
Helicobacter pylori in Native Americans in Northern Arizona †

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Considerations
2.2. Patients and Sample Collection
2.3. H. pylori Strains and DNA Isolation
2.4. Molecular Identification
2.5. CagA Gene Amplification
2.6. vacA Gene Amplification
2.7. Statistical Analysis
3. Results
3.1. Demographic Information in Positive Patients
3.2. PCR Amplification of cagA Gene Variable 3′ Region and EPIYA Motif Bioinformatic Analysis
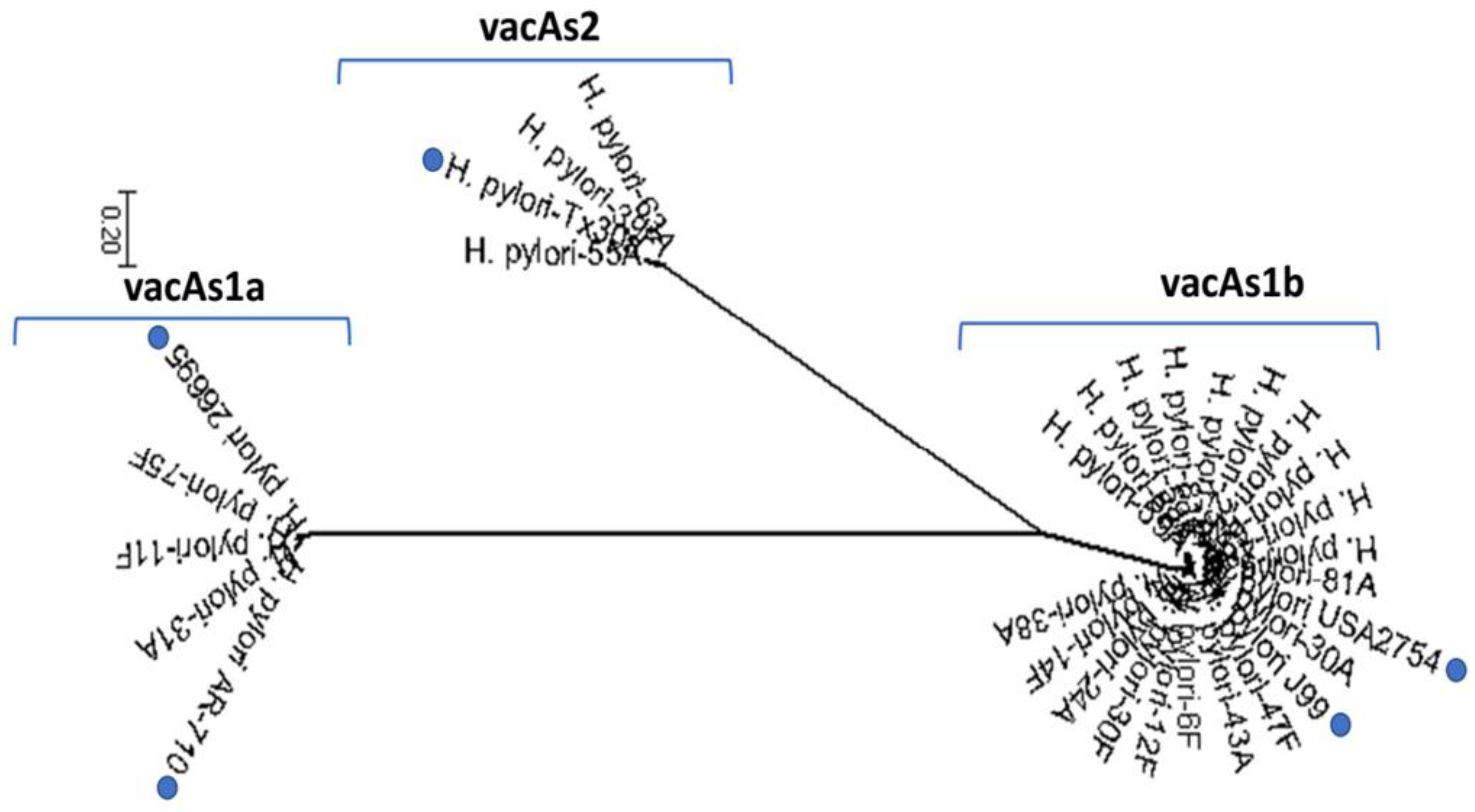
3.3. Presence of vacA in Native American Isolates
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Makola, D.; Peura, D.A.; Crowe, S.E. Helicobacter pylori infection and related gastrointestinal diseases. J. Clin. Gastroenterol. 2007, 41, 548–558. [Google Scholar] [CrossRef] [PubMed]
- De Martel, C.; Forman, D.; Plummer, D. Gastric Cancer. Epidemiology and Risk Factors. Gastroenterol. Clin. N. Am. 2013, 42, 219–240. [Google Scholar] [CrossRef] [PubMed]
- Perez-Perez, G.I.; Rothenbacher, D.; Brenner, H. Epidemiology of Helicobacter pylori Infection. Helicobacter 2004, 9, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Camargo, M.C.; Yepez, M.C.; Cerón, C.; Guerrero, N.; Bravo, L.E.; Correa, P.; Fontham, E.T. Age at acquisition of Helicobacter pylori infection: Comparison of two areas with contrasting risk of gastric cancer. Helicobacter 2004, 9, 262–270. [Google Scholar] [CrossRef]
- Harris, R.B.; Brown, H.E.; Begay, R.; Sanderson, P.; Chief, C.; Oren, E.; Monroy, F.P. Helicobacter pylori prevalence and risk factors in three rural Navajo communities of Northern Arizona: A cross-sectional study. Int. J. Environ. Res. Public Health 2022, 19, 797. [Google Scholar] [CrossRef]
- Everhart, J.E. Recent developments in the epidemiology of Helicobacter pylori. Gastroenterol. Clin. N. Am. 2000, 29, 559–578. [Google Scholar] [CrossRef]
- Blaser, M.J. Helicobacter pylori eradication and its implications for the future. Aliment. Pharmacol. Ther. 1997, 11, 103–107. [Google Scholar] [CrossRef]
- Bui, D.; Brown, H.; Harris, R.; Oren, E. Serologic evidence for fecal-oral transmission of Helicobacter pylori. Am. J. Trop. Med. Hyg. 2016, 94, 82–88. [Google Scholar] [CrossRef]
- Portal-Celhay, C.; Perez-Perez, G.I. Immune responses to Helicobacter pylori colonization: Mechanisms and clinical outcomes. Clin. Sci. 2006, 110, 305–314. [Google Scholar] [CrossRef]
- Ernst, P.B.; Gold, B.D. Helicobacter pylori in childhood: New insights into the immunopathogenesis of gastric disease and implications for managing infection in children. J. Pediatr. Gastroenterol. Nutr. 1999, 28, 462–473. [Google Scholar] [CrossRef]
