Metabolomic Alteration in the Mouse Distal Colonic Mucosa after Oral Gavage with Oxalobacter formigenes



Abstract
1. Introduction
2. Results and Discussion
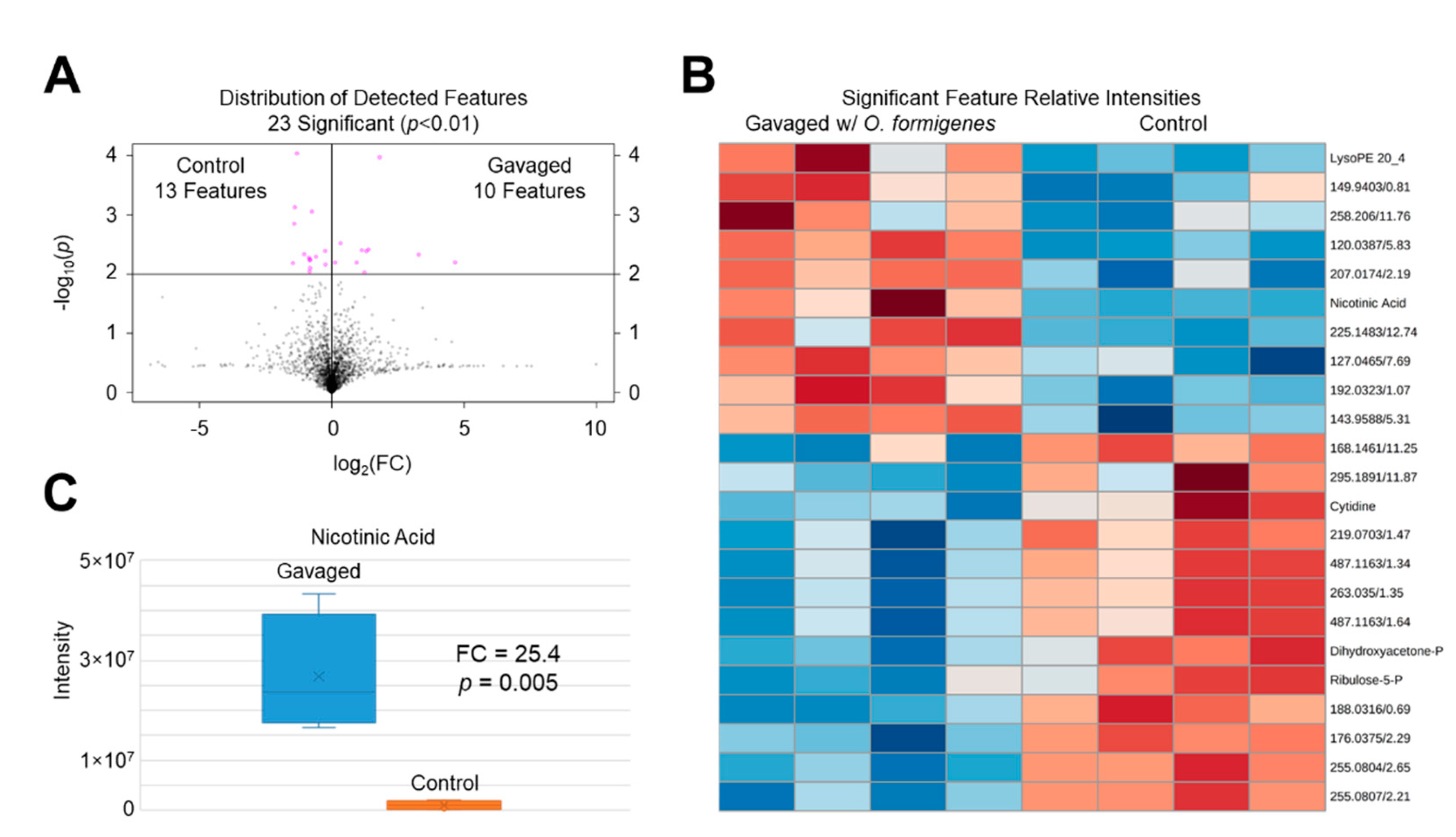
2.1. Distal Colonic Metabolome Alteration with O. formigenes Oral Gavage
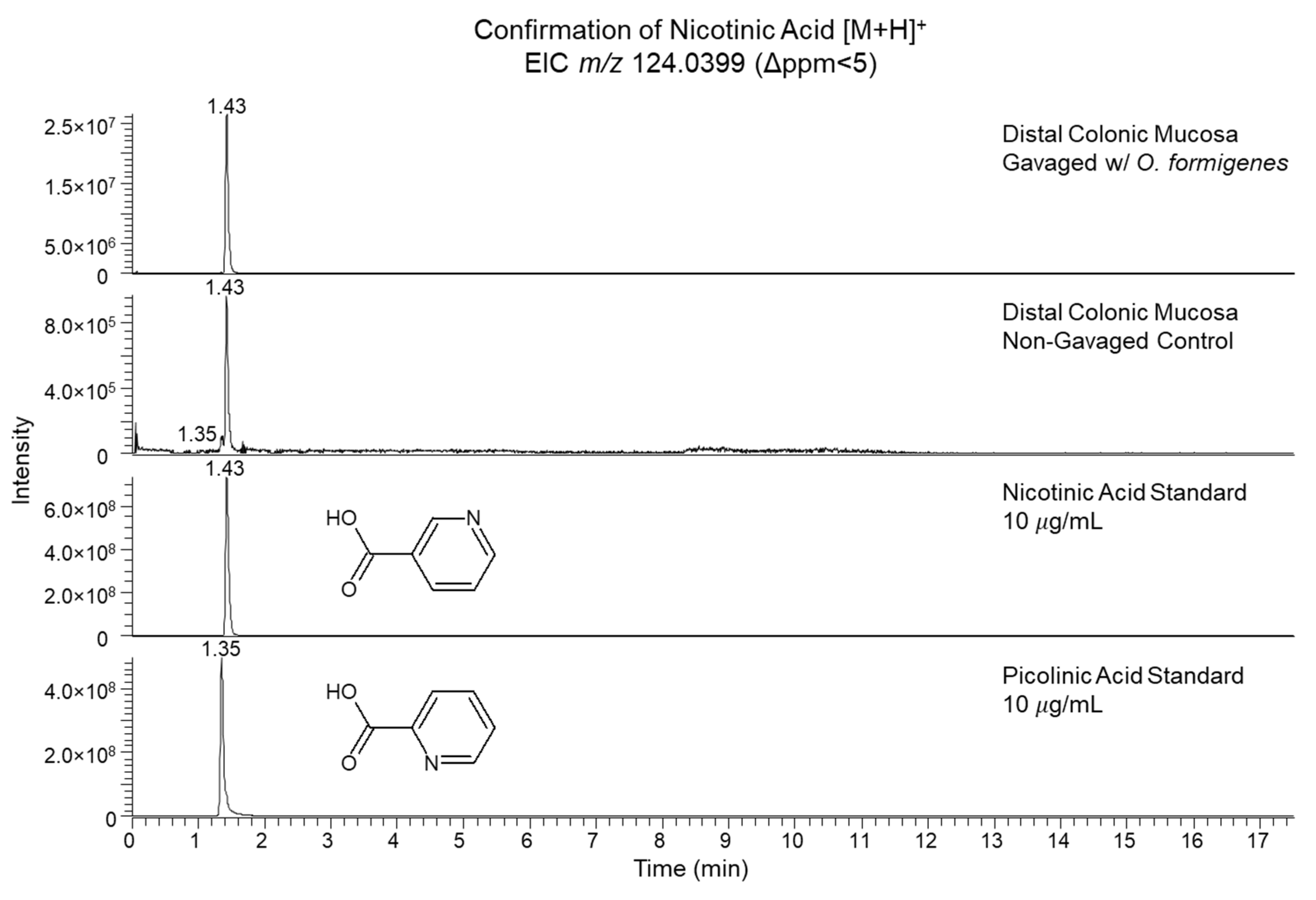
2.2. Assessment of the Gavage Effect on Significant Features
3. Materials and Methods
3.1. Use of Animals
3.2. Gavaging and Tissue Harvest
3.3. Pre-Extraction Sample Preparation
3.4. Metabolite Extraction
3.5. Analytical Instrumentation and Methodology
3.6. Data Processing, Quality Control, and Statistical Analysis
4. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Almeida, A.; Mitchell, A.L.; Boland, M.; Forster, S.C.; Gloor, G.B.; Tarkowska, A.; Lawley, T.D.; Finn, R.D. A new genomic Bluepr. of the human gut microbiota. Nature 2019, 568, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Nazzal, L. Enteric hyperoxaluria: Role of microbiota and antibiotics. Curr. Opin. Nephrol. Hypertens. 2019, 28, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Dawson, K.A.; Allison, M.J.; Hartman, P.A. Isolation and some characteristics of anaerobic oxalate-degrading bacteria from the rumen. Appl. Environ. Microbiol. 1980, 40, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Holmes, R.P.; Assimos, D.G. Glyoxylate synthesis, and its modulation and influence on oxalate synthesis. J. Urol. 1998, 160, 1617–1624. [Google Scholar] [CrossRef]
- Holmes, R.P.; Goodman, H.O.; Assimos, D.G. Contribution of dietary oxalate to urinary oxalate excretion. Kidney Int. 2001, 59, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Worcester, E.M.; Coe, F.L. Nephrolithiasis. Prim. Care 2008, 35, 369–391. [Google Scholar] [CrossRef]
- Miller, A.W.; Dearing, D. The metabolic and ecological interactions of oxalate-degrading bacteria in the Mammalian gut. Pathogens 2013, 2, 636–652. [Google Scholar] [CrossRef]
