Volatile Organic Compounds (VOCs) of Endophytic Fungi Growing on Extracts of the Host, Horseradish (Armoracia rusticana)

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Identified Fungi
2.2. Identified VOCs
2.3. Performance of the GC-MS Method and the Proposed Cultivation Technique
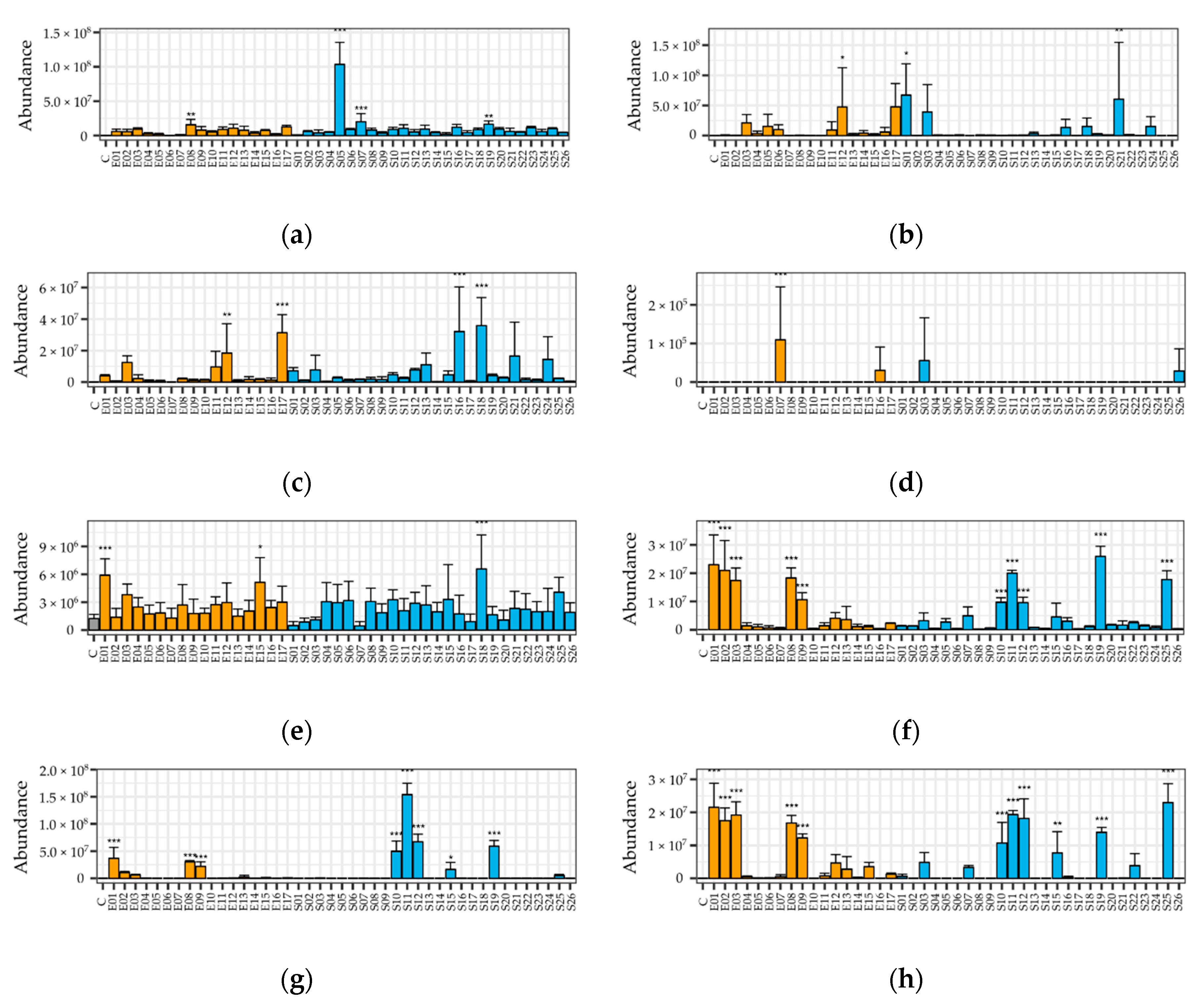
2.4. Differences among VOC Patterns of Fungi
3. Discussion
3.1. Fungi
3.2. Performance of Proposed Cultivation Technique
3.3. Glucosinolate Decomposition by Fungi
3.4. Lack of VOC Pattern Significant Differences between Host Endophytes and Soil Fungi
4. Materials and Methods
4.1. Chemicals
4.2. Standard Microbiological Media
4.3. Horseradish Extract Medium
4.4. Isolation of Fungal Strains
4.5. Fungal Inoculum Preparation and Headspace GC-MS Measurements
4.6. Headspace Measurement Parameters
4.7. Data Evaluation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, Z.; Li, G. A review of advances and new developments in the analysis of biological volatile organic compounds. Microchem. J. 2010, 95, 127–139. [Google Scholar] [CrossRef]
- Girotti, J.R.; Malbrán, I.; Lori, G.A.; Juárez, M.P. Use of solid phase microextraction coupled to capillary gas chromatography-mass spectrometry for screening Fusarium spp. based on their volatile sesquiterpenes. World Mycotoxin J. 2010, 3, 121–128. [Google Scholar] [CrossRef]
- Hung, R.; Lee, S.; Bennett, J.W. Fungal volatile organic compounds and their role in ecosystems. Appl. Microbiol. Biotechnol. 2015, 99, 3395–3405. [Google Scholar] [CrossRef] [PubMed]
- Chitarra, G.S.; Abee, T.; Rombouts, F.M.; Dijksterhuis, J. 1-Octen-3-ol inhibits conidia germination of Penicillium paneum despite of mild effects on membrane permeability, respiration, intracellular pH, and changes the protein composition. FEMS Microbiol. Ecol. 2005, 54, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Garcia, E.; Garzia, A.; Cordobés, S.; Espeso, E.A.; Ugalde, U. 8-Carbon oxylipins inhibit germination and growth, and stimulate aerial conidiation in Aspergillus nidulans. Fungal Biol. 2011, 115, 393–400. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Kalkhove, S.I.C.; Lugones, L.G.; Baars, J.J.P.; Wösten, H.A.B.; Bakker, P.A.H.M. Effects of the mushroom-volatile 1-octen-3-ol on dry bubble disease. Appl. Microbiol. Biotechnol. 2013, 97, 5535–5543. [Google Scholar] [CrossRef] [PubMed]
- Mercier, J.; Jiménez, J.I. Control of fungal decay of apples and peaches by the biofumigant fungus Muscodor albus. Postharvest Biol. Technol. 2004, 31, 1–8. [Google Scholar] [CrossRef]
