Abstract
Like with skin, both men and women—regardless of ethnicity—gradually lose satisfaction with changes in hair brought about by ageing. Especially when such transition is apparent by others, and that the hair condition has a significant role in an individual’s overall physical appearance and self-perception. Beyond the familiar age-related signs such as hair greying, hair loss, and hair fragility, this review includes current knowledge of biological processes underlying hair pigmentation and hair growth, highlights variations in gender and ethnicity, as well as delineates hair fibre diameter, ellipticity, and elasticity properties that collectively contribute to the characteristics of aged hair. Additionally, in view of the rising importance of enhancing scalp skin health to promote healthy hair growth, the latter part of the review focuses on age-associated alterations to the scalp skin and its microbiome. Consideration of the morphological changes in the hair fibre, biological processes occurring within the hair follicle and its enveloping scalp environment provide a unique, holistic overview of hair and scalp changes during ageing. Finally, after acknowledging the impact caused by chronological ageing and environmental stresses, it is important to recognise that healthy tresses are largely influenced by scalp skin care, and this stimulates the advancement of appropriate cosmetic solutions that help delay or improve the appearance of aged hair.
1. Introduction
The condition of the hair entails a significant role in an individual’s overall physical appearance and self-perception. Although human hair has evolved to be unnecessary for primary survival, poor hair health and hair loss can cause detrimental emotional, psychological, and social impacts. As early as can be traced, there have been archaeological discoveries and ancient written documentations testifying humanity’s obsession with the appearance of hair [1,2]. Experts have proposed that hair presented as healthy, shiny, and strong signal overall good physical health and reproductive potential [3,4,5]. A study conducted by the University of Göttingen further showed that variations in hair colour and hair styling influenced impressions of age, health, and attractiveness [6]. Hence, in addition to facial shape and skin condition, hair also strongly contributes to the perception of beauty.
To better comprehend hair ageing, it is paramount to grasp the different aspects and dynamicity of hair growth. Human hair normally follows a lifelong, continuous cyclic pattern of growth known as the hair growth cycle. It comprises primarily of four phases: anagen—characterised by rapid growth and proliferation, catagen—a brief regression stage, telogen—a quiescent stage, and exogen—the eventual shedding of the hair shaft [7] (Figure 1). As the longest phase of the hair cycle, the duration of anagen for scalp hairs is typically 2–8 years, whilst catagen and telogen lasts approximately 2 weeks and 2–3 months, respectively [8]. Multiple factors have been shown to influence the transition between the anagen and telogen phases. In healthy conditions, approximately 85–90% of human scalp follicles are in the anagen phase, and the rest are either in catagen or telogen [9]. However, with age, the length of the anagen phase tends to shorten, and the proportion of hair follicles in anagen, with respect to telogen follicles, declines [10]. Furthermore, distinct from murine hair growth, individual human hair follicles cycle independently and asynchronously, with each hair follicle potentially capable of undergoing 10–30 cycles in a lifetime. At any one time, most individuals have an estimated average of 100,000 hairs on their scalp, with normal shedding taking place at a rate of 100–150 strands per day [8,11].
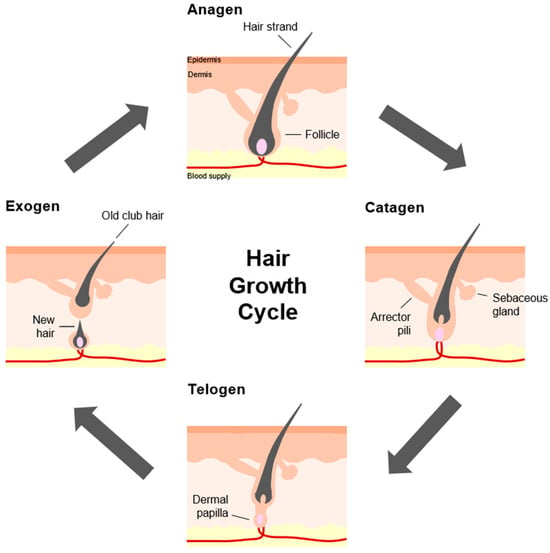
Figure 1.
Stages of the hair growth cycle. The hair growth cycle, also referred to as the hair follicle cycle, consists of the anagen, catagen, telogen, and exogen phases. (Starting from the top box; clockwise direction).
Depending on their location, hair follicles may vary in size and shape but are generally characterised by the same structural components [12]. A hair follicular unit, sometimes referred to as the pilosebaceous unit (PSU), is a naturally occurring structural unit of hair that resides within the dermis and the epidermis. It consists of 1–4 hair follicles, a sebaceous gland, and an arrector pili (Latin) muscle [7] (Figure 1). Longitudinally, the hair follicular unit is divided into three regions. The infundibulum refers to the uppermost segment of the hair follicle, which extends from the skin surface to the opening of the sebaceous duct. The middle portion between this sebaceous duct opening and where the arrector pili muscle contacts the hair follicle is the isthmus. The remaining lowermost part of the hair follicle and the hair bulb constitutes the inferior region [13]. The visible part of the hair shaft protrudes from the surface of skin. On the other end, embedded within the skin, each hair follicle is a highly compartmentalised structure made of epithelial components, such as the outer root sheath (ORS), the inner root sheath (IRS), the germinative matrix (GM), and the mesenchymal (or dermal) components, like the dermal papilla (DP) and the encapsulating dermal sheath (DS) [7] (Figure 2). Fundamentally, hair growth and follicle function are modulated by epithelial–mesenchymal interactions, as well as inter- and intra-compartmental communication [14]. It is important to mention that the ORS is a putative source of multipotent stem cells that can differentiate into several cell types such as keratinocytes and melanocytes. This stem cell pool is thought to reside in a distinct area, often called the bulge, positioned between the sebaceous gland ductal opening and the insertion of the arrector pili muscle [15]. At the bottommost follicular bulb region, the GM, which surrounds the DP, is composed of ‘transient-amplifying’ keratinocyte cells that rapidly divide and proliferate. As their volume increases, they push and move upwards and concentrically away, until they terminally differentiate and transform into a hardened material. Through a multi-step process of keratinisation and cornification, the hair shaft is formed and gradually emerges from the skin [7]. Composed primarily of keratin and macrofibrils, made up of rods of microfibrils woven together in a matrix, the hair shaft is another organisational structure consisting of the cuticle, the cortex, and occasionally the medulla. A medulla core is sometimes present in thick hairs. This is surrounded by the cortex, or cortical layer, which is the main structure of the hair shaft that determines the physical and mechanical properties of the hair fibre. Lastly, the outermost layer named the cuticle layer forms an overlapping scale-like barrier that shields against external physical and chemical damage [7,16] (Figure 2).
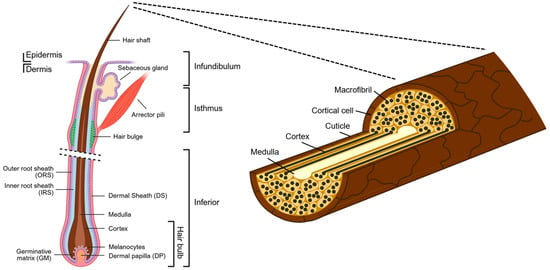
Figure 2.
Hair anatomy and hair structure. (Left) Schematic of a longitudinal cross-section of a human hair follicle, showing the hair structure arising from the dermis to its protrusion from the skin surface. Created in BioRender. Lim, Y. (2026) https://BioRender.com/xspkzwv (Right) The visible part of the hair, the hair shaft, consists of three layers—from innermost to outermost—the medulla, cortex, and cuticle.
2. Characteristics of Ageing Hair
2.1. Hair Greying
Greying of the hair, sometimes referred to as canities, is the most visible indicator of normal ageing. ‘Grey hairs’ refer to a mixture of pigmented and non-pigmented (‘white’) hairs on the scalp, or, in some cases, a single hair strand can display a gradual loss of melanin pigment from dark to light. The onset and progression of hair greying depend on chronological age, an individual’s health, and vary amongst ethnicity, gender, and genetics. Extrinsic environmental factors like climate, seasons, exposure to pollutants and chemical toxins, and lifestyle stress can equally affect hair pigmentation as well [17,18].
Keogh and Walsh first stated the 50-50-50 rule of thumb in 1965—whereby 50% of people exhibit 50% of grey hair by 50 years of age [19]. However, a more recent survey of healthy men and women volunteers revealed that a global range of only 6–23% of people experience 50% grey hair by 50 years of age [20]. This is likely due to a varying rate of hair greying amongst different ethnicities—beginning at an average of 34 ± 9.6 years in Caucasians, late 30 s in Asians, and 43.9 ± 10.3 years in Africans [18,20,21,22]. Generally, instances of hair greying arising before 20 years of age in Caucasians, before 25 years in Asians, or before 30 years of age in Africans is deemed to be premature [22,23].
The age of onset and rate of hair greying are largely dependent on ethnic and geographical origins, but irrespective of gender. Instead, hair greying in men and women differ in terms of the incidence of grey hair and pattern of hair greying [20,22]. Based on the previously mentioned 2012 survey, within a specified age range of 45 to 65 years, more men had grey hair and significantly higher greying intensity as compared to women [20]. Another Korean study reported that greying in men initially appears at the temples and sideburns, whilst grey hairs tend to develop first along the hairline or boundaries of the scalp in women [24]. Then, whether starting from the temporal or frontal regions of the scalp, grey hairs gradually spread to the vertex and parietal and lastly show up at the occipital region [18,25]. This varying rate and timing of hair greying, as well as other hair parameters, across the different parts of the scalp may be attributed to embryonic derivation, where the anterior scalp (frontal and temporoparietal areas) originates from the neural crest and the posterior scalp (consisting of the occipital area) is derived from the mesoderm [26].
The exact cellular and molecular mechanisms involved in human hair greying are not well elucidated, and the extent of genetic, metabolic, oxidative, and inflammatory contributions to greying remain unclear [25,27,28,29]. This likely arises from the insufficient understanding of physiological processes modulating human hair pigmentation [30]. Epidermal and hair follicular melanocytes may share a common origin and can be reactivated under certain circumstances [31], but their location, morphology, function, regulation, and susceptibility to chronological ageing are dissimilar [30,32,33]. Particularly, hair follicle pigmentation is closely coupled to the hair cycle [28,34] (Figure 1). Hair follicular melanocytes demonstrate an extraordinary capacity for the synthesis and storage of melanin during the anagen phase (stages III–VI). It is proposed that as few as 100 melanocytes per anagen scalp hair follicle can supply enough melanin to intensely pigment a hair fibre of up to 1.5 metres in length for 10 years [35]. In a human hair follicle, melanocytes and their precursors mainly reside at the follicular bulb matrix (above and around the tip of the dermal papilla; below the pre-cortical keratinocytes), the sub-bulge area of the ORS, and the bulge [34,36,37] (Figure 2). The bulbar melanocytes, together with the surrounding keratinocyte population, constitute the melanocytic or pigmentary unit (HFPU or hair follicle pigmentary unit). This unit, together with the melanocyte stem cell (MeSC) populations located along the hair follicle, contribute to hair fibre pigmentation, producing and transferring melanin to the cortical keratinocytes that continuously proliferate to form the hair shaft cortex, medulla, and cuticle [37,38,39].
The principal view is that hair greying stems from the reduced bulb melanin content and the depleted potential of the HFPU—as a combined result of reduced melanogenesis activity, aberrant melanin transfer to the hair shaft, and apoptosis of mature, differentiated bulbar melanocytes at the HFPU [28,40]. This phenomenon has been attributed to dysregulated antioxidant pathways and DNA damage brought about by exposure to environmental stresses like ultraviolet (UV) rays and pollutants, normal metabolism, inflammation, and genetic defects. Collectively, they trigger the formation of H2O2, as well as the overproduction and accumulation of free radicals and reactive oxygen species (ROS). Since mitochondria is a primary source of intracellular ROS, a mitochondrial “free radical theory of ageing” is commonly associated with hair greying [41]. Cumulative oxidative stress exacerbates apoptosis, causes defective oxidative stress protection, then leads to accumulated mitochondrial DNA damage that reduces bulbar melanocytes and causes depigmentation of the hair follicle [18,25,28,30,42,43].
Precise downstream mechanisms of how excessive oxidative stress and mitochondrial damage impact HFPU during human hair greying have yet to be identified. The process of melanin synthesis itself, or melanogenesis, inherently produces high ROS levels as a byproduct of a series of biochemical reactions involved in melanin production, such as the hydroxylation of tyrosine to L-DOPA (L-3,4-dihydroxyphenylalanine) and the oxidation of dopamine to melanin [28,30]. Previously, Van Neste suggested that the elongation rate of non-pigmented female hair follicle shafts was significantly faster than pigmented hairs when maintained in organ culture [44]. In an Asian cohort [45], hair shaft diameters were also comparatively higher in unpigmented hairs than pigmented hairs. Notably, elevated levels of keratins (KRTs) and keratin-associated proteins (KRTAPs), which form major components of the hair shaft and are essential for its mechanical properties and rigidity, were observed in these unpigmented hair bulbs. Microarray and RT-PCR analyses of the unpigmented and pigmented Asian hair bulbs showed that KRT6 (keratin 6), KRT14/16 (keratin 14/16), KRTAP4 (keratin-associated protein 4), and FGF7 (fibroblast growth factor 7) were associated with the induction of hair growth and were significantly upregulated in unpigmented hairs. Contrarily, FGF5 (fibroblast growth factor 5), a factor involved in the transition of the hair from the anagen to catagen phase and an inhibitor of hair growth [10], was downregulated in unpigmented hairs. On one hand, these findings reveal that a reduction in melanin-associated processes at the hair bulb may promote active growth by way of consolidating the follicle’s efforts to manufacture thicker fibrils at a faster rate. However, they also emphasise the high metabolic activity, redox, and genotoxic stress occurring in the hair follicle, which requires management by sophisticated antioxidant systems that include antioxidant enzymes, scavengers, etc. [25,28,30]. In addition to antioxidant metabolism in response to excessive levels of ROS, cells also recruit the DNA Damage Response (DDR) mechanism to detect and protect against the damaging effects of free radicals on DNA. One crucial component of the DDR is the Ataxia-Telangiectasia mutated (ATM) gene, which is a serine/threonine transducer kinase activated during oxidative stress. In mice, deficiency of ATM caused sensitisation and early differentiation of melanoblasts (a transient population of precursor cells derived from neural crest cells that migrate to develop into melanocytes), thereby leading to a diminished stem cell niche [46]. Correspondingly, when checked in scalp tissues collected from healthy donors, the incidence and expression level of ATM also correlated with the unpigmented status of human hair follicles [27].