- Peek, R.M.; Blaser, M.J. Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat. Rev. Cancer 2002, 2, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Boyle, P.; Levin, B. World Cancer Report 2008; International Agency for Research on Cancer (IARC): Lyon, France, 2008. [Google Scholar]
- Figura, N.; Tabaqchali, S. Bacterial pathogenic factors. Curr. Opin. Gastroenterol. 1996, 12, 11–15. [Google Scholar] [CrossRef]
- Con, S.A.; Takeuchi, H.; Con-Chin, G.R.; Con-Chin, V.G.; Yasuda, N.; Con-Wong, R. Role of bacterial and genetic factors in gastric cancer in Costa Rica. World J. Gastroenterol. 2009, 15, 211–228. [Google Scholar] [CrossRef] [PubMed]
- Kusters, J.G.; Van Vliet, A.H.M.; Kuipers, E.J. Pathogenesis of Helicobacter pylori infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef]
- Kao, C.Y.; Sheu, B.S.; Wu, J.J. Helicobacter pylori infection: An overview of bacterial virulence factors and pathogenesis. Biomed. J. 2016, 39, 14–23. [Google Scholar] [CrossRef]
- Argent, R.H.; Thomas, R.J.; Letley, D.P.; Rittig, M.G.; Hardie, K.R.; Atherton, J.C. Functional association between the Helicobacter pylori virulence factors vacA and cagA. J. Med. Microbiol. 2008, 57, 145–150. [Google Scholar] [CrossRef]
- Jones, K.R.; Joo, Y.M.; Jang, S.; Yoo, Y.-J.; Lee, H.S.; Chung, I.-S.; Olsen, C.H.; Whitmire, J.M.; Merrell, D.S.; Cha, J.-H. Polymorphism in the cagA EPIYA motif impacts development of gastric cancer. J. Clin. Microbiol. 2009, 47, 959–968. [Google Scholar] [CrossRef]
- Covacci, A.; Censini, S.; Bugnoli, M.; Petracca, R.; Burroni, D.; Macchia, G.; Massone, A.; Papini, E.; Xiang, Z.; Figura, N. Molecular characterization of the 128-kDa immunodominant antigen of Helicobacter pylori associated with cytotoxicity and duodenal ulcer. Proc. Natl. Acad. Sci. USA 1993, 90, 5791–5795. [Google Scholar] [CrossRef]
- Backert, S.; Selbach, M. Role of type IV secretion in Helicobacter pylori pathogenesis. Cell. Microbiol. 2008, 10, 1573–1581. [Google Scholar] [CrossRef]
- Blaser, M.J.; Perez, G.P.; Kleanthous, H.; Cover, T.; Peek, R.M.; Chyou, P.H.; Stemmermann, G.N.; Nomura, A. Infection with Helicobacter pylori strains possessing cagA is associated with an increased risk of developing adenocarcinoma of the stomach. Cancer Res. 1995, 55, 2111–2115. [Google Scholar]
- Acosta, N.; Quiroga, A.; Delgado, P.; Bravo, M.M.; Jaramillo, C. Helicobacter pylori cagA protein polymorphisms and their lack of association with pathogenesis. World. J. Gastroenterol. 2010, 16, 3936–3943. [Google Scholar] [CrossRef] [PubMed]
- Gwack, J.; Shin, A.; Kim, C.-S.; Ko, K.-P.; Kim, Y.; Jun, J.K.; Bae, J.; Park, S.K.; Hong, Y.-C.; Kang, D.; et al. CagA-producing Helicobacter pylori and increased risk of gastric cancer: A nested case-control study in Korea. Br. J. Cancer. 2006, 95, 639–641. [Google Scholar] [CrossRef] [PubMed]
