Possible Use of Blood Tryptophan Metabolites as Biomarkers for Coronary Heart Disease in Sudden Unexpected Death


Abstract
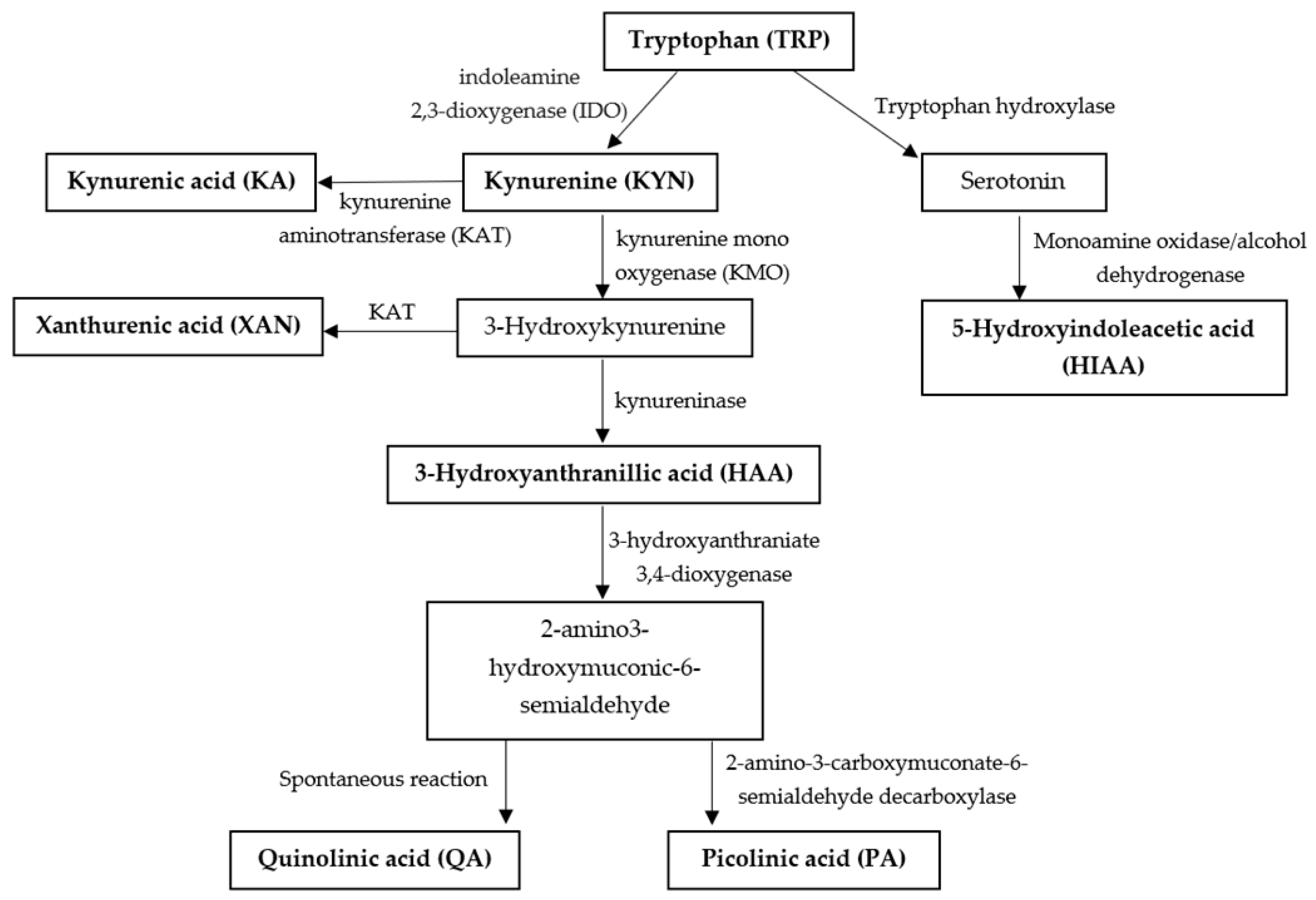
1. Introduction
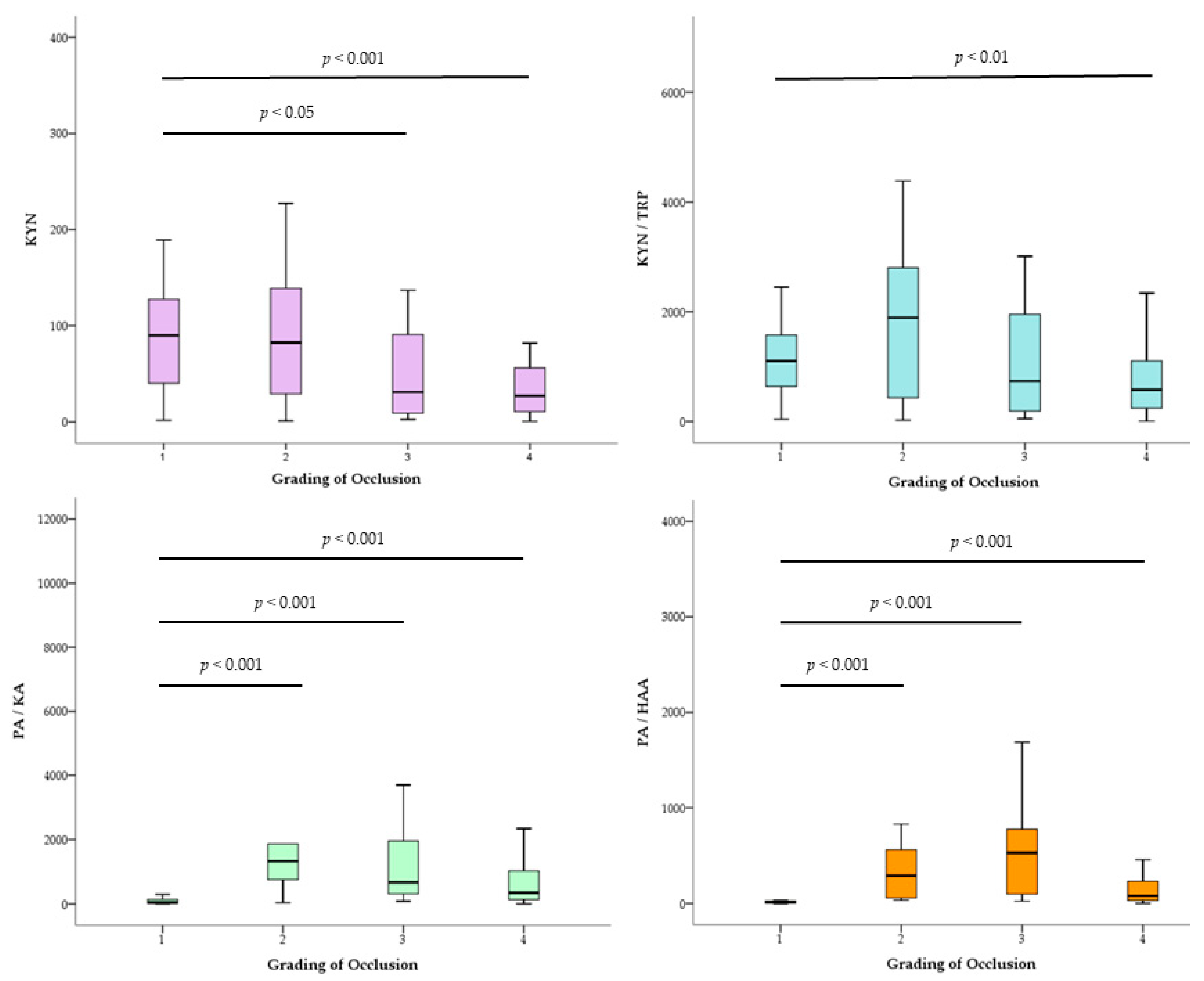
2. Results
3. Discussion
4. Materials and Methods
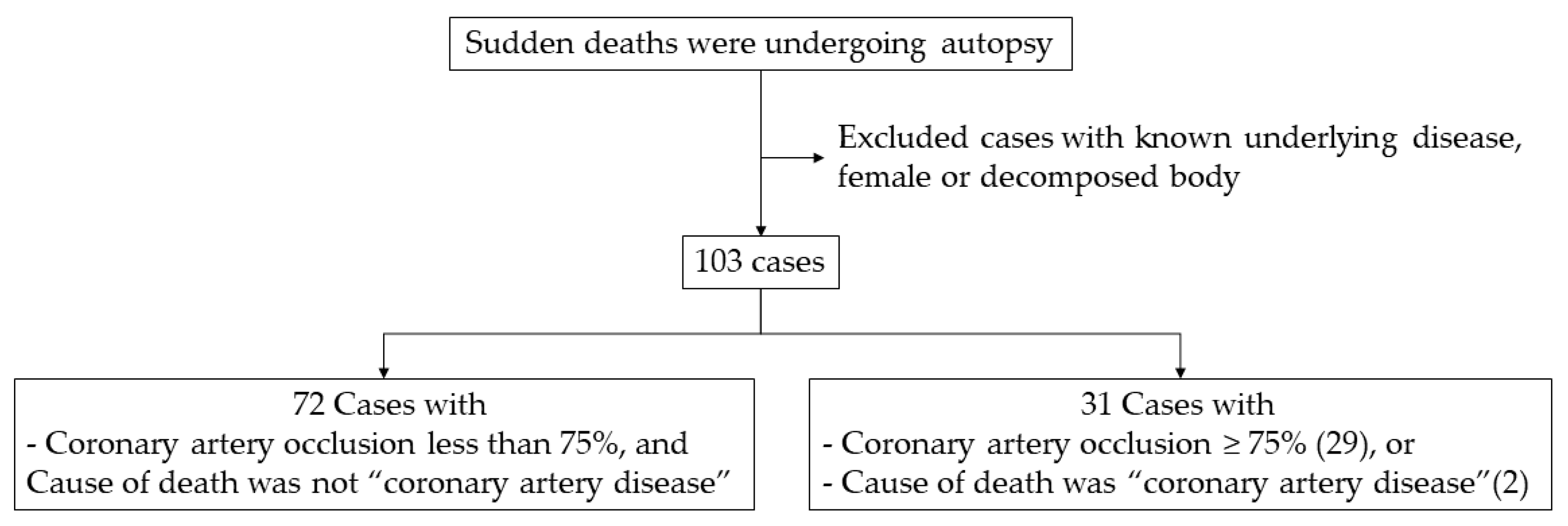
4.1. Subjects and Study Design
4.2. Collection and Specimen Preparation
4.3. Measurement of Tryptophan and Its Metabolites
4.4. Pathology
4.5. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CHD | Coronary heart disease |
| SUD | Sudden unexpected death |
| QA | Quinolinic acid |
| PA | Picolinic acid |
| HAA | 3-Hydroxyanthranillic acid |
| HIAA | 5-Hydroxyindoleacetic acid |
| KYN | Kynurenine |
| XAN | Xanthurenic acid |
| TRP | Tryptophan |
| KA | Kynurenic acid |
| IDO | Indoleamine 2, 3-dioxygenase |
References
- Kumar, V.; San, K.P.; Idwan, A.; Shah, N.; Hajar, S.; Norkahfi, M. A study of sudden natural deaths in medico legal autopsies in University Malaya Medical Centre (UMMC), Kuala Lumpur. J. Forensic Leg. Med. 2007, 14, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Udnoon, J.; Chirachariyavej, T.; Peonim, V. Sudden unexpected deaths in different age groups at Ramathibodi Hospital, Bangkok, Thailand: A retrospective autopsy study during 2003–2007. Southeast Asian J. Trop. Med. Public Health 2009, 40, 162. [Google Scholar] [PubMed]
- World Health Organization Cardiovascular Diseases (cvds). Available online: https://www.who.int/en/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 17 May 2017).