Another viewpoint frequently discussed is the role of MeSCs in hair pigmentation. The MeSC population, which resides within the hair bulge–sub-bulge niche and adheres to hair follicle stem cells (HFSCs), as well as the intra-bulbar melanocyte progenitor cells, gives rise to mature melanocytes located in the hair matrix at the bulb [28]. Little is known about how the MeSC populations replenish these HFPU melanocytes, or about the cell-cycle entry and quiescence of the MeSC population during hair cycling. Based on murine models, modulation of MeSCs is critically dependent on the master regulator of melanocyte development—MITF (microphthalmia-associated transcription factor)—and its downstream target Bcl-2 (B-cell lymphoma 2) [47]. TGF-β (transforming growth factor-β) and Col17A1 (collagen XVII) expressed by surrounding HFSCs also have important roles in maintaining MSCs’ quiescence and immaturity [48,49,50]. Additionally, several other signalling pathways such as SCF/c-kit (Stem Cell Factor and its receptor c-kit), Notch, Raf and Wnt/β-catenin (together with the transcription factor Pax3), NFIB (Nuclear factor I/B), EDNRB (Endothelin Receptor Type B), and CXCL12 (C-X-C motif chemokine ligand 12) were identified to be essential for melanocyte development and MeSC homeostasis [51,52,53,54,55,56]. Maintenance of this MeSC reservoir is viewed as imperative, because the reduction in MeSCs via differentiation and/or apoptosis is hypothesised to provoke the ‘irreversible’ greying of hair [25]. Up until then, loss of pigmentation due to initial defects in the HFPU is still considered reparable if MeSC niches are still maintained [30]. Keeping in mind the fundamental differences between mouse and human models, several notable regulators of human hair follicle pigmentation are not as essential as they are in murine hair follicle pigmentation, and vice versa. Nevertheless, MITF-M, Sox10, Pax3, TYR (tyrosinase), and TYRP1 (tyrosinase-related proteinase 1) were reportedly reduced or absent in unpigmented human hair bulbs compared to pigmented hair bulbs [57]. A subsequent microarray study of human hair follicles (of varying pigmentation) further supported these observations and revealed a corresponding decrease in KIT (tyrosinekinase receptor for SCF) in unpigmented follicles [58,59]. Other theories include the involvement of aberrant DDR pathways (e.g., nucleotide excision repair (NER)) in the MeSC population and melanocytes that cause the reduction in melanin production capacity [60], as well as decelerated melanocyte migration from the MeSC ORS/bulge reservoir to the follicular bulb [18]. A recent publication conducted in mice further substantiates that the loss of hair fibre pigmentation during ageing is due to the defective migration of MeSCs into the follicular bulb matrix. Using long-term lineage tracing in vivo, researchers from the NYU Grossman School of Medicine demonstrated how MSCs are exceptionally plastic and dynamic, translocating actively between compartments of the developing hair follicle. They found that MeSCs increasingly get trapped in the follicle bulge of aged murine follicles, and their location renders these cells incapable of regenerating or maturing into pigment-producing melanocytes [61].
Taken together, the hair greying phenomenon is not monocausal but the outcome of a myriad of factors [30]. Beyond the complex network of controls and mechanisms that regulate human hair pigmentation, it is vital to also account for the functionality of the whole HFPU unit in conjunction with its unique microenvironment within the scalp skin (more details in Section 3). On a positive note, this multifactorial process presents opportunities where pigmentation of the hair follicle can be reversed or restored as long as MeSC and melanocyte populations of the hair follicle are not completely lost. Indeed, reversal of hair greying is perhaps not too inconceivable, considering the correlation between hair pigmentation patterns (HPPs—a single hair fibre that exhibits greying and reversal transitions) and psychological stressors [62], suggesting that management of psychological stress can pose as a potential avenue for controlling hair greying.
2.2. Hair Loss During Ageing
Much like hair greying, hair loss is another common occurrence that is part of the natural ageing process. While an individual typically sheds approximately 100 hairs a day, excessive hair loss leads to balding. This loss or absence of hair from areas of the body where it is expected to be found is medically termed as alopecia [8,11,63]. Alopecia can manifest as localised—specific areas on the skin (in patches) or diffuse—general thinning, partial or complete loss of hair, and can be temporary or permanent [64]. The most prevalent type of hair loss is androgenetic alopecia (AGA), although other alopecia types are also observed as well, such as alopecia areata, traction alopecia, telogen effluvium, scarring (cicatricial) alopecia, etc. [65]. Since senescent alopecia (SA) and AGA are both age-related hair loss conditions, we have chosen to focus on these two alopecia types in the subsequent paragraphs. Regardless of the form of alopecia, hair loss and reduced hair growth is caused by disruption to the hair growth cycle and/or damage to hair follicles due to various systemic and local factors like genetic predisposition, lifestyle and nutrition choices, hormone imbalances, medical conditions, and ageing [8,66].
Located at the base of the hair follicle is the DP, which consists of a specialised population of mesenchymal cells formed during hair follicle development (Figure 2). As an instructive niche, DP cells maintain an active communication with its precursors and the epithelial compartment—to direct keratinocytes in follicle growth and production of hair shaft, as well as to regulate hair shape, size, and colour [67,68,69]. It was first reported that the size of the hair follicular bulb (including both DP and GM) was correlated to the health of human hair follicles, suggesting that the hair follicle size and volume of hair fibre could be determined by DP size [70]. A subsequent study by Elliott et al., 1999 [71] demonstrated that the reduction in DP volume, due to a decrease in DP cell number, was associated with vellus hairs. More clearly, in the context of progressive hair loss, the size of the follicle and hair it produces was shown to gradually reduce in successive hair cycles until the hair follicle reaches miniaturisation [72]. Several studies have proposed that hair loss treatments involving IGF-1 (insulin-like growth factor 1) and FGF-7 upregulate Wnt/β-catenin signalling in DP, thereby stimulating anagen entry and hair growth. Another well-studied growth factor, EGF (epidermal growth factor), also plays an important role in regulating hair follicle growth. However, recent findings indicate that EGFR signalling downregulates Wnt/β-catenin signalling to delay differentiation and lineage commitment, thereby helping to coordinate normal hair follicle formation [73]. Alternatively, because the telogen phase is more than just a resting state—it is a crucial period where the DP compacts in preparation and awaits signals that reactivate hair regeneration—attention has been focused on transitions to and from the resting telogen phase as part of successful maintenance of DP for hair growth [67]. Other emerging solutions include the inhibition of senescence of hair follicular DP cells—for example, the use of veratric acid [74]—as well as senolytic treatments with dasatinib and quercetin, which specifically target and effectively reduce the number of senescent DP cells and restore inductive properties of hair follicles, thereby allowing them to regenerate and promote new hair growth [75].
Epithelial–mesenchymal interactions between the DP and HFSCs are critical for powering the cyclic growth of the hair follicles and maintenance of follicle morphogenesis [76,77]. Most HFSCs reside within the bulge region of the ORS, which is located between the opening of the sebaceous gland and the attachment site of the arrector pili muscle (Figure 2). These bulge HFSCs are a multipotent adult stem cell population that have high proliferative potential. They harbour the ability to generate several epithelial lineages of the skin—in addition to new hair follicles, they are also able to give rise to the epidermis and sebaceous glands during wound healing [78,79]. Because the hair follicle is an integral component of the skin, the growth and activity of HFSCs are regulated by various neighbouring cells, such as the DP, adipocytes, immune cells, and nerves, which together form a specialised microenvironment called the HFSC niche [80]. Within this HFSC niche is the constantly opposing activating and inhibitory signals, mostly driven by the activating Wnt/β-catenin signalling and the inhibitory BMP (Bone Morphogenetic Protein) signalling. Transition between the activated state or quiescent state of HFSCs is dependent on the final readout summation of the counteracting signals. Other signalling pathways that further add to the HFSC niche’s complexity and heterogeneity include Hedgehog (Hh), Notch, PI3K/AKT, RAS/MAPK, TGF-β2, Foxp1, and oncostatin M [76,80,81,82]. Upon stimulation, quiescent HFSCs become activated, initiating the onset of anagen that drives the start of a new hair cycle.
Mostly demonstrated in murine studies, many changes have been noted in hair follicles during ageing. For example, the lengthening of the telogen phase whilst the anagen phase is shortened, extended dormancy and inability to re-enter the hair growth cycle, as well as the miniaturisation of hair follicles, are cited to result in shorter and more scattered hairs and eventually hair loss [8,81,83,84,85]. While it is still uncertain if the number of HFSCs varies with age, there is a general agreement that the function of these HFSCs diminish with advanced age [81,86,87,88]. In-depth in vivo fate analyses revealed that aged HFSCs are cyclically eliminated from the murine skin via the proteolysis of COL17A1/BP180 by neutrophil elastase in response to DNA damage. COL17A1 plays an essential role of maintaining HFSC health and function, and its depletion led to the stepwise miniaturisation of the murine hair follicles and eventual hair loss. In the same study, gene set enrichment analysis of telogen hair follicles between young and aged mice also showed that Wnt signalling was less activated in aged HFSCs [83]. Further supporting this view, Tiwari et al., 2021 [89] showed that Wnt target genes—Axin-2, Lef-1, Lgr-6, and c-Myc—were downregulated in HFSCs upon ageing. Nuclear localization of β-catenin was also markedly decreased, and the canonical Wnt signalling was antagonised by non-canonical Wnt5a-Cdc42 signalling in aged HFSCs. On the other hand, it is proposed that the changing activity and loss of function of aged HFSCs supersedes the importance of the HFSC population size. Murine aged HFSCs have been shown to grow slower in vitro, exhibiting decreased activity and capacity to form colonies as compared to young HFSCs [84]. While there is yet an established, direct causal relationship between alopecia occurrence and senescent HFSCs [86], the abovementioned studies provide hints at potential links—hence spurring stem cell-based therapy approaches focused on replenishing HFSCs or reactivating HFSCs with adequate signals and environments. From the transplantation of multipotent stem cells, derived from adipose tissues, bone marrow, umbilical cord, and hair follicles from unaffected scalp areas, to the application of stem cell-derived conditioned media and stem cell-derived exosomes, clinical studies have shown promising results with improved efficacies, albeit with their respective limitations [90]. Ultimately, in addition to the HFSCs’ actual activity, its niche and microenvironment have an essential role in influencing HFSCs and should not be overlooked [88].
Senescent/Senile Alopecia and Androgenetic Alopecia
Senescent alopecia or senile alopecia (SA) is described as a non-androgen-based, diffused hair loss that can involve all hairs of the human body (not only the scalp). It is thought to affect individuals above 50 years of age with no family history of patterned balding [91]. Separately, the more widely known AGA, or, male or female-patterned alopecia, is regarded as a genetically predetermined disorder, which is caused by an excessive sensitivity to androgens. Its prevalence differs across race and gender—possibly affecting up to 85% of men and approximately 40% in women. In some cases, hair thinning may begin as early as after puberty in adolescents [92,93]. To date, there still exists much debate to whether SA or AGA represent two distinct entities [94]. Various studies like Deng et al., 2023 [86] and Whiting, 2011 [91] have indicated significant overlap between SA and AGA in terms of reduced total hair counts, reduced hair follicle size and hair fibre diameter, decreased anagen-to-telogen ratios, and higher terminal-to-vellus hair ratios. Indeed, the incidence and prevalence of AGA increases with age. However, the diagnoses of AGA are not limited to older people. In the first comparative histopathology study of SA and AGA, Kligman, 1988 [95] reported that, while SA was characterised by a modest reduction in hair follicle size, AGA was observed to be associated with hair follicle miniaturisation, inflammation, and fibrosis. The absence of fibrosis in SA was a likely explanation why hair growth may still be stimulated by pharmacological means. A more recent study of gene expression profiles of scalp biopsies of SA and AGA male subjects, as well as control group (with no visible hair thinning), revealed that both phenotypes represent different entities [96]. As expected, hair development and function were the most significant physiological parameters altered in both SA and AGA. Notably, there was an upregulation of the androgen receptor (AR) in AGA, but not in SA. This corroborates previous studies that have demonstrated that genetic variability in the human AR is the main prerequisite of the early onset of AGA [97,98]. Additionally, unique to SA, genes involved in transcription (FOS, FYN, JUN, JUNB, MYC, NAB1), growth factors (CTGF, TGFα), and mitochondrial function (JAK2, PRKD3, AK2, TRAP1, TRIO, ATP12A, MLL4, STK22B) were significantly downregulated, whilst expression levels of oxidative stress and inflammatory response genes were upregulated [94,96].
With most studies conducted in AGA subject cohorts, major developments have been made in understanding androgen metabolism and how androgen-regulated factors operate through the DP [94]. This has led to the development of widely prescribed 5α-reductase inhibitors, which are known modulators of androgen metabolism, such as finasteride and dutasteride. However, there remains limited success with these hair loss treatments [99]. The primary concerns are that these oral medications may cause persistent adverse effects and that there is a lack of studies examining their safety and effects in females with AGA [100]. Additionally, in hair follicles of AGA men aged between 18 and 30 years, there were significantly higher levels of 5α-reductase types 1 and 2 receptors and ARs in the frontal follicles compared with the occipital follicles. This finding was also almost a two-fold increase compared to hair follicles collected from SA men [101]. This in part explains why the efficacy of using 5α-reductase inhibitors could decrease in men at advanced ages [94]. Another well-known treatment for AGA in both men and women is minoxidil. Initially developed as a hypertension medication to treat high blood pressure by dilating blood vessels, it was unexpectedly found to also promote hair growth, thereby driving its later development as a hair growth solution. Mechanisms of how minoxidil on the hair follicle are proposed to be multifarious and not fully understood, although animal studies have shown that topical minoxidil induces premature entry of resting hair follicles into anagen and prolongs the anagen phase, with a potentially similar mode of action in humans. Moreover, minoxidil is described to stimulate vascular endothelial growth factor (VEGF) expression, cell proliferation, and prostaglandin synthesis, whilst inhibiting collagen synthesis in various skin and hair follicle cell types [102,103]. Notably, VEGF has an important role in angiogenesis by promoting the formation of new blood vessels and ensuring an adequate supply of blood flow to the hair follicles. Overall, topical minoxidil is generally well tolerated, with more common side effects like skin irritation and itching. Yet a notable drawback is that it must be used indefinitely to provide continuous support and maintenance of any hair regrowth experienced [104].