- Hatch, M. Gut microbiota and oxalate homeostasis. Ann. Transl. Med. 2017, 5, 36. [Google Scholar] [CrossRef]
- Li, X.; Ellis, M.L.; Knight, J. Oxalobacter formigenes Colonization and Oxalate Dynamics in a Mouse Model. Appl. Environ. Microbiol. 2015, 81, 5048–5054. [Google Scholar] [CrossRef]
- Allison, M.J.; Dawson, K.A.; Mayberry, W.R.; Foss, J.G. Oxalobacter formigenes gen. nov., sp. nov.: oxalate-degrading anaerobes that inhabit the gastrointestinal tract. Arch. Microbiol. 1985, 141, 1–7. [Google Scholar]
- Hatch, M.; Cornelius, J.; Allison, M.; Sidhu, H.; Peck, A.; Freel, R.W. Oxalobacter sp. reduces urinary oxalate excretion by promoting enteric oxalate secretion. Kidney Int. 2006, 69, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Hatch, M.; Gjymishka, A.; Salido, E.C.; Allison, M.J.; Freel, R.W. Enteric oxalate elimination is induced and oxalate is normalized in a mouse model of primary hyperoxaluria following intestinal colonization with Oxalobacter. Am. J. Physiol. Gastrointest Liver Physiol. 2011, 300, G461–G469. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, J.; Schwartz, R.A.; Hegyi, V. Pellagra: Dermatitis, dementia, and diarrhea. Int. J. Dermatol. 2004, 43, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Redzic, S.; Gupta, V. Niacin Deficiency; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Chamberlain, C.A.; Hatch, M.; Garrett, T.J. Metabolomic and lipidomic characterization of Oxalobacter formigenes strains HC1 and OxWR by UHPLC-HRMS. Anal. Bioanal. Chem. 2019. [Google Scholar] [CrossRef]
- Chamberlain, C.A.; Rubio, V.Y.; Garrett, T.J. Strain-Level Differentiation of Bacteria by Paper Spray Ionization Mass Spectrometry. Anal. Chem. 2019, 91, 4964–4968. [Google Scholar] [CrossRef]
- Ellis, M.E.; Mobley, J.A.; Holmes, R.P.; Knight, J. Proteome Dynamics of the Specialist Oxalate Degrader Oxalobacter formigenes. J. Proteomics Bioinform. 2016, 9, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Patti, G.J.; Yanes, O.; Siuzdak, G. Innovation: Metabolomics: The apogee of the omics trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269. [Google Scholar] [CrossRef]