- Torres, J.; Lopez, L.; Lazcano, E.; Camorlinga, M.; Flores, L.; Muñoz, O. Trends in Helicobacter pylori infection and gastric cancer in Mexico. Cancer Epidemiol. Biomark. Prev. 2005, 14, 1874–1877. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Murata-Kamiya, N. Pathophysiological functions of the CagA oncoprotein during infection by Helicobacter pylori. Microbes Infect. 2011, 13, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Miernyk, K.; Morris, J.; Bruden, D.; McMahon, B.; Hurlburt, D.; Sacco, F.; Parkinson, A.; Hennessy, T.; Bruce, M. Characterization of Helicobacter pylori cagA and vacA genotypes among Alaskans and their correlation with clinical disease. J. Clin. Microbiol. 2011, 49, 3114–3121. [Google Scholar] [CrossRef] [PubMed]
- Higashi, H.; Tsutsumi, R.; Fujita, A.; Yamazaki, S.; Asaka, M.; Azuma, T.; Hatakeyama, M. Biological activity of the Helicobacter pylori virulence factor CagA is determined by variation in the tyrosine phosphorylation sites. Proc. Natl. Acad. Sci. USA 2002, 99, 14428–14433. [Google Scholar] [CrossRef]
- Rizzato, C.; Torres, J.; Plummer, M.; Munoz, N.; Franceschi, S.; Camorlinga-Ponce, M.; Fuentes-Panana, E.M.; Canzian, F.; Kato, I. Variations in Helicobacter pylori cytotoxin-associated genes and their influence in progression to gastric cancer: Implications for prevention. PLoS ONE 2012, 7, e29605. [Google Scholar] [CrossRef]
- Panayotopoulou, E.G.; Sgouras, D.N.; Papadakos, K.; Kalliaropoulos, A.; Papatheodoridis, G.; Mentis, A.F.; Archimandritis, A.J. Strategy to characterize the number and type of repeating EPIYA phosphorylation motifs in the carboxyl terminus of cagA protein in Helicobacter pylori clinical isolates. J. Clin. Microbiol. 2007, 45, 488–495. [Google Scholar] [CrossRef]
- Quiroga, A.J.; Huertas, A.; Combita, A.L.; Bravo, M.M. Variation in the number of EPIYA-C repeats in cagA protein from Colombian Helicobacter pylori strains and its ability middle to induce hummingbird phenotype in gastric epithelial cells. Biomedica 2010, 30, 251–258. [Google Scholar] [CrossRef]
- Batista, S.A.; Rocha, G.A.; Rocha, A.M.; Saraiva, I.E.; Cabral, M.M.; Oliveira, R.C.; Queiroz, D.M. Higher number of Helicobacter pylori cagA EPIYA C phosphorylation sites increases the risk of gastric cancer, but not duodenal ulcer. BMC Microbiol. 2011, 11, 61. [Google Scholar] [CrossRef]
- Basso, D.; Zambon, C.; Letley, D.P.; Stranges, A.; Marchet, A.; Rhead, J.L.; Schiavon, S.; Guariso, G.; Ceroti, M.; Nitti, D.; et al. Clinical Relevance of Helicobacter pylori cagA and vacA Gene Polymorphisms. Gastroenterology 2008, 135, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Atherton, J.C.; Cao, P.; Peek, R.M.; Tummuru, M.K.R.; Blaser, M.J.; Cover, T.L. Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori. Association of specific vacA types with cytotoxin production and peptic ulceration. J. Biol. Chem. 1995, 270, 17771–17777. [Google Scholar] [CrossRef] [PubMed]