It is important to also consider other potential hair loss factors like oxidative stress, follicular microinflammation, and fibrosis [94]. In assessing the effect of topical application of linolein hydroperoxides on murine skin, which causes free radicals that induce early catagen, there was increased apoptosis of murine hair follicle cells. Moreover, cultured human epidermal keratinocytes subjected to linolein hydroperoxides also showed an upregulation of apoptosis-associated gene expression [105]. In another study, isolated DP cells from AGA scalps grew significantly slower in culture than DP cells from non-balding scalps. The authors demonstrated that the loss of proliferative capacity in balding DP cells may be related to downregulated levels of proliferating cell nuclear antigen and Bmi-1, along with corresponding enhanced expressions of senescence-associated β-galactosidase, p16INK4a/pRb, and other markers of oxidative stress and DNA damage such as heat shock protein-27, superoxide dismutase catalase, ATM kinase, and Rad3-related protein. In particular, the association of cultured balding DP cells with increased p16INK4a/pRb implies a heightened sensitivity to environmental stress [106]. Beyond determination by genetics, progressive hair loss is equally susceptible to changes brought about by chronological ageing and hormones, as well as external environmental, dietary, and lifestyle factors. Furthermore, several other studies have proposed that a chronic, low-level, microscopic follicular inflammation, referred to as ‘microinflammation’, may contribute as a cause of hair loss [107,108,109]. This should not be confused with the classical, more destructive inflammatory process involved in scarring (cicatricial) alopecia disease [107]. Morphometric studies conducted by Whiting, 1993 [110] provided early evidence that AGA scalps with significant microinflammation and fibrosis had reduced likelihood of hair regrowth in response to treatment. It is unclear how inflammation is generated around the hair follicle, since it is likely a consequence of multiple factors triggered by a primary event. In more deteriorating conditions of hair loss, it has been suggested that there could be growing clusters of perifollicular macrophages and cumulative physiological degeneration of selected hair follicles, where malfunctioning or defective follicles are eventually removed—similar to a programmed organ deletion process [111].
2.3. Changes in Morphology and Properties of Hair During Ageing
Beyond the loss of pigmentation in hair fibres during ageing, greying hair is observed to be frequently coarser, wirier, and more unmanageable compared to its pigmented equivalent (Figure 3). The absence of melanin accentuates changes in the chemical and physical properties of the post-pigmented hair fibre [35,112]. More specifically, hair fibre properties like diameter, ellipticity, and fibre elasticity vary according to normal chronological ageing [94]. It has long been recognised that there are many underlying differences in fibre properties between ethnicities; hence, it is important to also expect ethnic-associated variations during hair ageing [42].
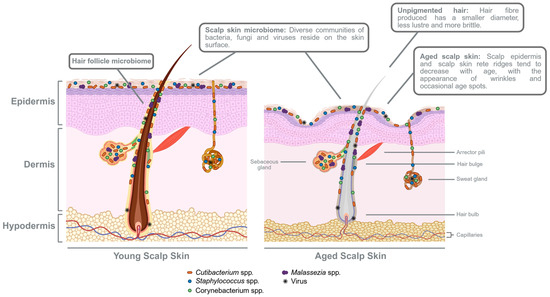
Figure 3.
Comparison of the hair follicle and scalp skin between healthy young and aged skins. With increasing age, scalp skin thickness decreases, while TEWL and blood microvasculature are also reduced. Other observations include diminished rete ridges and sebaceous glands, thus resulting in lower sebum secretion. Compared to the hair follicle in the young scalp skin, a less pigmented hair fibre is produced in the ageing hair follicle and appears to have a smaller hair diameter, less lustrous, and more susceptible to breakage. Healthy scalp skin hosts unique diverse communities of bacteria, fungi, viruses, and even mites that reside on the surface, within the scalp skin epidermis, and along the hair follicle. The scalp microbiota contributes to the protective skin barrier and influences its immune defence. Thus, changes in diversity and composition with age potentially influences hair and scalp health. Created in BioRender. Lim, Y. (2026) https://BioRender.com/keec1cq.
Age-associated hair diameter changes appear to have a significant contributory effect on the overall perception of hair thinning and are not solely due to reduced hair density [113]. Past studies have so far indicated a general trend towards hair shaft diameter decrease with increasing age. In Japanese and Caucasian male study cohorts, hair shaft diameter increases and peaks during the late teenage years. After which, from approximately 25 years of age to late 40s and up to 89 years, the hair shaft diameter steadily decreases [114,115,116]. Contrary to men, hair shaft diameters are not linearly proportional to age in women. Rather, both Caucasian and Asian women tend to follow a curvature that reaches a maximum at about 40–46 years, before it starts declining [113,114]. In addition, physiological and hormonal effects as women approach and experience menopause are suggested to also influence changes in female hair diameters. For example, a comparison between premenopausal and postmenopausal Caucasian women reported a significantly smaller diameter of the scalp hairs of postmenopausal women [117]. However, if hormonal level changes (throughout menopause) could be monitored together with scalp hair diameters, this physiological effect would be further validated. Recently, X-ray diffraction studies have suggested that the hair shaft shrinking phenomenon does not simply consist of producing a smaller shaft diameter but also involves the implementation of an ‘aged hair program’ that begins in the hair follicle. Age-associated thin hairs were found to exhibit changes in the expression of hair keratins and keratin-associated proteins, as well as alterations in the intermediate filament structural organisation that forms the hair cortex—the middle layer of hair shafts known to confer strength, texture, and colour. Furthermore, the thinner hairs exhibit a decreased hair ellipticity, fewer cuticle layers, and lower frequency of a medulla, the innermost core of the hair fibre that is usually absent in finer hairs. Collectively, these modifications could have resulted in modifications in mechanical properties such as increased rigidity, reduced viscosity, and decreased water diffusion coefficient that make hairs more susceptible to breakage and damage [118].
A hair fibre’s ability to stretch (elastic modulus or Young’s modulus), bend (bending modulus), and twist (torsional modulus) before permanent deformation has been shown to be proportional to hair fibre diameter [119,120,121]. As the hair shaft diameter reduces with age, the fibre’s ability to withstand these three stresses correspondingly decreases. A comparison among three different age groups—children (1–10 years), young adults (25–35 years), and the elderly (50–60 years)—demonstrated that elastic modulus and hardness were higher in young adults than in the elderly. This may in part explain the decreased tactile perception of hair with advanced age [122]. Considering the variations in gender, Caucasian and Asian men start to observe a deterioration in the mechanical properties of their scalp hairs in their 20s, while Caucasian and Asian women only exhibit signs of weakening mechanical properties in their early 40s [123].
Persaud and Kamath’s development of a single hair fibre torsional pendulum method additionally suggests that reinforcement of the hair cuticle layers has a more significant role towards strengthening torsional resistance than in elasticity [121]. This presents as another contributory factor to how hair’s physical properties can change. Indeed, there is a gradual loss of hair cuticles with age, as Plott et al., 2020 [124] found a decreased abundance of cuticular proteins with increasing age. This corroborates with an earlier key study by Thibaut et al., 2010 [125]. Given the known variations in hair cuticle characteristics across ethnicities, it is conceivable how the hair fibre’s ability to resist mechanical stresses at advanced ages may differ across populations.
Hair fibre curvature also has an important effect on the appearance of hair. In a study group of Japanese females, aged from 10 to 70 years, it was found that the decrease in hair lustre corresponded to irregular increase in hair fibre curvature during ageing [126]. This is likely due to an inhomogeneous amino acid deposition of hair keratins that varies the internal structure of the outer and inner regions of the fibre, giving rise to the curvatures [127]. Altogether, this lack of alignment within an assembly of hairs shows up as frizziness or a reduction in hair lustre. Moreover, notable increases in cysteic acid residues and decreases in cysteine found in grey hairs imply a weakening of the hair fibre’s strength and structure, which causes heightened sensitivity to weathering and reactivity to reducing and oxidising agents [33,128]. Lastly, without the photochemical protection due to loss of hair melanins, hair proteins progressively become degraded and lost, altering the composition of hair shaft layers. Caught in a negative cycle, grey or non-pigmented hairs increases sensitivity to ultraviolet radiation (UVR) [129].
Although hair lipids form a small part of the composition of hair, they are as essential as hair proteins, minimising moisture loss, providing a permeability barrier, and maintaining hair integrity. Besides its physical properties, lipid metabolism contributes by modulating modifications to signalling proteins, like Hhs and Wnts, which are necessary for hair follicle morphogenesis and hair growth cycling [130,131]. The two major sources of hair lipids are the follicular bulb matrix cells and follicular sebaceous glands (Figure 2; left). Amidst ongoing hair shaft production and melanogenesis, the hair bulb matrix also produces integral lipids, which are incorporated during keratinisation and the formation of the IRS. These lipids are embedded within the keratinised cell membranes, providing structural integrity and protection like a lipid envelope [132,133]. Overall, human scalp hair lipids mainly comprise fatty acids, wax esters, ceramides, squalene, and cholesterol. Together, they account for about 0.7–1.3% of the total chemical content of hair [94,131,132]. It was reported that because Asian hairs have more integral hair lipids, they exhibited significantly lesser damage compared to Caucasian and African hairs after exposure to UVR irradiation [131]. Nevertheless, the overall lipid content of the hair cuticle, cortex, and medulla has been shown to decrease during the pigmented-non-pigmented hair transition and with advancing age [134]. Importantly, 18-methyl eicosanoic acid (18-MEA), the most abundant fatty acid found in hair, is covalently bound to proteins of the cuticle cell outer layers via thioester bonds. This constitutes the fatty layer, sometimes referred to as the F-layer, which is responsible for the surface and tactile properties of the hair fibres and gives the hair a smooth, shiny, and silky texture [134,135]. 18-MEA in hair strands is easily damaged and lost through dyeing, perming, and other chemical processes and external factors. Through these daily stresses, the amount of 18-MEA naturally decreases with age, with studies showing more pronounced loss of total 18-MEA content starting from age 50 [136]. These hair lipids are distinct from the sebum production of the sebaceous glands that secrete a waxy, oily substance to lubricate emerging hair shafts, conferring shine, smoothness, and softness [137]. The amount of sebum production is dependent on the size of sebaceous glands, which is generally known to be low before puberty, rapidly increases at puberty, remains at relatively high levels, and declines at 45–50 years of age [138]. Decreasing exogenous and endogenous lipid contents of the hair cuticles, cortex, and medulla with advancing age, including diminishing 18-MEA levels and de novo dihydroceramide synthesis, causes unpigmented hair fibres to have an overall lower capacity of water absorption and higher water diffusion, thereby leading to greater stiffness and permeability [125,139,140,141,142]. This causes unpigmented hairs to be more susceptible to brushing and mechanical stresses, further accelerating loss of the cuticle barrier that prompts the occurrence of more hair breakage [143,144].
3. Ageing Scalp Skin
3.1. Age-Related Changes in the Scalp Skin
While variations in hair volume, density, pigmentation, and texture have been extensively studied and reported during chronological ageing, age-related changes in the scalp skin are relatively less understood. Similarly to other skin sites, the scalp skin likely evolves with age. On the other hand, it is prudent to first note the functional distinctions between the stratum corneum (SC) of the scalp skin, the face, and other sites of the body. In a study of Japanese patients with alopecia areata and AGA [145], physiological differences in water barrier function (TEWL values), skin surface hydration, and skin surface lipids were found to vary across the scalp, cheek, and volar forearm skin test sites. Moreover, superficial corneocytes of the scalp skin appeared significantly larger in size than the cheek skin, but smaller than the volar forearm skin. Given that corneocyte size is inversely correlated with epidermal proliferative activity, the epidermal proliferative activity of the scalp skin, with an estimated turnover rate of about 10 days, is hence slower than that of facial skin. With the growing prominence of scalp ageing and the acceleration of the ‘skinification’ of the hair initiative in recent years, the focus has now shifted to deeper understanding of, and better care suited for, the hair source—that is, the scalp skin—in order to stimulate or maintain healthy hair growth [146,147,148].
Conceivably, the intertwined structural arrangement of the scalp skin and hair fibres implies an interdependent relationship between both components. While the hair strands protect and shield the scalp from mechanical stresses and UVR, the scalp, conversely, serves as an incubatory environment that supports hair growth [94,149]. Continuous cycling of the hair follicles requires considerable remodelling of the scalp dermal environment. In fact, a transplant study in mice performed by Cao et al., 2016 [150] provided evidence of the importance of the hair follicle dermal environment in growth and maintenance of hair follicles. Young and aged murine follicles transplanted into young host nude mice began to establish blood vessel connections in 2 weeks and promoted hair shaft growth by the 4th week, implying that the young host dermal environment could have rejuvenated the aged follicles through efficient angiogenesis and that ageing was potentially reversible. Conversely, there was no regrowth of hair shafts observed when both young and aged follicles were implanted into aged host nude mice. Further substantiating this concept is an earlier referenced study by Ge et al., 2020 [88], who demonstrated how aged murine HFSCs could be reinvigorated when transplanted into the neonatal dermis, but young HFSCs are not well supported by the aged dermis and failed to regenerate hair follicles.
There are apparent differences in the physiological characteristics amongst androgen-sensitive (vertex) scalp areas, androgen-insensitive (occipital) scalp areas, and non-balding scalps, which indicate underlying alterations of the scalp skin structure and barrier function upon hair loss. While TEWL showed no significant changes, the vertex scalp of AGA male subjects exhibited lower SC hydration level and skin surface lipid content, both of which positively correlated to the severity of hair loss [145,151]. Two treatment studies conducted by The Procter and Gamble Company (P&G) directly demonstrated that the scalp condition can influence hair growth and physical attributes of post-emergent hair fibres. Likewise, this impact to the hair cuticles was reversible upon restoration of scalp skin health [149,152]. Further supporting this notion, earlier published epidemiological studies comparing healthy individuals and patients with psoriasis, dandruff or seborrheic dermatitis, and atopic dermatitis showed that these patients exhibited premature hair loss, as well as thicker cuticular edges, increased roughness and surface pitting, and decreased shine of hairs from diseased scalps [94,153]. Interestingly, these observations of perturbed hair cuticles were suggested to resemble hair cuticles of the chronologically aged; thus, hair produced from unhealthy scalps can be referred to as ‘prematurely aged’ hair [125,153].
One of the first few studies providing an overview of the changes in male and female scalps across ages [154] documented how the female scalp dermal and hypodermal thicknesses evolved with age—achieving maximum thickness at 35 years, declining until 70 years, and then thickening again by 85 years of age. Contrastingly, the male scalp dermis reaches maximum thickness at 55 years and decreases until 80 years of age. In general, the scalp epidermal thicknesses appeared to differ with age, but only total scalp thicknesses displayed both gender- and age-related variations. While probing into the biophysical characteristics of healthy Caucasian women [155], it was reported that Caucasian women of an older age group (mean age: 62 ± 2 years) exhibited significantly lower values of TEWL and decreased scalp surface temperatures at the scalp vertex, while there were no notable changes in the sebum and SC hydration levels or SC thicknesses compared to a younger age group (mean age: 30 ± 3 years). Despite no significant differences in Caucasian female scalp SC thicknesses, the thickness of the scalp epidermal layer was found to significantly reduce with age. In comparison, a similar study conducted in a Korean population evaluated potential differences between two age groups—20s (mean age: 24 years) and 50s (mean age: 54 years)—in both females and males [156]. As expected, all participants in the 50s age group had significant decreases in hair densities when compared to the 20s age group. However, it was discovered that there were more changes in scalp biophysical parameters between the 20s and 50s age groups of women than the 20s and 50s age groups of men. For instance, women from the 50s age group had significantly lower sebum output levels, increased skin firmness, increased degrees of yellowness and redness, and reduced pH levels at the vertex and temporal scalp regions. Contrarily, men in the 50s age group showed only reduced TEWL at the scalp occipital and more pronounced yellowness than men from the 20s age group. One notable similarity that both genders shared was a significant decrease in microvasculature of blood vessels (not lymphatics) in the aged scalp skin [157]. In view of these studies, it can be deduced that there are diverse findings on the physiological changes in the aged scalp for both men and women. Whether due to gender or ethnic differences, or to the specific selection of scalp sites or age range, certain structural and physiological characteristics of the ageing scalp mentioned—such as decreased TEWL, lower sebum secretion, a thinning epidermal layer, and reduced blood microvasculature—resemble skin ageing at other anatomical sites [145,154,156,158,159]. These differing characteristics are depicted in the summary visual comparing healthy young and aged human skin (Figure 3).