- Macías-Rubalcava, M.L.; Hernández-Bautista, B.E.; Oropeza, F.; Duarte, G.; González, M.C.; Glenn, A.E.; Hanlin, R.T.; Anaya, A.L. Allelochemical Effects of Volatile Compounds and Organic Extracts from Muscodor yucatanensis, a Tropical Endophytic Fungus from Bursera simaruba. J. Chem. Ecol. 2010, 36, 1122–1131. [Google Scholar] [CrossRef]
- Hung, R.; Lee, S.; Bennett, J.W. Arabidopsis thaliana as a model system for testing the effect of Trichoderma volatile organic compounds. Fungal Ecol. 2013, 6, 19–26. [Google Scholar] [CrossRef]
- Paul, D.; Park, K.S. Identification of Volatiles Produced by Cladosporium cladosporioides CL-1, a Fungal Biocontrol Agent That Promotes Plant Growth. Sensors 2013, 13, 13969–13977. [Google Scholar] [CrossRef]
- Kissen, R.; Rossiter, J.T.; Bones, A.M. The ‘mustard oil bomb’: Not so easy to assemble?! Localization, expression and distribution of the components of the myrosinase enzyme system. Phytochem. Rev. 2009, 8, 69–86. [Google Scholar] [CrossRef]
- Manici, L.M.; Lazzeri, L.; Palmieri, S. In Vitro Fungitoxic Activity of Some Glucosinolates and Their Enzyme-Derived Products toward Plant Pathogenic Fungi. J. Agric. Food Chem. 1997, 45, 2768–2773. [Google Scholar] [CrossRef]
- Zsolnai, T. The antimicrobial activity of thiocyanates and isothiocyantes 1. Arzneim. Forsch. Drug Res. 1966, 16, 870–876. [Google Scholar]
- Buxdorf, K.; Yaffe, H.; Barda, O.; Levy, M. The effects of glucosinolates and their breakdown products on necrotrophic fungi. PLoS ONE 2013, 8, e70771. [Google Scholar] [CrossRef]
- Bednarek, P.; Piślewska-Bednarek, M.; Svatoš, A.; Schneider, B.; Doubský, J.; Mansurova, M.; Humphry, M.; Consonni, C.; Panstruga, R.; Sanchez-Vallet, A.; et al. A glucosinolate metabolism pathway in living plant cells mediates broad-spectrum antifungal defense. Science 2009, 323, 101–106. [Google Scholar] [CrossRef]
- Hiruma, K.; Gerlach, N.; Sacristán, S.; Nakano, R.T.; Hacquard, S.; Kracher, B.; Neumann, U.; Ramírez, D.; Bucher, M.; O’Connell, R.J.; et al. Root Endophyte Colletotrichum tofieldiae Confers Plant Fitness Benefits that Are Phosphate Status Dependent. Cell 2016, 165, 464–474. [Google Scholar] [CrossRef]
- Frerigmann, H.; Piślewska-Bednarek, M.; Sánchez-Vallet, A.; Molina, A.; Glawischnig, E.; Gigolashvili, T.; Bednarek, P. Regulation of Pathogen-Triggered Tryptophan Metabolism in Arabidopsis thaliana by MYB Transcription Factors and Indole Glucosinolate Conversion Products. Mol. Plant 2016, 9, 682–695. [Google Scholar] [CrossRef]