- Rao, G.A.; Abraham, S. Triglyceride synthesis from dihydroxyacetone phosphate and palmitate by microsomes from mammary glands of lactating mice. Lipids 1978, 13, 95–98. [Google Scholar] [CrossRef]
- Coleman, R.A.; Mashek, D.G. Mammalian triacylglycerol metabolism: Synthesis, lipolysis, and signaling. Chem. Rev. 2011, 111, 6359–6386. [Google Scholar] [CrossRef]
- Hajra, A.K.; Larkins, L.K.; Das, A.K.; Hemati, N.; Erickson, R.L.; MacDougald, O.A. Induction of the peroxisomal glycerolipid-synthesizing enzymes during differentiation of 3T3-L1 adipocytes. Role in triacylglycerol synthesis. J. Biol. Chem. 2000, 275, 9441–9446. [Google Scholar] [CrossRef]
- Ahmadian, M.; Duncan, R.E.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Triacylglycerol metabolism in adipose tissue. Future Lipidol. 2007, 2, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Stincone, A.; Prigione, A.; Cramer, T.; Wamelink, M.M.; Campbell, K.; Cheung, E.; Olin-Sandoval, V.; Grüning, N.; Krüger, A.; Alam, M.T.; et al. The return of metabolism: Biochemistry and physiology of the pentose phosphate pathway. Biol. Rev. Camb. Philos. Soc. 2015, 90, 927–963. [Google Scholar] [CrossRef] [PubMed]
- Epand, R.M.; Epand, R.F. Lipid domains in bacterial membranes and the action of antimicrobial agents. Biochim. Biophys. Acta. 2009, 1788, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Law, S.H.; Chan, M.L.; Marathe, G.K.; Parveen, F.; Chen, C.H.; Ke, L.Y. An Updated Review of Lysophosphatidylcholine Metabolism in Human Diseases. Int. J. Mol. Sci. 2019, 20, 1149. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.B.; Karlsson, B.H.; Ozgen, M.; Palta, J.P. Inhibition of phospholipase D by lysophosphatidylethanolamine, a lipid-derived senescence retardant. Proc. Natl. Acad. Sci. USA 1997, 94, 12717–12721. [Google Scholar] [CrossRef] [PubMed]