A more recent histochemistry and immunohistochemistry study [160] examining scalp biopsies of Caucasian females aged between 19 and 81 years confirmed the reduction in epidermal thicknesses and particularly highlighted decreases in scalp skin rete ridges and sebaceous glands, and thus lower sebum secretion during ageing. Diminished rete ridges were similarly observed in the scalp skins of aged Korean men, although their epidermal thicknesses remained unchanged and dermal thicknesses decreased with age instead [161]. In greater detail, Williams et al., 2021 [160] revealed how the papillary reticular boundary of the younger female scalp skin (below 40 years of age) was more indistinguishable than the older female scalp skin (above 40 years of age), which displayed delineation of the papillary dermal architecture from the reticular dermis. This is in combination with significant changes in the expressions of ECM (extracellular matrix) proteins, including podoplanin (PCPN), versican (VCAN), and matrix metalloproteinase 1 (MMP1); papillary collagen organisation and deposition; as well as an elevated cartilage oligomeric matrix protein (COMP) level (a dermal sheath protein that facilitates fibrillar collagen assembly)—changes that were uniquely detected in the proteomic analysis of the aged dermal fibroblast culture secretome. These changes in the hair dermal environment and function appear to reflect the characteristics of solar elastosis, which imply that the female scalp dermis is equally susceptible to UV-induced photoaging [160].
Solar, or actinic, elastosis occurs when there is accumulation and disorganisation of abnormal elastotic fibres at the middle and upper dermal layers, which results in tissue stiffness and thickening [162,163]. In addition, upregulated MMP1, MMP2, and MMP3 levels in aged interfollicular dermal fibroblast and hair follicle dermal sheath cell cultures could also contribute to increased collagen degradation, as has been previously established in skin ageing [160,164,165]. There was also an overall higher prevalence of miniaturised hair follicles, resulting in markedly lesser hair follicles embedded within the dermal adipose tissues of aged scalp skins [160]. In human scalp skin, the bulbs of healthy, terminal anagen hair follicle typically penetrate the dermal white adipose tissue (dWAT) layer of the hypodermis [164]. These differences have also been included in Figure 3. It remains to be determined how the composition and volume of dWAT changes during scalp skin ageing, or how the scalp dWAT alters with human hair follicle cycling. But the significance of adipocytes in hair growth is gaining traction when treatments of autologous adipose-derived stromal vascular cells (ADSVCs) significantly improved alopecia areata symptoms by increasing hair densities and diameters in both men and women [166]. Additionally, another study demonstrated how dWAT could be modulating human hair growth and pigmentation through the hepatocyte growth factor (HGF)/c-Met signalling pathway [167]. Recalling the importance of HFSCs in hair growth, HFSC activity has also been associated with dWAT thickness, as well as regulated by mature adipocytes and immature adipocyte precursor cells in dWAT [81,168,169]. Also found within the dermis and hypodermis of the skin, the remodelling of vasculature and lymphatic capillary networks is in dynamic crosstalk with HFSCs and can affect one another during hair follicle regeneration [81,170,171,172].
The final notable feature is the increased areas of inflammatory infiltrate under the papillary dermis and around the sebaceous glands in aged Caucasian female scalp skin [160]. This indicates the presence of a low-grade inflammation in the ageing scalp skin, which has also been identified to be closely associated with skin ageing [173]. Frequently linked to increased levels of pro-inflammatory cytokines, proteolytic enzymes, and ROS, oxidative stress and free radical damage to DNA, proteins, and lipids within the interfollicular environment possibly elevate until antioxidant defences are exceeded, further driving the production of collagen-degrading MMPs [164,174]. Collectively, these documented changes present an aged hair phenotype. They accelerate collagen degradation and fibril fragmentation until it gives way to a physically modified interfollicular environment that hinders the active movement of the hair follicles throughout cycling and remodelling [160,164].
Similarities between dermal ageing and scalp ageing have spurred additional postulations that senescence may also be correlated with decreasing cell–cell interactions within scalp environment [164]. Single-cell transcriptomic analyses of young (28–37 years old) and aged (54–86 years old) healthy Caucasian inguinal iliac skin showed partial loss of dermal cell identities, reduced dermal fibroblast ‘priming’, and thus a corresponding decrease in predicted interactions between the dermal fibroblasts and other skin cell types [175]. Such loss of cellular identity during ageing has been previously described in mice [176]. With regard to the ageing scalp, a lack of interactions between the DS, DP, and undifferentiated bulb matrix keratinocytes can conceivably influence the growth and maintenance of the hair follicle. Furthermore, another murine study found that ageing weakened the commitment of the hair follicle epithelial bulge cells to the hair follicles and instead favoured the alternative differentiation into epidermal keratinocytes [83]. With additional hypothesised evacuation of the hair follicle DS cells as observed in alopecia patients [177], the continuous depletion of both follicular stem cell niche and DS population may jointly lead to age-related hair follicle miniaturisation. Taken all together, while much of the underlying structural, organisational and functional changes during scalp ageing still require complete elucidation. It is important to realise that the health and condition of the scalp skin can directly affect growth and homeostasis of hair follicles.
3.2. Scalp Skin Microbiome and Ageing
The human scalp is described as an anatomical area bordered anteriorly by the face, and laterally and posteriorly by the neck. It consists of soft tissue layers that envelope the cranium, extending from the supraorbital foramen margins to the occipital protuberance and superior nuchal lines [178]. The scalp skin is commonly characterised by high densities of hair follicles and sebaceous glands, which create a unique microenvironment of temperature, pH, moisture, and sebum content, providing a rich and conducive climate for microorganisms [94].
In general, the scalp skin is often described to exhibit a comparatively lower microbial diversity. The healthy scalp microbiome is mainly composed of particularly high abundances of Cutibacterium spp. (predominantly C. acnes), Staphylococcus spp. (majority being S. epidermidis), and Malessezia spp. (mostly made of M. restricta and M. globosa), alongside lesser abundant species such as Corynebacterium spp., Streptococcus spp., Acinetobacter spp., Prevotolla spp., as well as Ascomycota (Acremonium spp., Didymella bryoniae), Coniochaeta spp., Rhodotorula spp., and other Basidiomycota (like Cryptococcys liquefaciens and C. diffluens) [179,180,181,182]. These proportions vary across populations from different countries and genders. For example, a lower representation of the Cutibacterium spp. has been observed in scalp colonisation in males [183]. Typical scalp microbiome sampling methods include skin swabs, pore and tape strips, as well as cyanoacrylate gel biopsies [179,184]. These techniques have helped identify resident skin microbiome communities; however, they may fail to sample the deeper regions of the hair follicle epithelium and are unable to distinguish between hair follicular and skin microbiomes [179,184,185,186,187]. Alternatively, plucking hairs, as a means of sample collection, causes the detachment of the essential lower hair follicle mesenchyme from other components of the hair follicle epithelium and mesenchyme [188]. Studies examining the deeper, anaerobic regions of the scalp hair follicles are scarce. An early study by Matard et al., 2012 [189] captured the presence of bacterial biofilms, morphologically resembling C. acnes, below the infundibulum, by field emission scanning electron microscopy (FESEM) and confocal laser scanning microscopy (CLSM). More recently, another study coupling laser-capture microdissection (LCM) with 16S rRNA gene sequencing [184] additionally revealed variations in taxa abundances depending on the hair follicular compartments—namely, the infundibulum, isthmus, and the inferior sections (Figure 2). Interestingly, the isthmus and inferior segments were shown to share more similarities in identified genera than with the upper section of the hair follicle. The observed microbiome differences likely substantiate the follicular compartmental variations in terms of availability of nutrients, metabolites, antimicrobial peptide (AMP) production, and human hair follicle immunological characteristics, as previously reported, which in turn favour the growth of specific bacterial species [190,191]. With the hair follicle known to enjoy an immune-privileged status within the skin, there is growing evidence suggesting how microbial colonisation and dysbiosis at different follicular compartments, like the stem cell niche, can adversely affect hair follicle growth and cycling. Some examples of hair follicle-associated diseases that have been linked to specific anatomical areas of the hair follicle include acne vulgaris and hidradenitis suppurativa, which are mostly associated with the infundibulum, while alopecia areata is associated with the hair bulb region [192,193].
Compared to studies profiling the skin microbiota on the forehead, cheeks, forearms, axilla, inguinal creases, and feet, there are limited investigations on the human scalp, and even lesser reports on scalp skin commensals during ageing. Like various body sites, the scalp skin gradually experiences a reduction in sweat, sebum production, and immune function over time, leading to significant modifications in skin surface physiology like pH, SC hydration levels, decreased sebum content, and lipid composition [160,180,194,195,196]. These biophysical modifications may bring about changes in the skin ecology that influence growth of the resident skin microbiota. In a comparison between younger (21–37 years of age) and older (60–76 years of age) healthy Japanese women [181], the authors proposed that age-related modifications of the skin microbiome are skin-site dependent. Out of the four skin sites tested, the scalp exhibited the most significant difference in OTUs (Operational Taxonomic Units) or species numbers (α-diversity) between the younger and older cohorts, in contrast to the cheek, forehead, and forearm sites. Notably, while Cutibacterium still remained the most abundant genus on the scalp, previously minor species in the younger scalp, such as Acinetobacter, were significantly increased in the older scalp microbiomes. Their findings further identified normal oral cavity bacteria in the higher microbial diversity of the older female scalp microbiomes as well. This observation of a microbial signature in aged skin was intriguing given that the prevalence of oral cavity bacteria had also been reported in the skin microbiomes of atopic dermatitis patients, of which the mean age was 23.1 years [197]. Just as scalp microbiome diversity is correlated to decreased sebum secretion in aged skin [180], excessive sebum production at the scalp occipital region has been shown to be associated with the onset of sensitive scalp. Significant increases in free fatty acids, cholesterol esters, and squalene levels in the occipital regions of sensitive scalp subjects, compared to non-sensitive scalp subjects, have changed the scalp’s environment and pH, with a preference for Cutibacterium and supporting a lower microbial diversity [198]. However, it is not shown in this study which strains (pathogenic or non-pathogenic) of C. acnes were found to be elevated. Throughout the ageing process, ROS levels accumulate, and reduced presence of enzymatic and non-enzymatic protectors contributes to oxidative stress [199]. It is thus important to point out that the prevalence of Staphylococcus (particularly S. aureus) and Malassezia, which result in skin microbiome perturbation, has previously been reported as a source of oxidative stress [200,201,202]. Correspondingly, oxidative stress has in turn been shown to negatively impact the pre-emergent hair condition that causes pre-mature hair loss [94,203]. Striking the right balance between the skin microbiota and host environment (i.e., strain-specificity and context-dependence) is ultimately key, as in converse, there is new, promising data indicating that the commensal microbiome (also including S. aureus) is able to promote regeneration of hair follicles by stimulating keratinocyte HIF-1α (hypoxia-inducible factor 1-α) signalling and glutamine metabolism in the murine wound-induced hair follicle neogenesis (WIHN) model [204]. While the WIHN phenomenon is not evident in human skin, commensal microbes, such as S. aureus, are able to accelerate wound healing in mouse and human [204], which suggests an overlapping mechanism. Collectively, it appears that the ageing-related alteration of skin microbiota may be more correlated to changes in the skin’s biophysical parameters than ageing itself, as illustrated in Figure 3. In fact, these physiological changes in the skin are subject to both intrinsic and extrinsic ageing. Finally, it could be interesting to consider whether these distinctive microbial profiles could be used as an indicator to quantify the biological age, instead of the sole assessment based on an individual’s chronological age.
4. Hair and Scalp Care
Across ethnicities and societies, it is generally accepted that fading hair colour, hair volume loss, and changing hair texture, such as loss of hair lustre, increasing dryness, and brittleness, are common signs of ageing hair. In addition to the effect of intrinsic ageing, external factors like mechanical, UV, heat, and chemical stresses, to which we are inevitably exposed to over time, also inflict considerable damage on our hair strands [42]. Mechanical damage refers to the accumulative friction brought about by daily habits of combing, brushing, teasing, and styling. If ranked according to increasing severity, the next and most frequent type of damage would be excessive exposure to the sun and UVR, results in the ‘weathering’ effect of hair [205]. Hair strands at this stage appear less pigmented, stiffer, rougher in texture, and less shiny. An early study by Hoting et al., 1995 [206] demonstrated that it is less of visible light but more of UVA and UVB irradiation that is responsible for the photochemical impairment of hair. Both collectively cause degradation and loss of hair proteins and hair pigments. While UVB is confined to the cuticle layers of the hair shaft, UVA is able to penetrate past the cuticles and enter deep into the cortex. It is precisely UVB’s restricted depth of penetration that causes more destructive morphological damage to hair cuticles [207]. The most severe forms of damage are heat and chemical stresses, which include perming, straightening, Brazilian keratin treatments, dyeing, and bleaching of the hair. Permanent modification of hair bonds and colour requires the usage of chemicals and high heat, which tends to remove the protective hair cuticles, exposing the hair cortex and increasing the hair fibre’s susceptibility to breakage [42].
Correspondingly, also in progressive severity, mildly damaged hair is described as showing visible wear and tear, fraying in relatively intact hair cuticle scales—this largely results in only a loss of shine. Moderate damage is characterised as having cracks along the hair fibre, cuticle edges become uplifted and dislodged in some areas, and the loss of the essential 18-MEA lipid layer, leading to a dry, dull appearance and texture, a loss of elasticity and manageability, and an increased occurrence of frizzy and flyaway hairs [208]. Gradually, without protection from the cuticles and 18-MEA, the hair cortex is exposed and repeatedly subjected to further damage, becoming more porous and giving rise to fractured hairs and split ends. In the most severe cases, individuals experience hair breakage and hair loss [42,209].