- Szűcs, Z.; Plaszkó, T.; Cziáky, Z.; Kiss-Szikszai, A.; Emri, T.; Bertóti, R.; Sinka, L.T.; Vasas, G.; Gonda, S. Endophytic fungi from the roots of horseradish (Armoracia rusticana) and their interactions with the defensive metabolites of the glucosinolate—myrosinase—isothiocyanate system. BMC Plant Biol. 2018, 18, 85. [Google Scholar] [CrossRef] [PubMed]
- Bertóti, R.; Vasas, G.; Gonda, S.; Nguyen, N.M.; Szőke, É.; Jakab, Á.; Pócsi, I.; Emri, T. Glutathione protects Candida albicans against horseradish volatile oil. J. Basic Microbiol. 2016, 56, 1071–1079. [Google Scholar] [CrossRef]
- Burow, M.; Wittstock, U. Regulation and function of specifier proteins in plants. Phytochem. Rev. 2009, 8, 87–99. [Google Scholar] [CrossRef]
- Sellam, A.; Dongo, A.; Guillemette, T.; Hudhomme, P.; Simoneau, P. Transcriptional responses to exposure to the brassicaceous defence metabolites camalexin and allyl-isothiocyanate in the necrotrophic fungus Alternaria brassicicola. Mol. Plant Pathol. 2007, 8, 195–208. [Google Scholar] [CrossRef]
- Butrindr, B.; Niamsup, H.; Shank, L.; Rakariyatham, N. Myrosinase overproducing mutants of Aspergillus sp. NR463. Ann. Microbiol. 2004, 54, 493–501. [Google Scholar]
- Rakariyatham, N.; Sakorn, P. Biodegradation of glucosinolates in brown mustard seed meal (Brassica juncea) by Aspergillus sp. NR-4201 in liquid and solid-state cultures. Biodegradation 2002, 13, 395–399. [Google Scholar] [CrossRef]
- Smits, J.P.; Knol, W.; Bol, J. Glucosinolate degradation by Aspergillus clavatus and Fusarium oxysporum in liquid and solid-state fermentation. Appl. Microbiol. Biotechnol. 1993, 38, 696–701. [Google Scholar] [CrossRef]
- Albaser, A.; Kazana, E.; Bennett, M.H.; Cebeci, F.; Luang-In, V.; Spanu, P.D.; Rossiter, J.T. Discovery of a Bacterial Glycoside Hydrolase Family 3 (GH3) β-Glucosidase with Myrosinase Activity from a Citrobacter Strain Isolated from Soil. J. Agric. Food Chem. 2016, 64, 1520–1527. [Google Scholar] [CrossRef]
- Wieczorek, M.N.; Jeleń, H.H. Volatile Compounds of Selected Raw and Cooked Brassica Vegetables. Molecules 2019, 24, 391. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, H.; Umashankar, S.; Liang, X.; Lee, H.W.; Swarup, S.; Ong, C.N. Characterization of Plant Volatiles Reveals Distinct Metabolic Profiles and Pathways among 12 Brassicaceae Vegetables. Metabolites 2018, 8, 94. [Google Scholar] [CrossRef]
