Abstract
Topical sun protective cosmetics (sunscreens, pre- and post-sun) have been intensively developed and produced to protect human skin against solar irradiation-associated damages/pathologies. Unfortunately, routine cosmetics for sun protection containing synthetic organic and/or physical sunscreens could exert adverse effects towards human organisms and bring undesirable ecological changes. Terrestrial and marine plant species, being exposed to sun light for hundreds of millions of years, have evolved two pro-survival strategies: effective protection against/adaptation to its deleterious effects and the use of solar energy for photosynthesis/photo-biochemical reactions. Secondary plant metabolites (SPM) are primary sensors of solar energy and mediators of its use (photo-sensitisers) or neutralisation (photo-protectors). A similar double photo-protective/photo-sensitising system is built in within human skin. Modern development of toxicologically/ecologically safe yet effective sun-protective cosmetics attempts to pre-select photo-stable and non-phototoxic SPMs that provide broad UVA + UVB sunscreen, free radical scavenging and direct antioxidant defence, endogenous antioxidant rescue, induction of antioxidant enzymes (indirect antioxidant defence), and normalisation of metabolic and immune responses to UVA + UVB. Proper formulation of sun protective cosmetics should assure targeted delivery of photo-active SPMs to definite skin layers to invigorate the built in photo-chemical skin barrier.
1. Introduction
Solar UV radiation interacts with an extremely complex networks of cutaneous compartments dedicated to mechanical, photo-chemical, redox, metabolic, and immune defence of the skin and underlying organs. As a first step, UV induces photo-chemical reactions in the outmost superficial skin surface lipids. Then, UV photo-chemically modifies non-viable keratinocytes and intercellular lipids of the stratum corneum. Finally, it reaches directly or by means of photo-chemical mediators the viable layers of epidermis and the underlying dermal compartments [1]. Upon exposure to UV radiation or to photo-chemical mediators, skin keratinocytes mount characteristic inflammatory, metabolic, and proliferative responses in an attempt for self-protection from and adaptation to UV-induced damage [2]. Effective and safe skin protection against damages induced by solar irradiation remains among the unsolved problems of modern dermatology and cosmetology [3]. Currently, plants and marine organisms are considered valuable sources of anti-photo-ageing and photo-protective compounds for the development of cosmeceutical and topical pharmaceutical products [4,5,6,7]. Here, we will address this issue comparing natural evolution-derived photo-active systems/mechanisms in human skin, higher plants, and lower marine organisms.
2. Photo-Chemical Barrier in Plants versus Human Skin
2.1. Photo-Active Secondary Metabolites in Terrestrial and Marine Plants: Biosynthesis and Physiological Effects
Terrestrial and marine plants synthesise primary metabolites essential for their growth, division, and propagation (proteins and aminoacids, lipids and fatty acids, poly- and oligosaccharides). They account for more than 90–95% of a dry weight of higher and marine plants [8].
Terrestrial plants and lower eucariots, such as marine micro/macroalgae, sea weeds, coral gorgonians, etc., have evolved biosynthesis of secondary metabolites (SPMs), which do not participate directly in their growth, division, and propagation but are essential for defence and adaptation to continuously changing environmental conditions (to co-called biotic and abiotic stresses) [4,9,10,11]. For example, secondary metabolism in plants/marine organisms starts upon exposure to UV, X-rays or ultrasonic irradiation, extreme temperatures (cold or heat), heavy metals, salts, and organic toxins, mechanical damage, etc. [7]. It is also initiated by bacterial, viral or parasite infections, or by damage produced by herbivores and insects. In general, SPMs are known as protective, adaptive, connecting, and regulatory (hormone-like) substances. It is believed that SPMs play the role of the primitive “immune system” and “detoxification network” in plants and lower marine organisms (reviewed in [12]). A wealth of known so far SPMs (more than 5000 substances) are commonly divided in accordance with their chemical structure into three major classes:
- -
- phenylpropanoids (synonyms, ethylpropanoids) and their derivatives, such as simple polyphenolics (acids, alcohols, and aldehydes), aromatic/poly-aromatic polyphenols (flavonoids, stilbenes, curcuminoids, coumarins, etc.), and glycosides (glycoside moieties, are, mainly, rhamnose, mannose, rutinose, etc.). All these SPMs contain exclusively carbon, hydrogen, and oxygen atoms and multiple hydroxyl groups;
- -
- terpenoids containing long, mainly, unsaturated C-H chain and nucleus of non aromatic C-H cycle;
- -
- nitrogen-containing heterocycles, such as alkaloids, purines, pyrimidines, porphyrins, chlorophylls, flavins, etc.
A great majority of SPMs are synthesised upon exposure to solar UV irradiation [5,9,13,14].
Protective properties of secondary plant metabolites against solar light of broad spectral range from UV irradiation to visible and infrared light [10,14] are greatly similar for plant and mammalian cells. This possibility of universal defence is provided by peculiar chemical structure (chromophores) that absorbs solar light energy and dissipates it either in the form of thermal or fluorescent/phosphorescent light energy (sunscreen properties) or convert it into energy for chemical reactions (photo-sensitising properties).
For example, polyphenols like phenylpropanoids and their glycosides as well as bioflavonoids are synthesised in plant cells through phenylpropanoid pathway immediately upon exposure to UV [8,9,14,15]. Having molecular structure consisting of condensed aromatic 5- and 6-carbon rings with multiple OH-groups, they effectively absorb UVA + UVB (sun screen properties) (Figure 1) without promotion of further photo-chemical reactions. They have been considered safer than classical synthetic sunscreens [16], chemical structures of which are also aromatic polyphenols [17,18]. From a kinetic chemistry point of view, SPM phenylpropanoids and their off-spring polyphenols usually have rather high activation energy; therefore, they need UV irradiation to acquire an excited state. Due to their highly conjugated aromatic moieties with stable “π” configuration, their excited state lives longer and energy dissipates within the space of this molecule. Their aromatic rings are mainly in perpendicular positions that favour electron “entrap” within the molecule (Figure 1). Moreover, the presence of glycosyl moieties in SPMs provides increased photo-stability, hence many of the glycosylated metabolites are not susceptible to UVA destruction [19].
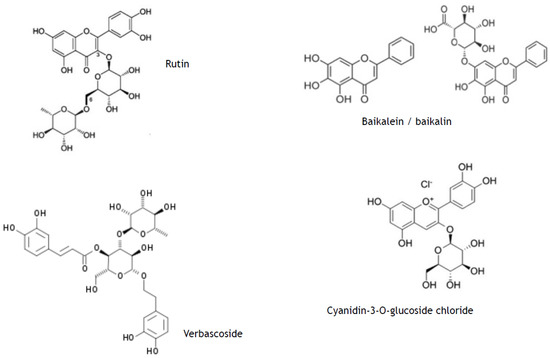
Figure 1.
Photo-protective polyphenolic secondary plant metabolites. Examples of plant-derived secondary polyphenolic metabolites (phenylpropanoid glycoside verbascoside; flavonoids (glycosylated and aglycon baicalein/baicalin), rutin, silibinin).
Other SPMs (stilbenes, catechins, flavonoids, and terpenoids) preferably protect plant molecules and structures against damaging effects of reactive oxygen species (superoxide anion-radicals, hydroxyl radicals, peroxides, and singlet oxygen) formed upon UV reaction with organic matter in the presence of molecular oxygen (type II photo-reactions) [5,13] (Figure 2).
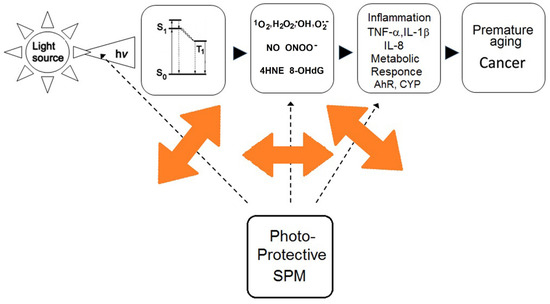
Figure 2.
Modes of photo-protection by polyphenolic secondary plant metabolites. Physical photo-protection by interaction with UV light quantum (sunscreen), free radical scavenging, antioxidant, and singlet oxygen quenching (photo-chemical protection), and inhibition of inflammatory and metabolic skin responses to UV light (biological protection).
In contrast, the majority of nitrogen-containing heterocycles (Figure 3) and terpenoids are photo-sensitisers, which use UV energy, visible or infra-red solar irradiation to promote biologically important free radical-driven photo-chemical reactions (Figure 4), such as photosynthesis or terpenoid- and polyphenol-containing polymer synthesis [9]. Of note that polyphenols also containing aldehyde group(s) could exert properties of photo-sensitisers due to decreased energy of photo-excitation [20,21] (Figure 5).
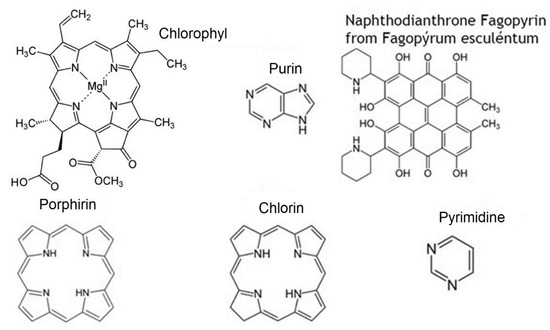
Figure 3.