Since our hair fibres are deemed as ‘dead’ and non-renewable upon production at the follicle, damage to our hair is irreversible and cumulative over time. Fortunately, there are readily available cosmetic solutions such as hair conditioners that temporarily confer ‘repairing’ or ‘restorative’ effects to damaged hair strands. Conditioners act primarily by neutralising the negative charges along the hair fibre with positive charges and providing lubrication to hair cuticles by increasing hydrophobicity. These anti-static and lubricating agents are broadly categorised into five main groups: polymers, oils, waxes, hydrolysed amino acids, and cationic molecules [209,210]. Out of all these materials, possibly the most frequently used are silicones. There exist different types of silicones with varying degrees of deposition, adherence, and washout capacity, which can be fine-tuned to achieve different conditioning performances [211]. For a natural option, botanical extracts and oils have long been used as an emollient to moisturise hair strands. Due to their natural composition consisting of myristic acid, stearic acid, linoleic acid, palmitic acid fatty acids, and antioxidants like tocopherol, Vitamin E, carotenoids, and cysteine, these extracts have also been associated with other anti-inflammatory, anti-bacterial, anti-fungal, and photoprotection properties as well [212,213]. Similarly, natural bee products containing mixtures of honey and royal jelly have demonstrated hair protective and repair effects towards hair fibres subjected to environmental stressors of extensive UVA exposure and particulate matter 10 (PM10) [214,215]. Regardless, an ideal hair conditioner restores fibre hydrophobicity, seals and protects hair cuticle scales, and mimics the natural 18-MEA lipid layer of the cuticles. Together, they help reduce friction and frizz, improve combability, and enhancing lustre, smoothness, and manageability of hair strands.
Another means to improve hair growth or prevent damage to hair shafts is to focus on the hair scalp. For decades, scalp care, for the most part, has been concerned with the treatment and reduction of hair dandruff. Spurred by COVID-19, lockdowns, and social restrictions in 2019, the ‘skinification’ of scalp and hair care increasingly gained popularity, with an influx of commercial products promoting the care of the scalp beyond basic needs [146,147,148]. Essentially, this concept of the skinification of hair revolves around providing our scalp with the same ingredients, nourishment, care, and attention as we give to facial skin. For example, there has been increasing integration of well-known skincare ingredients such as collagen, niacinamide, salicylic acid, hyaluronic acid, and copper peptides in new hair product launches, such as shampoos, in recent years [216]. It could involve addressing moisturising, hydration, sensitivity management, greasiness and oil control, exfoliating, and anti-ageing concerns, using familiar ingredients commonly incorporated into face serums, creams, and cosmetics. These, in addition to more commonly seen caffeine, adenosine, and biotin, can be formulated into hair products [217,218,219]. Although scientific evidence supporting these ingredients are varying and show limitations, further optimisation of dosage and delivery, as well as large-scale clinical validation is required. Alternatively, new and emerging stem cell-based and exosome therapies are still in the early stages of research and development and are still considered to be experimental solutions [217]. While the effect of probiotics on various skin conditions have yet to be thoroughly explored, several beneficial effects have been documented for topical probiotic treatment of several inflammatory skin diseases like acne, rosacea, psoriasis, atopic dermatitis, and dandruff/seborrheic dermatitis [220,221]. Particularly, a patent (EP2149368B1) describes the use of probiotic Lactobacillus paracasei for the management of dandruff by increasing the hydration of scalp skin and restoring the barrier function of the scalp [222]. It is proposed that the incorporation of probiotics and prebiotics in hair care products promotes the growth of beneficial bacteria and stimulates their production of specific metabolites and antimicrobial peptides, thereby restoring the balance of a healthy scalp skin microbiome community that enhances skin barrier function and modulates adverse immune responses [220,223]. To support this notion, clinical studies by Woo et al., 2022 [224], de Jesus et al., 2023 [225], Tsai et al., 2023 [226], and Mori-Ichioka et al., 2024 [227] showed that topical treatment with lysates of lactic acid bacteria or inactivated probiotic bacteria has led to improvements in greasy or dandruff-prone scalps, and increased hair growth.
Often, consumers look for natural, botanical extracts or phytochemicals as safe, effective alternatives to current hair medications and drugs [148,228,229]. Since single compounds derived from natural plant extracts have long been used in the development of dermatological treatments protecting against environmental stressors like ultraviolet rays and pollution, as well as the alleviation of skin inflammation and keloid scarring, it is conceivable how these extracts and compounds have been explored for their potential applications in hair health [230,231]. Previous studies have shown that well-selected, natural botanical extracts and compounds enhance cell proliferation, cell survival, and cell cycle progression, upregulate IGF-1, VEGF, HGF, and KGF (Keratinocyte Growth Factor, or FGF7) growth factors, and suppress cellular senescence and apoptosis, all of which contribute to the induction and lengthening of the anagen phase and the corresponding delayed entry into the telogen phase of the hair cycle. Furthermore, several of these natural extracts and phytochemicals have been found to be associated with the stimulation of diverse signalling pathways mediated by protein kinase B (PKB, or AKT), extracellular signal-regulated kinases (ERK), Wnt/β-catenin, sonic hedgehog (Shh), JNK (c-Jun-N-terminal kinase), and JAK (Janus-activated kinase)/STAT3 (Signal transducer and activator of transcription-3), as well as to downregulate TGFβ and BMP, and exhibit 5α-reductase inhibitory properties [230,231]. As mentioned above, these signalling pathways have been shown to be involved from the early stages of hair follicle induction and morphogenesis to the maintenance of homeostasis and hair regeneration processes [130,131,232,233]. For example, a natural bee product consisting of a combination of four honeys and royal jelly in a hair follicle–dermis–epidermis model potentially promoted the pigmentation of hair and hair growth by the upregulation of krüppel-like factor 4 (KLF4), which indicated a proliferation of hair bulb SC population [234]. Taken together, these options seek to improve scalp skin health, laying a solid foundation for the growth and production of lustrous and healthy tresses.
5. Concluding Remarks
Understanding hair growth and how the hair fibre is produced involves a multi-disciplinary approach: hair biology allows the elucidation of the hair follicle cycle regulation and cellular processes governing follicle growth; hair physics interrogates the mechanics and physical properties of the protruding, non-living hair shaft; and chemistry of hair investigates the structural assembly and chemical composition of the hair strand. However, the increasing number of studies on scalp health in recent years has expanded our understanding that achieving healthy hair and scalp requires a multifaceted overview, which involves not only the hair follicle but also its surrounding scalp skin environment and supporting skin microbiome. In these present times, it is no longer sufficient to consider individual aspects of hair science separately, but rather to embrace a holistic perspective, recognizing that all these factors can collectively influence age-related changes in the hair and scalp. Notably, various differentiations of the hair and scalp across gender and ethnicities that were addressed in the text should be contemplated as well. Equipped with more integrated knowledge, together with recent advances in omics platforms for the discovery of active ingredients, developing research on stem cell-based therapeutic solutions, and the optimisation of delivery systems of actives, they will all help drive future research in hair ageing. Ultimately, deeper and more well-rounded understanding can aid scientists to better match the performance of hair care formulations to the needs of their target consumers. Furthermore, continued investigation of these aspects will aid in the elucidation of hair ageing and pave the way for the identification of novel and more effective strategies to address the different aspects of ageing hair.
Author Contributions
Conceptualisation and writing—original draft preparation, review, editing, Y.S.L. and R.K. Revision and editing, C.B., C.N. and K.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
Yi Shan Lim is the employee of Louis Vuitton Moët Hennessy (LVMH) Fragrances & Cosmetics Singapore Pte. Ltd. Carine Nizard, Karl Pays, Cecilia Brun and Robin Kurfurst the authors are employees of LVMH Recherche or Parfums Christian Dior. The authors declare that this potential conflict of interest has been disclosed in the interest of full transparency and has striven to ensure that the data and analysis in this manuscript are presented objectively.
Correction Statement
This article has been republished with a minor correction to the readability of Figures 2 and 3. This change does not affect the scientific content of the article.
Abbreviations
The following abbreviations are used in this manuscript:
| PSU | Pilosebaceous Unit |
| ORS | Outer Root Sheath |
| IRS | Inner Root Sheath |
| GM | Germinative Matrix |
| DP | Dermal Papilla |
| DS | Dermal Sheath |
| HFPU | Hair Follicle Pigmentary Unit |
| MeSC | Melanocyte Stem Cell |
| UV | Ultraviolet |
| ROS | Reactive Oxygen Species |
| L-DOPA | L-3,4-dihydroxyphenylalanine |
| KRTs | Keratins |
| KRTAP | Keratin-associated Proteins |
| KRT6 | Keratin 6 |
| KRT14/16 | Keratin 14/16 |
| KRTAP4 | Keratin-associated Protein 4 |
| FGF7 | Fibroblast Growth Factor 7 |
| FGF5 | Fibroblast Growth Factor 5 |
| DDR | DNA Damage Response |
| ATM | Ataxia-Telangiectasia Mutated |
| HFSCs | Hair Follicle Stem Cells |
| MITF | Microphthalmia-associated Transcription Factor |
| Bcl-2 | B-cell Lymphoma 2 |
| TGF-β | Transforming Growth Factor-β |
| Col17A1 | Collagen XVII |
| SCF/c-Kit | Stem Cell Factor and its receptor c-Kit |
| NFIB | Nuclear Factor I/B |
| EDNRB | Endothelin Receptor Type B |
| CXCL12 | C-X-C Motif Chemokine Ligand 12 |
| TYR | Tyrosinase |
| TYRP1 | Tyrosinase-related Proteinase 1 |
| KIT | Tyrosine Kinase Receptor for SCF |
| NER | Nucleotide Excision Repair |
| HPP | Hair Pigmentation Patterns |
| AGA | Androgenetic Alopecia |
| SA | Senescent Alopecia |
| EGF | Epidermal Growth Factor |
| IGF-1 | Insulin-like Growth Factor 1 |
| BMP | Bone Morphogenetic Protein |
| Hh | Hedgehog |
| VEGF | Vascular Endothelial Growth Factor |
| UVR | Ultraviolet Radiation |
| 18-MEA | 18-methyleicosanoic Acid |
| SC | Stratum Corneum |
| ECM | Extracellular Matrix |
| PCPN | Podoplanin |
| VCAN | Versican |
| MMP1 | Matrix Metalloproteinase 1 |
| COMP | Cartilage Oligomeric Matrix Protein |
| dWAT | Dermal White Adipose Tissue |
| ADSVCs | Adipose-derived stromal vascular cells |
| HGF | Hepatocyte Growth Factor |
| FESEM | Field Emission Scanning Electron Microscopy |
| CLSM | Confocal Laser Scanning Microscopy |
| LCM | Laser-capture Microdissection |
| AMP | Antimicrobial Peptide |
| OTUs | Operational Taxonomic Units |
| HIF-1α | Hypoxia-inducible Factor 1-α |
| WIHN | Wound-induced Hair Follicle Neogenesis |
| PM10 | Particulate Matter 10 |
| KGF | Keratinocyte Growth Factor |
| PKB | Protein Kinase B |
| ERK | Extracellular Signal-regulated Kinases |
| Shh | Sonic Hedgehog |
| JNK | c-Jun N-terminal Kinase |
| JAK | Janus-activated Kinase |
| STAT3 | Signal Transducer and Activator of Transcription 3 |
| KLF4 | Krüppel-like Factor 4 |
References
- Trüeb, R.M. The value of hair cosmetics and pharmaceuticals. Dermatology 2001, 202, 275–282. [Google Scholar] [CrossRef]
- Trüeb, R.M.; Henry, J.P.; Davis, M.G.; Schwartz, J.R. Scalp condition impacts hair growth and retention via oxidative stress. Int. J. Trichology 2018, 10, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Etcoff, N. Survival of the Prettiest: The Science of Beauty; Anchor: Hamburg, Germany, 2000. [Google Scholar]
- Symons, D. Beauty is in the adaptions of the beholder: The evolutionary psychology of human female sexual attractiveness. In Sexual Nature, Sexual Culture; Abramson, P.R., Pinkerton, S.D., Eds.; The University of Chicago Press: Chicago, IL, USA, 1995; pp. 80–119. [Google Scholar]
- Hinsz, V.B.; Matz, D.C.; Patience, R.A. Does women’s hair signal reproductive potential? J. Exp. Soc. Psychol. 2001, 37, 166–172. [Google Scholar] [CrossRef]
- Fink, B.; Hufschmidt, C.; Hirn, T.; Will, S.; McKelvey, G.; Lankhof, J. Age, health and attractiveness perception of virtual (rendered) human hair. Front. Psychol. 2016, 22, 1893. [Google Scholar] [CrossRef]
- Lim, Y.S.; Harland, D.P.; Dawson, T.L., Jr. Wanted, dead and alive: Why a multidisciplinary approach is needed to unlock hair treatment potential. Exp. Dermatol. 2019, 28, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Natarelli, N.; Gahoonia, N.; Sivamani, R.K. Integrative and mechanistic approach to the hair growth cycle and hair loss. J. Clin. Med. 2023, 12, 893–923. [Google Scholar] [CrossRef]
- Jimenez, F.; Alam, M. The proportion of catagen and telogen hair follicles in occipital scalp of male androgenetic alopecia patients: Challenging the established dogma. Exp. Dermatol. 2024, 33, e70001. [Google Scholar] [CrossRef]
- Burg, D.; Yamamoto, M.; Namekata, M.; Haklani, J.; Koike, K.; Halasz, M. Promotion of anagen, increased hair density and reduction of hair fall in a clinical setting following identification of FGF5-inhibiting compounds via a novel 2-stage process. Clin. Cosmet. Investig. Dermatol. 2017, 10, 71–85. [Google Scholar] [CrossRef]
- Harrison, S.; Bergfeld, W. Diffuse hair loss: Its triggers and management. Cleve. Clin. J. Med. 2009, 76, 361–367. [Google Scholar] [CrossRef]
