Salivary Metabolomics: From Diagnostic Biomarker Discovery to Investigating Biological Function


Abstract
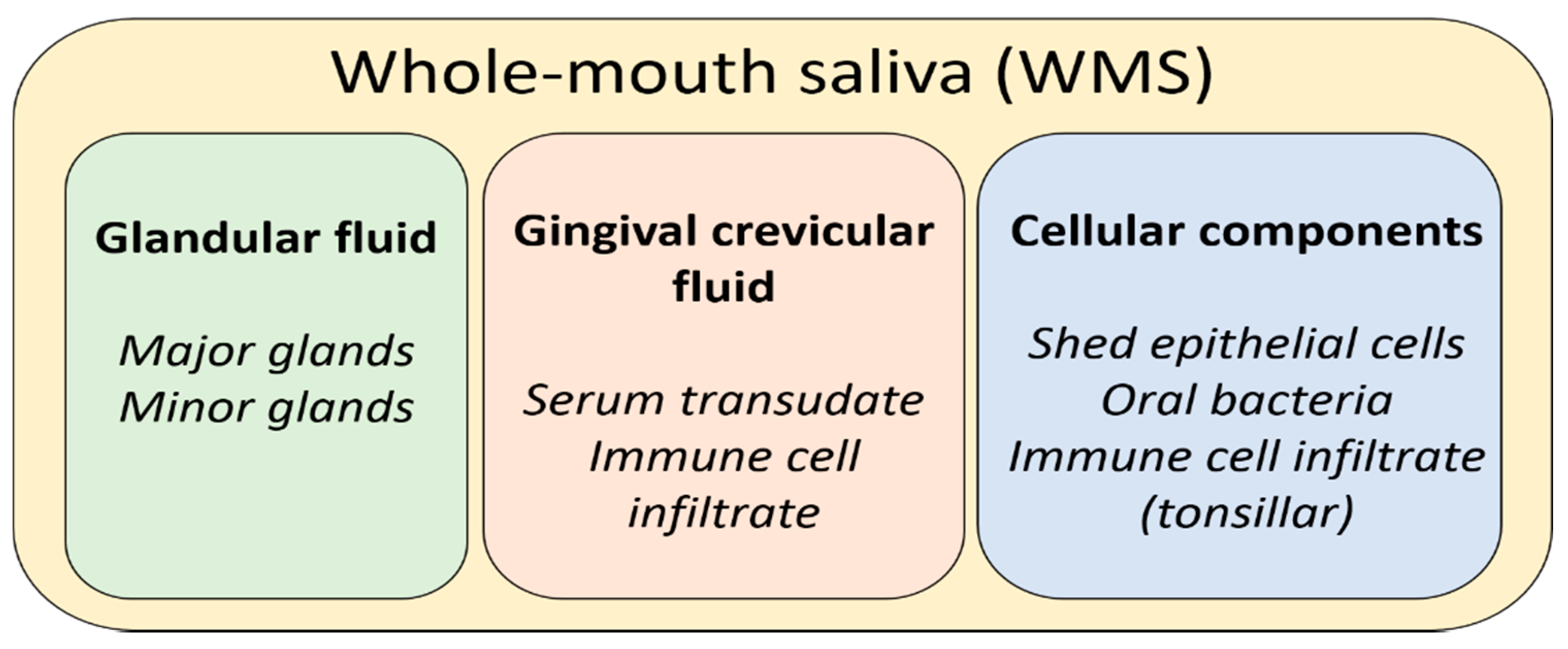
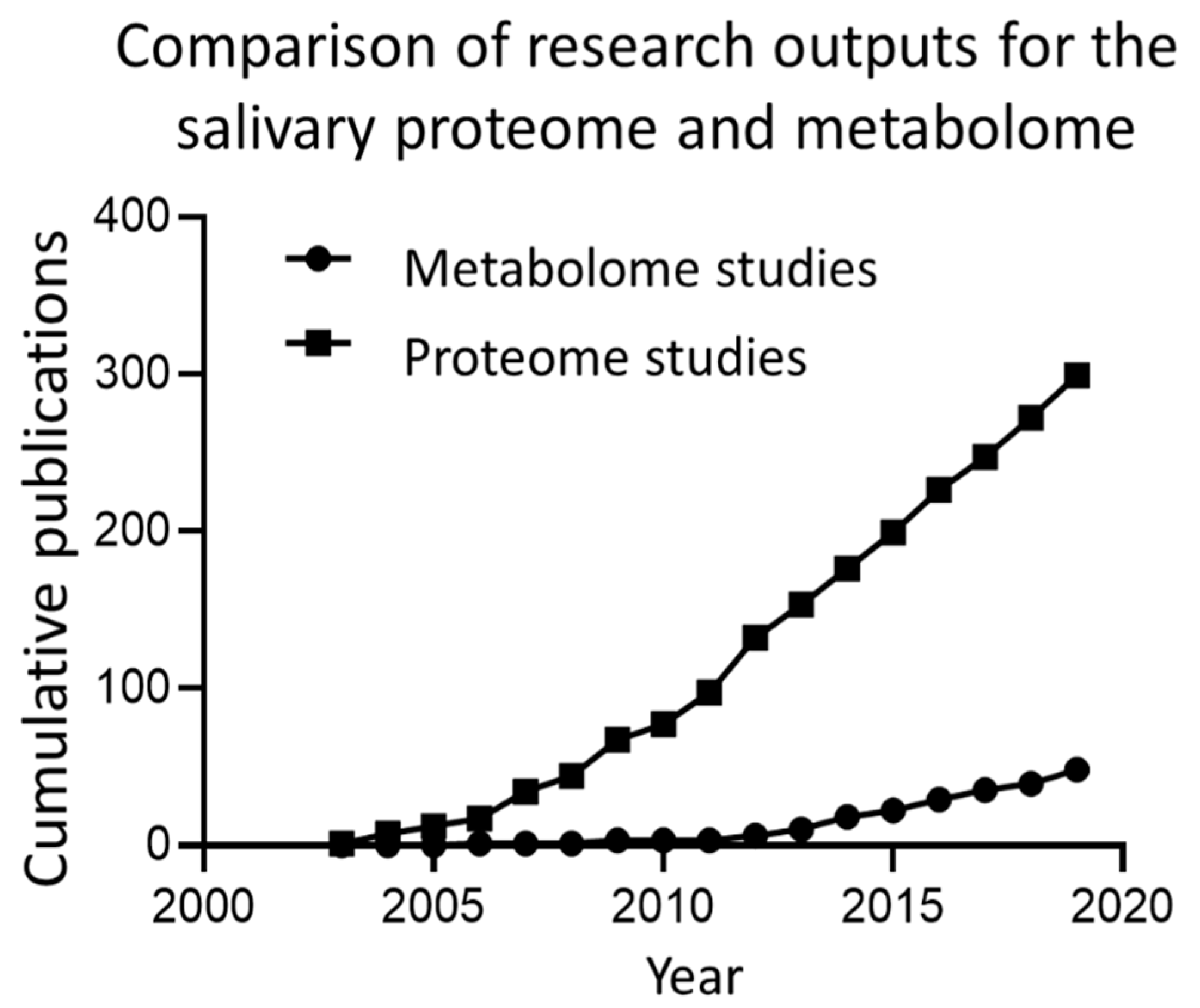
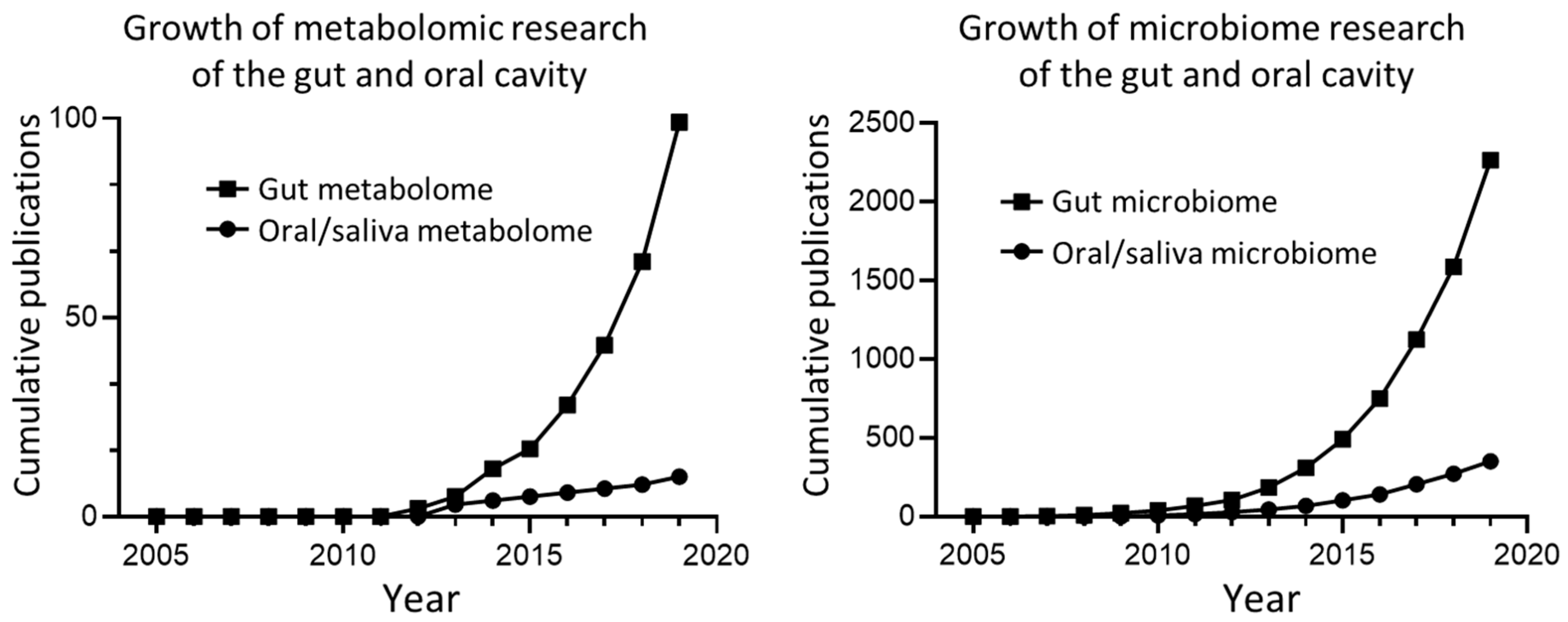
1. Introduction
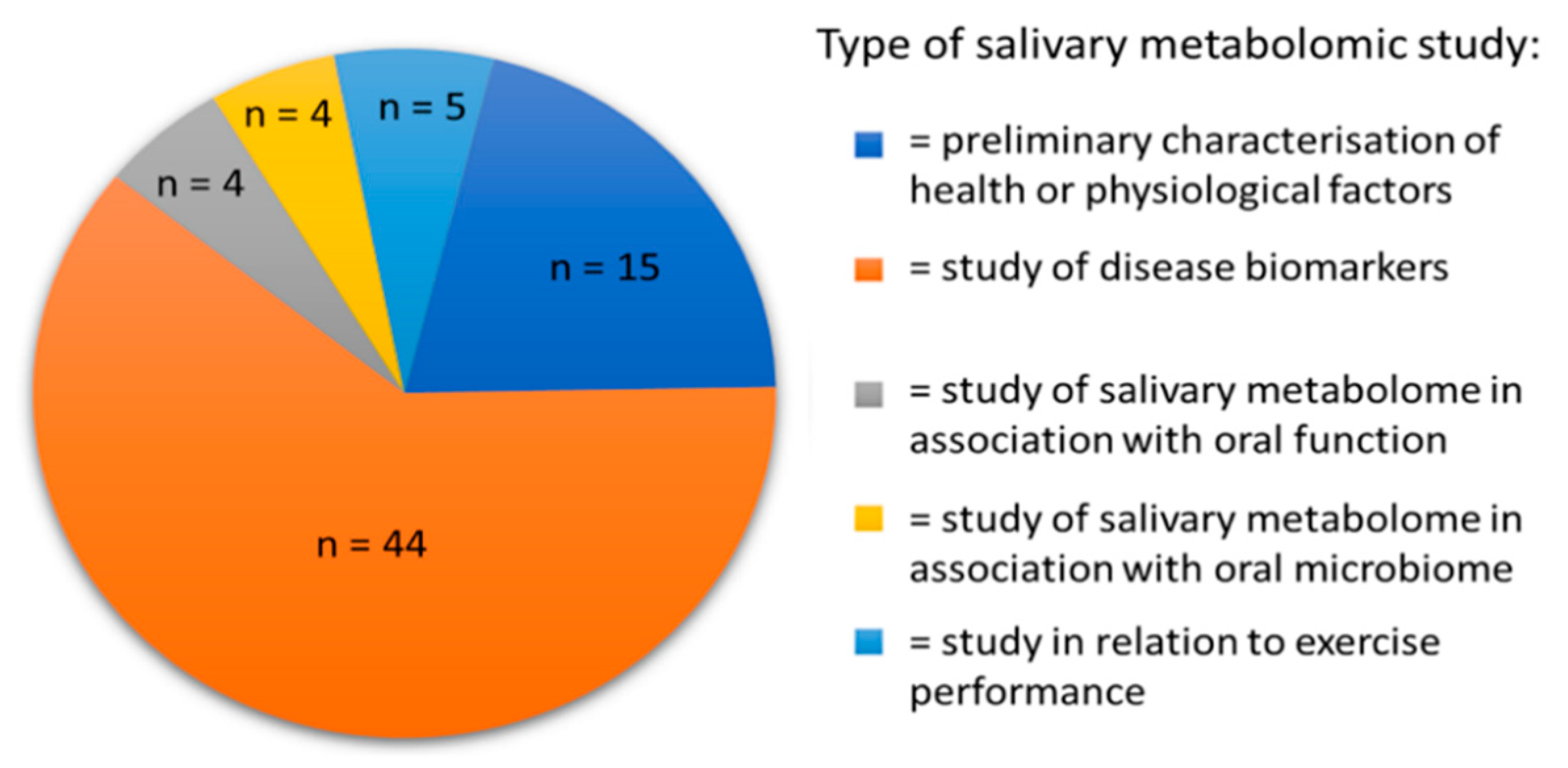
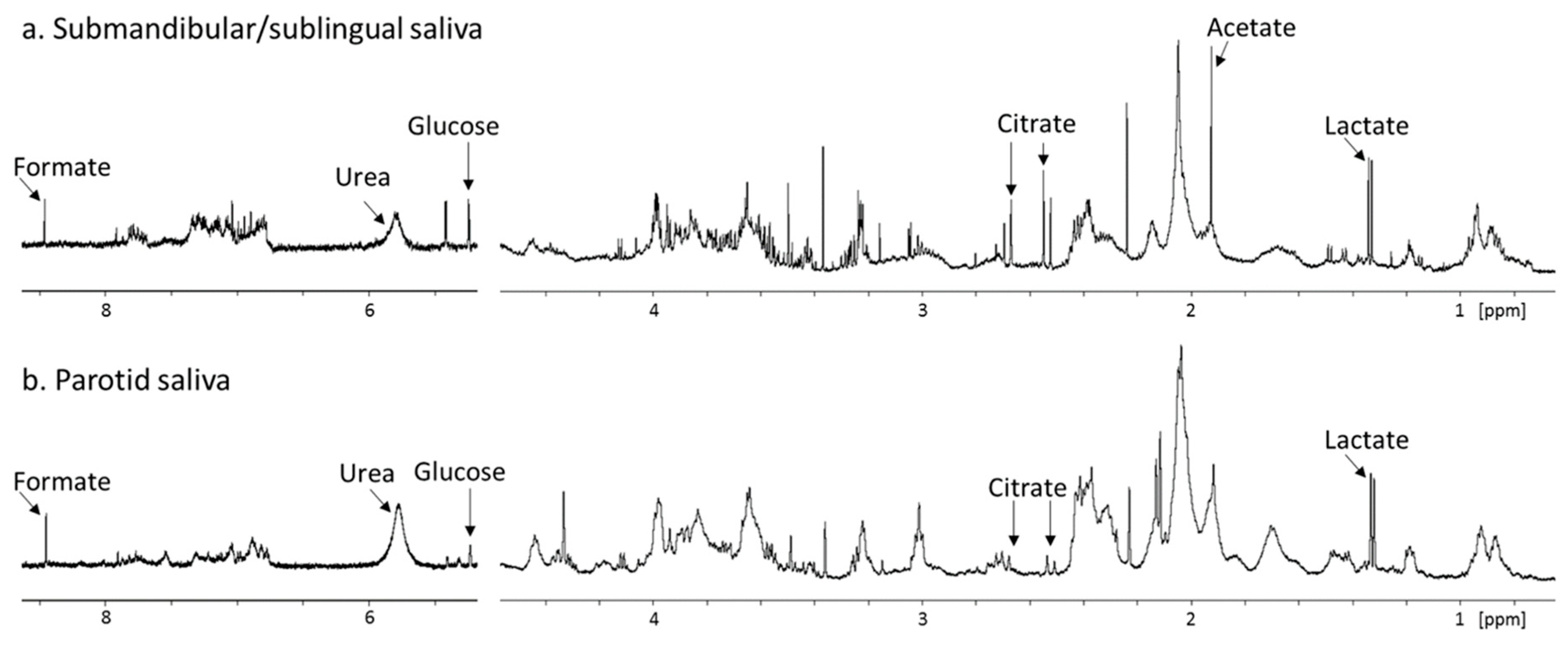
2. Approaches to the Study of Salivary Metabolites
3. Current Knowledge of the Salivary Metabolome
4. Role of Host–Microbiome Interactions in Modifying Salivary Metabolic Composition
5. Potential Physiological Significance of Salivary Metabolites
6. Future Directions for Salivary Metabolomics
7. Conclusions
Funding
Conflicts of Interest
References
- Proctor, G.B. The physiology of salivary secretion. Periodontol. 2016, 70, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Navazesh, M. Methods for collecting saliva. Ann. N. Y. Acad. Sci. 1993, 694, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, Y.C.; Chu, L.J.; Chen, Y.T.; Chi, L.M.; Chien, K.Y.; Chiang, W.F.; Chang, Y.T.; Chen, S.F.; Wang, W.S.; Chuang, Y.N. Variability assessment of 90 salivary proteins in intraday and interday samples from healthy donors by multiple reaction monitoring-mass spectrometry. PROTEOMICS–Clin. Appl. 2018, 12, 1700039. [Google Scholar] [CrossRef] [PubMed]
- Quintana, M.; Palicki, O.; Lucchi, G.; Ducoroy, P.; Chambon, C.; Salles, C.; Morzel, M. Inter-individual variability of protein patterns in saliva of healthy adults. J. Proteomics 2009, 72, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Goswami, Y.; Mishra, R.; Agrawal, A.P.; Agrawal, L.A. Salivary biomarkers: A review of powerful diagnostic tool. IOSR J. Dental Med. Sci. 2015, 14, 80–87. [Google Scholar]
- Al-Tarawneh, S.K.; Border, M.B.; Dibble, C.F.; Bencharit, S. Defining salivary biomarkers using mass spectrometry-based proteomics: A systematic review. OMICS 2011, 15, 353–361. [Google Scholar] [CrossRef]
- Huq, N.L.; Cross, K.J.; Ung, M.; Myroforidis, H.; Veith, P.D.; Chen, D.; Stanton, D.; He, H.; Ward, B.R.; Reynolds, E.C. A review of the salivary proteome and peptidome and saliva-derived peptide therapeutics. Int. J. Pept. Res. Ther. 2007, 13, 547–564. [Google Scholar] [CrossRef]
- Dame, Z.T.; Aziat, F.; Mandal, R.; Krishnamurthy, R.; Bouatra, S.; Borzouie, S.; Guo, A.C.; Sajed, T.; Deng, L.; Lin, H. The human saliva metabolome. Metabolomics 2015, 11, 1864–1883. [Google Scholar] [CrossRef]
- Yoshizawa, J.M.; Schafer, C.A.; Schafer, J.J.; Farrell, J.J.; Paster, B.J.; Wong, D.T. Salivary biomarkers: Toward future clinical and diagnostic utilities. Clin. Microbiol. Rev. 2013, 26, 781–791. [Google Scholar] [CrossRef]
- Pfaffe, T.; Cooper-White, J.; Beyerlein, P.; Kostner, K.; Punyadeera, C. Diagnostic potential of saliva: Current state and future applications. Clin. Chem. 2011, 57, 675–687. [Google Scholar] [CrossRef]
- Bellagambi, F.G.; Lomonaco, T.; Salvo, P.; Vivaldi, F.; Hangouët, M.; Ghimenti, S.; Biagini, D.; Di Francesco, F.; Fuoco, R.; Errachid, A. Saliva sampling: Methods and devices: An overview. TrAC Trends Analytical Chem. 2019, 124, 1–15. [Google Scholar] [CrossRef]
- Gardner, A.; Parkes, H.G.; Carpenter, G.H.; So, P.-W. Developing and standardizing a protocol for quantitative proton nuclear magnetic resonance (1h nmr) spectroscopy of saliva. J. Proteome Res. 2018, 17, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Stafford, M.; Horning, M.G.; Zlatkis, A. Profiles of volatile metabolites in body fluids. J. Chromatogr. A 1976, 126, 495–502. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Buckingham, M.J.; Sadler, P.J. High resolution 1h nmr studies of vertebrate blood and plasma. Biochem. J. 1983, 211, 605. [Google Scholar] [CrossRef] [PubMed]
- Bales, J.R.; Higham, D.P.; Howe, I.; Nicholson, J.K.; Sadler, P.J. Use of high-resolution proton nuclear magnetic resonance spectroscopy for rapid multi-component analysis of urine. Clin. Chem. 1984, 30, 426. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Wilson, I.D. High resolution proton magnetic resonance spectroscopy of biological fluids. Progr. Nuclear Magnet. Resonance Spectrosc. 1989, 21, 449–501. [Google Scholar] [CrossRef]
- Sequeira, S.; So, P.; Everett, J.; Elcombe, C.; Kelvin, A.; Nicholson, J. 1h-nmr spectroscopy of biofluids and the investigation of xenobiotic-induced changes in liver biochemistry. J. Pharm. Biomed. Anal. 1990, 8, 945–949. [Google Scholar] [CrossRef]
- Dan, K.; Kawano, K.; Terada, Y.; Iga, F.; Hirayasu, R. High resolution proton nuclear magnetic resonance studies of human saliva. Jap. J. Oral Biol. 1989, 31, 324–328. [Google Scholar] [CrossRef][Green Version]
- Harada, H.; Shimizu, H.; Maeiwa, M. 1h-nmr of human saliva. An application of nmr spectroscopy in forensic science. Forensic Sci. Int. 1987, 34, 189–195. [Google Scholar] [CrossRef]
- Yamada-Nosaka, A.; Fukutomi, S.; Uemura, S.; Hashida, T.; Fujishita, M.; Kobayashi, Y.; Kyogoku, Y. Preliminary nuclear magnetic resonance studies on human saliva. Arch. Oral Biol. 1991, 36, 697–701. [Google Scholar] [CrossRef]
