biol. 2013, 301, 359–401. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, D.; Harris, J.C.; Maroulis, S.; Mitchell, A.; Hughes, M.N.; Wadley, R.B.; Edwards, M.R. Nitrosative stress induced cytotoxicity in Giardia intestinalis. J. Appl. Microbiol. 2003, 95, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Müller, N.; Müller, J. Giardia . In Molecular Parasitology; Walochnik, J., Duchêne, M., Eds.; Springer: Vienna, Austria, 2016; pp. 93–114. [Google Scholar]
- Leitsch, D.; Burgess, A.G.; Dunn, L.A.; Krauer, K.G.; Tan, K.; Duchêne, M.; Upcroft, P.; Eckmann, L.; Upcroft, J.A. Pyruvate:ferredoxin oxidoreductase and thioredoxin reductase are involved in 5-nitroimidazole activation while flavin metabolism is linked to 5-nitroimidazole resistance in Giardia lamblia. J. Antimicrob. Chemother. 2011, 66, 1756–1765. [Google Scholar] [CrossRef] [PubMed]
- Dan, M.; Wang, A.L.; Wang, C.C. Inhibition of pyruvate-ferredoxin oxidoreductase gene expression in Giardia lamblia by a virus-mediated hammerhead ribozyme. Mol. Microbiol. 2000, 36, 447–456. [Google Scholar] [CrossRef]
- Müller, J.; Sterk, M.; Hemphill, A.; Müller, N. Characterization of Giardia lamblia WB C6 clones resistant to nitazoxanide and to metronidazole. J. Antimicrob. Chemother. 2007, 60, 280–287. [Google Scholar] [CrossRef]
- Müller, J.; Hemphill, A.; Müller, N. Physiological aspects of nitro drug resistance in Giardia lamblia. Int, J. Parasitol. Drugs Drug Resist. 2018, 8, 271–277. [Google Scholar] [CrossRef]
- Nillius, D.; Müller, J.; Müller, N. Nitroreductase (GlNR1) increases susceptibility of Giardia lamblia and Escherichia coli to nitro drugs. J. Antimicrob. Chemother. 2011, 66, 1029–1035. [Google Scholar] [CrossRef]
- Ansell, B.R.; Baker, L.; Emery, S.J.; McConville, M.J.; Svard, S.G.; Gasser, R.B.; Jex, A.R. Transcriptomics indicates active and passive metronidazole resistance mechanisms in three seminal Giardia lines. Front. Microbiol. 2017, 8, 398. [Google Scholar] [CrossRef]
- Müller, J.; Schildknecht, P.; Müller, N. Metabolism of nitro drugs metronidazole and nitazoxanide in Giardia lamblia: Characterization of a novel nitroreductase (GlNR2). J. Antimicrob. Chemother. 2013, 68, 1781–1789. [Google Scholar] [CrossRef]
- Müller, J.; Rout, S.; Leitsch, D.; Vaithilingam, J.; Hehl, A.; Müller, N. Comparative characterisation of two nitroreductases from Giardia lamblia as potential activators of nitro compounds. Int. J. Parasitol. Drugs Drug Resist. 2015, 5, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Müller, N. Nitroreductases of bacterial origin in Giardia lamblia: Potential role in detoxification of xenobiotics. MicrobiologyOpen 2019, 8, e904. [Google Scholar] [CrossRef] [PubMed]