- Gangwer, K.A.; Shaffer, C.L.; Suerbaum, S.; Lacy, D.B.; Cover, T.L.; Bordenstein, S.R. Molecular evolution of the Helicobacter pylori vacuolating toxin gene vacA. J. Bacteriol. 2010, 192, 6126–6135. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.C.; De Corvelo Oliveira, T.C.; Demachki, S.; De Araújo, M.T.F.; Assumpcão, M.B.; Vilar, S.C.A.J.; Freitas, F.B.; Barbosa, H.P.M.; Fecury, A.A.; Amaral, R.K.C.D.; et al. Clinical and pathological importance of vacA allele heterogeneity and cagA status in peptic ulcer disease in patients from North Brazil. Mem. Inst. Oswaldo Cruz. 2005, 100, 875–881. [Google Scholar] [CrossRef]
- Chang, Y.H.; Wang, L.; Lee, M.S.; Cheng, C.W.; Wu, C.Y.; Shiau, M.Y. Genotypic characterization of Helicobacter pylori cagA and vacA from biopsy specimens of patients with gastroduodenal diseases. Mt. Sinai J. Med. 2006, 73, 622–626. [Google Scholar]
- Arévalo-Galvis, A.; Trespalacios-Rangel, A.A.; Otero, W.; Mercado-Reyes, M.M.; Poutou-Piñales, R.A. Prevalence of cagA, vacA, and iceA genes in H. pylori strains isolated from Colombian patients with functional dispepsia. Pol. J. Microbiol. 2012, 61, 33–40. [Google Scholar] [CrossRef]
- Parkinson, A.J.; Bruce, M.G.; Zulz, T. International circumpolar international surveillance, an Arctic network for surveillance of infectious diseases. Emerg. Infect. Dis. 2008, 14, 18–24. [Google Scholar] [CrossRef]
- Nolen, L.D.; Bruden, D.; Miernyk, K.; McMahon, B.J.; Sacco, F.; Varner, W.; Mezzetti, T.; Hurlburt, D.; Tiesinga, J.; Bruce, M.G. H. pylori-associated pathologic findings among Alaska native patients. Int. J. Circumpolar Health 2018, 77, 1510715. [Google Scholar] [CrossRef]
- Stancioiu, F.; Ahmed, S.H. Helicobacter pylori: Findings in a Native American Population. IHS Prim. Care Provid. 2005, 30, 59–63. [Google Scholar]
- Kelly, J.J.; Lanier, A.P.; Schade, T.; Brantley, J.; Starkey, B.M. Cancer disparities among Alaska native people, 1970–2011. Prev. Chronic Dis. 2014, 11, E221. [Google Scholar] [CrossRef]
- Tveit, A.H.; Bruce, M.G.; Bruden, D.L.; Morris, J.; Reasonover, A.; Hurlburt, D.A.; Hennessy, T.W.; McMahon, B. Alaska sentinel surveillance study of Helicobacter pylori isolates from Alaska native persons from 2000 to 2008. J. Clin. Microbiol. 2011, 49, 3638–3643. [Google Scholar] [CrossRef] [PubMed]
- Navajo Epidemiology Report. Cancer Among the Navajo, 2005–2013. Available online: https://www.nec.navajo-nsn.gov/Epi-Reports (accessed on 15 June 2021).