- Müller-Maatsch, J.; Gurtner, K.; Carle, R.; Björn Steingass, C. Investigation into the removal of glucosinolates and volatiles from anthocyanin-rich extracts of red cabbage. Food Chem. 2019, 278, 406–414. [Google Scholar] [CrossRef]
- Wei, S.; Xiao, X.; Wei, L.; Li, L.; Li, G.; Liu, F.; Xie, J.; Yu, J.; Zhong, Y. Development and comprehensive HS-SPME/GC–MS analysis optimization, comparison, and evaluation of different cabbage cultivars (Brassica oleracea L. var. capitata L.) volatile components. Food Chem. 2021, 340, 128166. [Google Scholar] [CrossRef]
- Gonda, S.; Kiss-Szikszai, A.; Szűcs, Z.; Nguyen, N.M.; Vasas, G. Myrosinase Compatible Simultaneous Determination of Glucosinolates and Allyl Isothiocyanate by Capillary Electrophoresis Micellar Electrokinetic Chromatography (CE-MEKC). Phytochem. Anal. 2016, 27, 191–198. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information (NCBI). Basic Local Alignment Search Tool; National Library of Medicine (US); National Center for Biotechnology Information: Bethesda, MD, USA, 2020. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 19 September 2020).
- Strobel, G. Muscodor species—Endophytes with biological promise. Phytochem. Rev. 2011, 10, 165–172. [Google Scholar] [CrossRef]
- Uslu, S.; Celik, M.B. Combustion and emission characteristics of isoamyl alcohol-gasoline blends in spark ignition engine. Fuel 2020, 262, 116496. [Google Scholar] [CrossRef]
- Hari Krishna, S.; Divakar, S.; Prapulla, S.G.; Karanth, N.G. Enzymatic synthesis of isoamyl acetate using immobilized lipase from Rhizomucor miehei. J. Biotechnol. 2001, 87, 193–201. [Google Scholar] [CrossRef]
- Junker, C.; Draeger, S.; Schulz, B. A fine line—Endophytes or pathogens in Arabidopsis thaliana. Fungal Ecol. 2012, 5, 657–662. [Google Scholar] [CrossRef]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun. 2019, 10, 2369. [Google Scholar] [CrossRef] [PubMed]
- Schoen, H.R.; Knighton, W.B.; Peyton, B.M. Endophytic fungal production rates of volatile organic compounds are highest under microaerophilic conditions. Microbiology (Reading, Engl.) 2017, 163, 1767–1777. [Google Scholar] [CrossRef]
- Sridharan, A.P.; Thankappan, S.; Karthikeyan, G.; Uthandi, S. Comprehensive profiling of the VOCs of Trichoderma longibrachiatum EF5 while interacting with Sclerotium rolfsii and Macrophomina phaseolina. Microbiol. Res. 2020, 236, 126436. [Google Scholar] [CrossRef]