- Paus, R.; Cotsarelis, G. The biology of hair follicles. N. Engl. J. Med. 1999, 341, 491–497. [Google Scholar] [CrossRef]
- Bichakjian, C.K.; Johnson, T.M. Chapter 1—Anatomy of the skin. In Local Flaps in Facial Reconstruction, 2nd ed.; Baker, S.R., Ed.; Mosby: Maryland Heights, MO, USA, 2007; pp. 3–13. [Google Scholar] [CrossRef]
- Lindberg, M.; Forslind, B. Formation and structure: An introduction to hair. In Skin, Hair, and Nails, 1st ed.; Forslind, B., Lindberg, M., Eds.; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Li, H.; Masieri, F.F.; Schneider, M.; Bartella, A.; Gaus, S.; Hahnel, S.; Zimmerer, R.; Sack, U.; Maksimovic-Ivanic, D.; Mijatovic, S.; et al. The middle part of the plucked hair follicle outer root sheath is identified as an area rich in lineage-specific stem cell markers. Biomolecules 2021, 11, 154–169. [Google Scholar] [CrossRef]
- Leerunyakul, K.; Suchonwanit, P. Asian hair: A review of structures, properties, and distinctive disorders. Clin. Cosmet. Investig. Dermatol. 2020, 13, 309–318. [Google Scholar] [CrossRef]
- Trüeb, R.M. Possibilities and limitations for reversal of age-related pigment loss. In Aging Hair; Trüeb, R.M., Tobin, D.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 207–213. [Google Scholar] [CrossRef]
- Tobin, D.J.; Paus, R. Graying: Gerontobiology of the hair follicle pigmentary unit. Exp. Gerontol. 2001, 36, 29–54. [Google Scholar] [CrossRef] [PubMed]
- Keogh, E.V.; Walsh, R.J. Rate of greying of human hair. Nature 1965, 207, 877–878. [Google Scholar] [CrossRef]
- Panhard, S.; Lozano, I.; Loussouarn, G. Greying of the human hair: A worldwide survey, revisiting the ‘50′ rule of thumb. Br. J. Dermatol. 2012, 167, 865–873. [Google Scholar] [CrossRef]
- Pandhi, D.; Khanna, D. Premature graying of hair. Indian J. Dermatol. Venereol. Leprol. 2013, 79, 641–653. [Google Scholar] [CrossRef]
- Maymone, M.B.C.; Laughter, M.; Pollock, S.; Khan, I.; Marques, T.; Abdat, R.; Goldberg, L.J.; Vashi, N.A. Hair aging in different races and ethnicities. J. Clin. Aesthet. Dermatol. 2021, 14, 38–44. [Google Scholar] [PubMed]
- Kumar, A.B.; Shamim, H.; Nagaraju, U. Premature graying of hair: Review with updates. Int. J. Trichology 2018, 10, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.J.; Paik, S.H.; Choi, J.W.; Lee, J.H.; Cho, S.; Kim, K.H.; Eun, H.C.; Oh, S.K. Hair graying pattern depends on gender, onset age and smoking habits. Acta Derm. Venereol. 2012, 92, 160–161. [Google Scholar] [CrossRef]
- Jo, S.K.; Lee, J.Y.; Lee, Y.; Kim, C.D.; Lee, J.; Lee, Y.H. Three streams for the mechanism of hair graying. Ann. Dermatol. 2018, 30, 397–401. [Google Scholar] [CrossRef]
- Rutnin, S.; Chanprapaph, K.; Pakornphadungsit, K.; Leerunyakul, K.; Visessiri, Y.; Srisont, S.; Suchonwanit, P. Variation of hair follicle counts among different scalp areas: A quantitative histopathology study. Skin Appendage Disord. 2021, 8, 24–30. [Google Scholar] [CrossRef]
- Sikkink, S.K.; Mine, S.; Freis, O.; Danoux, L.; Tobin, D.J. Stress-sensing in the human greying hair follicle: Ataxia Telangiectasia Mutated (ATM) depletion in hair bulb melanocytes in canities-prone scalp. Sci. Rep. 2020, 10, 18711. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.D.B.; Nicu, C.; Picard, M.; Chéret, J.; Bedogni, B.; Tobin, D.J.; Paus, R. The biology of human hair greying. Biol. Rev. Camb. Philos. Soc. 2021, 96, 107–128. [Google Scholar] [CrossRef]
- Wang, S.; Kang, Y.; Qi, F.; Jin, H. Genetics of hair graying with age. Ageing Res. Rev. 2023, 89, 101977. [Google Scholar] [CrossRef]
- Paus, R.; Sevilla, A.; Grichnik, J.M. Human hair graying revisited: Principles, misconceptions, and key research frontiers. J. Investig. Dermatol. 2024, 144, 474–491. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Shen, L.Y.; Wang, G.C. Role of hair follicles in the repigmentation of vitiligo. J. Investig. Dermatol. 1991, 97, 410–416. [Google Scholar] [CrossRef]
- Kauser, S.; Westgate, G.E.; Green, M.R.; Tobin, D.J. Human hair follicle and epidermal melanocytes exhibit striking differences in their aging profile which involve catalase. J. Investig. Dermatol. 2011, 131, 979–982. [Google Scholar] [CrossRef] [PubMed]
- Trüeb, R.M. Oxidative stress in ageing of hair. Int. J. Trichology 2009, 1, 6–14. [Google Scholar] [CrossRef]
- Oh, J.W.; Kloepper, J.; Langan, E.A.; Kim, Y.; Yeo, J.; Kim, M.J.; Hsi, T.; Rose, C.; Yoon, G.S.; Lee, S.; et al. A guide to studying human hair cycling in vivo. J. Investig. Dermatol. 2016, 136, 34–44. [Google Scholar] [CrossRef]
- Tobin, D.J. Aging of the hair follicle pigmentation system. Int. J. Trichology 2009, 1, 83–93. [Google Scholar] [CrossRef]
- Tobin, D.J.; Bystryn, J.C. Different populations of melanocytes are present in hair follicles and epidermis. Pigment Cell Res. 1996, 9, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Flores, A.; Saeb-Lima, M.; Cassarino, D.S. Histopathology of aging of the hair follicle. J. Cutan. Pathol. 2019, 46, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Wortsman, J.; Plonka, P.M.; Schallreuter, K.U.; Paus, R.; Tobin, D.J. Hair follicle pigmentation. J. Investig. Dermatol. 2005, 124, 13–21. [Google Scholar] [CrossRef]
- Ng, K.J.; Lim, J.; Tan, Y.N.; Quek, D.; Lim, Z.; Pantelireis, N.; Clavel, C. Sox2 in the dermal papilla regulates hair follicle pigmentation. Cell Rep. 2022, 40, 111100. [Google Scholar] [CrossRef]
- Commo, S.; Gaillard, O.; Bernard, B.A. Human hair greying is linked to a specific depletion of hair follicle melanocytes affecting both the bulb and the outer root sheath. Br. J. Dermatol. 2004, 150, 435–443. [Google Scholar] [CrossRef]
- Arck, P.C.; Overall, R.; Spatz, K.; Liezman, C.; Handjiski, B.; Klapp, B.F.; Birch-Machin, M.A.; Peters, E.M.J. Towards a “free radical theory of graying”: Melanocyte apoptosis in the aging human hair follicle is an indicator of oxidative stress induced tissue damage. FASEB J. 2006, 20, 1567–1569. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.S.; Dawson, T.L. Ageing hair in Asians and Caucasians. In Practical Aspects of Hair Transplantation in Asians; Pathomvanich, D., Imagawa, K., Eds.; Springer: Tokyo, Japan, 2018. [Google Scholar] [CrossRef]
- Tobin, D.J.; Hagen, E.; Botchkarev, V.A.; Paus, R. Do hair bulb melanocytes undergo apoptosis during hair follicle regression (catagen)? J. Investig. Dermatol. 1998, 111, 941–947. [Google Scholar] [CrossRef]
- Van Neste, D. Thickness, medullation and growth rate of female scalp hair are subject to significant variation according to pigmentation and scalp location during ageing. Eur. J. Dermatol. 2004, 14, 28–32. [Google Scholar]
- Choi, H.I.; Choi, G.I.; Kim, E.K.; Choi, Y.J.; Sohn, K.C.; Lee, Y.; Kim, C.D.; Yoon, T.J.; Sohn, H.J.; Han, S.H.; et al. Hair greying is associated with active hair growth. Br. J. Dermatol. 2011, 165, 1183–1189. [Google Scholar] [CrossRef]
- Inomata, K.; Aoto, T.; Nguyen, T.B.; Okamoto, N.; Tanimura, S.; Wakayama, T.; Iseki, S.; Hara, E.; Masunaga, T.; Shimizu, H.; et al. Genotoxic stress abrogates renewal of melanocyte stem cells by triggering their differentiation. Cell 2009, 137, 1088–1099. [Google Scholar] [CrossRef]
- Nishimura, E.K.; Granter, S.R.; Fisher, D.E. Mechanisms of hair greying: Incomplete melanocyte stem cell maintenance in the niche. Science 2005, 307, 720–724. [Google Scholar] [CrossRef]
- Nishimura, E.K.; Suzuki, M.; Igras, V.; Du, J.; Lonning, S.; Miyachi, Y.; Roes, J.; Beermann, F.; Fisher, D.E. Key roles for Transforming Growth Factor β in melanocyte stem cell maintenance. Cell Stem Cell 2010, 6, 130–140. [Google Scholar] [CrossRef]
- Nishimura, E.K. Melanocyte stem cells: A melanocyte reservoir in hair follicles for hair and skin pigmentation. Pigment Cell Melanoma Res. 2011, 24, 401–410. [Google Scholar] [CrossRef]
- Tanimura, S.; Tadokoro, Y.; Inomata, K.; Nguyen, T.B.; Nishie, W.; Yamazaki, S.; Nakauchi, H.; Tanaka, Y.; McMillan, J.R.; Sawamura, D.; et al. Hair follicle stem cells provide a functional niche for melanocyte stem cells. Cell Stem Cell 2011, 8, 177–187. [Google Scholar] [CrossRef]
- Botchkareva, N.V.; Khlgatian, M.; Longley, B.J.; Botchkarev, V.A.; Gilchrest, B.A. SCF/c-kit signaling is required for cyclic regeneration of the hair pigmentation unit. FASEB J. 2001, 15, 645–658. [Google Scholar] [CrossRef]
- Moriyama, M.; Osawa, M.; Mak, S.; Ohtsuka, T.; Yamamoto, N.; Han, H.; Delmas, V.; Kageyama, R.; Beermann, F.; Larue, L.; et al. Notch signalling via Hes1 transcription factor maintains survival of melanoblasts and melanocyte stem cells. J. Cell Biol. 2006, 173, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Schouwey, K.; Delmas, V.; Larue, L.; Zimber-Strobl, U.; Strobl, L.J.; Radtke, F.; Beermann, F. Notch1 and Notch2 receptors influence progressive hair graying in a dose-dependent manner. Dev. Dyn. 2007, 236, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Valluet, A.; Druillennec, S.; Barbotin, C.; Dorard, C.; Monsoro-Burg, A.; Larcher, M.; Pouponnot, C.; Baccarini, M.; Larue, L.; Eychène, A. B-Raf and C-Raf are required for melanocyte stem cell self-maintenance. Cell Rep. 2012, 2, 774–780. [Google Scholar] [CrossRef]
- Kubic, J.D.; Young, K.P.; Plummer, R.S.; Ludvik, A.E.; Lang, D. Pigmentation PAX-ways: The role of Pax3 in melanogenesis, melanocyte stem cell maintenance, and disease. Pigment Cell Melanoma Res. 2008, 21, 627–645. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zuo, Y.; Li, S.; Li, C. Melanocyte stem cells in the skin: Origin, biological characteristics, homeostatic maintenance and therapeutic potential. Clin. Transl. Med. 2024, 14, e1720. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Yoon, T.J.; Lee, Y.H. Changing expression of the genes related to human hair graying. Eur. J. Dermatol. 2008, 18, 397–399. [Google Scholar] [PubMed]
- Peters, E.M.J.; Liezmann, C.; Spatz, K.; Ungethüm, U.; Kuban, R.; Daniltchenko, M.; Kruse, J.; Imfeld, D.; Klapp, B.F.; Campiche, R. Profiling mRNA of the graying human hair follicle constitutes a promising state-of-the-art tool to assess its aging: An exemplary report. J. Investig. Dermatol. 2013, 133, 1150–1160. [Google Scholar] [CrossRef] [PubMed]
- Adav, S.S.; Ng, K.W. Recent omics advances in hair aging biology and hair biomarker analysis. Ageing Res. Rev. 2023, 91, 102041. [Google Scholar] [CrossRef]
- Yu, M.; Bell, R.H.; Ho, M.M.; Leung, G.; Haegert, A.; Carr, N.; Shapiro, J.; McElwee, K.J. Deficiency in Nucleotide Excision Repair family gene activity, especially ERCC3, is associated with non-pigmented hair fiber growth. PLoS ONE 2012, 7, e34185. [Google Scholar] [CrossRef]
- Sun, Q.; Lee, W.; Hu, H.; Ogawa, T.; De Leon, S.; Katehis, I.; Lim, C.H.; Takeo, M.; Cammer, M.; Taketo, M.M.; et al. Dedifferentiation maintains melanocyte stem cells in a dynamic niche. Nature 2023, 616, 774–782. [Google Scholar] [CrossRef]
- Rosenberg, A.M.; Rausser, S.; Ren, J.; Mosharov, E.V.; Sturm, G.; Ogden, R.T.; Patel, P.; Soni, R.K.; Lacefield, C.; Tobin, D.J.; et al. Quantitative mapping of human hair greying and reversal in relation to life stress. eLife 2021, 10, e67437. [Google Scholar] [CrossRef]
- Singh, S.; Muthuvel, K. Practical approach to hair loss diagnosis. Indian J. Plast. Surg. 2021, 54, 399–403. [Google Scholar] [CrossRef]
- Phillips, T.G.; Slomiany, W.P.; Allison, R. Hair loss: Common causes and treatment. Am. Fam. Physician 2017, 96, 371–378. [Google Scholar]
- Qi, J.; Garza, L.A. An overview of alopecias. Cold Spring Harb. Perspect. Med. 2014, 4, a013615. [Google Scholar] [CrossRef]
- Gokce, N.; Basgoz, N.; Kenanoglu, S.; Akalin, H.; Ozkul, Y.; Ergoren, M.C.; Becarri, T.; Bertelli, M.; Dundar, M. An overview of the genetic aspects of hair loss and its connection with nutrition. J. Prev. Med. Hyg. 2022, 63, E228–E238. [Google Scholar] [CrossRef] [PubMed]
- Morgan, B.A. The dermal papilla: An instructive niche for epithelial stem and progenitor cells in development and regeneration of the hair follicle. Cold Spring Harb. Perspect. Med. 2014, 4, a015180. [Google Scholar] [CrossRef]
- Lim, J.; Ng, K.J.; Clavel, C. Chapter Four—Dermal papilla regulation of hair growth and pigmentation. In Advances in Stem Cells and their Niches; Perez-Moreno, M., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 115–138. [Google Scholar] [CrossRef]
- Zhang, H.; Qiu, X.; Liao, X. Dermal papilla cells: From basic research to translational applications. Biology 2024, 13, 842–860. [Google Scholar] [CrossRef]
- Van Scott, E.J.; Eckel, T.M. Geometric relationships between the matrix of the hair bulb and its dermal papilla in normal and alopecic scalp. J. Investig. Dermatol. 1958, 31, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Elliott, K.; Stephenson, T.J.; Messenger, A.G. Differences in hair follicle dermal papilla volume are due to extracellular matrix volume and cell number: Implications for the control of hair follicle size and androgen responses. J. Investig. Dermatol. 1999, 113, 873–877. [Google Scholar] [CrossRef]
- Miranda, B.H.; Tobin, D.J.; Sharpe, D.T.; Randall, V.A. Intermediate hair follicles: A new more clinically relevant model for hair growth investigations. Br. J. Dermatol. 2010, 163, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Abdin, R.; Zhang, Y.; Jimenez, J.J. Treatment of androgenetic alopecia using PRP to target dysregulated mechanisms and pathways. Front. Med. 2022, 16, 843127. [Google Scholar] [CrossRef]
- You, J.; Jang, Y.; Sim, J.; Ryu, D.; Cho, E.; Park, D.; Jung, E. Anti-hair loss effect of veratric acid on dermal papilla cells. Int. J. Mol. Sci. 2025, 26, 2240. [Google Scholar] [CrossRef]
- Pappalardo, A.; Kim, J.Y.; Abaci, H.E.; Christiano, A.M. Restoration of hair follicle inductive properties by depletion of senescent cells. Aging Cell 2025, 24, e14353. [Google Scholar] [CrossRef]
- Dong, T.; Li, Y.; Jin, S.; Yang, F.; Xiong, R.; Dai, Y.; Song, X.; Guan, C. Progress on mitochondria and hair follicle development in androgenetic alopecia: Relationships and therapeutic perspectives. Stem Cell Res. Ther. 2025, 16, 44–65. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Y.; Xiong, Y.; Li, Y. Pathogenesis and regenerative therapy in vitiligo and alopecia areata: Focus on hair follicle. Front. Med. 2025, 11, 1510363. [Google Scholar] [CrossRef]
- Joulai Veijouye, S.; Yari, A.; Heidari, F.; Sajedi, N.; Ghoroghi Moghani, F.; Nobakht, M. Bulge region as a putative hair follicle stem cells niche: A brief review. Iran. J. Public Health 2017, 46, 1167–1175. [Google Scholar]