- Leitsch, D.; Müller, J.; Müller, N. Evaluation of Giardia lamblia thioredoxin reductase as drug activating enzyme and as drug target. Int. J. Parasitol. Drugs Drug Resist. 2016, 6, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Braga, S.; Heller, M.; Müller, N. Resistance formation to nitro drugs in Giardia lamblia: No common markers identified by comparative proteomics. Int. J. Parasitol. Drugs Drug Resist. 2019, 9, 112–119. [Google Scholar] [CrossRef]
- Vermathen, M.; Müller, J.; Furrer, J.; Müller, N.; Vermathen, P. 1H HR-MAS NMR spectroscopy to study the metabolome of the protozoan parasite Giardia lamblia. Talanta 2018, 188, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Power, W.P. High-Resolution Magic Angle Spinning-enabling applications of NMR spectroscopy to semi-solid phases. Annu Rep. NMR Spectro. 2010, 72, 111–156. [Google Scholar] [CrossRef]
- Dietz, C.; Ehret, F.; Palmas, F.; Vandergrift, L.A.; Jiang, Y.; Schmitt, V.; Dufner, V.; Habbel, P.; Nowak, J.; Cheng, L.L. Applications of high-resolution magic angle spinning MRS in biomedical studies II-Human diseases. NMR Biomed. 2017, 30, e3784. [Google Scholar] [CrossRef]
- Kaebisch, E.; Fuss, T.L.; Vandergrift, L.A.; Toews, K.; Habbel, P.; Cheng, L.L. Applications of high-resolution magic angle spinning MRS in biomedical studies I-cell line and animal models. NMR Biomed. 2017, 30, e3700. [Google Scholar] [CrossRef]
- Santos, A.D.C.; Fonseca, F.A.; Liao, L.M.; Alcantara, G.B.; Barison, A. High-resolution magic angle spinning nuclear magnetic resonance in foodstuff analysis. Trac-Trend Anal. Chem. 2015, 73, 10–18. [Google Scholar] [CrossRef]
- Vermathen, M.; Marzorati, M.; Baumgartner, D.; Good, C.; Vermathen, P. Investigation of different apple cultivars by High Resolution Magic Angle Spinning NMR. A feasibility study. J. Agr. Food Chem. 2011, 59, 12784–12793. [Google Scholar] [CrossRef]
- Vermathen, M.; Marzorati, M.; Diserens, G.; Baumgartner, D.; Good, C.; Gasser, F.; Vermathen, P. Metabolic profiling of apples from different production systems before and after controlled atmosphere (CA) storage studied by 1H high resolution-magic angle spinning (HR-MAS) NMR. Food Chem. 2017, 233, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Duarte, I.F.; Ladeirinha, A.F.; Lamego, I.; Gil, A.M.; Carvalho, L.; Carreira, I.M.; Melo, J.B. Potential markers of cisplatin treatment response unveiled by NMR metabolomics of human lung cells. Mol. Pharm. 2013, 10, 4242–4251. [Google Scholar] [CrossRef] [PubMed]
- Lamego, I.; Duarte, I.F.; Marques, M.P.; Gil, A.M. Metabolic markers of MG-63 osteosarcoma cell line response to doxorubicin and methotrexate treatment: Comparison to cisplatin. J. Proteome Res. 2014, 13, 6033–6045. [Google Scholar] [CrossRef] [PubMed]
- Vermathen, M.; Paul, L.E.; Diserens, G.; Vermathen, P.; Furrer, J. 1H HR-MAS NMR Based Metabolic Profiling of Cells in Response to Treatment with a Hexacationic Ruthenium Metallaprism as Potential Anticancer Drug. PLoS ONE 2015, 10, e0128478. [Google Scholar] [CrossRef]