Nitrogen-containing heterocycle secondary plant metabolites possess photo-sensitising properties. Examples of biologically important secondary plant metabolites (nitrogen-containing heterocycles) with photo-sensitising action.
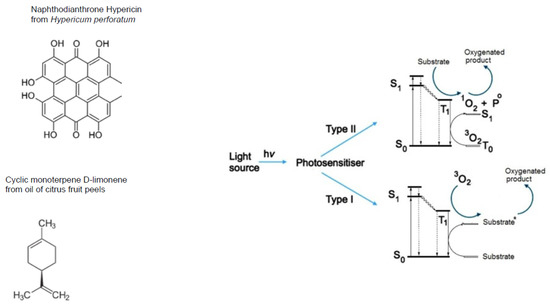
Figure 4.
Excitation of photo-sensitisers. Scheme explaining the process of solar light (UV, visible, infrared) -excitation of secondary plant metabolites followed by photo-chemical reactions.
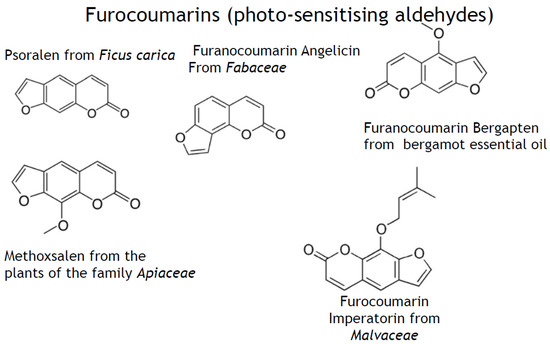
Figure 5.
Polyaromatic secondary plant metabolites with aldehyde group(s) are mainly photo-sensitisers. Examples of polyaromatic aldehyde-containing secondary plant metabolites with prevalently photo-sensitising properties.
When whole botanical extracts are used for topical photo-protection, there is a great risk of adverse photo-sensitising effects of definite plant-derived secondary metabolites that lead to phyto-photo dermatosis or allergic reaction [12].
The UV-protecting systems in plants and lower marine organisms resemble a cutaneous photo-chemical barrier in humans (Figure 6) consisting of photo-screens (pheomelanin, melatonin, and proteins rich of aromatic amino acids) [22,23] and photo-sensitisers (amino acid tryptophan and derivatives, retinoic acid, eumelanin, porphyrins, flavins, haemoglobin, purines, and pyrimidines) [24,25,26,27].
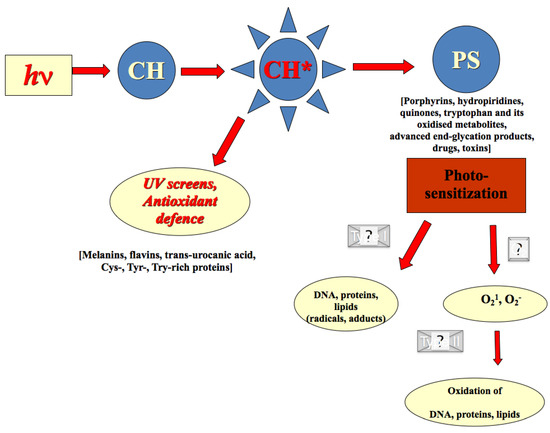
Figure 6.
Endogenous photo-protectors and photo-sensitisers within human skin. Quantum of solar energy (hν) is absorbed by endogenous chromophore (CH), which becomes excited (CH*) and highly reactive either directly with DNA and proteins (type I reaction) or it reacts first with molecular oxygen (type II reaction) converting it in singlet oxygen, thus initiating non-enzymatic oxidation reaction. Examples of endogenous cutaneous photo-protectors and photo-sensitisers are in the brackets.
The endogenous cutaneous barrier efficacy depends on the photo-protector/photo-sensitiser balance that gradually deteriorates in ageing or ailing human skin [25,28].
2.2. Photo-Chemical Barrier in Human Skin
A very peculiar photo-chemical barrier of the human skin aims to detoxify/eliminate solar light-modified low molecular weight substances.
The outmost external photo-chemical barrier encountered by environmental radiations is the skin surface lipids, a protective hydrophobic film occurring more abundantly in the most photo-exposed cutaneous districts [1]. The lipid barrier is a mixture of epidermal lipids, deriving: (a) from the exfoliating stratum corneum, mainly composed of keratinocyte debris phospholipids and their products of hydrolysis, and (b) from lipids, triglycerides, sterols, and lipophilic vitamins (vitamin E and coenzyme Q10) produced by sebaceous glands. The lipids synthesised in sebocytes are rich in highly oxidisable triterpenoid squalene, a highly lipophilic molecule presents uniquely on human skin, which is regarded not only as a water-insulating and skin-smoothening factor, but also most importantly as a sacrificial antioxidant [29,30]. Under UV exposure, following the rapid degradation of the lipophilic antioxidants, squalene remains a main guardian protecting precious epidermal unsaturated phospholipid moieties from the UV-induced free radical-driven oxidative damage. The short-lived, hydrophilic, low-molecular weight oxidative by-products of squalene oxidation, able to rapidly diffuse to viable epidermal and dermal layers, have been proven to be an early signal triggering the adaptive skin immune response to UV-irradiation [31] as well as a cutaneous metabolic response to solar-simulating UV [32]. Although squalene has long been considered a double edge sword: if produced by sebaceous glands or applied to the skin in excessive amounts, squalene could give an uncontrolled rise of toxic aldehydes and epoxides, which are not sufficiently metabolised by the built in skin antioxidant systems. In this case, squalene may be responsible for the pro-inflammatory, pro-ageing, and tumour-promoting cutaneous photo-oxidative damage (reviewed in [1]). The rate of squalene degradation has been therefore proposed as a feasible parameter to measure the efficacy of sun-protecting formulations [33]. A role for selected plant-derived polyphenols in the modulation of squalene oxidation under UV stimuli is anticipated [7,32].
Several photo-chemical mediators of UV signalling to skin cells, such as products of tryptophan photo-oxidation [34], 4-hydroxy-2-nonenal, the final product of membrane-bound arachidonic acid oxidation [35], 9-hydroxyoctadecadienoic acid, the product of photo-oxidation of linoleum acid [36], and photo-oxidised squalene [32] have been identified so far.
At underlying cellular skin layers (epidermis and derma), the UV interaction starts from absorption of UV light by endogenous chromophores such as trans-urocanic acid, melanins, porphyrins, flavins, quinones, tryptophan and its photo-oxidised metabolites, such as N′-formylkinurenine and 6-formylindolo[3,2-b]carbazole, hydropirydines, and advanced glycation end-products [22,23,24,25]. This interaction reduces UV penetration, hence the chromophores play a protective role of sacrificial sunscreens. Recently, the utmost importance of epidermal and dermal UV chromophore rich proteins including the late cornified envelope proteins, keratin associated proteins, and elastic fibre-associated components as endogenous sacrificial sunscreens have been described [37].
2.3. Redox Barrier of Human Skin
Human skin, being the largest organ in the organism exposed to a hostile environment, secures redox protection from the air- and UV-born free radicals and reactive species (so called redox barrier of the skin). In the viable layers of the epidermis, lipid-soluble antioxidants, mainly alpha-tocopherol, and antioxidant enzymes like catalase (CAT), superoxide dismutases (SOD), glutathione peroxidases (GPx), mainly GPx3, and peroxyredoxins (Px) are abundantly expressed [38,39]. The homeostatic role of mitochondrial manganese SOD (MnSOD) towards keratinocyte response to UV irradiation mediated through the epidermal growth factor receptor (EGFR) has recently been discovered [40].
To effectively counteract insults coming from the environment, the epidermis of normal human skin contains high levels of water and lipid-soluble antioxidants such as glutathione, thioredoxin, vitamin C, uric acid, alpha-tocopherol, squalene, and coenzyme Q10, distributed in a gradient with the highest concentration on the deepest epidermal layers [38,39].
Endogenous intracutaneous melatonin and its metabolites, which are formed in enzymatic and non-enzymatic oxidation reactions, appear to represent the most potent antioxidants against UV irradiation-induced oxidative stress and carcinogenesis due to their hydroxyl radical scavenging properties [41,42]. Melatonin-derived metabolites exerting antioxidant properties (6-hydroxymelatonin, N(1)-acetyl-N(2)-formyl-5-methoxykynuramine, and 5-methoxytryptamine) were detected in three types of normal human skin cells, keratinocytes, melanocytes, and dermal fibroblasts [43]. The extracellular space of dermis contains large amounts of water-soluble antioxidants such as ascorbic acid, uric acid, and glutathione. In addition, several key structural proteins abundantly present in the stratum corneum, hair, and papillary dermis and characterised by the high content of aminoacids-chromophores, namely, cysteine, tryptophan, and tyrosine, appear to have significant impact on cutaneous antioxidant defence protecting skin structures against photo-chemical oxidative stress [37]. As part of an adaptive response, abnormal levels of ROS also induce the expression of ROS-detoxifying enzymes, mainly through a nuclear factor erythroid 2-related factor 2 (Nrf2)-dependent mechanism [44].