- Milroy, C.M. Coronary artery disease, sudden death and implications for forensic pathology practice. Diagn. Histopathol. 2017, 23, 479–485. [Google Scholar] [CrossRef]
- Basso, C.; Aguilera, B.; Banner, J.; Cohle, S.; d’Amati, G.; de Gouveia, R.H.; di Gioia, C.; Fabre, A.; Gallagher, P.J.; Leone, O. Guidelines for autopsy investigation of sudden cardiac death: 2017 update from the Association for European Cardiovascular Pathology. Virchows Arch. 2017, 471, 691–705. [Google Scholar] [CrossRef]
- Antman, E.; Bassand, J.-P.; Klein, W.; Ohman, M.; Sendon, J.L.L.; Rydén, L.; Simoons, M.; Tendera, M. Myocardial infarction redefined—A consensus document of the Joint European Society of Cardiology/American College of Cardiology committee for the redefinition of myocardial infarction: The Joint European Society of Cardiology/American College of Cardiology Committee. J. Am. Coll. Cardiol. 2000, 36, 959–969. [Google Scholar]
- Jaffe, A.S.; Ravkilde, J.; Roberts, R.; Naslund, U.; Apple, F.S.; Galvani, M.; Katus, H. It’s time for a change to a troponin standard. Am. Heart Assoc 2000, 102, 1216–1220. [Google Scholar] [CrossRef]
- Rahimi, R.; Dahili, N.D.; Zainun, K.A.; Kasim, N.A.M.; Noor, S.M. Post mortem troponin T analysis in sudden death: Is it useful? Malays. J. Pathol. 2018, 40, 143–148. [Google Scholar]
- Wirleitner, B.; Rudzite, V.; Neurauter, G.; Murr, C.; Kalnins, U.; Erglis, A.; Trusinskis, K.; Fuchs, D. Immune activation and degradation of tryptophan in coronary heart disease. Eur. J. Clin. Investig. 2003, 33, 550–554. [Google Scholar] [CrossRef]
- Song, P.; Ramprasath, T.; Wang, H.; Zou, M.-H. Abnormal kynurenine pathway of tryptophan catabolism in cardiovascular diseases. Cell. Mol. Life Sci. 2017, 74, 2899–2916. [Google Scholar] [CrossRef]
- Dehhaghi, M.; Kazemi Shariat Panahi, H.; Guillemin, G.J. Microorganisms, tryptophan metabolism, and kynurenine pathway: A complex interconnected loop influencing human health status. Int. J. Tryptophan Res. 2019, 12, 1178646919852996. [Google Scholar] [CrossRef]
- Ketelhuth, D.F. The immunometabolic role of indoleamine 2, 3-dioxygenase in atherosclerotic cardiovascular disease: Immune homeostatic mechanisms in the artery wall. Cardiovasc. Res. 2019, 115, 1408–1415. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.M.; Lee, S.Y.; Kim, Y.S.; Seo, J.S.; Lee, Y.S.; Seo, J.W. Guidelines for forensic assessment of natural unexpected cardiovascular death. Basic Appl. Pathol. 2008, 1, 155–163. [Google Scholar] [CrossRef]
- Ohashi, H.; Iizuka, H.; Yoshihara, S.; Otani, H.; Kume, M.; Sadamoto, K.; Ichiba, H.; Fukushima, T. Determination of l-tryptophan and l-kynurenine in human serum by using LC-MS after derivatization with (R)-DBD-PyNCS. Int. J. Tryptophan Res. 2013, 6, IJTR. S11459. [Google Scholar] [CrossRef]