- Sacco, F.; Bruce, M.G.; McMahon, B.J.; Bruden, D. A prospective evaluation of 200 upper endoscopies performed in Alaska Native persons. Int. J. Circumpolar Health 2007, 66, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, A.J.; Gold, B.D.; Bulkow, L.; Wainwright, R.B.; Swaminathan, B.; Khanna, B.; Petersen, K.M.; Fitzgerald, M.A. High prevalence of Helicobacter pylori in the Alaska Native population and association with low serum ferritin levels in young adults. Clin. Diagn. Lab. Immunol. 2000, 7, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Keck, J.W.; Miernyk, K.M.; Bulkow, L.R.; Kelly, J.J.; McMahon, B.J.; Sacco, F.; Hennessy, T.W.; Bruce, M.G. Helicobacter pylori infection and markers of gastric cancer risk in Alaska Native persons: A retrospective case-control study. Can. J. Gastroenterol. Hepatol. 2014, 28, 305–310. [Google Scholar] [CrossRef]
- McMahon, B.J.; Bruce, M.G.; Koch, A.; Goodman, K.J.; Tsukanov, V.; Mulvad, G.; Borresen, M.L.; Sacco, F.; Barrett, D.C.; Westby, S.; et al. The diagnosis and treatment of Helicobacter pylori infection in Arctic regions with a high prevalence of infection: Expert Commentary. Epidemiol. Infect. 2016, 144, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Miernyk, K.M.; Bruden, D.; Rudolph, K.M.; Hurlburt, D.A.; Sacco, F.; McMahon, B.J.; Bruce, M.G. Presence of cagPAI genes and characterization of vacA s, i and m regions in Helicobacter pylori isolated from Alaskans and their association with clinical pathologies. J. Med. Microbiol. 2020, 69, 218–227. [Google Scholar] [CrossRef]
- Atrisco-Morales, J.; Martínez-Santos, V.I.; Román-Román, A.; Alarcón-Millán, J.; De Sampedro-Reyes, J.; Carmen, I.C.-D.; Martínez-Carrillo, D.N.; Fernández-Tilapa, G. vacA s1m1 genotype and cagA EPIYA-ABC pattern are predominant among Helicobacter pylori strains isolated from Mexican patients with chronic gastritis. J. Med. Microbiol. 2018, 67, 314–324. [Google Scholar] [CrossRef]
- Román-Román, A.; Martínez-Carrillo, D.N.; Atrisco-Morales, J.; Azúcar-Heziquio, J.C.; Cuevas-Caballero, A.S.; Castañón-Sánchez, C.A.; Reyes-Ríos, R.; Betancourt-Linares, R.; Reyes-Navarrete, S.; Carmen, I.C.-D.; et al. Helicobacter pylori vacA s1m1 genotype but not cagA or babA2 increase the risk of ulcer and gastric cancer in patients from Southern Mexico. Gut Pathog. 2017, 9, 1–12. [Google Scholar] [CrossRef]
- Engstrand, L.; Nguyen, A.M.; Graham, D.Y.; el Zaatari, F.A. Reverse transcription and polymerase chain reaction amplification of rRNA for detection of Helicobacter species. J. Clin. Microbiol. 1992, 30, 2295–2301. [Google Scholar] [CrossRef]