- Silwood, C.; Lynch, E.; Claxson, A.; Grootveld, M. 1h and 13c nmr spectroscopic analysis of human saliva. J. Dent. Res. 2002, 81, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Sánchez, B.; Priego-Capote, F.; Luque de Castro, M.D. Study of sample preparation for metabolomic profiling of human saliva by liquid chromatography–time of flight/mass spectrometry. J. Chromatogr. A 2012, 1248, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.; Wong, D.T.; Hirayama, A.; Soga, T.; Tomita, M. Capillary electrophoresis mass spectrometry-based saliva metabolomics identified oral, breast and pancreatic cancer-specific profiles. Metabolomics 2010, 6, 78–95. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. ‘Metabonomics’: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological nmr spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Reid, D.G.; Bazin, D.; Daudon, M.; Duer, M.J. Solid state nmr of salivary calculi: Proline-rich salivary proteins, citrate, polysaccharides, lipids, and organic–mineral interactions. Comptes Rendus Chimie 2016, 19, 1665–1671. [Google Scholar] [CrossRef]
- Buko, A.M. Capillary electrophoresis mass spectrometry based metabolomics. J. Appl. Bioanalysis 2017, 3, 1303. [Google Scholar] [CrossRef]
- Ren, J.-L.; Zhang, A.-H.; Kong, L.; Wang, X.-J. Advances in mass spectrometry-based metabolomics for investigation of metabolites. RSC Adv. 2018, 8, 22335–22350. [Google Scholar] [CrossRef]
- Aretz, I.; Meierhofer, D. Advantages and pitfalls of mass spectrometry based metabolome profiling in systems biology. Int. J. Mol. Sci. 2016, 17, 632. [Google Scholar] [CrossRef]
- Emwas, A.-H.M. The strengths and weaknesses of nmr spectroscopy and mass spectrometry with particular focus on metabolomics research. In Metabonomics; Springer: New York, NY, USA, 2015; pp. 161–193. [Google Scholar]
- Takis, P.G.; Ghini, V.; Tenori, L.; Turano, P.; Luchinat, C. Uniqueness of the nmr approach to metabolomics. TrAC Trends Analy. Chem. 2018, 120, 1–9. [Google Scholar] [CrossRef]
- Markley, J.L.; Brüschweiler, R.; Edison, A.S.; Eghbalnia, H.R.; Powers, R.; Raftery, D.; Wishart, D.S. The future of nmr-based metabolomics. Curr. Opin. Biotechnol. 2017, 43, 34–40. [Google Scholar] [CrossRef]
- Marshall, D.D.; Powers, R. Beyond the paradigm: Combining mass spectrometry and nuclear magnetic resonance for metabolomics. Prog. Nuclear Magn. Resonance Spectrosc. 2017, 100, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Beale, D.J.; Jones, O.A.; Karpe, A.V.; Dayalan, S.; Oh, D.Y.; Kouremenos, K.A.; Ahmed, W.; Palombo, E.A. A review of analytical techniques and their application in disease diagnosis in breathomics and salivaomics research. Int. J. Mol. Sci. 2017, 18, 24. [Google Scholar] [CrossRef] [PubMed]
- Lohavanichbutr, P.; Zhang, Y.; Wang, P.; Gu, H.; Gowda, G.N.; Djukovic, D.; Buas, M.F.; Raftery, D.; Chen, C. Salivary metabolite profiling distinguishes patients with oral cavity squamous cell carcinoma from normal controls. PLoS ONE 2018, 13, e0204249. [Google Scholar] [CrossRef] [PubMed]
- Dona, A.C.; Jiménez, B.; Schäfer, H.; Humpfer, E.; Spraul, M.; Lewis, M.R.; Pearce, J.T.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Precision high-throughput proton nmr spectroscopy of human urine, serum, and plasma for large-scale metabolic phenotyping. Anal. Chem. 2014, 86, 9887–9894. [Google Scholar] [CrossRef]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.D.; Bundy, J.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for nmr spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060. [Google Scholar] [CrossRef]
- Chan, E.C.Y.; Pasikanti, K.K.; Nicholson, J.K. Global urinary metabolic profiling procedures using gas chromatography–mass spectrometry. Nat. Protoc. 2011, 6, 1483. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Wang, X. Saliva metabolomics opens door to biomarker discovery, disease diagnosis, and treatment. Appl. Biochem. Biotechnol. 2012, 168, 1718–1727. [Google Scholar] [CrossRef]
- Aimetti, M.; Cacciatore, S.; Graziano, A.; Tenori, L. Metabonomic analysis of saliva reveals generalized chronic periodontitis signature. Metabolomics 2012, 8, 465–474. [Google Scholar] [CrossRef]
- Romano, F.; Meoni, G.; Manavella, V.; Baima, G.; Tenori, L.; Cacciatore, S.; Aimetti, M. Analysis of salivary phenotypes of generalized aggressive and chronic periodontitis through nuclear magnetic resonance-based metabolomics. J. Periodontol. 2018, 89, 1452–1460. [Google Scholar] [CrossRef]
- Wei, J.; Xie, G.; Zhou, Z.; Shi, P.; Qiu, Y.; Zheng, X.; Chen, T.; Su, M.; Zhao, A.; Jia, W. Salivary metabolite signatures of oral cancer and leukoplakia. Int. J. Cancer 2011, 129, 2207–2217. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, A.; Geddes, T.; Han, B.; Bahado-Singh, R.O.; Wilson, G.D.; Imam, K.; Maddens, M.; Graham, S.F. Diagnostic biomarkers of alzheimer’s disease as identified in saliva using 1h nmr-based metabolomics. J. Alzheimers Dis. 2017, 58, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Figueira, J.; Jonsson, P.; Adolfsson, A.N.; Adolfsson, R.; Nyberg, L.; Öhman, A. Nmr analysis of the human saliva metabolome distinguishes dementia patients from matched controls. Mol. Biosyst. 2016, 12, 2562–2571. [Google Scholar] [CrossRef] [PubMed]