- Casarrubia, S.; Sapienza, S.; Fritz, H.; Daghino, S.; Rosenkranz, M.; Schnitzler, J.-P.; Martin, F.; Perotto, S.; Martino, E. Ecologically Different Fungi Affect Arabidopsis Development: Contribution of Soluble and Volatile Compounds. PLoS ONE 2016, 11, e0168236. [Google Scholar] [CrossRef]
- Singh, S.K.; Strobel, G.A.; Knighton, B.; Geary, B.; Sears, J.; Ezra, D. An Endophytic Phomopsis sp. Possessing Bioactivity and Fuel Potential with its Volatile Organic Compounds. Microb. Ecol. 2011, 61, 729–739. [Google Scholar] [CrossRef]
- Rojas-Flores, C.; Ventura-Aguilar, R.I.; Bautista-Baños, S.; Revah, S.; Saucedo-Lucero, J.O. Estimating CO2 and VOCs production of Colletotrichum fragariae and Rhizopus stolonifer grown in cold stored strawberry fruit. Microbiol. Res. 2019, 228, 126327. [Google Scholar] [CrossRef]
- Ndagijimana, M.; Chaves-López, C.; Corsetti, A.; Tofalo, R.; Sergi, M.; Paparella, A.; Guerzoni, M.E.; Suzzi, G. Growth and metabolites production by Penicillium brevicompactum in yoghurt. Int. J. Food Microbiol. 2008, 127, 276–283. [Google Scholar] [CrossRef]
- Heddergott, C.; Calvo, A.M.; Latgé, J.P. The Volatome of Aspergillus fumigatus. Eukaryot. Cell 2014, 13, 1014–1025. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.S.; Mallik, A.U.; Setliff, E. Growth stimulation of ectomycorrhizal fungi by root exudates of Brassicaceae plants: Role of degraded compounds of indole glucosinolates. J. Chem. Ecol. 2003, 29, 1337–1355. [Google Scholar] [CrossRef]
- Schreiner, M.; Krumbein, A.; Knorr, D.; Smetanska, I. Enhanced Glucosinolates in Root Exudates of Brassica rapa ssp. rapa Mediated by Salicylic Acid and Methyl Jasmonate. J. Agric. Food Chem. 2011, 59, 1400–1405. [Google Scholar] [CrossRef]
- Bressan, M.; Roncato, M.-A.; Bellvert, F.; Comte, G.; Haichar, F.E.Z.; Achouak, W.; Berge, O. Exogenous glucosinolate produced by Arabidopsis thaliana has an impact on microbes in the rhizosphere and plant roots. ISME J. 2009, 3, 1243–1257. [Google Scholar] [CrossRef]
- Reese, E.T.; Clapp, R.C.; Mandels, M. A thioglucosidase in fungi. Arch. Biochem. Biophys. 1958, 75, 228–242. [Google Scholar] [CrossRef]
- Ohtsuru, M.; Hata, T. General characteristics of the intracellular myrosinase from Aspergillus niger. Agric. Biol. Chem. 1973, 37, 2543–2548. [Google Scholar] [CrossRef]
- Croat, J.R.; Karki, B.; Berhow, M.; Iten, L.; Muthukumarappan, K.; Gibbons, W.R. Utilizing pretreatment and fungal incubation to enhance the nutritional value of canola meal. J. Appl. Microbiol. 2017, 123, 362–371. [Google Scholar] [CrossRef]