2.4. Effects of Secondary Plant Metabolites on Photo-Chemical and Redox Barriers of Human Skin
The transcription factor Nrf2 (nuclear factor-E2-related factor 2) is widely considered a key switch of cell defence against environmental abiotic stresses, such as UV radiation, pollutants, xenobiotics both low molecular weight toxins and drugs, heavy metals, ozone, oxidative stress, etc. [45,46]. Genes regulated by this nuclear factor include detoxifying phase II enzymes, DNA repair, inflammatory signalling, and antioxidant response. Recently, Nrf2-regulated gene expression in response to solar UVA + UVB radiation in cultivated human skin cells has been reported [47]. Due to these findings, Nrf2 has been suggested as a target for modulation of skin chronological and premature photo-damage-associated skin ageing [48,49].
3. Metabolic Barrier of Human Skin
The skin is an excellent biochemical/toxicological barrier against transcutaneous penetration of low molecular weight xenobiotics and drugs. Topically applied SPMs are a spectacular example of xenobiotics to be detoxified and eliminated through activation of skin-located metabolic enzymes. In addition, these enzymes participate in biotransformation followed by elimination of toxic endogenous metabolites, such as hormones, oxidised/photo-oxidised lipids and amino acids.
The biotransformation is initiated by the oxidative phase I enzymes (cytochrome 450-CYPs- superfamily, flavoprotein mono-oxygenase, cycloxygenase, alcohol dehydrogenase, amine oxidases, epoxide hydroxylase, and others). An oxidised toxin is subjected to reductive or conjugative modification by the phase II enzymes (glutathione-S-transferases (GSTs), UDP-glucoronosyl transferases (UGTs), catechol-O-methyl transferases (COMT), N-acetyl transferases (NATs), and sulfo transferases). Expression of xenobiotic metabolising enzymes is regulated by nuclear receptors, transcription factors, and epigenetic mechanisms including microRNAs. It is common knowledge now that phase I enzymatic reactions are less relevant than metabolic transformations provided by phase II enzymes in the skin (reviewed recently in [50]). Of relevance, both Cyp1A1 and Cyp1B1 are highly inducible in human keratinocytes not only by polyaromatic chemicals, including environmental organic toxins, drugs, SPMs, cosmetics, but also by UVB irradiation [34,51,52]. The metabolising enzymes are located mostly in keratinocytes within the epidermis [53], although these enzymes are also expressed in cutaneous immune cells, melanocytes, and fibroblasts [50,54].
Their induction and activity are regulated by the aryl hydrocarbon receptor (AhR), a transcriptional factor shuttling from cytoplasm to nucleus. For a long time, the AhR has been considered a sensor and a primary regulator of the adaptive metabolic response to a wide range of xenobiotics. It also plays a leading role in the metabolic degradation of toxic endobiotics. For example, prostaglandins, lipoxins, and other eicosanoids serve as AhR ligands-activators of its signal transduction pathway [55]. There is also growing evidence that the physiological role of AhR in the skin is not limited to xenobiotic metabolism, but it extends to numerous functions of cutaneous cells such as breakdown of endogenous metabolites, proliferation, cell-to-cell contacts, immune and inflammatory responses, and melanogenesis [56]. Among the endogenous AhR ligands identified so far, there are the products of tryptophan photo-oxidation, squalene photo-oxidation, oestrogen, prostaglandins, bilirubin/biliverdin, and kinurenic acid [32,57,58]. Apart from metabolic enzymes, a number of growth factors, cytokines, chemokines and their receptors are down-stream gene targets for activated AhR [45,59]. AhR is also functionally connected with epidermal growth factor receptor, presumably, through the NF-kappaB-regulated pathway [60], thus influencing the epithelial cell proliferation. AhR can also cross-talk and directly interact with proteins involved in major redox-regulated signalling pathways such as NF-kappaB and various kinases, such as Src, JNK, p38, MAPK and with oestrogen receptors to mediate oestrogen metabolism [61]. Recent studies have unraveled unsuspected physiological roles and novel alternative ligand-specific pathways for this receptor that allowed for hypothesising numerous pharmacological roles of AhR ligands useful for the development of a new generation of photo-protective drugs and cosmetics [62,63].
3.1. Metabolic Responses in Human Skin to Solar Irradiation
The tryptophan photo-oxidation products, firstly formylindolo[3,2-b]carbazole (FICZ), were identified as endogenous ligands of AhR hence mediators of CYP-related metabolic responses to UV in keratinocytes and melanocytes [34]. A recent publication has shown that squalene, a major component of human skin surface lipids, becomes an AhR ligand and, as a consequence, a CYP1A1 and CYP1B1 inducer, following photo-oxidation by solar UV [32]. In this context, peroxidated squalene has been suggested as another endogenous AhR-CYP activator in the skin, and also a potential contributor to pro-inflammatory functions of epidermal keratinocytes amid their response to squalene peroxide with a significant upregulation and de novo synthesis of IL-6 and TNF-alpha [32].
3.2. Effects of Secondary Plant Metabolites on Metabolic Responses to Solar Irradiation
A number of dietary redox active polyphenols (resveratrol, quercetin, curcumin, etc.), indoles, tryptophan metabolites, bilirubin, and oxidised products of lipid metabolism have been suggested as non-toxic ligands-activators or ligands-inhibitors of the AhR expression by competitive and non-competitive pathways [32,56]. Among a number of plant polyphenols used for topical application, exclusively phenylpropanoid verbascoside and flavonoid quercetin were strong inhibitors of UV- or FICZ-upregulated AhR-CYP1A1-CYP1B1 axis in human keratinocytes [63,64], suggesting their potency as topical UV protectors. Several plant-derived polyphenols have been shown to suppress UV-stimulated CYP1A1 and CYP1B1 enzymes, which have been suggested as a positive sign to prevent biotransformation of pro-carcinogens into ultimate carcinogens in the skin [65]. Several other SPMs induced over-expression of these metabolic enzymes that could be described in terms of increased risk of non-melanoma skin cancer development [66,67]. On the other hand, induction of phase I enzymes could accelerate SPMs metabolism in the skin. There are data confirming that AhR receptor activation/inhibition by definite plant polyphenols correlate with the consequent CYP1 behaviour [68]. Resveratrol and its glycoside polydatin augment metabolic and inflammatory responses of human keratinocytes to UVA + UVB radiation or to exogenous FICZ [69], which allowed for issuing a warning about possible risks of resveratrol in sun protective cosmetics.
4. Immune System in Human Skin
The skin has been evolved to assure appropriate multilevel defence of entire human organisms against mechanical damage, foreign biological invaders, high molecular weight and low molecular weight substances, and physical changes in the environment, such as temperature and solar irradiation. Numerous microbial and viral organisms, high molecular weight haptens and UV-modified substances are guarded by the innate skin-resided immune system as well as by the acquired induced-by-stimulus immunity consisting of keratinocytes, endotheliocytes, tissue-located and recruited professional immune cells (lymphocytes, monocytes, granulocytes, dendritic cells, and Langerhans cells). The cellular components communicate by means of cytokines and chemokines to provide acute immune/inflammatory response targeting the invader [22] (Figure 7).
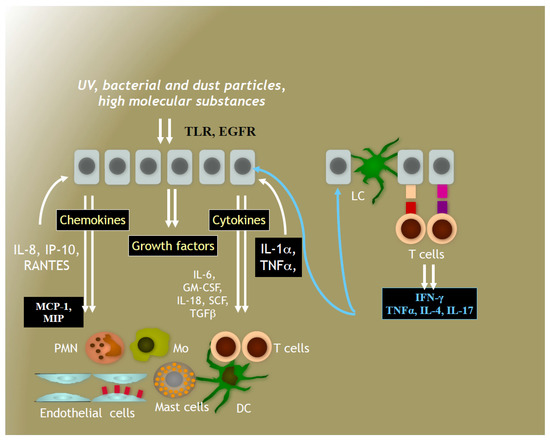
Figure 7.
Immune system in human skin. Skin immune system consists of cells-residents (mainly, keratinocytes, tissue macrophages, dendritic and Langerhans cells) and recruited professional immune cells, such as lymphocytes, granulocytes, and monocytes. Immune cells react to foreign invaders and UV radiation by synthesising de novo a plethora of cytokines and chemokines as well as growth factors. UV-responsive cellular receptors, such as Toll-like (TLR), epidermal growth factor receptor (EGFR), aryl hydrocarbon receptor (AhR) initiate molecular events with the aid of specialised nuclear factors (NF-kappaB and Nrf2), which lead to metabolic and inflammatory response of the skin in order to protect intra-cutaneous homeostasis. Photo-protective secondary plant metabolites could modulate these molecular events, thus enhancing the photo-protection.