- Fidalgo, T.K.; Freitas-Fernandes, L.B.; Angeli, R.; Muniz, A.M.; Gonsalves, E.; Santos, R.; Nadal, J.; Almeida, F.C.; Valente, A.P.; Souza, I.P. Salivary metabolite signatures of children with and without dental caries lesions. Metabolomics 2013, 9, 657–666. [Google Scholar] [CrossRef]
- Pereira, J.; Duarte, D.; Carneiro, T.; Ferreira, S.; Cunha, B.; Soares, D.; Costa, A.; Gil, A. Saliva nmr metabolomics: Analytical issues in paediatric oral health research. Oral Dis. 2019, 25, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Mikkonen, J.J.W.; Singh, S.P.; Akhi, R.; Salo, T.; Lappalainen, R.; González-Arriagada, W.A.; Ajudarte Lopes, M.; Kullaa, A.M.; Myllymaa, S. Potential role of nuclear magnetic resonance spectroscopy to identify salivary metabolite alterations in patients with head and neck cancer. Oncol. Lett. 2018, 16, 6795–6800. [Google Scholar] [CrossRef]
- Wang, Q.; Gao, P.; Wang, X.; Duan, Y. The early diagnosis and monitoring of squamous cell carcinoma via saliva metabolomics. Sci. Rep. 2014, 4, 6802. [Google Scholar] [CrossRef]
- Wang, Q.; Gao, P.; Wang, X.; Duan, Y. Investigation and identification of potential biomarkers in human saliva for the early diagnosis of oral squamous cell carcinoma. Clin. Chim. Acta 2014, 427, 79–85. [Google Scholar] [CrossRef]
- Yan, S.-K.; Wei, B.-J.; Lin, Z.-Y.; Yang, Y.; Zhou, Z.-T.; Zhang, W.-D. A metabonomic approach to the diagnosis of oral squamous cell carcinoma, oral lichen planus and oral leukoplakia. Oral Oncol. 2008, 44, 477–483. [Google Scholar] [CrossRef]
- Ishikawa, S.; Sugimoto, M.; Edamatsu, K.; Sugano, A.; Kitabatake, K.; Iino, M. Discrimination of oral squamous cell carcinoma from oral lichen planus by salivary metabolomics. Oral Dis. 2019. [Google Scholar] [CrossRef]
- Ishikawa, S.; Wong, D.T.; Sugimoto, M.; Gleber-Netto, F.O.; Li, F.; Tu, M.; Zhang, Y.; Akin, D.; Iino, M. Identification of salivary metabolites for oral squamous cell carcinoma and oral epithelial dysplasia screening from persistent suspicious oral mucosal lesions. Clin. Oral Investig. 2019, 23, 3557–3563. [Google Scholar] [CrossRef] [PubMed]
- Babay, N.; Alshehri, F.; Al Rowis, R. Majors highlights of the new 2017 classification of periodontal and peri-implant diseases and conditions. Saudi. Dent. J. 2019, 31, 303–305. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.P.; Saxena, M.; Saimbi, C.S.; Siddiqui, M.H.; Roy, R. Post-periodontal surgery propounds early repair salivary biomarkers by 1 h nmr based metabolomics. Metabolomics 2019, 15, 141. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, S.; Sugimoto, M.; Kitabatake, K.; Sugano, A.; Nakamura, M.; Kaneko, M.; Ota, S.; Hiwatari, K.; Enomoto, A.; Soga, T. Identification of salivary metabolomic biomarkers for oral cancer screening. Sci. Rep. 2016, 6, 31520. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, G.; Ramani, P.; Patankar, S. Serum metabolomics in oral leukoplakia and oral squamous cell carcinoma. J. Cancer Res. Ther. 2017, 13, 556. [Google Scholar] [CrossRef]
- Ohshima, M.; Sugahara, K.; Kasahara, K.; Katakura, A. Metabolomic analysis of the saliva of japanese patients with oral squamous cell carcinoma. Oncol. Rep. 2017, 37, 2727–2734. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Funchain, P.; Retuerto, M.; Jurevic, R.J.; Fowler, N.; Burkey, B.; Eng, C.; Ghannoum, M.A. Metabolomic analysis identifies differentially produced oral metabolites, including the oncometabolite 2-hydroxyglutarate, in patients with head and neck squamous cell carcinoma. BBA Clinical 2017, 7, 8–15. [Google Scholar] [CrossRef]
- Murata, T.; Yanagisawa, T.; Kurihara, T.; Kaneko, M.; Ota, S.; Enomoto, A.; Tomita, M.; Sugimoto, M.; Sunamura, M.; Hayashida, T. Salivary metabolomics with alternative decision tree-based machine learning methods for breast cancer discrimination. Breast Cancer Res. Treat. 2019, 177, 591–601. [Google Scholar] [CrossRef]
- Liebsch, C.; Pitchika, V.; Pink, C.; Samietz, S.; Kastenmüller, G.; Artati, A.; Suhre, K.; Adamski, J.; Nauck, M.; Völzke, H. The saliva metabolome in association to oral health status. J. Dent. Res. 2019, 0022034519842853. [Google Scholar] [CrossRef]
- Huang, Y.; Zhu, M.; Li, Z.; Sa, R.; Chu, Q.; Zhang, Q.; Zhang, H.; Tang, W.; Zhang, M.; Yin, H. Mass spectrometry-based metabolomic profiling identifies alterations in salivary redox status and fatty acid metabolism in response to inflammation and oxidative stress in periodontal disease. Free Radic. Biol. Med. 2014, 70, 223–232. [Google Scholar] [CrossRef]
- Singh, M.P.; Saxena, M.; Saimbi, C.S.; Arif, J.M.; Roy, R. Metabolic profiling by 1h nmr spectroscopy of saliva shows clear distinction between control and diseased case of periodontitis. Metabolomics 2017, 13, 137. [Google Scholar] [CrossRef]