- Xing, Y.; Guo, H.; Xiang, F.; Li, Y. Recent progress in hair follicle stem cell markers and their regulatory roles. World J. Stem Cells 2024, 16, 126–136. [Google Scholar] [CrossRef]
- Chen, C.; Huang, W.; Wang, E.H.C.; Tai, K.; Lin, S. Functional complexity of hair follicle stem cell niche and therapeutic targeting of niche dysfunction for hair regeneration. J. Biomed Sci. 2020, 27, 43–54. [Google Scholar] [CrossRef]
- Jang, H.; Jo, Y.; Lee, J.H.; Choi, S. Aging of hair follicle stem cells and their niches. BMB Rep. 2023, 56, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Choi, S. Deciphering the molecular mechanisms of stem cell dynamics in hair follicle regeneration. Exp. Mol. Med. 2024, 56, 110–117. [Google Scholar] [CrossRef]
- Matsumura, H.; Mohri, Y.; Binh, N.T.; Morinaga, H.; Fukuda, M.; Ito, M.; Kurata, S.; Hoeijmakers, J.; Nishimura, E.K. Hair follicle aging is driven by transepidermal elimination of stem cells via COL17A1 proteolysis. Science 2016, 351, aad4395. [Google Scholar] [CrossRef] [PubMed]
- Keyes, B.E.; Segal, J.P.; Heller, E.; Lien, W.; Chang, C.; Guo, X.; Oristian, D.S.; Zheng, D.; Fuchs, E. Nfatc1 orchestrates aging in hair follicle stem cells. Proc. Natl. Acad. Sci. USA 2013, 110, E4950–E4959. [Google Scholar] [CrossRef] [PubMed]
- Messenger, A.G. Hair through the female life cycle. Br. J. Dermatol. 2011, 165, 2–6. [Google Scholar] [CrossRef]
- Deng, Y.; Wang, M.; He, Y.; Liu, F.; Chen, L.; Xiong, X. Cellular senescence: Ageing and androgenetic alopecia. Dermatology 2023, 239, 533–541. [Google Scholar] [CrossRef]
- Ji, J.; Ho, B.S.; Qian, G.; Xie, X.; Bigliardi, P.L.; Bigliardi-Qi, M. Aging in hair follicle stem cells and niche microenvironment. J. Dermatol. 2017, 44, 1097–1104. [Google Scholar] [CrossRef]
- Ge, Y.; Miao, Y.; Gur-Cohen, S.; Gomez, N.; Yang, H.; Nikolova, M.; Polak, L.; Hu, Y.; Verma, A.; Elemento, O.; et al. The aging microenvironment dictates stem cell behavior. Proc. Natl. Acad. Sci. USA 2020, 117, 5339–5350. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.L.; Mishra, P.; Martin, N.; George, N.O.; Sakk, V.; Soller, K.; Nalapareddy, K.; Nattamai, K.; Scharffetter-Kochanek, K.; Florian, M.C.; et al. A Wnt5a-Cdc42 axis controls aging and rejuvenation of hair-follicle stem cells. Aging 2021, 13, 4778–4793. [Google Scholar] [CrossRef] [PubMed]
- Egger, A.; Tomic-Canic, M.; Tosti, A. Advances in stem cell-based therapy for hair loss. CellR4 Repair Replace. Regen. Reprogram. 2020, 8, e2894. [Google Scholar] [PubMed]
- Whiting, D.A. How real is senescent alopecia? A histopathologic approach. Clin. Dermatol. 2011, 29, 49–53. [Google Scholar] [CrossRef]
- Ntshingila, S.; Oputu, O.; Arowolo, A.T.; Khumalo, N.P. Androgenetic alopecia: An update. JAAD Int. 2023, 13, 150–158. [Google Scholar] [CrossRef]
- Chen, S.; Xie, X.; Zhang, G.; Zhang, Y. Comorbidities in androgenetic alopecia: A comprehensive review. Dermatol. Ther. 2022, 12, 2233–2247. [Google Scholar] [CrossRef]
- Trüeb, R.M.; Rezende, H.D.; Dias, M.F.R.G. A comment on the science of hair aging. Int. J. Trichology 2018, 10, 245–254. [Google Scholar] [CrossRef]
- Kligman, A.M. The comparative histopathology of male-pattern baldness and senescent baldness. Clin. Dermatol. 1988, 6, 108–118. [Google Scholar] [CrossRef]
- Karnik, P.; Shah, S.; Dvorkin-Wininger, Y.; Oshtory, S.; Mirmirani, P. Microarray analysis of androgenetic and senescent alopecia: Comparison of gene expression shows two distinct profiles. J. Dermatol. Sci. 2013, 72, 183–186. [Google Scholar] [CrossRef]
- Hillmer, A.M.; Hanneken, S.; Ritzmann, S.; Becker, T.; Freudenberg, J.; Brockschmidt, F.F.; Flaquer, A.; Freudenberg-Hua, Y.; Jamra, R.A.; Metzen, C.; et al. Genetic variation in the human androgen receptor gene is the major determinant of common early-onset androgenetic alopecia. Am. J. Hum. Genet. 2005, 77, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Brockschimidt, F.F.; Kiefer, A.K.; Stefansson, H.; Nyholt, D.R.; Song, K.; Vermeulen, S.H.; Kanoni, S.; Glass, D.; Medland, S.E.; et al. Six novel susceptibility Loci for early-onset androgenetic alopecia and their unexpected association with common diseases. PLoS Genet. 2012, 8, e1002746. [Google Scholar] [CrossRef] [PubMed]
- Courtney, A.; Triwongwarant, D.; Chim, I.; Eisman, S.; Sinclair, R. Evaluating 5 alpha reductase inhibitors for the treatment of male androgenetic alopecia. Expert Opin. Pharmacother. 2023, 24, 1919–1922. [Google Scholar] [CrossRef] [PubMed]
- Hirshburg, J.M.; Kelsey, P.A.; Therrien, C.A.; Gavino, A.C.; Reichenberg, J.S. Adverse effects and safety of 5-alpha reductase inhibitors (Finasteride, Dustasteride): A systematic review. J. Clin. Aesthet. Dermatol. 2016, 9, 56–62. [Google Scholar]
- Sawaya, M.E.; Price, V.H. Different levels of 5α-reductase type I & II, aromatase, and androgen receptor in hair follicles of women and men with androgenetic alopecia. J. Investig. Dermatol. 1997, 109, 296–300. [Google Scholar] [CrossRef]
- Messenger, A.G.; Rundegren, J. Minoxidil: Mechanisms of action on hair growth. Br. J. Dermatol. 2004, 150, 186–194. [Google Scholar] [CrossRef]
- Gupta, A.K.; Taluker, M.; Venkataraman, M.; Bamimore, M.A. Minoxidil: A comprehensive review. J. Dermatol. Treat. 2022, 33, 1896–1906. [Google Scholar] [CrossRef] [PubMed]
- Orasan, M.S.; Bolfa, P.; Coneac, A.; Muresan, A.; Mihu, C. Topical products for human hair regeneration: A comparative study on an animal model. Ann. Dermatol. 2016, 28, 65–73. [Google Scholar] [CrossRef]
- Naito, A.; Midorikawa, T.; Yoshino, T.; Ohdera, M. Lipid peroxides induce early onset of catagen phase in murine hair cycles. Int. J. Mol. Med. 2008, 22, 725–729. [Google Scholar] [CrossRef]
- Bahta, A.W.; Farjo, N.; Farjo, B.; Philpott, M.P. Premature senescence of balding dermal papilla cells in vitro is associated with p16INK4a expression. J. Investig. Dermatol. 2008, 128, 1088–1094. [Google Scholar] [CrossRef]
- Jaworsky, C.; Kligman, A.M.; Murphy, G.F. Characterization of inflammatory infiltrates in male pattern alopecia: Implications for pathogenesis. Br. J. Dermatol. 1992, 127, 239–246. [Google Scholar] [CrossRef]
- Mahé, Y.F.; Michelet, J.F.; Billoni, N.; Jarrousse, F.; Buan, B.; Commo, S.; Saint-Léger, D.; Bernard, B.A. Androgenetic alopecia and microinflammation. Int. J. Dermatol. 2000, 39, 576–584. [Google Scholar] [CrossRef]
- Lee, Y.I.; Choi, S.; Roh, W.S.; Lee, J.H.; Kim, T. Cellular senescence and inflammaging in the skin microenvironment. Int. J. Mol. Sci. 2021, 22, 3849. [Google Scholar] [CrossRef] [PubMed]
- Whiting, D.A. Diagnostic and predictive value of horizontal sections of scalp biopsy specimens in male pattern androgenetic alopecia. J. Am. Acad. Dermatol. 1993, 28 Pt 1, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Eichmüller, S.; van der Veen, C.; Moll, I.; Hermes, B.; Hofmann, U.; Müller-Röver, S.; Paus, R. Clusters of perifollicular macrophages in normal murine skin: Physiological degeneration of selected hair follicles by programmed organ deletion. J. Histochem. Cytochem. 1998, 46, 361–370. [Google Scholar] [CrossRef]
- Van Neste, D.; Tobin, D.J. Hair cycle and hair pigmentation: Dynamic interactions and changes associated with aging. Micron 2004, 35, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Robbins, C.; Mirmirani, P.; Messenger, A.G.; Birch, M.P.; Youngquist, R.S.; Tamura, M.; Filloon, T.; Luo, F.; Dawson, T.L., Jr. What women want—Quantifying the perception of hair amount: An analysis of hair diameter and density changes with age in Caucasian women. Br. J. Dermatol. 2012, 167, 324–332. [Google Scholar] [CrossRef]
- Otsuka, H.; Nemoto, T. Study on Japanese hair. J. Jpn. Cosmet. Sci. Soc. 1988, 12, 192–197. [Google Scholar]
- Trotter, M.; Dawson, H.L. The hair of French Canadians. Am. J. Phys. Anthrop. 1934, 18, 443–456. [Google Scholar] [CrossRef]
- Courtois, M.; Loussouarn, G.; Hourseau, C.; Grollier, J.F. Ageing and hair cycles. Br. J. Dermatol. 1995, 132, 86–93. [Google Scholar] [CrossRef]
- Mirmirani, P.; Luo, F.; Youngquist, S.R.; Fisher, B.K.; Li, J.; Oblong, J.; Dawson, T.L., Jr. Hair growth parameters in pre-and postmenopausal women. In Aging Hair; Trüeb, R.M., Tobin, D.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 49–60. [Google Scholar] [CrossRef]
- Baltenneck, F.; Genty, G.; Samra, E.B.; Richena, M.; Harland, D.P.; Clerens, S.; Leccia, E.; Le Blach, M.; Doucet, J.; Michelet, J.; et al. Age-associated thin hair displays molecular, structural and mechanical characteristic changes. J. Struct. Biol. 2022, 214, 107908. [Google Scholar] [CrossRef]
- Scott, G.V.; Robbins, C.R. Stiffness of human hair fibers. J. Soc. Cosmet. Chem. 1978, 29, 469–485. [Google Scholar]
- Robbins, C.R.; Crawford, R.J. Cuticle damage and the tensile properties of human hair. J. Soc. Cosmet. Chem. 1991, 42, 49–58. [Google Scholar]
- Persaud, D.; Kamath, Y.K. Torsional method for evaluating hair damage and performance of hair care products. J. Cosmet. Sci. 2004, 55, S65–S77. [Google Scholar]
- Tang, W.; Zhang, S.G.; Zhang, J.K.; Chen, S.; Zhu, H.; Ge, S.R. Ageing effects on the diameter, nanomechanical properties and tactile perception of human hair. Int. J. Cosmet. Sci. 2016, 38, 155–163. [Google Scholar] [CrossRef]
- Kim, S.N.; Lee, S.Y.; Choi, M.H.; Joo, K.M.; Kim, S.H.; Koh, J.S.; Park, W.S. Characteristic features of ageing in Korean women’s hair and scalp. Br. J. Dermatol. 2013, 168, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Plott, T.J.; Karim, N.; Durbin-Johnson, B.P.; Swift, D.P.; Youngquist, R.S.; Salemi, M.; Phinney, B.S.; Rocke, D.M.; Davis, M.G.; Parker, G.J.; et al. Age-related changes in hair shaft protein profiling and genetically variant peptides. Forensic Sci. Int. Genet. 2020, 47, 102309. [Google Scholar] [CrossRef]
- Thibaut, S.; Becker, E.D.; Bernard, B.A.; Huart, M.; Fiat, F.; Baghdadli, N.; Luengo, G.S.; Leroy, F.; Angevin, P.; Kermoal, A.M.; et al. Chronological ageing of human hair keratin fibres. Int. J. Cosmet. Sci. 2010, 32, 422–434. [Google Scholar] [CrossRef]
- Nagase, S.; Kajiura, Y.; Mamada, A.; Abe, H.; Shibuichi, S.; Satoh, N.; Itou, T.; Shinohara, Y.; Amemiya, Y. Changes in structure and geometric properties of human hair by aging. J. Cosmet. Sci. 2010, 60, 637–648. [Google Scholar]
- Nagase, S.; Tsuchiya, M.; Matsui, T.; Shibuichi, S.; Tsujimura, H.; Masukawa, Y.; Satoh, N.; Itou, T.; Koike, K.; Tsujii, K. Characterization of curved hair of Japanese women with reference to internal structures and amino acid composition. J. Cosmet. Sci. 2008, 59, 317–332. [Google Scholar] [PubMed]
- Hollfelder, B.; Blankenburg, G.; Wolfram, L.J.; Höcker, H. Chemical and physical properties of pigmented and non-pigmented hair (‘grey hair’). Int. J. Cosmet. Sci. 1995, 17, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, A.C.S.; Joekes, I. Hair color changes and protein damage caused by ultraviolet radiation. J. Photochem. Photobiol. B 2004, 74, 109–117. [Google Scholar] [CrossRef]
- Stenn, K.S.; Karnik, P. Lipids to the top of hair biology. J. Investig. Dermatol. 2010, 130, 1205–1207. [Google Scholar] [CrossRef]
- Ji, J.H.; Park, T.S.; Lee, H.J.; Kim, Y.D.; Pi, L.Q.; Jin, X.H.; Lee, W. The ethnic differences of the damage of hair and integral hair lipid after ultra violet radiation. Ann. Dermatol. 2013, 25, 54–60. [Google Scholar] [CrossRef]
- Lee, W. Integral hair lipid in human hair follicle. J. Dermatol. Sci. 2011, 64, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Harel, S.; Christiano, A.M. Genetics of structural hair disorders. J. Investig. Dermatol. 2012, 132, E22–E26. [Google Scholar] [CrossRef] [PubMed]
- Coderch, L.; Alonso, C.; Garcia, M.T.; Pérez, L.; Martí, M. Hair lipid structure: Effect of surfactants. Cosmetics 2023, 10, 107. [Google Scholar] [CrossRef]
- Tokunaga, S.; Tanamachi, H.; Ishikawa, K. Degradation of hair surface: Importance of 18-MEA and epicuticle. Cosmetics 2019, 6, 31. [Google Scholar] [CrossRef]
- Song, S.; Park, H.S.; Lim, B.T.; Son, S.K. Biomimetics through bioconjugation of 16-methylheptadecanoic acid to damaged hair for hair barrier recovery. Sci. Rep. 2024, 14, 27387. [Google Scholar] [CrossRef]
- Thiboutot, D. Regulation of human sebaceous glands. J. Investig. Dermatol. 2004, 123, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nicolaides, N.; Rothman, S. Studies on the chemical composition of human hair fat. II. The overall composition with regard to age, sex and race. J. Investig. Dermatol. 1953, 21, 9–14. [Google Scholar] [CrossRef]
- Oliver, M.A.; Martí, M.; Coderch, L.; Carrer, V.; Kreuzer, M.; Barba, C. Lipid loses and barrier function modifications of the brown-to-white hair transition. Skin Res. Technol. 2019, 25, 517–525. [Google Scholar] [CrossRef]
- Breakspear, S.; Smith, J.R.; Luengo, G. Effect of the covalently linked fatty acid 18-MEA on the nanotribology of hair’s outermost surface. J. Struct. Biol. 2005, 149, 235–242. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, Y.D.; Pi, L.; Lee, S.Y.; Hong, H.; Lee, W. Comparison of hair shaft damage after chemical treatment in Asian, White European, and African hair. Int. J. Dermatol. 2014, 53, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Masukawa, Y.; Tanamachi, H.; Tsujimura, H.; Mamada, A.; Imokawa, G. Characterization of hair lipid images by argon sputter etching-scanning electron microscopy. Lipids 2006, 41, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Mamada, A.; Breakspear, S.; Itou, T.; Tanji, N. Age-dependent changes in damage processes of hair cuticle. J. Cosmet. Dermatol. 2015, 14, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Vyumvuhore, R.; Verzeaux, L.; Gilardeau, S.; Bordes, S.; Aymard, E.; Manfait, M.; Closs, B. Investigation of the molecular signature of greying hair shafts. Int. J. Cosmet. Sci. 2021, 43, 332–340. [Google Scholar] [CrossRef]
- O’goshi, K.; Iguchi, M.; Tagami, H. Functional analysis of the stratum corneum of scalp skin: Studies in patients with alopecia areata and androgenetic alopecia. Arch. Dermatol. Res. 2000, 292, 605–611. [Google Scholar] [CrossRef]
- Lam-Phaure, L. Formulating on Trend: Skinification of Hair. Cosmetics & Toiletries. 2022. Available online: https://www.cosmeticsandtoiletries.com/formulas-products/formulating-basics/article/22288816/cosmetics-toiletries-magazine-formulating-on-trend-skinification-of-hair (accessed on 3 July 2025).