- Cheong, J.E.; Sun, L. Targeting the IDO1/TDO2–KYN–AhR pathway for cancer immunotherapy–challenges and opportunities. Trends Pharmacol. Sci. 2018, 39, 307–325. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, K.; Ketelhuth, D. The role of the kynurenine pathway of tryptophan metabolism in cardiovascular disease. Hamostaseologie 2015, 35, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.-B.; Dougherty, D.M. Assessment of the human kynurenine pathway: Comparisons and clinical implications of ethnic and gender differences in plasma tryptophan, kynurenine metabolites, and enzyme expressions at baseline and after acute tryptophan loading and depletion. Int. J. Tryptophan Res. 2016, 9, IJTR. S38189. [Google Scholar] [CrossRef]
- Adaway, J.E.; Dobson, R.; Walsh, J.; Cuthbertson, D.J.; Monaghan, P.J.; Trainer, P.J.; Valle, J.W.; Keevil, B.G. Serum and plasma 5-hydroxyindoleacetic acid as an alternative to 24-h urine 5-hydroxyindoleacetic acid measurement. Ann. Clin. Biochem. 2016, 53, 554–560. [Google Scholar] [CrossRef]
- Zuwała-Jagiello, J.; Pazgan-Simon, M.; Simon, K.; Warwas, M. Picolinic acid in patients with chronic hepatitis C infection: A preliminary report. Mediat. Inflamm. 2012, 2012, 762863. [Google Scholar] [CrossRef][Green Version]
- Basile, A.S.; Saito, K.; Al-Mardini, H.; Record, C.O.; Hughes, R.D.; Harrison, P.; Williams, R.; Li, Y.; Heyes, M.P. The relationship between plasma and brain quinolinic acid levels and the severity of hepatic encephalopathy. Gastroenterology 1995, 108, 818–823. [Google Scholar] [CrossRef]
- Poesen, R.; Mutsaers, H.A.; Windey, K.; van den Broek, P.H.; Verweij, V.; Augustijns, P.; Kuypers, D.; Jansen, J.; Evenepoel, P.; Verbeke, K. The influence of dietary protein intake on mammalian tryptophan and phenolic metabolites. PLoS ONE 2015, 10, e0140820. [Google Scholar] [CrossRef]
- Brundin, L.; Sellgren, C.; Lim, C.; Grit, J.; Pålsson, E.; Landen, M.; Samuelsson, M.; Lundgren, K.; Brundin, P.; Fuchs, D. An enzyme in the kynurenine pathway that governs vulnerability to suicidal behavior by regulating excitotoxicity and neuroinflammation. Transl. Psychiatry 2016, 6, e865. [Google Scholar] [CrossRef] [PubMed]
- Lugo-Huitrón, R.; Ugalde Muñiz, P.; Pineda, B.; Pedraza-Chaverrí, J.; Ríos, C.; Pérez-De La Cruz, V. Quinolinic acid: An endogenous neurotoxin with multiple targets. Oxidative Med. Cell. Longev. 2013, 2013, 104024. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.; Coggan, S.; Smythe, G. The physiological action of picolinic acid in the human brain. Int. J. Tryptophan Res. 2009, 2, IJTR. S2469. [Google Scholar] [CrossRef]
- Leipnitz, G.; Schumacher, C.; Scussiato, K.; Dalcin, K.B.; Wannmacher, C.M.; Wyse, A.T.; Dutra-Filho, C.S.; Wajner, M.; Latini, A. Quinolinic acid reduces the antioxidant defenses in cerebral cortex of young rats. Int. J. Dev. Neurosci. 2005, 23, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, D.; Song, P.; Zou, M.-H. Deregulated tryptophan-kynurenine pathway is linked to inflammation, oxidative stress, and immune activation pathway in cardiovascular diseases. Front. Biosci. 2015, 20, 1116. [Google Scholar]