- Toracchio, S.; Caruso, R.A.; Perconti, S.; Rigoli, L.; Betri, E.; Neri, M.; Verginelli, F.; Mariani-Costantini, R. Evolutionarily-Related Helicobacter pylori Genotypes and Gastric Intraepithelial Neoplasia in a High-Risk Area of Northern Italy. Microorganisms 2020, 8, 324. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Kodama, T.; Kashima, K.; Graham, D.Y.; Sepulveda, A.R. Variants of the 3′ region of the cagA gene in Helicobacter pylori isolates from patients with different H. pylori-associated diseases. J. Clin. Microbiol. 1998, 36, 2258–2263. [Google Scholar] [CrossRef] [PubMed]
- Rudi, J.; Kolb, C.; Maiwald, M.; Kuck, D.; Sieg, A.; Galle, P.R.; Stremmel, W. Diversity of Helicobacter pylori vacA and cagA genes and relationship to vacA and cagA protein expression, cytotoxin production, and associated diseases. J. Clin. Microbiol. 1998, 36, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Argent, R.H.; Zhang, Y.; Atherton, J.C. Simple method for determination of the number of Helicobacter pylori cagA variable-region EPIYA tyrosine phosphorylation motifs by PCR. J. Clin. Microbiol. 2005, 43, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Melkonian, S.C.; Pete, D.; Jim, M.A.; Haverkamp, D.; Wiggins, C.L.; Bruce, M.G.; White, M.C. Gastric Cancer Among American Indian and Alaska Native Populations in the United States, 2005–2016. Am. J. Gastroenterol. 2020, 115, 1989–1997. [Google Scholar] [CrossRef] [PubMed]
- Wiggins, C.L.; Perdue, D.G.; Henderson, J.A.; Bruce, M.G.; Lanier, A.P.; Kelley, J.J.; Seals, B.F.; Espey, D.K. Gastric cancer among American Indians and Alaska natives in the USA, 1999–2004. Cancer 2008, 113, 1225–1233. [Google Scholar] [CrossRef][Green Version]
- Goodman, K.J.; Jacobson, K.; Veldhuyzen van Zanten, S. Helicobacter pylori infection in Canadian and related Arctic Aboriginal populations. Can. J. Gastroenterol. 2008, 22, 289–295. [Google Scholar] [CrossRef]
- Yang, P.; Zhou, Y.; Chen, B.; Wan, H.-W.; Jia, G.-Q.; Bai, H.-L.; Wu, X.-T. Overweight, obesity and gastric cancer risk: Results from a meta-analysis of cohort studies. Eur. J. Cancer 2009, 45, 2867–2873. [Google Scholar] [CrossRef]
- Yusefi, A.R.; Bagheri Lankarani, K.; Bastani, P.; Radinmanesh, M.; Kavosi, Z. Risk Factors for Gastric Cancer: A Systematic Review. Asian Pac. J. Cancer Prev. 2018, 19, 591–603. [Google Scholar] [CrossRef]
- Carmack, A.M.; Schade, T.L.; Sallison, I.; Provost, E.M.; Kelly, J.J. Cancer in Alaska Native People: 1969–2013 the 45-Year Report; Alaska Native Tumor Registry, Alaska Native Epidemiology Center, Alaska Native Tribal Health Consortium: Anchorage, AK, USA, 2015. [Google Scholar]
- Parsonnet, J.; Friedman, G.D.; Orentreich, N.; Vogelman, H. Risk for gastric cancer in people with cagA positive or cagA negative Helicobacter pylori infection. Gut 1997, 40, 297–301. [Google Scholar] [CrossRef]
- Sahara, S.; Sugimoto, M.; Vilaichone, R.-K.; Mahachai, V.; Miyajima, H.; Furuta, T.; Yamaoka, Y. Role of Helicobacter pylori cagA EPIYA motif and vacA genotypes for the development of gastrointestinal diseases in Southeast Asian countries: A meta-analysis. BMC. Infect. Dis. 2012, 12, 223–230. [Google Scholar] [CrossRef]