- Brogi, S.; Fiorillo, A.; Chemi, G.; Butini, S.; Lalle, M.; Ilari, A.; Gemma, S.; Campiani, G. Structural characterization of Giardia duodenalis thioredoxin reductase (gTrxR) and computational analysis of its interaction with NBDHEX. Eur. J. Med. Chem. 2017, 135, 479–490. [Google Scholar] [CrossRef]
- Takei, M.; Ando, Y.; Saitoh, W.; Tanimoto, T.; Kiyosawa, N.; Manabe, S.; Sanbuissho, A.; Okazaki, O.; Iwabuchi, H.; Yamoto, T.; et al. Ethylene glycol monomethyl ether-induced toxicity is mediated through the inhibition of flavoprotein dehydrogenase enzyme family. Toxicol. Sci. 2010, 118, 643–652. [Google Scholar] [CrossRef]
- Morris, S.M., Jr. Recent advances in arginine metabolism. Curr. Opin. Clin. Nutr. Metab. Care. 2004, 7, 45–51. [Google Scholar] [CrossRef]
- Knodler, L.A.; Sekyere, E.O.; Stewart, T.S.; Schofield, P.J.; Edwards, M.R. Cloning and expression of a prokaryotic enzyme, arginine deiminase, from a primitive eukaryote Giardia intestinalis. J. Biol. Chem. 1998, 273, 4470–4477. [Google Scholar] [CrossRef]
- Touz, M.C.; Rópolo, A.S.; Rivero, M.R.; Vranych, C.V.; Conrad, J.T.; Svard, S.G.; Nash, T.E. Arginine deiminase has multiple regulatory roles in the biology of Giardia lamblia. J. Cell Sci. 2008, 121, 2930–2938. [Google Scholar] [CrossRef]
- Ropolo, A.S.; Feliziani, C.; Touz, M.C. Unusual proteins in Giardia duodenalis and their role in survival. Adv. Parasitol. 2019, 106, 1–50. [Google Scholar] [CrossRef]
- Ortega-Pierres, M.G.; Arguello-Garcia, R. Giardia duodenalis: Role of secreted molecules as virulent factors in the cytotoxic effect on epithelial cells. Adv. Parasitol. 2019, 106, 129–169. [Google Scholar] [CrossRef] [PubMed]
- Bringaud, F.; Ebikeme, C.; Boshart, M. Acetate and succinate production in amoebae, helminths, diplomonads, trichomonads and trypanosomatids: Common and diverse metabolic strategies used by parasitic lower eukaryotes. Parasitology 2010, 137, 1315–1331. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Nillius, D.; Hehl, A.; Hemphill, A.; Müller, N. Stable expression of Escherichia coli beta-glucuronidase A (GusA) in Giardia lamblia: Application to high-throughput drug susceptibility testing. J. Antimicrob. Chemother 2009, 64, 1187–1191. [Google Scholar] [CrossRef] [PubMed]
- Bénéré, E.; da Luz, R.A.; Vermeersch, M.; Cos, P.; Maes, L. A new quantitative in vitro microculture method for Giardia duodenalis trophozoites. J. Microbiol. Methods 2007, 71, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Su, L.H.; Lee, G.A.; Huang, Y.C.; Chen, Y.H.; Sun, C.H. Neomycin and puromycin affect gene expression in Giardia lamblia stable transfection. Mol. Biochem Parasitol 2007, 156, 124–135. [Google Scholar] [CrossRef]
- R_Core_Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in 1H NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef]
- Van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef]