- Liu, G.; Chen, S.; Zhong, J.; Teng, K.; Yin, Y. Crosstalk between tryptophan metabolism and cardiovascular disease, mechanisms, and therapeutic implications. Oxidative Med. Cell. Longev. 2017, 2017, 1602074. [Google Scholar] [CrossRef]
- Zhang, L.; Ovchinnikova, O.; Jönsson, A.; Lundberg, A.M.; Berg, M.; Hansson, G.K.; Ketelhuth, D.F. The tryptophan metabolite 3-hydroxyanthranilic acid lowers plasma lipids and decreases atherosclerosis in hypercholesterolaemic mice. Eur. Heart J. 2012, 33, 2025–2034. [Google Scholar] [CrossRef]
- Giles, G.I.; Collins, C.A.; Stone, T.W.; Jacob, C. Electrochemical and in vitro evaluation of the redox-properties of kynurenine species. Biochem. Biophys. Res. Commun. 2003, 300, 719–724. [Google Scholar] [CrossRef]
- Darlington, L.G.; Forrest, C.M.; Mackay, G.M.; Smith, R.A.; Smith, A.J.; Stoy, N.; Stone, T.W. On the biological importance of the 3-hydroxyanthranilic acid: Anthranilic acid ratio. Int. J. Tryptophan Res. 2010, 3, IJTR. S4282. [Google Scholar] [CrossRef]
- Meier, M.A.; Ottiger, M.; Vögeli, A.; Steuer, C.; Bernasconi, L.; Thomann, R.; Christ-Crain, M.; Henzen, C.; Hoess, C.; Zimmerli, W. Activation of the tryptophan/serotonin pathway is associated with severity and predicts outcomes in pneumonia: Results of a long-term cohort study. Clin. Chem. Lab. Med. (Cclm) 2017, 55, 1060–1069. [Google Scholar] [CrossRef]
- Heyes, M.P.; Saito, K.; Major, E.O.; Milstien, S.; Markey, S.P.; Vickers, J.H. A mechanism of quinolinic acid formation by brain in inflammatory neurological disease: Attenuation of synthesis from L-tryptophan by 6-chlorotryptophan and 4-chloro-3-hydroxyanthranilate. Brain 1993, 116, 1425–1450. [Google Scholar] [CrossRef] [PubMed]
- Zuo, H.; Ueland, P.M.; Ulvik, A.; Eussen, S.J.; Vollset, S.E.; Nygård, O.; Midttun, Ø.; Theofylaktopoulou, D.; Meyer, K.; Tell, G.S. Plasma biomarkers of inflammation, the kynurenine pathway, and risks of all-cause, cancer, and cardiovascular disease mortality: The Hordaland Health Study. Am. J. Epidemiol. 2016, 183, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Gobaille, S.; Kemmel, V.; Brumaru, D.; Dugave, C.; Aunis, D.; Maitre, M. Xanthurenic acid distribution, transport, accumulation and release in the rat brain. J. Neurochem. 2008, 105, 982–993. [Google Scholar] [CrossRef] [PubMed]
- Reyes Ocampo, J.; Lugo Huitrón, R.; González-Esquivel, D.; Ugalde-Muñiz, P.; Jiménez-Anguiano, A.; Pineda, B.; Pedraza-Chaverri, J.; Ríos, C.; Pérez de la Cruz, V. Kynurenines with neuroactive and redox properties: Relevance to aging and brain diseases. Oxidative Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Haneda, M.; Yoshino, M. Prooxidant action of xanthurenic acid and quinoline compounds: Role of transition metals in the generation of reactive oxygen species and enhanced formation of 8-hydroxy-2′-deoxyguanosine in DNA. Biometals 2006, 19, 429–435. [Google Scholar] [CrossRef]