4.1. Immune Responses to Solar Irradiation
Immune system of the skin suffers the most upon exposure to UV radiation and/or photo-modified metabolites derived from surface skin lipids, damaged skin cells and extracellular matrix. The finely tuned system of defence against foreigners, such as high molecular weight molecules, dust and bacterial particles, protozoa, fungi, and viruses, becomes confused and its responses are greatly altered either being suppressed (immune deficit) or inadequately enhanced (inflammation). Chronic exposure to solar irradiation inevitably results in the immune deficit of a cutaneous immune system that is commonly regarded as a causative reason for UV-induced carcinogenesis [2,3,28,65]. Immune responses of cutaneous cells could be made evident and quantified by the measurements of cytokines, chemokines, and growth factors’ expression, by the expression of receptors responsive to UV light, such as membrane-bound epidermal growth factor receptor, Toll-like receptors, G2A receptors and many others, or intra-cellular receptors, for example, NF-kappaB and AhR [28,34,64,69]. A majority of skin pathologies and skin ageing is attributed to these alterations in the local immune system. Therefore, abnormally expressed soluble or cell-bound substances and cutaneous immune cells have become targets for the search of pathogenically relevant drugs and photo-protective cosmetics.
4.2. Effects of Secondary Plant Metabolites on Immune Responses to Solar Irradiation
A wealth of publications on the photo-protective effects of plant-derived substances, mainly SPMs, have been done during the last two decades due to growing public and medicinal society concern about severe damage associated with solar and artificial UV irradiation [3,4,5,7,65]. Experimental data suggest that a variety of SPMs, mainly, polyphenols extracted from green tea, grape seed, soy beans, or Curcuma longa root, when taken orally or applied topically, do exert chemo-prevention of skin inflammation (erythema and sunburned lesions) followed by a significant decrease in non-malignant and malignant skin tumours derived from exposure to carcinogenic doses of UV [70,71]. In a clinical study, the soybean isoflavone genistein effectively inhibited UVB-induced erythema when applied onto the skin before UVB irradiation [72]. Phytochemicals containing a plethora of SPMs directly exert their chemo-prevention of properties by diminishing the levels of pro-inflammatory mediators such as free radicals, inorganic and organic peroxides, and aldehydes generated by UV irradiation [8,15]. Oxidation end-products of sebum and keratinocyte cell membrane lipid moieties represent key biochemical mediators of the inflammatory reaction induced by UV irradiation [1,73,74], among which 4-hydroxynonenal (4-HNE) is the most thoroughly characterised. Green tea polyphenols, theaflavins, and other SPMs could act as sacrificial nucleophiles trapping 4-HNE efficiently, the reason why tea and red wine polyphenols protect various types of cells in vitro, against HNE-induced intracellular oxidative stress and cytotoxicity [75]. In addition, a significant inhibition of HNE-induced mitochondrial ROS production is provided by quercetin, epigallocatechin gallate (EGCG), theaflavins and their gallate esters [76]. The most documented applications for skin damage protection under photo-irradiation anti-proliferative therapy concern standardised green tea extracts, due to their potent anti-inflammatory and photo-protective properties [67]. Green tea epicatechin derivatives, possessing free radical scavenging, antioxidant, anti-inflammatory, as well as direct anti-carcinogenic properties, have shown to be protective against UVB- or PUVA-induced photo-toxicity, with inflammation control through the suppression of proteasome function, NF-kappaB activity, and cytokine release [66]. Carotenoids, being a significant part of terpenoid SPMs, including beta-carotene, lycopene, lutein, zeaxanthin, and astaxanthin, have been studied for many years as remedies for cutaneous photo-protection [28]. The systemic intake of carotenoids displayed photo-protective effects after cutaneous accumulation. The mechanistic studies have revealed that carotenoids accumulated in the skin were light photon absorbers, served as sacrificial antioxidants, and quenchers of excited states and singlet oxygen. Recent publications have suggested bixin, an apocarotenoid from seeds of the achiote tree acted as a potent UV protector for cultured skin cells as well as after oral intake. Bixin mainly stimulated Toll-like receptor 4, an ancient system of immune response to bacterial particles, which is closely interconnected with Nrf2-dependent mechanism of adaptation/protection from deleterious effects of solar radiation. [25,28].
5. Pre-Selection of Sun-Protective Secondary Plant Metabolites
Usually, to achieve an optimal composition of topical sunscreens labeled as products with high broad-band UVA + UVB protection, several synthetic molecules having polyphenol nature-inspired moieties (derivatives of benzoic or cinnamic acids) are combined [17,18]. To meet requirements of regulatory bodies for claimed SPF-B and SPF-A values, these synthetic substances should be added to sunscreen cosmetics/drugs in high concentrations ranging from 10 to 25% [18] that sharply increases risks of adverse cutaneous reactions to poly-aromatic synthetics [12] and might have negative environmental impact.
The very first attempts to substitute synthetic sunscreens with the natural ones, for example, leaf extracts of Baccharis antioquensis containing quercetin, kaempferol, and caffeic acid glycosides [77] have been conducted. It has been published recently [23] that the glycosylated plant phenylpropanoids (polyphenols) baicalein and verbascoside protected cultured human keratinocytes against a broad band UVA + UBV simulating solar UV irradiation better than benzophenone-3. According to this paper and other publications [23,68], baicalein and verbascoside protected cell viability via multiple pathways: partly by physical UVA + UVB absorption, partly by direct free radical scavenging (chemical protection), and/or targeting adaptive/pro-survival, such as Nrf2-connected pathways in keratinocytes (biological protection).
Theoretically, SPMs could modify skin-UV interaction at several crucial points: (a) by absorption of UVA + UVB (screen action); (b) by inhibition of UV-induced free radical reactions in skin cells and extracellular matrix (scavenging and direct antioxidant chain-breaking effects); (c) by protection of skin surface lipid antioxidants, such as alpha-tocopherol, coenzyme Q10, and squalene (antioxidant rescue action); (d) by induction of endogenous antioxidant systems in keratinocytes and fibroblasts (indirect antioxidant effects); (e) by attenuation of inflammatory responses in cutaneous immune cells (keratinocytes/leukocytes/dendritic cells); (f) by modulation of excessive metabolic and proliferative UV-induced stress responses (anti-stress effects), and (g) by attenuation of UV-related immune suppression (immuno-modulation) [23,78,79,80].
The problem of photo-stability of sunscreens has been drawing close attention for quite some time because it might seriously affect desired durable photo-protection and recommendations regarding frequency of re-applications. Previous studies have shown that synthetic sunscreens lost a significant part of their protection when exposed to UV irradiation [81,82]. Unfortunately, natural UVA + UVB screens could also be susceptible to photo-degradation mainly by the UVA range of solar radiation [23,83]. To suppress photo-instability of natural chromophores based on SPM, several technological approaches have been applied in the course of formulation of fully natural sun protective cosmetics.
6. Formulation of Sun Protective Cosmetics Based on Secondary Plant Metabolites
Organic (synthetic) and mineral UV filters pose relevant challenges related to their photo-stability in vitro and in vivo under UV irradiation [81,82], their homogeneous distribution onto the skin [83,84,85], and their capability to reach the viable layers of epidermis and obtain sufficient accumulation in the stratum corneum [86] while also avoiding their systemic penetration due to the widespread concerns on their toxicity [87]. A major role is played by vehicles, which undergo a continuous technological evolution in the highly competitive cosmetic world, moving from the traditional vaseline or oil-in-water (o/w) emulsion delivery systems. This search for state-of-the-art formulations goes along with the attempt to develop innovative experimental methods to reliably assess effective penetration, homogeneity, stability, and bioavailability of the sunscreens in the various cosmetic micro-environments with different physical and chemical features [88,89]. A large potential portfolio of industrial applications of secondary plant metabolites promises breakthrough sunscreening properties, with reduced toxicity and appealing environmental sustainability. The natural UV filters are nevertheless also limited by several factors, including their low solubility with consequence difficulties in the cosmetic formulation, their fragile chemo- and photo-stability in the given cosmetic moiety, their reduced capability to penetrate the stratum corneum, and their limited bioavailability in the target skin layers [83,90]. These major drawbacks have been addressed by different formulations strategies, ranging from mere changes of vehicle viscosity to liposome or cyclodextrin complexation [90,91].