- Pecháček, R.; Velíšek, J.; Hrabcová, H. Decomposition products of allyl isothiocyanate in aqueous solutions. J. Agric. Food Chem. 1997, 45, 4584–4588. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Oresic, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: https://www.r-project.org (accessed on 22 June 2020).
- Wenig, P.; Odermatt, J. OpenChrom: A cross-platform open source software for the mass spectrometric analysis of chromatographic data. BMC Bioinform. 2010, 11, 405. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Isolate | Species | Sequenced Gene(s) | Isolate | Species | Sequenced Gene |
|---|---|---|---|---|---|
| E1 | Fusarium oxysporum species complex | ITS, eF1 | S1 | Notophoma sp. | ITS |
| E2 | Macrophomina phaseolina | ITS, β-tubulin | S2 | Penicillium sp. | ITS |
| E3 | Fusarium oxysporum species complex | ITS, eF1 | S3 | Curvularia sp. | ITS |
| E4 | Setophoma terrestris | ITS, α-actin | S4 | Curvularia sp. | ITS |
| E5 | Paraphoma radicina | ITS, α-actin | S5 | Curvularia sp. | ITS |
| E6 | Paraphoma radicina | ITS, α-actin | S6 | Aspergillus sp. | α-actin |
| E7 | Oidiodendron cerealis | ITS | S7 | Fusarium sp. | ITS |
| E8 | Fusarium sp. | ITS | S8 | Fusarium sp. | ITS |
| E9 | Fusarium sp. | ITS | S9 | Penicillium sp. | ITS |
| E10 | Fusarium sp. | ITS | S10 | Fusarium sp. | ITS |
| E11 | Plectosphaerella sp. | ITS | S11 | Fusarium sp. | ITS |
| E12 | Plectosphaerella sp. | ITS | S12 | Fusarium sp. | ITS |
| E13 | Pyrenochaeta sp. | ITS | S13 | Penicillium sp. | ITS |
| E14 | Volutella sp. | ITS | S14 | Penicillium sp. | ITS |
| E15 | Phomopsis sp. | ITS | S15 | Fusarium sp. | ITS |
| E16 | Cadophora sp. | ITS | S16 | Fusarium sp. | ITS |
| E17 | Colletotrichum sp. | ITS | S17 | Penicillium sp. | ITS |
| S18 | Penicillium sp. | ITS | |||
| S19 | Fusarium sp. | ITS | |||
| S20 | Penicillium sp. | ITS | |||
| S21 | Paraphoma sp. | ITS | |||
| S22 | Penicillium sp. | ITS | |||
| S23 | Penicillium sp. | ITS | |||
| S24 | Aspergillus sp. | calmodulin | |||
| S25 | Fusarium sp. | ITS | |||
| S26 | Penicillium sp. | ITS |
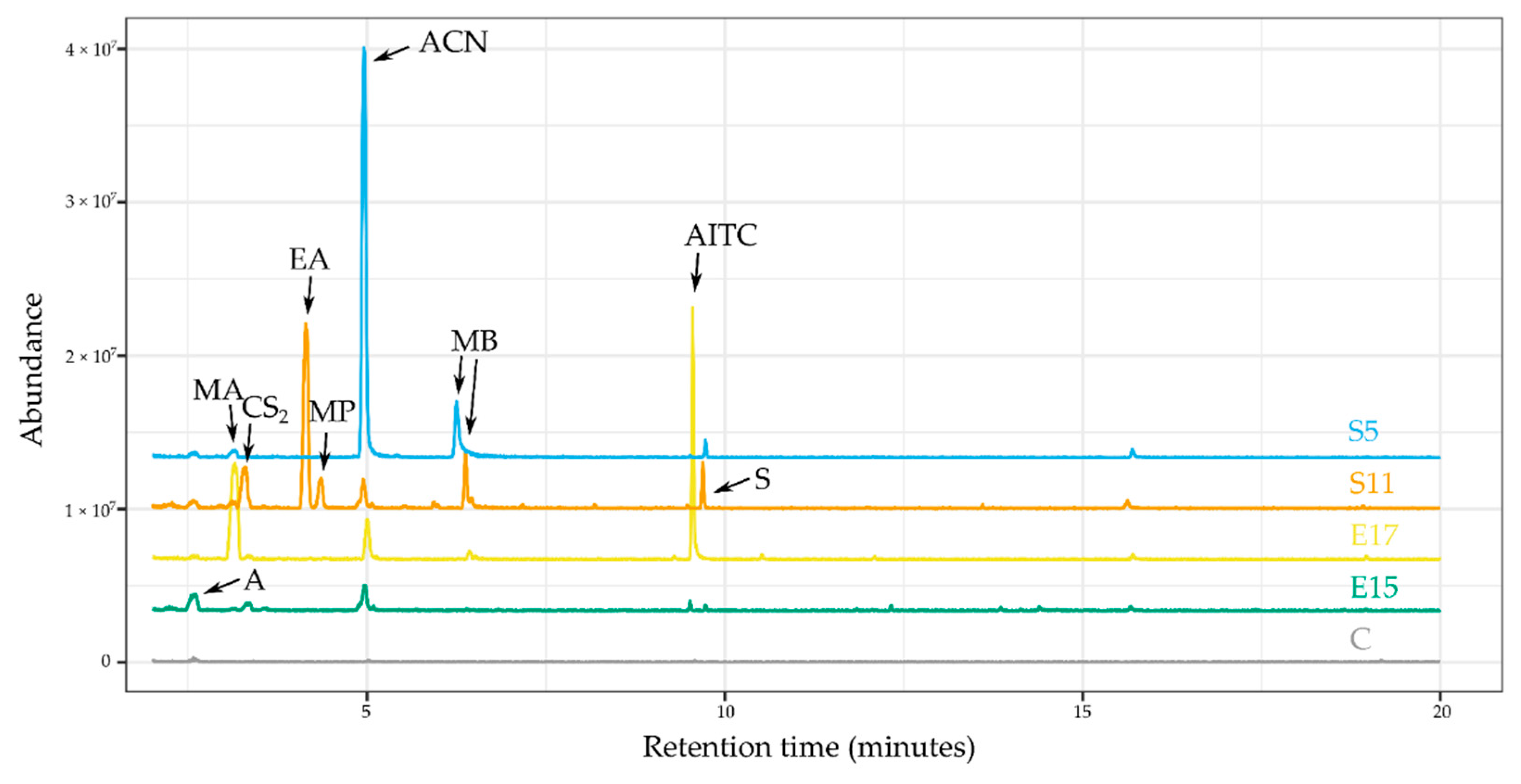
| Compound | Chemical Class | Retention Time (min) | Most Abundant Peaks * (m/z) | Identification Method (Similarity Score) |
|---|---|---|---|---|
| methyl-1-propanol | alcohol | 4.42 | 56, 74 | NIST library (818) |
| methyl-1-butanol isomer 1 | alcohol | 6.49 | 55, 57, 70 | NIST library (880) |