- Konradsson-Geuken, Å.; Wu, H.; Gash, C.; Alexander, K.; Campbell, A.; Sozeri, Y.; Pellicciari, R.; Schwarcz, R.; Bruno, J. Cortical kynurenic acid bi-directionally modulates prefrontal glutamate levels as assessed by microdialysis and rapid electrochemistry. Neuroscience 2010, 169, 1848–1859. [Google Scholar] [CrossRef]
- Zmarowski, A.; Wu, H.Q.; Brooks, J.; Potter, M.; Pellicciari, R.; Schwarcz, R.; Bruno, J. Astrocyte-derived kynurenic acid modulates basal and evoked cortical acetylcholine release. Eur. J. Neurosci. 2009, 29, 529–538. [Google Scholar] [CrossRef]
- Hardeland, R.; Zsizsik, B.; Poeggeler, B.; Fuhrberg, B.; Holst, S.; Coto-Montes, A. Indole-3-pyruvic and-propionic acids, kynurenic acid, and related metabolites as luminophores and free-radical scavengers. In Tryptophan, Serotonin, and Melatonin; Springer: Berlin, Germany, 1999; pp. 389–395. [Google Scholar]
- Pedersen, E.R.; Tuseth, N.; Eussen, S.J.; Ueland, P.M.; Strand, E.; Svingen, G.F.T.; Midttun, Ø.; Meyer, K.; Mellgren, G.; Ulvik, A. Associations of plasma kynurenines with risk of acute myocardial infarction in patients with stable angina pectoris. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 455–462. [Google Scholar] [CrossRef]
- Schmidt, S.K.; Ebel, S.; Keil, E.; Woite, C.; Ernst, J.F.; Benzin, A.E.; Rupp, J.; Däubener, W. Regulation of IDO activity by oxygen supply: Inhibitory effects on antimicrobial and immunoregulatory functions. PLoS ONE 2013, 8, e63301. [Google Scholar] [CrossRef]
- Chen, T.; Zheng, X.; Ma, X.; Bao, Y.; Ni, Y.; Hu, C.; Rajani, C.; Huang, F.; Zhao, A.; Jia, W. Tryptophan predicts the risk for future type 2 diabetes. PLoS ONE 2016, 11, e0162192. [Google Scholar] [CrossRef]
- Zhou, C.C.; Yang, X.; Hua, X.; Liu, J.; Fan, M.B.; Li, G.Q.; Song, J.; Xu, T.Y.; Li, Z.Y.; Guan, Y.F. Hepatic NAD+ deficiency as a therapeutic target for non-alcoholic fatty liver disease in ageing. Br. J. Pharmacol. 2016, 173, 2352–2368. [Google Scholar] [CrossRef] [PubMed]
- Haruki, H.; Hovius, R.; Pedersen, M.G.; Johnsson, K. Tetrahydrobiopterin biosynthesis as a potential target of the kynurenine pathway metabolite xanthurenic acid. J. Biol. Chem. 2016, 291, 652–657. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Metabolites | Range | Total (n = 103) | CHD Group (n = 31) | Control Group (n = 72) | p-Value |
|---|---|---|---|---|---|
| TRP (mM) | 0.04–0.17 | 0.06 ± 0.03 | 0.06 ± 0.02 | 0.07 ± 0.03 | 0.076 |
| KYN (mM) | 0.70–306.87 | 71.68 ± 60.88 | 41.98 ± 47.68 | 84.47 ± 61.74 | <0.001 |
| KA (mM) | 0–0.07 | 0.005 ± 0.01 | 0.003 ± 0.004 | 0.007 ± 0.01 | 0.078 |
| XAN (mM) | 0–0.28 | 0.03 ± 0.04 | 0.03 ± 0.04 | 0.02 ± 0.04 | 0.131 |
| HAA (mM) | 0–0.88 | 0.02 ± 0.09 | 0.01 ± 0.03 | 0.02 ± 0.10 | 0.062 |
| QA (mM) | 0.04–4.07 | 0.61 ± 0.85 | 0.30 ± 0.23 | 0.74 ± 0.98 | 0.124 |