- Fukuwatari, T.; Shibata, K. Nutritional aspect of tryptophan metabolism. Int. J. Tryptophan. Res. 2013, 6, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, K.J. The relationship of pellagra to corn and the low availability of niacin in cereals. Experientia. Suppl. 1983, 44, 197–222. [Google Scholar] [CrossRef]
- Savvidou, S. Pellagra: A non-eradicated old disease. Clin. Pract. 2014, 4, 637. [Google Scholar] [CrossRef]
- Fu, L.; Doreswamy, V.; Prakash, R. The biochemical pathways of central nervous system neural degeneration in niacin deficiency. Neural. Regen. Res. 2014, 9, 1509–1513. [Google Scholar] [CrossRef]
- Morowitz, M.J.; Carlisle, E.M.; Alverdy, J.C. Contributions of intestinal bacteria to nutrition and metabolism in the critically ill. Surg. Clin. N. Am. 2011, 91, 771–785. [Google Scholar] [CrossRef]
- Chamberlain, C.A.; Hatch, M.; Garrett, T.J. Metabolomic profiling of oxalate-degrading probiotic Lactobacillus acidophilus and Lactobacillus gasseri. PLoS ONE 2019, 14, e0222393. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, C.A.; Rubio, V.Y.; Garrett, T.J. Impact of matrix effects and ionization efficiency in non-quantitative untargeted metabolomics. Metabolomics 2019, 15, 135. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Diedrich, J.; Chu, Y.Y.; Yates, J.R. Extracting Accurate Precursor Information for Tandem Mass Spectra by RawConverter. Anal. Chem. 2015, 87, 11361–11367. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Pfann, C.; Herbold, C.; Daims, H.; Loy, A.; et al. METLIN: A Technology Platform for Identifying Knowns and Unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef] [PubMed]
- Wehrens, R.; Hageman, J.A.; van Eeuwijk, F.; Kooke, R.; Flood, P.J.; Wijnker, E.; Keurentjes, J.J.; Lommen, A.; van Eekelen, H.D.; Hall, R.D.; et al. Improved batch correction in untargeted MS-based metabolomics. Metabolomics 2016, 12, 88. [Google Scholar] [CrossRef]