- Gawron, K.; Wojtowicz, W.; Łazarz-Bartyzel, K.; Łamasz, A.; Qasem, B.; Mydel, P.; Chomyszyn-Gajewska, M.; Potempa, J.; Mlynarz, P. Metabolomic status of the oral cavity in chronic periodontitis. In Vivo 2019, 33, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Rzeznik, M.; Triba, M.N.; Levy, P.; Jungo, S.; Botosoa, E.; Duchemann, B.; Le Moyec, L.; Bernaudin, J.-F.; Savarin, P.; Guez, D. Identification of a discriminative metabolomic fingerprint of potential clinical relevance in saliva of patients with periodontitis using 1h nuclear magnetic resonance (nmr) spectroscopy. PLoS ONE 2017, 12, e0182767. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, G.; Saegusa, J.; Irino, Y.; Tanaka, S.; Tsuda, K.; Takahashi, S.; Sendo, S.; Morinobu, A. Metabolomics analysis of saliva from patients with primary sjögren’s syndrome. Clin. Exp. Immunol. 2015, 182, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Mikkonen, J.J.; Herrala, M.; Soininen, P.; Lappalainen, R.; Tjäderhane, L.; Seitsalo, H.; Niemelä, R.; Salo, T.; Kullaa, A.M.; Myllymaa, S. Metabolic profiling of saliva in patients with primary sjögren’s syndrome. Metabolomics 2013, 3, 1. [Google Scholar]
- Tsuruoka, M.; Hara, J.; Hirayama, A.; Sugimoto, M.; Soga, T.; Shankle, W.R.; Tomita, M. Capillary electrophoresis-mass spectrometry-based metabolome analysis of serum and saliva from neurodegenerative dementia patients. Electrophoresis 2013, 34, 2865–2872. [Google Scholar] [CrossRef]
- Liang, Q.; Liu, H.; Zhang, T.; Jiang, Y.; Xing, H.; Zhang, A.-h. Metabolomics-based screening of salivary biomarkers for early diagnosis of alzheimer’s disease. Rsc. Adv. 2015, 5, 96074–96079. [Google Scholar] [CrossRef]
- Huan, T.; Tran, T.; Zheng, J.; Sapkota, S.; MacDonald, S.W.; Camicioli, R.; Dixon, R.A.; Li, L. Metabolomics analyses of saliva detect novel biomarkers of alzheimer’s disease. J. Alzheimers Dis. 2018, 1–16. [Google Scholar] [CrossRef]
- Marksteiner, J.; Oberacher, H.; Humpel, C. Acyl-alkyl-phosphatidlycholines are decreased in saliva of patients with alzheimer’s disease as identified by targeted metabolomics. J. Alzheimers Dis. 2019, 68, 1–7. [Google Scholar] [CrossRef]
- Liang, Q.; Liu, H.; Li, X.; Zhang, A.-H. High-throughput metabolomics analysis discovers salivary biomarkers for predicting mild cognitive impairment and alzheimer’s disease. Rsc. Adv. 2016, 6, 75499–75504. [Google Scholar] [CrossRef]
- Zheng, J.; Dixon, R.A.; Li, L. Development of isotope labeling lc–ms for human salivary metabolomics and application to profiling metabolome changes associated with mild cognitive impairment. Anal. Chem. 2012, 84, 10802–10811. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Goyal, V.; Kumaran, S.S.; Dwivedi, S.; Srivastava, A.; Jagannathan, N. Quantitative metabolomics of saliva using proton nmr spectroscopy in patients with parkinson’s disease and healthy controls. Neurol. Sci. 2020, 41, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Francavilla, R.; Ercolini, D.; Piccolo, M.; Vannini, L.; Siragusa, S.; De Filippis, F.; De Pasquale, I.; Di Cagno, R.; Di Toma, M.; Gozzi, G.; et al. Salivary microbiota and metabolome associated with celiac disease. Appl. Environ. Microbiol. 2014, 80, 3416–3425. [Google Scholar] [CrossRef] [PubMed]
- Duchemann, B.; Triba, M.N.; Guez, D.; Rzeznik, M.; Savarin, P.; Nunes, H.; Valeyre, D.; Bernaudin, J.-F.; Le Moyec, L. Nuclear magnetic resonance spectroscopic analysis of salivary metabolome in sarcoidosis. Sarcoidosis Vasc. diffus. Lung Dis. 2016, 33, 10–16. [Google Scholar]
- Li, Y.; Wang, D.; Zeng, C.; Liu, Y.; Huang, G.; Mei, Z. Salivary metabolomics profile of patients with recurrent aphthous ulcer as revealed by liquid chromatography–tandem mass spectrometry. J. Int. Med. Res. 2018, 46, 1052–1062. [Google Scholar] [CrossRef]
- Munshi, S.U.; Rewari, B.B.; Bhavesh, N.S.; Jameel, S. Nuclear magnetic resonance based profiling of biofluids reveals metabolic dysregulation in hiv-infected persons and those on anti-retroviral therapy. PLoS ONE 2013, 8, e64298. [Google Scholar] [CrossRef]
- Gilany, K.; Mohamadkhani, A.; Chashmniam, S.; Shahnazari, P.; Amini, M.; Arjmand, B.; Malekzadeh, R.; Ghoochani, N.M.; Fatemeh, B. Metabolomics analysis of the saliva in patients with chronic hepatitis b using nuclear magnetic resonance: A pilot study. Iran. J. Basic Med. Sci. 2019, 22, 1044–1049. [Google Scholar]
- Yatsuoka, W.; Ueno, T.; Miyano, K.; Uezono, Y.; Enomoto, A.; Kaneko, M.; Ota, S.; Soga, T.; Sugimoto, M.; Ushijima, T. Metabolomic profiling reveals salivary hypotaurine as a potential early detection marker for medication-related osteonecrosis of the jaw. PLoS ONE 2019, 14, 1–12. [Google Scholar] [CrossRef]
- Grimaldi, M.; Palisi, A.; Rossi, G.; Stillitano, I.; Faiella, F.; Montoro, P.; Rodriquez, M.; Palladino, R.; D’Ursi, A.M.; Romano, R. Saliva of patients affected by salivary gland tumour: An nmr metabolomics analysis. J. Pharm. Biomed. Anal. 2018, 160, 436–442. [Google Scholar] [CrossRef]
- Goodson, J.M.; Hardt, M.; Hartman, M.-L.; Schulte, F.; Tavares, M.; Al-Mutawa, S.; Ariga, J.; Soparkar, P.; Behbehani, J.; Behbehani, K. Identification of salivary and plasma biomarkers for obesity in children by non-targeted metabolomic analysis. BioRxiv 2018, 371815. [Google Scholar]
- Troisi, J.; Belmonte, F.; Bisogno, A.; Pierri, L.; Colucci, A.; Scala, G.; Cavallo, P.; Mandato, C.; Di Nuzzi, A.; Di Michele, L. Metabolomic salivary signature of pediatric obesity related liver disease and metabolic syndrome. Nutrients 2019, 11, 274. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Hu, H.; Huang, R. A pilot study of the metabolomic profiles of saliva from female orthodontic patients with external apical root resorption. Clin. Chim. Acta 2018, 478, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Dawes, C. Circadian rhythms in human salivary flow rate and composition. J. Physiol. 1972, 220, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Dallmann, R.; Viola, A.U.; Tarokh, L.; Cajochen, C.; Brown, S.A. The human circadian metabolome. Proceed. Nati. Acad. Sci. USA 2012, 109, 2625–2629. [Google Scholar] [CrossRef] [PubMed]