Many up-to-date studies support the advantages of formulation strategies searching for a synergic photo-protective effect among natural plant sources, able to exert synergic anti-photo-ageing, anti-inflammatory, and antioxidant activities [90]. Several vegetable oils have been proposed as photo-stability enhancers [92] and skin-friendly carriers by increasing the penetration of both synthetic and natural UV filters. Vegetable oils provide hydration to the sun-exposed skin and, in addition, can have an SPF themselves [93]. Plant oils based on renewable vegetable resources proved to be effective enhancers for chemical sunscreens. Soybean and olive oils displayed optimal performance as skin penetration enhancers for dihydroquercetin, a very effective inhibitor of skin photo-damage hitherto limited by its solubility in aqueous solvents inferior to 0.1% [94]. Future industrial-scale use shall rely on the appropriate standardisation of the vegetable sources, variable in their fatty acid composition and in the quality grade of raw materials. Rice bran oil and raspberry seed oil, formulated as advanced nano-structured lipid carriers with antioxidant properties, have shown minimal side effects and the ability to release controlled mounts of the encapsulated synthetic broad-spectrum filters butyl-methoxydibenzoylmethane and octocrylene, which were then needed in lower amounts [95]. Lipid micro- and nano-carriers represent the evolution of the pioneer liposome-based carriers. These were first found to enhance the skin penetration and absorption of the lipophilic sunscreens such as ethyl hexyl methoxycinnamate, as compared to the traditional oil/water emulsions supplied with emulsifiers [96], or the stratum corneum bio-adhesiveness of padimate O [97], although limited in the industrial applications by the unresolved problem of liposome instability. Nano-emulsifier particles also show the ability to inhibit trans-cutaneous absorption of the systemically toxic and immune-suppressing chemical filters [98]. Major limitations lay in the uncertain regulatory status of many polymers, and the elevated costs of large scale production [98,99].
Solid lipid nano-particles and nanostructure lipid carriers, in forms already available on the cosmetic market, have been proven to be effective and sustainable (solvent-free) carrier systems for natural lipid-soluble sun-protecting factors like lutein, by increasing its stability and inhibiting its physical-chemical degradation, while exerting skin hydrating and elasticity boosting essential cosmetic functions [99]. The encapsulation of flavonoids, such as rutin, with limited solubility in aqueous solvents and scarce permeation in the stratum corneum, showed the promising advantages of gelatin nano-particles entrapping rutin, which, when coupled, enhanced sun protection with an elevated skin compatibility of the new vehicle [100]. Genistein was successfully encapsulated in Vitamin E nano-particle emulsions with photo-chemo-preventive efficacy [101]. Mono-oleate lipid nano-particles display a liquid crystal gel structure in water-rich environments. They were feasible for the co-formulation with other cosmetic ingredients, have been shown to enhance trans-epidermal penetration and bio-adhesiveness of lipophilic natural compounds like Vitamin K [102]. Surfactant Transcutol®, coupled with penetration enhancers like sucrose esters, are among the cosmetic excipients enabling increased solubility, penetration, and bioavailability, of attractive UV-protecting plant actives against UVB-induced oxidative stress, like Silybum marianum extracts [103].
In spite of the ongoing intense biological investigations and technological improvements, so far, no entirely natural UV filter has been officially approved for the EU market (Annex VI, Directive. EC-1223/2009). Waiting for the expected breakthroughs in the cosmetic regulations in the application of natural sunscreens alone, the advantages of a synergy between synthetic and plant-derived UV filters have been widely assessed, based on the antioxidant and anti-inflammatory properties of the plant molecules, which are able to enhance the sunscreen efficacy by protecting its photo-degradation and by boosting the target tissue endogenous defence from UV-driven inflammation [104]. In contrast, chemical filters are also being investigated as low-dose stabilisers for plant sunscreens, as in the case of green tea catechins. For example, epigallocatechin-3-gallate in oil/water emulsions was the most effectively protected from UV degradation by low concentrations of the water-soluble UVB filter Benzophenone-4 [83].
The combination of chemical sunscreens with melatonin, rutin, lipophilic antioxidants proved their efficacy as sun protection boosters and inhibitors of skin photo-oxidative damage [90,105]. For the conventional synthetic organic filters, many of which are potential endocrine disruptors, the inhibition of their deep penetration in the dermis remains a primary clinical goal. This has been traditionally attempted by increasing of vehicle viscosity. More effective increase of synthetic UV-filters safety has been recently achieved with the co-formulation of beta-carotene and resveratrol. These plant-derived SPMs inhibited deep transcutaneous penetration of synthetic UV filters and prolonged their accumulation in the stratum corneum [87]. This may represent a feasible approach to the reduction of repeated sunscreen applications to ensure sunscreen compliance [106].
The reduction of UV filter concentrations in sunscreen cosmetics, achieved through innovative delivery strategies or by the combination or substitution with plant-derived natural filters, will also have a major impact on environmental sustainability, at risk for the high concentration of endocrine disrupting chemical UV-filters in aquatic environments. Concerns have been raised, however, on the safety of the long-term application of the novel vehicles, and on the environmental impact of cosmetic micro- and nano-particles [107].
7. Conclusions: Advantages and Disadvantages of Fully Natural Sun Protective Cosmetics
The idea to develop fully natural sun protective cosmeceuticals and skin care products is one of the most attractive yet challenging health concepts in the modern highly technological world. This idea is an “evolution inspired” because it is based on striking similarities of endogenous UV protection/adaptation in humans, terrestrial and marine plants. Numerous SPMs synthesised by plants/marine organisms on request in response to challenges of the hostile environment, first of all, solar irradiation seems to be excellent candidates to prevent, decelerate or even revert skin damage provided by solar radiation [108]. Photo-protective plant-derived SPMs have a chemical structure of chromophores capable of effectively absorbing solar light energy within a wide range of biologically damaging UVA and UVB spectra (sunscreen properties). After physical interaction with solar energy, they could maintain their initial chemical structure for quite some time that is a feature of their photo-stability. Due to photo-stability, plant-derived SPMs should not theoretically exert photo-toxic effects against skin cells and extracellular matrix. Very peculiar redox characteristics and metal chelating properties make SPMs excellent free radical scavengers and direct antioxidants to prevent and repair photo-oxidative damage provided by solar radiation [109]. A combination of sunscreen and redox protection allows for SPMs to rescue endogenous skin-located low molecular antioxidants (antioxidant rescue properties). Being polyaromatic xenobiotics, SPMs interact with endogenous cutaneous antioxidant and detoxifying enzymes constructing metabolic and redox barriers of the skin. This interaction leads to restoration of normal antioxidant defence (indirect antioxidant action) [110] and to modulation of phase I and phase II metabolic enzymes to eliminate toxic photo-oxidised metabolites of lipid degradation and SPMs decomposition (detox action). Structural similarity of SPMs and human bioactive molecules results in the ability of SPMs to interact with multiple receptors on skin immune cells and to alter molecular pathways leading to their response to UV irradiation that attenuates UV-associated pro-inflammatory signalling (anti-inflammatory action). To achieve all the above-mentioned positive properties and actions of SPM, there should be serious research on mechanisms/dose-dependent anti-UV effects of single isolated SPM, of its combination with other potential photo-protectors, as well as their anti-UV effects within a final sun protective cosmetic product.
Conflicts of Interest
C.D.L. and W.M. work for MEDENA AG, a company producing sun protective cosmetics. L.K. is currently a scientific consultant for the company. The other authors have no conflict of interests.
References
- De Luca, C.; Valacchi, G. Surface lipids as multifunctional mediators of skin responses to environmental stimuli. Mediat. Inflamm. 2010, 2010, 321494. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, V.A.; Potapovich, A.I.; Lulli, D.; Stancato, A.; De Luca, C.; Pastore, S.; Korkina, L. Modulation of human keratinocyte responses to solar UV by plant polyphenols as a basis for chemoprevention of non-melanoma skin cancers. Curr. Med. Chem. 2013, 20, 869–879. [Google Scholar] [PubMed]
- Scharffetter-Kochanek, K.; Wlaschek, M.; Brenneisen, P.; Schauen, M.; Blaudschun, R.; Wenk, J. UV-induced reactive oxygen species in photocarcinogenesis and photoaging. Biol. Chem. 1997, 378, 1247–1257. [Google Scholar] [PubMed]
- Pallela, R.; Young, Y.N.; Kim, S.K. Anti-photoaging and photoprotective compounds derived from marine organisms. Mar. Drugs 2010, 8, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- Chanchal, D.; Swarnlata, S. Herbal photoprotective formulations and their evaluation. Open Nat. Prod. J. 2009, 2, 71–76. [Google Scholar] [CrossRef]
- Saewan, N.; Jimtaisong, A. Natural products as photoprotection. J. Cosmet. Dermatol. 2015, 14, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Korkina, L.; Mayer, W.; De Luca, C. Meristem plant cells as a sustainable source of redox actives for skin rejuvenation. Biomolecules 2017, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Douglas, C.J. Phenylpropanoid metabolism and lignin biosynthesis: From weeds to trees. Trends Plant Sci. 1996, 1, 171–178. [Google Scholar] [CrossRef]
- Korkina, L.G. Phenylpropanoids as naturally occurring antioxidants: From plant defence to human health. Cell. Mol. Biol. 2007, 53, 15–25. [Google Scholar] [PubMed]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [PubMed]
- Groniger, A.; Sinha, R.P.; Klisch, M.; Häder, D.P. Photoprotective compounds in cyanobacteria, phytoplankton and macroalgae—A database. J. Photochem. Photobiol. B Biol. 2000, 58, 115–122. [Google Scholar] [CrossRef]
- Korkina, L.G.; Pastore, S.; De Luca, C.; Kostyuk, V.A. Metabolism of plant polyphenols in the skin: Beneficial versus deleterious effects. Curr. Drug Metab. 2008, 9, 710–729. [Google Scholar] [CrossRef] [PubMed]
- Perez-Sancez, A.; Barrajon-Catalan, E.; Caturla, N.; Castillo, J.; Benavente-Garcia, O.; Alcaraz, M.; Micol, V. Protective effects of citrus and rosemary extracts on UV-induced damage in skin cell model and human volunteers. J. Photochem. Photobiol. B 2014, 136, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Frohnmeyer, H.; Staiger, D. Ultraviolet-B radiation-mediated responses in plants. Balancing damage and protection. Plant Physiol. 2003, 133, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Hahlbrock, K.; Scheel, D. Physiology and molecular biology of phenylpropanoid metabolism. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1989, 4, 347–369. [Google Scholar] [CrossRef]
- Moyal, D.D.; Fourtanier, A.M. Broad-spectrum sunscreens provide better protection from solar ultraviolet-stimulated radiation and natural sunlight-induced immunosuppression in human beings. J. Am. Acad. Dermatol. 2008, 58, S149–S154. [Google Scholar] [CrossRef] [PubMed]
- Wolf, R.; Wolf, D.; Morganti, P.; Ruocco, V. Sunscreens. Clin. Dermatol. 2001, 9, 452–459. [Google Scholar] [CrossRef]
- Commission recommendation of 22 September 2006 on the efficacy of sunscreen products and the claims made relating thereto. Off. J. Eur. Union 2006, 265, 39–43.