- Knol, M.J.; Pestman, W.R.; Grobbee, D.E. The (mis)use of overlap of confidence intervals to assess effect modification. Eur. J. Epidemiol 2011, 26, 253–254. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Characteristics | Accession | References |
|---|---|---|---|
| WBC6 | Wildtype strain (WT) | No transgene | |
| C4 | Nitro drug-resistant strain derived from WT | No transgene | |
| GusA | WT overexpressing E. coli glucuronidase A | E. coli K-12 CDS 1785 074 –1785 074 | |
| NR1 | WT overexpressing G. lamblia Fd-NR2 (“NR1”) Higher susceptibility to nitro drugs in G. lamblia and E. coli | Giardia DB 22677 | [19,23] |
| NR2 | WT overexpressing G. lamblia Fd-NR1 (“NR2”) Better resistance to nitro drugs in E. coli | Giardia DB 6175 | [21,23] |
| NR3 | WT overexpressing G. lamblia NR family protein (“NR3”) | Giardia DB 15307 | [23] |
| TrxR | WT overexpressing G. lamblia TrxR wildtype | Giardia DB 9827 | [24] |
| TrxR DN | WT overexpressing G. lamblia TrxR-dominant negative allele Higher susceptibility to oxygen | Giardia DB 9827 | [24] This work |
| Metabolite | WT 1 n = 10 | WT 2 n = 4 | TrxR n = 10 | TrxR_DN n = 10 | GusA n = 6 | NR1 n = 5 | NR2 n = 6 | NR3 n = 5 |
|---|---|---|---|---|---|---|---|---|
| Carbohydrates and related | ||||||||
| Acetate | 0.77 ± 0.05 | 0.85 ± 0.03 | 0.57 ± 0.03 | 0.65 ± 0.04 * | 0.65 ± 0.02 | 0.49 ± 0.02 * | 0.58 ± 0.02 + | 0.58 ± 0.02 + |
| Citrate | 0.17 ± 0.07 | 0.20 ± 0.01 | 0.18 ± 0.09 | 0.24 ± 0.06 | 0.11 ± 0.05 | 0.32 ± 0.10 | 0.18 ± 0.08 | 0.16 ± 0.05 |
| Glucose-1-phosphate | 4.46 ± 0.65 | 1.65 ± 0.1 * | 4.59 ± 0.40 | 5.16 ± 0.66 | 1.69 ± 0.19 | 2.31 ± 0.13 ° | 1.71 ± 0.24 | 1.88 ± 0.14 |
| Amino acids | ||||||||
| Alanine | 6.02 ± 0.77 | 7.63 ± 0.33 * | 4.88 ± 0.88 | 5.20 ± 0.79 | 6.95 ± 0.30 | 5.40 ± 0.25 | 6.00 ± 0.48 | 6.80 ± 0.36 |
| Asparagine | 0.18 ± 0.02 | 0.26 ± 0.03 * | 0.19 ± 0.04 | 0.22 ± 0.03 | 0.22 ± 0.02 | 0.18 ± 0.07 | 0.21 ± 0.03 | 0.26 ± 0.02 |
| Citrulline | 0.19 ± 0.09 | 0.26 ± 0.09 | 0.14 ± 0.11 | 0.18 ± 0.05 | 0.26 ± 0.07 | 1.16 ± 0.14 * | 0.46 ± 0.18 | 0.24 ± 0.13 |
| Cysteine | 0.56 ± 0.12 | 0.29 ± 0.07 + | 0.40 ± 0.13 | 0.32 ± 0.04 | 0.22 ± 0.09 | 0.38 ± 0.07 | 0.29 ± 0.15 | 0.29 ± 0.14 |
| Cystine (Cys-Cys) | 0.12 ± 0.04 | 0.13 ± 0.03 | 0.12 ± 0.05 | 0.15 ± 0.04 | 0.24 ± 0.05 | 0.47 ± 0.04 + | 0.42 ± 0.06 + | 0.27 ± 0.04 |
| Glutamate | 7.03 ± 0.28 | 4.94 ± 0.31 * | 7.27 ± 0.25 | 7.12 ± 0.24 | 5.32 ± 0.15 | 5.35 ± 0.15 | 5.14 ± 0.12 | 5.41 ± 0.20 |
| Glutamine | 0.48 ± 0.12 | 0.72 ± 0.24 | 0.45 ± 0.14 | 0.48 ± 0.09 | 0.94 ± 0.10 | 0.34 ± 0.09 * | 0.66 ± 0.32 | 0.91 ± 0.11 |
| Glycine | 2.18 ± 0.10 | 2.85 ± 0.03 | 2.38 ± 0.09 | 2.21 ± 0.12 | 2.86 ± 0.06 | 2.40 ± 0.11 | 2.65 ± 0.03 | 2.96 ± 0.07 |
| Histidine | 0.19 ± 0.04 | 0.28 ± 0.02 | 0.14 ± 0.04 | 0.16 ± 0.04 | 0.26 ± 0.02 | 0.14 ± 0.05 | 0.21 ± 0.03 | 0.22 ± 0.04 |
| Isoleucine | 0.42 ± 0.18 | 0.68 ± 0.15 | 0.35 ± 0.10 | 0.35 ± 0.10 | 0.72 ± 0.12 | 0.68 ± 0.04 | 0.88 ± 0.11 | 0.77 ± 0.08 |