- Bertram, H.C.; Eggers, N.; Eller, N. Potential of human saliva for nuclear magnetic resonance-based metabolomics and for health-related biomarker identification. Anal. Chem. 2009, 81, 9188–9193. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, N.; Hoshi, N.; Masahiro, S.; Enomoto, A.; Ota, S.; Kaneko, M.; Soga, T.; Tomita, M.; Kimoto, K. Effects of inter-day and intra-day variation on salivary metabolomic profiles. Clin. Chim. Acta 2019, 489, 41–48. [Google Scholar] [CrossRef]
- Wallner-Liebmann, S.; Tenori, L.; Mazzoleni, A.; Dieber-Rotheneder, M.; Konrad, M.; Hofmann, P.; Luchinat, C.; Turano, P.; Zatloukal, K. Individual human metabolic phenotype analyzed by 1h nmr of saliva samples. J. Proteome Res. 2016, 15, 1787–1793. [Google Scholar] [CrossRef]
- Walsh, M.C.; Brennan, L.; Malthouse, J.P.G.; Roche, H.M.; Gibney, M.J. Effect of acute dietary standardization on the urinary, plasma, and salivary metabolomic profiles of healthy humans. Am. J. Clin. Nutrit. 2006, 84, 531–539. [Google Scholar] [CrossRef]
- Bader, M.; Stolle, T.; Jennerwein, M.; Hauck, J.r.; Sahin, B.; Hofmann, T. Chemosensate-induced modulation of the salivary proteome and metabolome alters the sensory perception of salt taste and odor-active thiols. J. Agric. Food Chem. 2018, 66, 7740–7749. [Google Scholar] [CrossRef]
- Takeda, I.; Stretch, C.; Barnaby, P.; Bhatnager, K.; Rankin, K.; Fu, H.; Weljie, A.; Jha, N.; Slupsky, C. Understanding the human salivary metabolome. NMR Biomed. 2009, 22, 577–584. [Google Scholar] [CrossRef]
- Okuma, N.; Saita, M.; Hoshi, N.; Soga, T.; Tomita, M.; Sugimoto, M.; Kimoto, K. Effect of masticatory stimulation on the quantity and quality of saliva and the salivary metabolomic profile. PLoS ONE 2017, 12, e0183109. [Google Scholar] [CrossRef] [PubMed]
- Neyraud, E.; Tremblay-Franco, M.; Gregoire, S.; Berdeaux, O.; Canlet, C. Relationships between the metabolome and the fatty acid composition of human saliva; effects of stimulation. Metabolomics 2013, 9, 213–222. [Google Scholar] [CrossRef]
- Sugimoto, M.; Saruta, J.; Matsuki, C.; To, M.; Onuma, H.; Kaneko, M.; Soga, T.; Tomita, M.; Tsukinoki, K. Physiological and environmental parameters associated with mass spectrometry-based salivary metabolomic profiles. Metabolomics 2013, 9, 454–463. [Google Scholar] [CrossRef]
- Gardner, A.; Parkes, H.G.; So, P.-W.; Carpenter, G.H. Determining bacterial and host contributions to the human salivary metabolome. J. Oral Microbiol. 2019, 11, 1617014. [Google Scholar] [CrossRef]
- Figueira, J.; Gouveia-Figueira, S.; Öhman, C.; Lif Holgerson, P.; Nording, M.L.; Öhman, A. Metabolite quantification by nmr and lc-ms/ms reveals differences between unstimulated, stimulated, and pure parotid saliva. J. Pharm. Biomed. Anal. 2017, 140, 295–300. [Google Scholar] [CrossRef]
- Ripps, H.; Shen, W. Review: Taurine: A “very essential” amino acid. Mol. Vis. 2012, 18, 2673–2686. [Google Scholar]
- Schuller-Levis, G.B.; Park, E. Taurine: New implications for an old amino acid. FEMS Microbiol. Lett. 2003, 226, 195–202. [Google Scholar] [CrossRef]
- Miller, B.F.; Muntz, J.A.; Bradel, S. Decomposition of carbohydrate substrates by dental plaque material. J. Dent. Res. 1940, 19, 473–478. [Google Scholar] [CrossRef]
- Takahashi, N.; Washio, J.; Mayanagi, G. Metabolomics of supragingival plaque and oral bacteria. J. Dent. Res. 2010, 89, 1383–1388. [Google Scholar] [CrossRef]
- Takahashi, N.; Washio, J. Metabolomic effects of xylitol and fluoride on plaque biofilm in vivo. J. Dent. Res. 2011, 90, 1463–1468. [Google Scholar] [CrossRef]
- Luke, G.; Gough, H.; Beeley, J.; Geddes, D. Human salivary sugar clearance after sugar rinses and intake of foodstuffs. Caries Res. 1999, 33, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Linke, H.A.B.; Moss, S.J.; Arav, L.; Chiu, P.M. Intra-oral lactic acid production during clearance of different foods containing various carbohydrates. Z. Ernahrungswiss. 1997, 36, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N. Oral microbiome metabolism: From “who are they?” to “what are they doing?”. J. Dent. Res. 2015, 94, 1628–1637. [Google Scholar] [CrossRef] [PubMed]
- Vital, M.; Howe, A.C.; Tiedje, J.M. Revealing the bacterial butyrate synthesis pathways by analyzing (meta)genomic data. mBio 2014, 5, e00889-14. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Wishnok, J.S.; Blusztajn, J.K. Formation of methylamines from ingested choline and lecithin. J. Pharmacol. Exp. Ther. 1983, 225, 320. [Google Scholar]
- Ruhl, S. The scientific exploration of saliva in the post-proteomic era: From database back to basic function. Expert Review Proteomics 2012, 9, 85–96. [Google Scholar] [CrossRef]