- Sicinschi, L.A.; Correa, P.; Peek, R.M.; Camargo, M.; Piazuelo, M.; Romero-Gallo, J.; Hobbs, S.; Krishna, U.; Delgado, A.; Mera, R.; et al. CagA C-terminal variations in Helicobacter pylori strains from Colombian patients with gastric precancerous lesions. Clin. Microbiol. Infect. 2010, 16, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Beltrán-Anaya, F.O.; Poblete, T.M.; Román-Román, A.; Reyes, S.; De Sampedro, J.; Peralta-Zaragoza, O.; Rodríguez, M.; Del Moral-Hernández, O.; Illades-Aguiar, B.; Fernández-Tilapa, G. The EPIYA-ABCC motif pattern in cagA of Helicobacter pylori is associated with peptic ulcer and gastric cancer in Mexican population. BMC Gastroenterol. 2014, 14, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Leon, A.; Atherton, J.C.; Argent, R.H.; Puente, J.L.; Torres, J. Heterogeneity in the activity of Mexican Helicobacter pylori strains in gastric epithelial cells and its association with diversity in the cagA gene. Infect. Immun. 2007, 75, 3445–3454. [Google Scholar] [CrossRef] [PubMed]
- Devi, S.M.; Ahmed, I.; Khan, A.A.; Rahman, S.A.; Alvi, A.; Sechi, L.A.; Ahmed, N. Genomes of Helicobacter pylori from native Peruvians suggest admixture of ancestral and modern lineages and reveal a western type cag-pathogenicity island. BMC Genom. 2006, 7, 191. [Google Scholar] [CrossRef] [PubMed]
- Camorlinga-Ponce, M.; Perez-Perez, G.; González-Valencia, G.; Mendoza, I.; Peñaloza-Espinosa, R.; Ramos, I.; Kersulyte, D.; Reyes-Leon, A.; Romo, C.; Granados, J.; et al. Helicobacter pylori genotyping from american indigenous groups shows novel amerindian vacA and cagA alleles and Asian, African and European admixture. PLoS ONE 2011, 6, e27212. [Google Scholar] [CrossRef]
- Sugimoto, M.; Yamaoka, Y. The association of vacA genotype and Helicobacter pylori-related disease in Latin American and African populations. Clin. Microbiol. Infect. 2009, 15, 835–842. [Google Scholar] [CrossRef]
- McClain, M.S.; Beckett, A.C.; Cover, T.L. Helicobacter pylori vacuolating toxin and gastric cancer. Toxins 2017, 9, 316. [Google Scholar] [CrossRef]
- Kersulyte, D.; Kalia, A.; Gilman, R.H.; Mendez, M.; Herrera, P.; Cabrera, L.; Velapatino, B.; Balqui, J.; De La Vega, F.P.P.; Ulloa, C.A.R.; et al. Helicobacter pylori from Peruvian Amerindians: Traces of human migrations in strains from remote Amazon, and genome sequence of an Amerind strain. PLoS ONE 2010, 5, e15076. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Primer | Use | Sequence | Target | Reference |
|---|---|---|---|---|
| 16S1/16S2 | Amplification | 5′ GCTAAGAGATCAGCCTATGTCC 3′ 5′ TGGCAATCAGCGTCAGGTAATG 3′ | 16S rRNA | [51] |
| VA1-F/VA1-R | Amplification and sequencing | 5′ ATGGAAATACAACAAACACAC 3′ 5′ CTGCTTGAATGCGCCAAAC 3′ | vacA s | [33,50] |
| mF1/mR1 | Amplification and sequencing | 5′ ACCGCTCATBAAGATYAAYARCGCTC 3′ 5′ GCTAGGCGCTCTTTGAATTGC 3′ | vacA m | [52] |
| glmM-F/glmM-R | Amplification | 5′ TAACCGAAGACATGCGCTG 3′ 5′ CATGAAAGATTTCTTCAATCAATCGCT 3′ | glmM | [52] |