- Dembitsky, V.M. Astonishing diversity of natural surfactants: 5. Biologically active glycosides of aromatic metabolites. Lipids 2005, 40, 869–900. [Google Scholar] [CrossRef] [PubMed]
- Ju, K.Y.; Kang, J.; Chang, J.H.; Lee, J.K. Clue to understanding the Janus behavior of eumelanin: Investigating the relationships between hierarchical assembly structure of eumelanin and its photo physical properties. Biomacromolecules 2016, 17, 2860–2872. [Google Scholar] [CrossRef] [PubMed]
- Bilkis, I.; Silman, I.; Weiner, L. Generation of reactive oxygen species by photosensitizers and their modes of action on proteins. Curr. Med. Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Korkina, L. Metabolic and redox barriers in the skin exposed to drugs and xenobiotics. Exp. Opin. Drug Metab. Toxicol. 2016, 12, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, V.; Potapovich, A.; Albuhaydar, A.R.; Mayer, W.; De Luca, C.; Korkina, L. Natural substances for prevention of skin photoageing: Screening systems in the development of sunscreen and rejuvenation cosmetics. Rejuvenation Res. 2017. [Google Scholar] [CrossRef]
- Herrmann, G.; Wlaschek, M.; Bolsen, K.; Prenzel, K.; Goerz, G.; Scharffetter-Kochanek, K. Photosensitisation of uroporphyrin augments the ultraviolet A-induced synthesis of matrix metalloproteinases in human dermal fibroblasts. J. Investig. Dermatol. 1996, 107, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Wondrak, G.T.; Jacobson, M.K.; Jacobson, E.L. Endogenous UVA-photosensitizers: Mediators of skin photodamage and novel targets for skin photoprotection. Photochem. Photobiol. Sci. 2006, 5, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Douki, T.; Ravanat, J.L.; Di Mascio, P. Sensitised formation of oxidatively generated damage to cellular DNA by UVA radiation. Photochem. Photobiol. Sci. 2009, 8, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Lamore, S.D.; Azimian, S.; Horn, D.; Anglin, B.L.; Uchida, K.; Cabello, C.M.; Wondrak, G.T. The malondialdehyde-derived fluorophore DPH-lysine is a potential sensitizer of UVA-induced photooxidative stress in human skin cells. J. Photochem. Photobiol. 2010, 101, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, S.E.; Wondrak, G.T. TLR4-directed molecular strategies targeting skin photodamage and carcinogenesis. Curr. Med. Chem. 2017. [Google Scholar] [CrossRef] [PubMed]
- De Luca, C.; Picardo, M.; Breathnach, A.; Passi, S. Lipoperoxidase activity of Pityrosporum: Characterisation of by-products and possible role in Pityriasis versicolor. Exp. Dermatol. 1996, 5, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Ekanayake Mudiyanselage, S.; Hamburger, M.; Elsner, P.; Thiele, J.J. Ultraviolet A induces generation of squalene monohydroperoxide isomers in human sebum and skin surface lipids in vitro and in vivo. J. Investig. Dermatol. 2003, 120, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Picardo, M.; Mastrofrancesco, A.; Biro, T. Sebaceous gland—A major player in skin homeostasis. Exp. Dermatol. 2015, 24, 485–486. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, V.; Potapovich, A.; Stancato, A.; De Luca, C.; Lulli, D.; Pastore, S.; Korkina, L. Photo-oxidation products of skin surface squalene mediate metabolic and inflammatory responses to solar UV in human keratinocytes. PLoS ONE 2012, 7, e44472. [Google Scholar] [CrossRef] [PubMed]
- Auffray, B. Protection against singlet oxygen, the main actor of sebum squalene peroxidation during sun exposure, using Commiphora myrrha essential oil. Int. J. Cosmet. Sci. 2007, 29, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Rannug, A.; Fritsche, E. The aryl hydrocarbon receptor and light. Biol. Chem. 2006, 387, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Shiraishi, M.; Naito, Y.; Torii, Y.; Nakamura, Y.; Osawa, T. Activation of stress signaling pathways by the end product of lipid peroxidation. J. Biol. Chem. 1999, 274, 2234–2242. [Google Scholar] [CrossRef] [PubMed]
- Hattori, T.; Obinata, H.; Ogawa, A.; Kishi, M.; Tatei, K.; Ishikawa, O.; Izumi, T. G2A plays proinflammatory roles in human keratinocytes under oxidative stress as a receptor for 9-hydroxyoctadecadenoic acid. J. Investig. Dermatol. 2008, 128, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Hibbert, S.A.; Watson, R.E.B.; Gibbs, N.K.; Costello, P.; Baldock, C.; Weiss, A.S.; Griffiths, C.E.; Sherratt, M.J. A potential role for endogenous proteins as sacrificial sunscreens and antioxidants in human tissues. Redox Biol. 2015, 5, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.J.; Han, J.; Kim, B.H.; Lee, Y.S.; Kim, T.Y. Superoxide dismutase 3 suppresses hyaluronic acid fragments mediated skin inflammation by inhibition of toll-like receptor 4 signaling pathway: Superoxide dismutase 3 inhibits reactive oxygen species-induced trafficking of toll-like receptor 4 to lipid rafts. Antioxid. Redox Signal. 2012, 16, 297–313. [Google Scholar] [PubMed]
- Thiele, J.; Barland, C.O.; Ghadially, R.; Elias, P. Permeability and antioxidant barriers in aged skin. In Skin Aging; Gilchrest, B., Krutmann, J., Eds.; Springer: Berlin, Germany, 2006. [Google Scholar]
- Packer, L.; Valacchi, G. Antioxidants and the response of skin to oxidative stress: Vitamin E as an indicator. Skin Pharmacol. Appl. Skin Physiol. 2002, 15, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Semak, I.; Zbytek, B.; Pisarchik, A.; Li, W.; Zjawiony, J.; Tuckey, R.C. Cytochromes P450 and skin cancer: Role of local endocrine pathways. Anticancer Agents Med. Chem. 2014, 14, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Kleszczynski, K.; Semak, I.; Janjetovic, Z.; Zmijewski, M.A.; Kim, T.K.; Slominski, R.M.; Reiter, R.J.; Fischer, T.W. Local melatoninergic system as the protector of skin integrity. Int. J. Mol. Sci. 2014, 15, 17705–17732. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Kleszczynski, K.; Janjetovic, Z.; Sweatman, T.; Lin, Z.; Li, W.; Reiter, R.J.; Fischer, T.W.; Slominski, A.T. Metabolism of melatonin and biological activity of intermediates of melatoninergic pathway in human skin cells. FASEB J. 2013, 27, 2742–2755. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Surh, Y.J. Nrf2 as a novel molecular target for chemoprevention. Cancer Lett. 2005, 224, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Role of Nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed]
- Schafer, M.; Werner, S. Nrf2-A regulator of keratinocyte redox signaling. Free Radic. Biol. Med. 2015, 88, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Rojo de la Vega, M.; Krajisnik, A.; Zhang, D.D.; Wondrak, G.T. Targeting NRF2 for improved skin barrier function and photoprotection: Focus on the achiote-derived apocarotenoid bixin. Nutrients 2017, 9, 1371. [Google Scholar] [CrossRef] [PubMed]
- Hirota, A.; Kawachi, Y.; Yamamoto, M.; Koga, T.; Hamada, K.; Otsuka, F. Acceleration of UVB-induced photoageing in nrf2 gene-deficient mice. Exp. Dermatol. 2011, 20, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Bosch, R.; Philips, N.; Suarez-Perez, J.A.; Juarranz, A.; Devmurari, A.; Chalensouk-Khaosaat, J.; Gonzalez, S. Mechanisms of Photoaging and Cutaneous Photocarcinogenesis, and Photoprotective Strategies with Phytochemicals. Antioxidants 2015, 4, 248–268. [Google Scholar] [CrossRef] [PubMed]