- Cleaver, L.M.; Moazzez, R.; Carpenter, G.H. Mixed aerobic-anaerobic incubation conditions induce proteolytic activity from in vitro salivary biofilms. J. Oral Microbiol. 2019, 11, 1643206. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Fernández, J.; Redondo-Blanco, S.; Gutierrez-del-Rio, I.; Miguelez, E.M.; Villar, C.J.; Lombo, F. Colon microbiota fermentation of dietary prebiotics towards short-chain fatty acids and their roles as anti-inflammatory and antitumour agents: A review. J. Funct. Foods 2016, 25, 511–522. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- van de Wouw, M.; Schellekens, H.; Dinan, T.G.; Cryan, J.F. Microbiota-gut-brain axis: Modulator of host metabolism and appetite. J. Nutri. 2017, 147, 727–745. [Google Scholar] [CrossRef] [PubMed]
- Byrne, C.S.; Chambers, E.S.; Morrison, D.J.; Frost, G. The role of short chain fatty acids in appetite regulation and energy homeostasis. Int. J. Obes. (Lond.) 2015, 39, 1331–1338. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Veledo, S.; Vendrell, J. Gut microbiota-derived succinate: Friend or foe in human metabolic diseases? Rev. Endocr. Metab. Disord. 2019, 20, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Garrote, G.L.; Abraham, A.G.; Rumbo, M. Is lactate an undervalued functional component of fermented food products? Front. Microbiol. 2015, 6, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 2015, 74, 13–22. [Google Scholar] [CrossRef]
- Alcock, J.; Maley, C.C.; Aktipis, C.A. Is eating behavior manipulated by the gastrointestinal microbiota? Evolutionary pressures and potential mechanisms. BioEssays News Rev. Mol. Cell. and Dev. Biol. 2014, 36, 940–949. [Google Scholar] [CrossRef]
- Mounayar, R.; Morzel, M.; Brignot, H.; Tremblay-Franco, M.; Canlet, C.; Lucchi, G.; Ducoroy, P.; Feron, G.; Neyraud, E. Salivary markers of taste sensitivity to oleic acid: A combined proteomics and metabolomics approach. Metabolomics 2014, 10, 688–696. [Google Scholar] [CrossRef]
- Mounayar, R.; Morzel, M.; Brignot, H.; Tremblay-Franco, M.; Canlet, C.; Lucchi, G.; Ducoroy, P.; Feron, G.; Neyraud, E. Nutri-metabolomics applied to taste perception phenotype: Human subjects with high and low sensitivity to taste of fat differ in salivary response to oleic acid. OMICS 2014, 18, 666–672. [Google Scholar] [CrossRef]
- Rodrigues, L.; Costa, G.; Cordeiro, C.; Pinheiro, C.; Amado, F.; Lamy, E. Salivary proteome and glucose levels are related with sweet taste sensitivity in young adults. Food Nutr. Res. 2017, 61, 1389208. [Google Scholar] [CrossRef]
- Manley, K.J.; Haryono, R.Y.; Keast, R.S. Taste changes and saliva composition in chronic kidney disease. Renal Soci. Austr. J. 2012, 8, 56–60. [Google Scholar]
- Feron, G. Unstimulated saliva: Background noise in taste molecules. J. Text. Stud. 2019, 50, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Spielman, A. Interaction of saliva and taste. J. Dent. Res. 1990, 69, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Billat, L.V. Use of blood lactate measurements for prediction of exercise performance and for control of training. Sports Med. 1996, 22, 157–175. [Google Scholar] [CrossRef] [PubMed]
- Ohkuwa, T.; Itoh, H.; Yamazaki, Y.; Sato, Y. Salivary and blood lactate after supramaximal exercise in sprinters and long-distance runners. Scand. J. Med. Sci. Sports 1995, 5, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; Almeida, A.; Caperuto, E.; Martins Jr, E.; Rosa, L.C. Effects of a 30-km race upon salivary lactate correlation with blood lactate. Comp. Biochem. Physiol. Part B: Biochem. Molecular Biol. 2006, 145, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Ra, S.-G.; Maeda, S.; Higashino, R.; Imai, T.; Miyakawa, S. Metabolomics of salivary fatigue markers in soccer players after consecutive games. Appl. Physiol. Nutrit. Meta. 2014, 39, 1120–1126. [Google Scholar] [CrossRef]
- Cicero, D.; Di Marino, S.; Dinallo, V.; Pieri, M.; Summa, V.; Desideri, A.; Bernardini, A.; Perondi, F.; D’Ottavio, S. A small sided game session affects salivary metabolite levels in young soccer players. Biomed. Spectro. Imag. 2016, 5, 55–70. [Google Scholar] [CrossRef]
- Santone, C.; Dinallo, V.; Paci, M.; D’Ottavio, S.; Barbato, G.; Bernardini, S. Saliva metabolomics by nmr for the evaluation of sport performance. J. Pharm. Biomed. Anal. 2014, 88, 441–446. [Google Scholar] [CrossRef]
- Malkar, A.; Devenport, N.A.; Martin, H.J.; Patel, P.; Turner, M.A.; Watson, P.; Maughan, R.J.; Reid, H.J.; Sharp, B.L.; Thomas, C.P. Metabolic profiling of human saliva before and after induced physiological stress by ultra-high performance liquid chromatography–ion mobility–mass spectrometry. Metabolomics 2013, 9, 1192–1201. [Google Scholar] [CrossRef]
- Pitti, E.; Petrella, G.; Di Marino, S.; Summa, V.; Perrone, M.; D’Ottavio, S.; Bernardini, A.; Cicero, D.O. Salivary metabolome and soccer match: Challenges for understanding exercise induced changes. Metabolites 2019, 9, 141. [Google Scholar] [CrossRef]
- De Ryck, T.; Grootaert, C.; Jaspaert, L.; Kerckhof, F.-M.; Van Gele, M.; De Schrijver, J.; Van den Abbeele, P.; Swift, S.; Bracke, M.; Van de Wiele, T.; et al. Development of an oral mucosa model to study host–microbiome interactions during wound healing. Appl. Microbiol. Biotechnol. 2014, 98, 6831–6846. [Google Scholar] [CrossRef] [PubMed]