| methyl-1-butanol isomer 2 | alcohol | 6.55 | 55, 57, 70 | NIST library (908) |
| styrene | aromatic | 9.80 | 104, 78, 51 | NIST library (855) |
| benzaldehyde | aromatic | 11.17 | 51, 77, 106 | NIST library (819) |
| methyl formate | ester | 2.09 | 60 | authentic standard |
| methyl acetate | ester | 2.99 | 74 | NIST library (926) |
| ethyl acetate | ester | 4.20 | 61, 70, 88 | authentic standard |
| ethyl propionate | ester | 6.03 | 57 | NIST library (762) |
| propyl acetate | ester | 6.09 | 61, 73 | NIST library (711) |
| methyl-1-butanol acetate | ester | 9.32 | 72, 99 | NIST library (703) |
| acetone | ketone | 2.58 | 58 | authentic standard |
| beta-phellandrene | monoterpene | 12.41 | 77, 93, 136 | NIST library (852) |
| allyl cyanide (ACN) | nitrile | 5.02 | 52, 67 | authentic standard |
| phenylpropionitrile (PCN) | nitrile | 15.85 | 65, 91, 131 | authentic standard |
| acetic acid | organic acid | 3.58 | 60 | NIST library (661) |
| dimethyl sulfide (Me2S) | organo-sulfur | 2.88 | 62 | authentic standard |
| carbon disulfide (CS2) | organo-sulfur | 3.14 | 76 | authentic standard |
| allyl isothiocyanate (AITC) | organo-sulfur | 9.58 | 60, 72, 99 | authentic standard |
| 2-phenylethyl isothiocyanate (PEITC) | organo-sulfur | 19.18 | 65, 91, 163 | authentic standard |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plaszkó, T.; Szűcs, Z.; Kállai, Z.; Csoma, H.; Vasas, G.; Gonda, S. Volatile Organic Compounds (VOCs) of Endophytic Fungi Growing on Extracts of the Host, Horseradish (Armoracia rusticana). Metabolites 2020, 10, 451. https://doi.org/10.3390/metabo10110451
Plaszkó T, Szűcs Z, Kállai Z, Csoma H, Vasas G, Gonda S. Volatile Organic Compounds (VOCs) of Endophytic Fungi Growing on Extracts of the Host, Horseradish (Armoracia rusticana). Metabolites. 2020; 10(11):451. https://doi.org/10.3390/metabo10110451
Chicago/Turabian StylePlaszkó, Tamás, Zsolt Szűcs, Zoltán Kállai, Hajnalka Csoma, Gábor Vasas, and Sándor Gonda. 2020. "Volatile Organic Compounds (VOCs) of Endophytic Fungi Growing on Extracts of the Host, Horseradish (Armoracia rusticana)" Metabolites 10, no. 11: 451. https://doi.org/10.3390/metabo10110451
APA StylePlaszkó, T., Szűcs, Z., Kállai, Z., Csoma, H., Vasas, G., & Gonda, S. (2020). Volatile Organic Compounds (VOCs) of Endophytic Fungi Growing on Extracts of the Host, Horseradish (Armoracia rusticana). Metabolites, 10(11), 451. https://doi.org/10.3390/metabo10110451