- Oesch, F.; Fabian, E.; Guth, K.; Landsiedel, R. Xenobiotic-metabolizing enzymes in the skin of rat, mouse, pig, guinea, and in human skin models. Arch. Toxicol. 2014, 88, 2135–2190. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K.; Matsui, M.S.; Mukhtar, H. Ultraviolet-B exposure of human skin induces cytochromes P450 1A1 and 1B1. J. Investig. Dermatol. 2000, 114, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Pavek, P.; Dvorak, Z. Xenobiotic-induced transcriptional regulation of xenobiotic metabolizing enzymes of the cytochrome P450 superfamily in human extrahepatic tissues. Curr. Drug Metab. 2008, 9, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Baron, J.M.; Holler, D.; Schiffer, R.; Frankenberg, S.; Neis, M.; Merk, H.F.; Jugert, F.K. Expression of multiple cytochrome P450 enzymes and multidrug resistance-associated transport proteins in human skin keratinocytes. J. Investig. Dermatol. 2001, 116, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Gundert-Remy, U.; Bernauer, U.; Blomeke, B.; Döring, B.; Fabian, E.; Goebel, C.; Hessel, S.; Jäckh, C.; Lampen, A.; Oesch, F.; et al. Extrahepatic metabolism at the body’s internal-external interfaces. Drug Metab. Rev. 2014, 46, 291–324. [Google Scholar] [CrossRef] [PubMed]
- Chiaro, C.R.; Patel, R.D.; Perdew, G.H. 12(R)-hydroxy-5(Z),8(Z),10(E),14(Z)-eicosatetraenoic acid [12(R)-HETE], an arachidonic acid derivative, is an activator of the aryl hydrocarbon receptor. Mol. Pharmacol. 2008, 74, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.P.; Bradfield, C.A. The search for endogenous activators of aryl hydrocarbon receptor. Chem. Res. Toxicol. 2008, 21, 102–116. [Google Scholar] [CrossRef] [PubMed]
- Stejskalova, L.; Dvorak, Z.; Pavek, P. Endogenous and exogenous ligands of aryl hydrocarbon receptor: Current state of art. Curr. Drug Metab. 2011, 12, 198–212. [Google Scholar] [CrossRef] [PubMed]
- Kalthoff, S.; Ehmer, U.; Freiberg, N.; Manns, M.P.; Strassburg, C.P. Interaction between oxidative stress sensor Nrf2 and xenobiotic-activated aryl hydrocarbon receptor in the regulation of the human phase II detoxifying UDP-glucuronosyltransferase 1A10. J. Biol. Chem. 2010, 285, 5993–6002. [Google Scholar] [CrossRef] [PubMed]
- Swanson, H.I. Cytochrome P450 expression in human keratinocytes: An aryl hydrocarbon receptor perspective. Chem. Biol. Interact. 2004, 149, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Puga, A.; Ma, C.; Marlowe, J.L. The aryl hydrocarbon receptor cross-talks with multiple signal transduction pathways. Biochem. Pharmacol. 1999, 77, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Ohtake, F.; Takeyama, K.; Matsumoto, T.; Kitagawa, H.; Yamamoto, Y.; Nohara, K.; Tohyama, C.; Krust, A.; Mimura, J.; Chambon, P.; et al. Modulation of estrogen receptor signaling by association with the activated dioxin receptor. Nature 2003, 423, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Guyot, E.; Chevallier, A.; Barouki, R.; Coumoul, X. The AhR twist: Ligand-dependent AhR signaling and pharmaco-toxicological implications. Drug Discov. Today 2013, 18, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Busbee, P.B.; Busbee, M.; Rouse, M.; Nagarkatti, M.; Nagarkatti, P.S. Use of natural AhR ligands as potential therapeutic modalities against inflammatory disorders. Nutr. Rev. 2013, 71, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Pastore, S.; Lulli, D.; Potapovich, A.I.; Fidanza, P.; Kostyuk, V.A.; Dellambra, E.; De Luca, C.; Maurelli, R.; Korkina, L. Differential modulation of stress-inflammation responses by plant polyphenols in cultured normal human keratinocytes and immortalized HaCaT cells. J. Dermatol. Sci. 2011, 63, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K. UV-induced immune suppression and photocarcinogenesis: Chemoprevention by dietary botanical agents. Cancer Lett. 2007, 255, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K.; Afaq, F.; Peres, A.; Mukhar, H. Green tea polyphenol (−)-epigallocatechin-3-gallate treatment of human skin inhibits ultraviolet radiation-induced oxidative stress. Carcinogenesis 2001, 22, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Afaq, F.; Adhami, V.M.; Ahmad, N.; Mukhtar, H. Inhibition of ultraviolet B-mediated activation of nuclear factor κB in normal human epidermal keratinocytes by green tea constituent (−)-epigallocatechin-3-gallate. Oncogene 2003, 22, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Potapovich, A.I.; Lulli, D.; Fidanza, P.; Kostyuk, V.A.; De Luca, C.; Pastore, S.; Korkina, L. Plant polyphenols differentially modulate inflammatory responses of human keratinocytes by interfering with activation of transcriptional factors NFκB and AhR and EGFR-ERK pathways independently of their direct redox properties. Toxicol. Appl. Pharmacol. 2011, 255, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Pastore, S.; Lulli, D.; Pascarella, A.; Maurelli, R.; Dellambra, E.; Potapovich, A.; Kostyuk, V.; De Luca, C.; Korkina, L. Resveratrol enhances solar UV induced responses in normal human epidermal keratinocytes. Photochem. Photobiol. 2012, 88, 1522–1530. [Google Scholar] [CrossRef] [PubMed]
- Wright, T.I.; Spencer, J.M.; Flowers, F.P. Chemoprevention of nonmelanoma skin cancer. J. Am. Acad. Dermatol. 2006, 54, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Huang, M.T.; Ferraro, T.; Wong, C.Q.; Lou, Y.R.; Reuhl, K.; Iatropulos, M.; Yang, C.S.; Connery, A.H. Inhibitory effect of green tea in the drinking water on tumorigenesis by ultraviolet light and 12-O-tetradecanoylphorbol-13-acetate in the skin of SKH-1 mice. Cancer Res. 1992, 52, 1162–1170. [Google Scholar] [PubMed]
- Wei, H.; Saladi, R.; Lu, Y.; Wang, Y.; Palep, S.R.; Moore, J.; Phelps, R.; Shyong, E.; Lebwohl, M.G. Isoflavone genistein: Photoprotection and clinical implications in dermatology. J. Nutr. 2003, 133, 3811S–3819S. [Google Scholar] [CrossRef] [PubMed]
- Picardo, M.; Zompetta, C.; De Luca, C.; Cirone, M.; Faggioni, A.; Nazzaro-Porro, M.; Passi, S.; Prota, G. Role of skin surface lipids in UV-induced epidermal cell changes. Arch. Dermatol. Res. 1991, 283, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Uchino, T.; Tokunaga, H.; Onodera, H.; Ando, M. Effect of squalene monohydroperoxide on cytotoxicity and cytokine release in a three-dimensional human skin model and human epidermal keratinocytes. Biol. Pharm. Bull. 2002, 25, 605–610. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhu, Q.; Zheng, Z.P.; Cheng, K.W.; Wu, J.J.; Zhang, S.; Tang, Y.S.; Sze, K.H.; Chen, J.; Chen, F.; Wang, M. Natural polyphenols as direct trapping agents of lipid peroxidation-derived acrolein and 4-hydroxy-trans-2-nonenal. Chem. Res. Toxicol. 2009, 22, 1721–1727. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Kumagai, T.; Torii, Y.; Nakamura, Y.; Osawa, T.; Uchida, K. Anticarcinogenic antioxidants as inhibitors against intracellular oxidative stress. Free Radic. Res. 2001, 35, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Mejia-Giraldo, J.C.; Winkler, R.; Gallardo, C.; Sanchez-Zapata, A.M.; Puertas-Majia, M.A. Photoprotective potential of Baccharis antioquensis (Asteraceae) as natural sunscreens. Photochem. Photobiol. 2016, 92, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Potapovich, A.I.; Kostyuk, V.A.; Kostyuk, T.V.; De Luca, C.; Korkina, L.G. Effects of pre- and post-treatment with plant polyphenols on human keratinocyte responses to solar UV. Inflamm. Res. 2013, 62, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Lulli, D.; Patapovich, A.; Maurelli, L.; Dellambra, E.; Pressi, G.; Kostyuk, V.; Dal Toso, R.; De Luca, C.; Pastore, S.; Korkina, L. Anti-inflammatory effects of concentrated ethanol extracts of Edelweiss (Leontopodium alpinum Cass.) callus cultures towards human keratinocytes and endothelial cells. Mediat. Inflamm. 2012, 2012, 498373. [Google Scholar] [CrossRef]
- Pastore, S.; Lulli, D.; Maurelli, R.; Dellambra, E.; De Luca, C.; Korkina, L. Resveratrol induces long-lasting IL-8 expression and peculiar EGFR activation/distribution in human keratinocytes: Mechanisms and implications for skin administration. PLoS ONE 2013, 8, e59632. [Google Scholar] [CrossRef] [PubMed]