- Jeng, J.H.; Chan, C.P.; Ho, Y.S.; Lan, W.H.; Hsieh, C.C.; Chang, M.C. Effects of butyrate and propionate on the adhesion, growth, cell cycle kinetics, and protein synthesis of cultured human gingival fibroblasts. J. Periodontol. 1999, 70, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Niederman, R.; Buyle-Bodin, Y.; Lu, B.-Y.; Robinson, P.; Naleway, C. Short-chain carboxylic acid concentration in human gingival crevicular fluid. J. Dent. Res. 1997, 76, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Gültekin, S.E.; Sengüven, B.; Sofuoğlu, A.; Taner, L.; Koch, M. Effect of the topical use of the antioxidant taurine on the two basement membrane proteins of regenerating oral gingival epithelium. J. Periodontol. 2012, 83, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Sree, S.L.; Sethupathy, S. Evaluation of the efficacy of taurine as an antioxidant in the management of patients with chronic periodontitis. Dent. Res. J. (Isfahan) 2014, 11, 228. [Google Scholar]
- Gardner, A. Endogenous salivary citrate is associated with enhanced rheological properties following oral capsaicin-stimulation. Exp. Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Vijay, A.; Inui, T.; Dodds, M.; Proctor, G.; Carpenter, G. Factors that influence the extensional rheological property of saliva. PLoS ONE 2015, 10, e0135792. [Google Scholar] [CrossRef]
- Meldrum, O.W.; Yakubov, G.E.; Bonilla, M.R.; Deshmukh, O.; McGuckin, M.A.; Gidley, M.J. Mucin gel assembly is controlled by a collective action of non-mucin proteins, disulfide bridges, ca2+-mediated links, and hydrogen bonding. Sci. Rep. 2018, 8, 5802. [Google Scholar] [CrossRef]
- Chen, E.Y.; Yang, N.; Quinton, P.M.; Chin, W.-C. A new role for bicarbonate in mucus formation. American J. Physiol. -Lung Cell. Molecul. Physiol. 2010, 299, L542–L549. [Google Scholar] [CrossRef]
- Duran-Pinedo, A.E.; Frias-Lopez, J. Beyond microbial community composition: Functional activities of the oral microbiome in health and disease. Microb. Infect. 2015, 17, 505–516. [Google Scholar] [CrossRef]
- Nyvad, B.; Crielaard, W.; Mira, A.; Takahashi, N.; Beighton, D. Dental caries from a molecular microbiological perspective. Caries Res. 2013, 47, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Wade, W.G. The oral microbiome in health and disease. Pharmacol. Res. 2013, 69, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Zierer, J.; Jackson, M.A.; Kastenmüller, G.; Mangino, M.; Long, T.; Telenti, A.; Mohney, R.P.; Small, K.S.; Bell, J.T.; Steves, C.J.; et al. The fecal metabolome as a functional readout of the gut microbiome. Nat. Genet. 2018, 50, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Zaura, E.; Brandt, B.W.; Prodan, A.; Teixeira de Mattos, M.J.; Imangaliyev, S.; Kool, J.; Buijs, M.J.; Jagers, F.L.; Hennequin-Hoenderdos, N.L.; Slot, D.E.; et al. On the ecosystemic network of saliva in healthy young adults. ISME J. 2017, 11, 1218–1231. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease (n. Disease vs. n. Control) | Metabolomic Technique | Biomarkers Identified (Raised/Lowered in Disease) | Reference |
|---|---|---|---|
| Oral Cancer (69 v. 87)) | Capillary electrophoresis time-of-flight mass spectrometry (CE-TOF-MS) | Alanine, taurine, leucine/isoleucine, histidine, valine, tryptophan, glutamate, threonine, carnitine, pipecolic acid | [23] |
| (24 v. 44) | CE-TOF-MS | 3-phosphoglycerate, carnosine, phosphoenolpyruvate, dihydroxyacetonephosphate Pipecolate, s-adenosylmethionine | [55] |
| (37 v. 34) | Ultraperformance liquid chromatography coupled with quadrupole-TOFMS (UPLC-QTOFMS) | Lactate, n-Eicosanoic acid Valine, γ-aminobutyric acid (GABA), Phenylalanine | [42] |
| (30 v. 30) | Reverse phase liquid chromatography and hydrophilic interaction chromatography with TOF-MS | Propionylcholine, succinic acid, lactic acid Acetylphenylalanine, carnitine, phytosphingosine | [48] |
| (101 v. 35) | 1H-NMR and LC-MS/MS | Glycine, proline | [34] |
| (22 v. 18) | UPLC-QTOFMS | 1-methylhistidine, inositol 1,3,4-triphosphate, dglycerate-2-phosphate, 4-nitroquinoline-1-oxide, 2-oxoarginine, norcocaine nitroxide, sphinganine-1-phosphate, pseudouridine L-homocysteic acid, ubiquinone, neuraminic acid, estradiol valerate | [56] |
| (22 v. 18) | CE-MS | Choline, hydroxyphenylacetic acid, 2-hydroxy-4-methylvaleric acid, valine, 3-phenyllactic acid, leucine, hexanoic acid, octanoic acid, terephthalic acid, γ-butyrobetaine, 3-(4-hydroxyphenyl)propionic acid, isoleucine, tryptophan, 3-phenylpropionic acid, butyric acid, cadaverine, 2-oxoisovaleric acid, trimethyllysine, taurine, glycolic acid, 3-hydroxybutyric acid, heptanoic acid, alanine Urea | [57] |
| (7 v. 7) | LC-MS/MS, LC-MS, GC-MS | Guanosine-3-monophosphate, N-acetylputrescine, 2-Hydroxyglutarate, Adenosine-3-monophosphate, Uracil, Spermidine, 3-Glycosyl tryptopha | [58] |
| Oral Leukoplakia (32 v. 34) | UPLC-QTOFMS | Isoleucine, phenylalanine, threonine, homocysteine, n-tetradecanoic acid, 4-methoxyphenylacetic acid | [42] |
| Breast Cancer (30 v. 87) | CE-TOF-MS | Alanine, leucine/isoleucine, valine, glutamate Taurine, Threonine | [23] |
| (124 v. 42) | CE- and LC-MS | Polyamines, spermine | [59] |