| cagA-F1/ cagA-R1 | Amplification | 5′ GATAACAGGCAAGCTTTTTGAGG 3′ 5′ CTGCAAAAGAATGTTTGGCAG 3′ | 5′ cagA | [53] |
| CAGTF/ CAGTR | Amplification | 5′ ACCCTAGTCGGTAATGGGTTA 3 5′ GTAATTGTCTAGTTTCGC 3′ | 3′ cagA | [53] |
| cagA-2/ cagA-4 | Sequencing | 5′ GGAACCCTAGTCGGTAATG 3′ 5′ ATCTTTGAGCTTGTCTATCG 3′ | 3′ cagA | [54,55] |
| Total | Positive | Negative | p Value | |
|---|---|---|---|---|
| Total | 96 | 22 | 74 | |
| Age (mean) | 55.6 | 54.0 | 56.1 | 0.57 * |
| Gender | 0.08 # | |||
| Male | 25 | 10 | 15 | |
| Female | 71 | 12 | 59 | |
| Reason for Endoscopy | ||||
| Epigastric Pain | 17 | 5 | 12 | 0.153 § |
| Abdominal Pain | 13 | 4 | 9 | |
| GERD | 10 | 1 | 9 | |
| Bloating | 10 | 2 | 8 | |
| Right Upper Quadrant Pain | 6 | 2 | 4 | |
| Nausea | 5 | 1 | 4 | |
| Left Upper Quadrant Pain | 4 | 1 | 3 | |
| Gastritis | 3 | 1 | 2 | |
| GI Bleed | 2 | 2 | 0 | |
| Constipation | 2 | 0 | 2 | |
| Gastric Finding | ||||
| Normal | 14 | 2 | 12 | 0.21 § |
| Gastritis | 60 | 13 | 47 | |
| Gastric Atrophy & Fundal Gastritis | 4 | 3 | 1 | |
| Erosive Gastropathy | 4 | 3 | 1 | |
| Gastritis & Intestinal Metaplasia w/o Dysplasia | 7 | 2 | 5 |
| cagA | vacA Genotype | ||||||
|---|---|---|---|---|---|---|---|
| Gastric Disease | EPIYA | s1a,s1b | s1a | s1b | s2 | m1 | m2 |
| Normal | |||||||
| 0 | 0 | 0 | 1 | 0 | 2 | ||
| Gastritis | |||||||
| AB (1) | 0 | 0 | 1 | 1 | 1 | 1 | |
| ABC (7) | 1 | 1 | 9 | 1 | 8 | 2 | |
| Gastritis & ulcer | |||||||
| Erosive gastropathy | |||||||
| ABC (2) | 1 | 1 | 1 | 0 | 2 | 0 | |
| Gastric atrophy & Fundal gastritis | |||||||
| ABC (2) | 0 | 1 | 0 | 1 | 0 | ||
| ABCC (1) | 0 | 1 | 0 | 0 | 1 | 0 | |
| Gastric & Intestinal Metaplasia | |||||||
| ABCC (1) | 0 | 0 | 1 | 0 | 1 | 0 | |
| Gastritis & Intestinal metaplasia without dysplasia | |||||||
| ABCC (1) | 0 | 0 | 1 | 0 | 1 | 0 | |
| TOTAL | (15/20) | 2 | 3 | 14 | 3 | 15 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monroy, F.P.; Brown, H.E.; Sanderson, P.R.; Jarrin, G.; Mbegbu, M.; Kyman, S.; Harris, R.B. Helicobacter pylori in Native Americans in Northern Arizona. Diseases 2022, 10, 19. https://doi.org/10.3390/diseases10020019
Monroy FP, Brown HE, Sanderson PR, Jarrin G, Mbegbu M, Kyman S, Harris RB. Helicobacter pylori in Native Americans in Northern Arizona. Diseases. 2022; 10(2):19. https://doi.org/10.3390/diseases10020019
Chicago/Turabian StyleMonroy, Fernando P., Heidi E. Brown, Priscilla R. Sanderson, Gregory Jarrin, Mimi Mbegbu, Shari Kyman, and Robin B. Harris. 2022. "Helicobacter pylori in Native Americans in Northern Arizona" Diseases 10, no. 2: 19. https://doi.org/10.3390/diseases10020019
APA StyleMonroy, F. P., Brown, H. E., Sanderson, P. R., Jarrin, G., Mbegbu, M., Kyman, S., & Harris, R. B. (2022). Helicobacter pylori in Native Americans in Northern Arizona. Diseases, 10(2), 19. https://doi.org/10.3390/diseases10020019