- Maier, H.; Schauberger, G.; Brunnhofer, K.; Honigsmann, H. Change in ultraviolet absorbance of sunscreens by exposure to solar-simulated radiation. J. Investig. Dermatol. 2001, 117, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Marrot, L.; Belaidi, J.P.; Lejeune, F.; Maunier, J.R.; Asselineau, D.; Bernerd, F. Photostability of sunscreen products influences the efficiency of protection with regard to UV-induced genotoxic or photoageing-related endpoints. Br. J. Dermatol. 2004, 151, 1234–1244. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, A.; Marchetti, N.; Scalia, S. Photodegradation of (−)-epigallocatechin-3-gallate in topical cream formulations and its photostabilization. J. Pharm. Biomed. Anal. 2011, 56, 692–697. [Google Scholar] [CrossRef] [PubMed]
- Lademann, J.; Rudolph, A.; Jacobi, U.; Weigmann, H.J.; Schaefer, H.; Sterry, W.; Meinke, M. Influence of nonhomogeneous distribution of topically applied UV filters on sun protection factors. J. Biomed. Opt. 2004, 9, 1358–1362. [Google Scholar] [CrossRef] [PubMed]
- Sohn, M.; Hêche, A.; Herzog, B.; Imanidis, G. Film thickness frequency distribution of different vehicles determines sunscreen efficacy. Biomed. Opt. 2014, 19, 115005. [Google Scholar] [CrossRef] [PubMed]
- Durand, L.; Habran, N.; Henschel, V.; Amighi, K. In vitro evaluation of the cutaneous penetration of sprayable sunscreen emulsions with high concentrations of UV filters. Int. J. Cosmet. Sci. 2009, 31, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Freitas, J.V.; Praça, F.S.; Bentley, M.V.; Gaspar, L.R. Trans-resveratrol and beta-carotene from sunscreens penetrate viable skin layers and reduce cutaneous penetration of UV-filters. Int. J. Pharm. 2015, 30, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Stiefel, C.; Schwack, W.; Nguyen, Y.-T.H. Photostability of Cosmetic UV Filters on Mammalian Skin under UV Exposure. Photochem. Photobiol. 2015, 91, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Roussel, L.; Gilbert, E.; Salmon, D.; Serre, C.; Gabard, B.; Haftek, H.; Maibach, H.I.; Pirot, F. Measurement, analysis and prediction of topical UV filter bioavailability. Int. J. Pharm. 2015, 478, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Radice, M.; Manfredini, S.; Ziosi, P.; Dissette, V.; Buso, P.; Fallacara, A.; Vertuani, S. Herbal extracts, lichens and biomolecules as natural photo-protection alternatives to synthetic UV filters. A systematic review. Fitoterapia 2016, 114, 144–162. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.S.; Ozzetti, R.A.; Vergnanini, A.L.; de Brito-Gitirana, L.; Volpato, N.M.; de Freitas, Z.M.; Ricci-Júnior, E.; dos Santos, E.P. Evaluation of octyl p-methoxycinnamate included in liposomes and cyclodextrins in anti-solar preparations: Preparations, characterizations and in vitro penetration studies. Int. J. Nanomed. 2012, 7, 3045–3058. [Google Scholar]
- L’alloret, F.; Candau, D.; Seité, S.; Pygmalion, M.J.; Ruiz, L.; Josso, M.; Meaudre, H.; Gauchet, L.; Pena, A.M.; Colonna, A. New combination of ultraviolet absorbers in an oily emollient increases sunscreen efficacy and photostability. Dermatol. Ther. 2012, 2, 4. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kaur, C.D.; Saraf, S. In vitro sun protection factor determination of herbal oils used in cosmetics. Pharmacogn. Res. 2010, 2, 22–25. [Google Scholar]
- Cˇižinauskas, V.; Elie, N.; Brunelle, A.; Briedis, V. Skin Penetration Enhancement by Natural Oils for Dihydroquercetin Delivery. Molecules 2017, 22, 1536. [Google Scholar] [CrossRef] [PubMed]
- Niculae, G.; Lacatusu, I.; Badea, N.; Stan, R.; Vasile, B.S.; Meghea, A. Rice bran and raspberry seed oil-based nanocarriers with self-antioxidative properties as safe photoprotective formulations. Photochem. Photobiol. Sci. 2014, 13, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Ramón, E.; Alonso, C.; Coderch, L.; De La Maza, A.; Lopez, O.; Parra, L.; Notario, J. Liposomes as Alternative Vehicles for Sun Filter Formulations. Drug Deliv. 2005, 12, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Ediriwickrema, A.; Yang, F.; Lewis, J.; Girardi, M.; Saltzman, W.M. A sunblock based on bioadhesive nanoparticles. Nat. Mater. 2015, 14, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Luppi, B.; Cerchiara, T.; Bigucci, F.; Basile, R.; Zecchi, V. Polymeric nanoparticles composed of fatty acids and polyvinylalcohol for topical application of sunscreens. J. Pharm. Pharmacol. 2004, 56, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Mitri, K.; Shegokar, R.; Gohla, S.; Anselmi, C.; Müller, R.H. Lipid nanocarriers for dermal delivery of lutein: Preparation, characterization, stability and performance. Int. J. Pharm. 2011, 414, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.A.; Peres, D.D.; Graziola, F.; Chacra, N.A.; Araújo, G.L.; Flórido, A.C.; Mota, J.; Rosado, C.; Velasco, M.V.; Rodrigues, L.M.; et al. Cutaneous biocompatible rutin-loaded gelatin-based nanoparticles increase the SPF of the association of UVA and UVB filters. Eur. J. Pharm. Sci. 2016, 81, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Brownlow, B.; Nagaraj, V.J.; Nayel, A.; Joshi, M.; Elbayoumi, T. Development and in vitro evaluation of vitamin E-enriched nanoemulsion vehicles loaded with genistein for chemoprevention against UVB-induced skin damage. J. Pharm. Sci. 2015, 104, 3510–3523. [Google Scholar] [CrossRef] [PubMed]
- Lopes, L.B.; Speretta, F.F.F.; Vitoria, M.; Bentley, L.B. Enhancement of skin penetration of vitamin K using monoolein-based liquid crystalline systems. Eur. J. Pharm. Sci. 2007, 32, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Fehér, P.; Ujhelyi, Z.; Váradi, J.; Fenyvesi, F.; Róka, E.; Juhász, B.; Varga, B.; Bombicz, M.; Priksz, D.; Bácskay, I.; Vecsernyés, M. Efficacy of Pre- and Post-Treatment by Topical Formulations Containing Dissolved and Suspended Silybum marianum against UVB-Induced Oxidative Stress in Guinea Pig and on HaCaT Keratinocytes. Molecules 2016, 21, 1269. [Google Scholar] [CrossRef] [PubMed]
- Steenvoorden, D.P.; van Henegouwen, G.M. The use of endogenous antioxidants to improve photoprotection. J. Photochem. Photobiol. B. 1997, 41, 1–10. [Google Scholar] [CrossRef]
- Sierra, A.F.; Ramírez, M.L.; Campmany, A.C.; Martínez, A.R.; Naveros, B.C. In vivo and in vitro evaluation of the use of a newly developed melatonin loaded emulsion combined with UV filters as a protective agent against skin irradiation. J. Dermatol. Sci. 2013, 69, 202–214. [Google Scholar] [CrossRef] [PubMed]
- De Villa, D.; da Silva Nagatomi, A.R.; Paese, K.; Guterres, S.; Ferreira Cestari, T. Reapplication Improves the Amount of Sunscreen, not its Regularity, under Real Life Conditions. Photochem. Photobiol. 2011, 87, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Stiefel, C.; Schwack, W. Photoprotection in changing times—UV filter efficacy and safety, sensitization processes and regulatory aspects. Int. J. Cosmet. Sci. 2015, 37, 2–30. [Google Scholar] [CrossRef] [PubMed]
- Baldisserotto, A.; Buso, P.; Radice, M.; Dissette, V.; Lampronti, I.; Gambari, R.; Manfredini, S.; Vertuani, S. Moringa oleifera leaf extracts as mutifunctional ingredients for “natural and organic” sunscreens and photoprotective preparations. Molecules 2018, 23, 664. [Google Scholar] [CrossRef] [PubMed]
- Girotti, A.W. Photosensitized oxidation of membrane lipids: Reaction pathways, cytotoxic effects, and cytoprotective mechanisms. J. Photochem. Photobiol. B 2001, 63, 103–113. [Google Scholar] [CrossRef]
- Shen, T.; Chen, X.M.; Harder, B.; Long, M.; Wang, X.N.; Lou, H.X.; Wondrak, G.T.; Ren, D.M.; Zhang, D.D. Plant extracts of the family Lauraceae: A potential resource for chemopreventive agents that activate the nuclear factor-erythroid 2-related factor 2/antioxidant response element pathway. Planta Med. 2014, 80, 426–434. [Google Scholar] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).