| Prostate Cancer (18 v. 87) | CE-TOF-MS | Alanine, leucine/isoleucine, histidine, valine, tryptophan, glutamic acid, threonine, carnitine Taurine | [23] |
| Periodontal Disease (11 v. 87) | CE-TOF-MS | Alanine, leucine/isoleucine, valine, tryptophan, threonine Pipecolic acid, taurine | [23] |
| (21 v. 22) | Proton nuclear magnetic resonance spectroscopy (1H-NMR) | Acetate, GABA, propionate, n-butyrate, succinate, trimethylamine, valine Pyruvate, n-acetyl groups | [40] |
| (n = 909, significant correlations between [metabolite] and disease severity) | Ultra-high performance liquid chromatography and tandem MS (UHPLC-MS/MS) | Phenylpropionate, phenylacetate | [60] |
| (50 v. 50) | GC-MS and LC-MS | Hydroxyeicosatetraenoic acid, prostaglandins, isoprostanes, thromboxane Hydroxyoctadecadienoic acid, prostacylcin | [61] |
| (62 v. 52) | 1H-NMR | Alanine, valine, leucine, glutamate, succinate, lactate Pyruvate | [62] |
| (30 v. 15) | 1H-NMR | Methanol, acetone Taurine, lactate | [63] |
| (26 v. 25) | 1H-NMR | Butyrate GABA, lactate, methanol, threonine | [64] |
| Periodontal Disease (Aggressive) (28 v. 39) | 1H-NMR | Isoleucine, proline, valine, tyrosine, phenylalanine Lactate, pyruvate, n-acetyl groups | [41] |
| Periodontal Disease (Chronic) (33 v. 39) | 1H-NMR | Formate, phenylalanine, tyrosine Sarcosine, Lactate, pyruvate, n-acetyl groups | [41] |
| Dental Caries (15 v. 18) | 1H-NMR | Propionate, butyrate Saccharides | [45] |
| (15 v. 23) | 1H-NMR | Extensive list of amino acids and organic acids Butyrate, acetone | [46] |
| Primary Sjögren’s Syndrome (12 v. 21) | Gas chromatography-MS (GC-MS) | 39 metabolites including organic acids, amino acids and carbohydrates | [65] |
| (15 v. 15) | 1H-NMR | Alanine, glycine, butyrate, taurine, phenylalanine, choline, tyrosine | [66] |
| Dementia (10 v. 9) | CE-TOF-MS | Tyrosine, arginine | [67] |
| (17 v. 34) | 1H-NMR | Acetate, histamine, propionate Dimethyl sulfone, glycerol, succinate, taurine | [44] |
| Alzheimer’s Disease (256 v. 218) | Faster ultra-performance liquid chromatography-MS (FUPLC-MS) | Sphinganine-1-phosphate, ornithine, phenyllactic acid Inosine, 3-dehydrocarnitine, hypoxanthine | [68] |
| (32 v. 45) | LC-MS | Histidinyl-Phenylalanine, methylguanosine, choline-cytidine | [69] |
| (9 v. 12) | 1H-NMR | Acetone, propionate | [43] |
| (25 v. 25) | Flow injection analysis- tandem mass spec. (DFI-MS/MS) | acyl-alkyl phosphatidylcholines | [70] |
| Alzheimer’s Disease (relative to mild cognitive impairment; (660 v. 586) | (FUPLC-MS) | Sphinganine-1-phosphate, cytidine, pyroglutamic acid, L-glutamic acid, L-tryptophan, ornithine, phenyllactic acid 3-dehydrocarnitine, hypoxanthine, Inosine | [71] |
| Mild cognitive impairment relative to health (20 v. 20) | LC-MS | Taurine, several putative di- and tripeptides | [72] |
| Parkinson’s disease (76 v. 37) | 1H-NMR | Histidine, tyrosine, phenylalanine, acetate, n-acetyl groups, propionate, Isoleucine, lysine, glycine, valine, GABA, fucose, trimethylamine, trimethylamine-oxide, alanine, acetoin | [73] |
| Celiac disease (13 v. 13) | GC-MS | Carbon disulfide, 1-chlorodecane, trichloromethane Alcohols, phenols, ketones | [74] |
| Sarcoidosis (24 v. 45) | 1H-NMR | Lactate, acetate Methanol, n-acetyl groups | [75] |
| Recurrent aphthous ulceration (45v. 49) | LC-MS/MS | Tryptophan, 5-methoxytryptamine, 5-methoxytryptophan Estrone sulfate, 17β-Estradiol 3-sulfate, Dehydroepiandrosterone sulfate | [76] |
| Untreated HIV (21 v. 8) | 1H-NMR | Butyrate, Lactate | [77] |
| Treated HIV (Antiretroviral therapy) (12 v. 8) | 1H-NMR | Butyrate, Lactate, propionate | [77] |
| Hepatitis B (20 v. 16) | 1H-NMR | Propionate, acetate, succinate, putrescine, tyrosine Lactate, pyruvate, butyrate, 4-hydroxybenzoate, 4-pyridoxoate | [78] |
| Medication- related osteonecrosis of the jaw (17 v. 18) | CE-MS | Hypotaurine | [79] |
| Parotid gland tumour (36 v. 23) | 1H-NMR | Alanine, threonine, leucine, serine, valine Formate, succinate | [80] |
| Obesity (22 v. 46) | HPLC-MS/MS and GC-MS | Creatinine, phosphate, cadaverine, dipeptides, putrescine | [81] |
| Paediatric obesity (18 v. 23) | GC-MS | Palmitic acid, myristic acid, urea, N-acetyl galactosamine | [82] |
| External apical root resorption in orthodontic therapy (8 v. 11) | 1H-NMR | Butyrate, urea, glucose, formate, fumarate | [83] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gardner, A.; Carpenter, G.; So, P.-W. Salivary Metabolomics: From Diagnostic Biomarker Discovery to Investigating Biological Function. Metabolites 2020, 10, 47. https://doi.org/10.3390/metabo10020047
Gardner A, Carpenter G, So P-W. Salivary Metabolomics: From Diagnostic Biomarker Discovery to Investigating Biological Function. Metabolites. 2020; 10(2):47. https://doi.org/10.3390/metabo10020047
Chicago/Turabian StyleGardner, Alexander, Guy Carpenter, and Po-Wah So. 2020. "Salivary Metabolomics: From Diagnostic Biomarker Discovery to Investigating Biological Function" Metabolites 10, no. 2: 47. https://doi.org/10.3390/metabo10020047
APA StyleGardner, A., Carpenter, G., & So, P.-W. (2020). Salivary Metabolomics: From Diagnostic Biomarker Discovery to Investigating Biological Function. Metabolites, 10(2), 47. https://doi.org/10.3390/metabo10020047