- van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genomics 2006, 7, 142. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Significant Identified Metabolites | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| m/z | Metabolite (PubChem CID) | ID Method | Chemical Formula (M) | Ion | Δppm | RT | p-Value | FC | Response | ||
| 244.0925 | Cytidine (6175) | m/z, RT | C9H13N3O5 | [M + H]+ | 3.3 | 1.55 | 4.1 × 10−3 | 1.2 | Reduced | ||
| 171.0053 | Dihydroxyacetone Phosphate (668) | m/z, RT | C3H7O6P | [M + H]+ | 3.5 | 0.74 | 4.6 × 10−3 | 2.1 | Reduced | ||
| 502.2925 | LysoPE (20:4) (42607465) | m/z, RT | C25H44NO7P | [M + H]+ | 1.8 | 12.73 | 4.7 × 10−3 | 9.7 | Elevated | ||
| 124.0395 | Nicotinic Acid (938) | m/z, RT | C6H5NO2 | [M + H]+ | 3.2 | 1.43 | 6.3 × 10−3 | 25.2 | Elevated | ||
| 231.0261 | Ribulose-5-Phosphate (439184) | m/z, RT | C5H11O8P | [M + H]+ | 3.9 | 0.78 | 9.1 × 10−3 | 1.8 | Reduced | ||
| Significant Unidentified Metabolites | |||||||||||
| m/z | (13C/12C) ×100 | # Carbons | Sulfur | Charge | Putative Formula (M) | Ion | Δppm | RT | p-Value | FC | Response |
| 127.0465 | 9.89 ± 1.59 | 8.99 ± 1.44 | Y | +2 | C8H16N2O5S | [M + 2H]2+ | 1.6 | 7.69 | 9.4 × 10−3 | 2.4 | Elevated |
| 143.9588 a | 15.54 ± 8.27 | 14.13 ± 7.51 | n | +1 | Undetermined | 5.31 | 4.1 × 10−3 | 2.5 | Elevated | ||
| 176.0375 | 5.71 ± 1.30 | 5.19 ± 1.18 | Y | +1 | C6H9NO3S | [M + H]+ | 0.6 | 2.29 | 8.8 × 10−4 | 1.7 | Reduced |
| 192.0323 | 7.48 ± 3.31 | 6.80 ± 3.01 | n | +1 | C8H10O2P | [M + Na]+ | 4.7 | 1.07 | 3.9 × 10−3 | 2.2 | Elevated |
| 219.0703 | 17.66 ± 4.85 | 16.06 ± 4.41 | Y | +2 | C15H24N4O9S | [M + 2H]2+ | 0.9 | 1.47 | 5.3 × 10−3 | 1.8 | Reduced |
| 255.0804 b | 22.85 ± 5.95 | 20.77 ± 5.41 | Y | +2 | C18H28N4O11S | [M + 2H]2+ | 2.4 | 2.65 | 9.2 × 10−5 | 2.5 | Reduced |
| 255.0807 b | 17.95 ± 6.93 | 16.32 ± 6.30 | Y | +2 | C18H28N4O11S | [M + 2H]2+ | 1.2 | 2.21 | 7.5 × 10−4 | 2.6 | Reduced |
| 263.0350 | 18.85 ± 2.29 | 17.14 ± 2.08 | Y | +2 | C19H22N2O10S2 | [M + H + Na]2+ | 3.8 | 1.35 | 5.7 × 10−3 | 1.8 | Reduced |