| Leucine | 4.27 ± 0.18 | 5.89 ± 0.10 * | 3.92 ± 0.16 | 3.76 ± 0.17 | 5.95 ± 0.06 | 4.09 ± 0.18 | 5.52 ± 0.15 | 5.19 ± 0.14 |
| Lysine | 4.67 ± 0.36 | 4.26 ± 0.18 | 5.45 ± 0.24 | 5.02 ± 0.24 ° | 4.37 ± 0.27 | 4.68 ± 0.18 | 4.08 ± 0.24 | 4.33 ± 0.25 |
| Methionine | 0.95 ± 0.08 | 1.06 ± 0.07 | 1.02 ± 0.05 | 0.98 ± 0.05 | 1.02 ± 0.05 | 1.30 ± 0.04 ° | 1.15 ± 0.05 | 1.09 ± 0.05 |
| Ornithine | 1.84 ± 0.13 | 1.71 ± 0.03 | 1.90 ± 0.11 | 1.72 ± 0.13 | 1.66 ± 0.02 | 2.65 ± 0.17 * | 1.92 ± 0.12 ° | 1.81 ± 0.08 |
| Phenylalanine | 0.30 ± 0.02 | 0.33 ± 0.02 | 0.28 ± 0.02 | 0.27 ± 0.04 | 0.29 ± 0.01 | 0.20 ± 0.04 + | 0.25 ± 0.04 | 0.28 ± 0.02 |
| Pipecolic acid | 0.64 ± 0.08 | 0.69 ± 0.12 | 0.73 ± 0.08 | 0.72 ± 0.11 | 1.00 ± 0.04 | 1.23 ± 0.06 * | 1.07 ± 0.07 | 1.00 ± 0.10 |
| Proline | 0.56 ± 0.12 | 0.40 ± 0.22 | 0.50 ± 0.12 | 0.50 ± 0.12 | 0.68 ± 0.08 | 0.54 ± 0.05 | 0.64 ± 0.17 | 0.66 ± 0.09 |
| Threonine | 2.96 ± 0.12 | 2.61 ± 0.07 + | 3.27 ± 0.12 | 3.03 ± 0.14 | 2.75 ± 0.11 | 2.50 ± 0.07 | 2.83 ± 0.14 | 3.00 ± 0.07 |
| Tryptophan | 0.11 ± 0.03 | 0.07 ± 0.03 | 0.09 ± 0.02 | 0.10 ± 0.03 | 0.05 ± 0.02 | 0.04 ± 0.02 | 0.08 ± 0.04 | 0.03 ± 0.01 |
| Tyrosine | 0.38 ± 0.04 | 0.37 ± 0.02 | 0.37 ± 0.02 | 0.34 ± 0.02 | 0.36 ± 0.02 | 0.26 ± 0.02 * | 0.32 ± 0.07 | 0.36 ± 0.02 |
| Valine | 4.24 ± 0.19 | 5.85 ± 0.08 * | 4.72 ± 0.20 | 4.46 ± 0.26 | 5.87 ± 0.13 | 5.27 ± 0.19 | 5.94 ± 0.25 | 5.93 ± 0.16 |
| Cofactors | ||||||||
| Adenosine triphosphate | 0.25 ± 0.03 | 0.22 ± 0.02 | 0.27 ± 0.04 | 0.25 ± 0.03 | 0.24 ± 0.02 | 0.26 ± 0.02 | 0.30 ± 0.05 | 0.25 ± 0.02 |
| Nicotinamide-adenine-dinucleotide | 0.08 ± 0.03 | 0.06 ± 0.01 | 0.08 ± 0.01 | 0.08 ± 0.02 | 0.07 ± 0.03 | 0.07 ± 0.02 | 0.08 ± 0.03 | 0.08 ± 0.01 |
| Other | ||||||||
| Trimethylamine | 1.18 ± 0.04 | 1.09 ± 0.02 + | 1.16 ± 0.06 | 1.16 ± 0.06 | 1.12 ± 0.03 | 0.71 ± 0.03 * | 0.87 ± 0.08 ° | 1.04 ± 0.03 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, J.; Vermathen, M.; Leitsch, D.; Vermathen, P.; Müller, N. Metabolomic Profiling of Wildtype and Transgenic Giardia lamblia Strains by 1H HR-MAS NMR Spectroscopy. Metabolites 2020, 10, 53. https://doi.org/10.3390/metabo10020053
Müller J, Vermathen M, Leitsch D, Vermathen P, Müller N. Metabolomic Profiling of Wildtype and Transgenic Giardia lamblia Strains by 1H HR-MAS NMR Spectroscopy. Metabolites. 2020; 10(2):53. https://doi.org/10.3390/metabo10020053
Chicago/Turabian StyleMüller, Joachim, Martina Vermathen, David Leitsch, Peter Vermathen, and Norbert Müller. 2020. "Metabolomic Profiling of Wildtype and Transgenic Giardia lamblia Strains by 1H HR-MAS NMR Spectroscopy" Metabolites 10, no. 2: 53. https://doi.org/10.3390/metabo10020053
APA StyleMüller, J., Vermathen, M., Leitsch, D., Vermathen, P., & Müller, N. (2020). Metabolomic Profiling of Wildtype and Transgenic Giardia lamblia Strains by 1H HR-MAS NMR Spectroscopy. Metabolites, 10(2), 53. https://doi.org/10.3390/metabo10020053