| PA (mM) | 0–7.81 | 0.74 ± 1.11 | 0.83 ± 1.04 | 0.70 ± 1.15 | 0.165 |
| HIAA (mM) | 0–364.59 | 22.75 ± 44.12 | 27.45 ± 70.41 | 20.73 ± 26.22 | 0.018 |
| KYN/XAN (×104) | 0–17.83 | 1.48 ± 2.66 | 0.73 ± 1.51 | 1.78 ± 2.97 | 0.001 |
| KYN/TRP (×103) | 0–7.00 | 1.15 ± 1.08 | 0.72 ± 0.70 | 1.34 ± 1.16 | 0.002 |
| XAN/KA | 0–229.84 | 18.83 ± 36.76 | 32.22 ± 53.56 | 13.06 ± 24.85 | 0.015 |
| HAA/XAN | 0–32.33 | 1.94 ± 3.94 | 1.17 ± 1.92 | 2.27 ± 4.52 | 0.029 |
| QA/XAN (×103) | 0–1.30 | 0.11 ± 0.23 | 0.05 ± 0.07 | 0.14 ± 0.26 | 0.018 |
| PA/HAA (×103) | 0–3.33 | 0.25 ± 0.52 | 0.33 ± 0.68 | 0.22 ± 0.44 | 0.043 |
| PA/HIAA | 0–4.37 | 0.13 ± 0.46 | 0.29 ± 0.81 | 0.06 ± 0.14 | 0.028 |
| PA/KYN | 0–0.45 | 0.03 ± 0.06 | 0.05 ± 0.09 | 0.02 ± 0.04 | 0.001 |
| PA/KA (×104) | 0–1.11 | 0.07 ± 0.13 | 0.06 ± 0.07 | 0.07 ± 0.15 | 0.044 |
| Age (years) | 21–86 | 54.48 ± 13.88 | 58.16 ± 13.34 | 52.89 ± 13.90 | 0.056 |
| Metabolites | Correlation Coefficient (r) | p-Value |
|---|---|---|
| TRP | −0.356 | <0.001 |
| KYN | −0.358 | <0.001 |
| KA | −0.322 | 0.001 |
| HAA | −0.309 | 0.001 |
| QA | −0.293 | 0.003 |
| PA | 0.361 | <0.001 |
| TRP/KA | 0.215 | 0.029 |
| KYN/XAN | −0.275 | 0.005 |
| KYN/TRP | −0.253 | 0.01 |
| XAN/KA | 0.251 | 0.01 |
| HAA/TRP | −0.228 | 0.02 |
| HAA/XAN | −0.207 | 0.036 |
| QA/PA | −0.425 | <0.001 |
| QA/XAN | −0.202 | 0.04 |
| QA/TRP | −0.20 | 0.043 |
| PA/HAA | 0.437 | <0.001 |
| PA/HIAA | 0.383 | <0.001 |
| PA/KYN | 0.533 | <0.001 |
| PA/XAN | 0.247 | 0.012 |
| PA/TRP | 0.403 | <0.001 |
| PA/KA | 0.45 | <0.001 |
| HIAA/KYN | 0.278 | 0.004 |
| Metabolites | Trend Correlation with Coronary Occlusion Degree |
|---|---|
| TRP | − |
| KYN | − |
| KA | − |
| XAN | + |
| HAA | − |
| QA | − |
| PA | + |
| HIAA | + |
| KYN/TRP | − |
| PA/KA | + |
| PA/HAA | + |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santisukwongchote, K.; Amornlertwatana, Y.; Sastraruji, T.; Jaikang, C. Possible Use of Blood Tryptophan Metabolites as Biomarkers for Coronary Heart Disease in Sudden Unexpected Death. Metabolites 2020, 10, 6. https://doi.org/10.3390/metabo10010006
Santisukwongchote K, Amornlertwatana Y, Sastraruji T, Jaikang C. Possible Use of Blood Tryptophan Metabolites as Biomarkers for Coronary Heart Disease in Sudden Unexpected Death. Metabolites. 2020; 10(1):6. https://doi.org/10.3390/metabo10010006
Chicago/Turabian StyleSantisukwongchote, Kobchai, Yutti Amornlertwatana, Thanapat Sastraruji, and Churdsak Jaikang. 2020. "Possible Use of Blood Tryptophan Metabolites as Biomarkers for Coronary Heart Disease in Sudden Unexpected Death" Metabolites 10, no. 1: 6. https://doi.org/10.3390/metabo10010006
APA StyleSantisukwongchote, K., Amornlertwatana, Y., Sastraruji, T., & Jaikang, C. (2020). Possible Use of Blood Tryptophan Metabolites as Biomarkers for Coronary Heart Disease in Sudden Unexpected Death. Metabolites, 10(1), 6. https://doi.org/10.3390/metabo10010006