| 487.1163 c | 18.97 ± 1.64 | 17.24 ± 1.50 | Y | +1 | C16H30N4O5S4 | [M + H]+ | 1.8 | 1.34 | 5.8 × 10−3 | 1.8 | Reduced |
| 487.1163 c | 18.69 ± 2.18 | 16.99 ± 1.98 | Y | +1 | C16H30N4O5S4 | [M + H]+ | 1.8 | 1.64 | 7.9 × 10−3 | 1.8 | Reduced |
| 120.0387 | Undetermined | C15H10O3 | [M + 2H]2+ | 0.8 | 5.83 | 1.1 × 10−4 | 3.5 | Elevated | |||
| 149.9403 a | Undetermined | 0.81 | 3.0 × 10−3 | 1.3 | Elevated | ||||||
| 168.1461 a | Undetermined | 11.25 | 6.5 × 10−3 | 2.8 | Reduced | ||||||
| 188.0316 | Undetermined | C6H9N3S2 | [M + H]+ | 2.7 | 0.69 | 1.4 × 10−3 | 2.7 | Reduced | |||
| 207.0174 | Undetermined | C4H6N4O4S | [M + H]+ | 4.3 | 2.19 | 3.8 × 10−3 | 2.6 | Elevated | |||
| 225.1483 | Undetermined | C13H20O3 | [M + H]+ | 0.9 | 12.74 | 6.4 × 10−3 | 1.9 | Elevated | |||
| 258.2060 | Undetermined | C14H27NO3 | [M + H]+ | 1.5 | 11.76 | 6.4 × 10−3 | 1.1 | Elevated | |||
| 295.1891 | Undetermined | C17H26O4 | [M + H]+ | 4.4 | 11.87 | 5.1 × 10−3 | 1.5 | Reduced | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chamberlain, C.A.; Hatch, M.; Garrett, T.J. Metabolomic Alteration in the Mouse Distal Colonic Mucosa after Oral Gavage with Oxalobacter formigenes. Metabolites 2020, 10, 405. https://doi.org/10.3390/metabo10100405
Chamberlain CA, Hatch M, Garrett TJ. Metabolomic Alteration in the Mouse Distal Colonic Mucosa after Oral Gavage with Oxalobacter formigenes. Metabolites. 2020; 10(10):405. https://doi.org/10.3390/metabo10100405
Chicago/Turabian StyleChamberlain, Casey A., Marguerite Hatch, and Timothy J. Garrett. 2020. "Metabolomic Alteration in the Mouse Distal Colonic Mucosa after Oral Gavage with Oxalobacter formigenes" Metabolites 10, no. 10: 405. https://doi.org/10.3390/metabo10100405
APA StyleChamberlain, C. A., Hatch, M., & Garrett, T. J. (2020). Metabolomic Alteration in the Mouse Distal Colonic Mucosa after Oral Gavage with Oxalobacter formigenes. Metabolites, 10(10), 405. https://doi.org/10.3390/metabo10100405