- Chia, V. The ‘Skinification’ of Hair Is the Biggest Beauty Trend Right Now—But What Is It? CNA Lifestyle. 2021. Available online: https://cnalifestyle.channelnewsasia.com/women/beauty-trend-skinification-hair-skincare-haircare-products-280901 (accessed on 3 July 2025).
- Hasegawa, R. The Future of Haircare, Styling & Colour: 2024 [Industry Report]; Mintel: London, UK, 2024. [Google Scholar]
- Schwartz, J.R.; Henry, J.P.; Kerr, K.M.; Flagler, M.J.; Page, S.H.; Redman-Furey, N. Incubatory environment of the scalp impacts pre-emergent hair to affect post-emergent hair cuticle integrity. J. Cosmet. Dermatol. 2018, 17, 105–111. [Google Scholar] [CrossRef]
- Cao, W.; Li, L.; Kaijura, S.; Amoh, Y.; Tan, Y.; Liu, F.; Hoffman, R.M. Aging hair follicles rejuvenated by transplantation to a young subcutaneous environment. Cell Cycle 2016, 15, 1093–1098. [Google Scholar] [CrossRef]
- Chanprapaph, K.; Sutharaphan, T.; Suchonwanit, P. Scalp biophysical characteristics in males with androgenetic alopecia: A comparative study with healthy controls. Clin. Interv. Aging 2021, 16, 781–787. [Google Scholar] [CrossRef]
- Schwartz, J.R.; Henry, J.P.; Kerr, K.M.; Mizoguchi, H.; Li, L. The role of oxidative damage in poor scalp health: Ramifications to causality and associated hair growth. Int. J. Cosmet. Sci. 2015, 37, 9–15. [Google Scholar] [CrossRef]
- Tosti, A.; Schwartz, J.R. Role of scalp health in achieving optimal hair growth and retention. Int. J. Cosmet. Sci. 2021, 43, S1–S8. [Google Scholar] [CrossRef]
- Hori, H.; Moretti, G.; Rebora, A.; Crovato, F. The thickness of human scalp: Normal and bald. J. Investig. Dermatol. 1972, 58, 396–399. [Google Scholar] [CrossRef]
- Florence, P.; Cornillon, C.; D’arras, M.; Flament, F.; Panhard, S.; Diridollou, S.; Loussouarn, G. Functional and structural age-related changes in the scalp skin of Caucasian women. Skin Res. Technol. 2013, 19, 384–393. [Google Scholar] [CrossRef]
- Lee, Y.I.; Kim, J.; Park, S.R.; Ham, S.; Lee, H.J.; Park, C.R.; Kim, H.N.; Kang, B.H.; Jung, I.; Suk, J.M.; et al. Age-related changes in scalp biophysical parameters: A comparative analysis of the 20s and 50s age groups. Skin Res. Technol. 2023, 29, e13433. [Google Scholar] [CrossRef]
- Piérard-Franchimont, C.; Loussouarn, G.; Panhard, S.; Saint Léger, D.; Mellul, M.; Piérard, G.E. Immunohistochemical patterns in the interfollicular Caucasian scalps: Influences of age, gender, and alopecia. Biomed Res. Int. 2013, 2013, 769489. [Google Scholar] [CrossRef]
- Zhang, S.; Duan, E. Fighting against skin aging. Cell Transplant. 2018, 27, 729–738. [Google Scholar] [CrossRef]
- Gunin, A.G.; Petrov, V.V.; Vasilieva, O.V.; Golubtsova, N.N. Age-related changes of blood vessels in the human dermis. Adv. Gerontol. 2015, 5, 65–71. [Google Scholar] [CrossRef]
- Williams, R.; Westgate, G.; Pawlus, A.D.; Sikkink, S.K.; Thornton, M.J. Age-related changes in female scalp dermal sheath and dermal fibroblasts: How the hair follicle environment impacts hair ageing. J. Investig. Dermatol. 2021, 141, 1041–1051. [Google Scholar] [CrossRef]
- Jo, S.J.; Kim, J.Y.; Jang, S.; Choi, S.J.; Kim, K.H.; Kwon, O. Decrease of versican levels in the follicular dermal papilla is a remarkable aging-associated change of human hair follicles. J. Dermatol. Sci. 2016, 84, 354–357. [Google Scholar] [CrossRef]
- Weihermann, A.C.; Lorencini, M.; Brohem, C.A.; de Carvalho, C.M. Elastin structure and its involvement in skin photoageing. Int. J. Cosmet. Sci. 2017, 39, 241–247. [Google Scholar] [CrossRef]
- Johnston, R.B. Chapter 12—Disorders of elastic tissue. In Weedon’s Skin Pathology Essentials E-Book, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 255–265. [Google Scholar]
- Williams, R.; Pawlus, A.D.; Thorton, M.J. Getting under the skin of hair aging: The impact of the hair follicle environment. Exp. Dermatol. 2020, 29, 588–597. [Google Scholar] [CrossRef]
- Freitas-Rodríguez, S.; Folgueras, A.R.; López-Otín, C. The role of matrix metalloproteinases in aging: Tissue remodeling and beyond. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 2015–2025. [Google Scholar] [CrossRef]
- Anderi, R.; Makdissy, N.; Azar, A.; Rizk, F.; Hamade, A. Cellular therapy with human autologous adipose-derived adult cells of stromal vascular fraction for alopecia areata. Stem Cell Res. Ther. 2018, 9, 141–150. [Google Scholar] [CrossRef]
- Nicu, C.; O’Sullivan, J.D.B.; Ramos, R.; Timperi, L.; Lai, T.; Farjo, N.; Farjo, B.; Pople, J.; Bhogal, R.; Hardman, J.A.; et al. Dermal adipose tissue secretes HGF to promote human hair growth and pigmentation. J. Investig. Dermatol. 2021, 141, 1633–1645. [Google Scholar] [CrossRef]
- Festa, E.; Fretz, J.; Berry, R.; Schmidt, B.; Rodeheffer, M.; Horowitz, M.; Horsley, V. Adipocyte lineage cells contribute to the skin stem cell niche to drive hair cycling. Cell 2011, 146, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Shook, B.; Gonzalez, G.R.; Ebmeier, S.; Grisotti, G.; Zwick, R.; Horsley, V. The role of adipocytes in tissue regeneration and stem cell niches. Annu. Rev. Cell Dev. Biol. 2016, 32, 609–631. [Google Scholar] [CrossRef] [PubMed]
- Li, K.N.; Jain, P.; He, C.H.; Eun, F.C.; Kang, S.; Tumbar, T. Skin vasculature and hair follicle cross-talking associated with stem cell activation and tissue homeostasis. Elife 2019, 8, e45977. [Google Scholar] [CrossRef]
- Gur-Cohen, S.; Yang, H.; Baksh, S.C.; Miao, Y.; Levorse, J.; Kataru, R.P.; Liu, X.; de la Cruz-Racelis, J.; Mehrara, B.J.; Fuchs, E. Stem cell-driven lymphatic remodeling coordinates tissue regeneration. Science 2019, 366, 1218–1225. [Google Scholar] [CrossRef]
- Peña-Jimenez, D.; Fontenete, S.; Megias, D.; Fustero-Torre, C.; Graña-Castro, O.; Castellana, D.; Loewe, R.; Perez-Moreno, M. Lymphatic vessels interact dynamically with the hair follicle stem cell niche during skin regeneration in vivo. EMBO J. 2019, 38, e101688. [Google Scholar] [CrossRef]
- Pająk, J.; Nowicka, D.; Szepietowski, J.C. Inflammaging and immunosenescence as part of skin aging—A narrative review. Int. J. Mol. Sci. 2023, 24, 7784. [Google Scholar] [CrossRef]
- Cole, M.A.; Quan, T.; Voorhees, J.J.; Fisher, G.J. Extracellular matrix regulation of fibroblast function: Redefining our perspective on skin aging. J. Cell Commun. Signal. 2018, 12, 35–43. [Google Scholar] [CrossRef]
- Solé-Boldo, L.; Raddatz, G.; Schütz, S.; Mallm, J.; Rippe, K.; Lonsdorf, A.S.; Rodriguez-Paredes, M.; Lyko, F. Single-cell transcriptomes of the human skin reveal age-related loss of fibroblast priming. Commun. Biol. 2020, 3, 188. [Google Scholar] [CrossRef]
- Salzer, M.C.; Lafzi, A.; Berenguer-Llergo, A.; Youssif, C.; Castellanos, A.; Solanas, G.; Peixoto, F.O.; Attolini, C.S.; Prats, N.; Aguilera, M.; et al. Identity noise and adipogenic traits characterize dermal fibroblast aging. Cell 2018, 175, 1575–1590.e22. [Google Scholar] [CrossRef]
- Jahoda, C.A.B. Cell movement in the hair follicle dermis—More than a two-way street? J. Investig. Dermatol. 2003, 121, ix–xi. [Google Scholar] [CrossRef]
- Tajran, J.; Gosman, A.A. Anatomy, Head and Neck, Scalp. StatPearls. 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK551565/ (accessed on 4 July 2025).
- Polak-Witka, K.; Rudnicka, L.; Blume-Peytavi, U.; Vogt, A. The role of the microbiome in scalp hair follicle biology and disease. Exp. Dermatol. 2020, 29, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Shibagaki, N.; Suda, W.; Clavaud, C.; Bastien, P.; Takayasu, L.; Iioka, E.; Kurokawa, R.; Yamashita, N.; Hattori, Y.; Shindo, C.; et al. Aging-related changes in the diversity of women’s skin microbiomes associated with oral bacteria. Sci. Rep. 2017, 7, 10567. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.; Mittal, P.; Clavaud, C.; Dhakan, D.B.; Roy, N.; Breton, L.; Misra, N.; Sharma, V.K. Longitudinal study of the scalp microbiome suggests coconut oil to enrich healthy scalp commensals. Sci. Rep. 2021, 11, 7220. [Google Scholar] [CrossRef]
- Park, H.K.; Ha, M.; Park, S.; Kim, M.N.; Kim, B.J.; Kim, W. Characterization of the fungal microbiota (mycobiome) in healthy and dandruff-afflicted human scalps. PLoS ONE 2012, 7, e32847. [Google Scholar] [CrossRef]
- Perez-Perez, G.I.; Gao, Z.; Jourdain, R.; Ramirez, J.; Gany, F.; Clavaud, C.; Demaude, J.; Breton, L.; Blaser, M.J. Body site is a more determinant factor than human population diversity in the healthy skin microbiome. PLoS ONE 2016, 11, e0151990. [Google Scholar] [CrossRef] [PubMed]
- Lousada, M.B.; Edelkamp, J.; Lachnit, T.; Fehrholz, M.; Jimenez, F.; Paus, R. Laser capture microdissection as a method for investigating the human hair follicle microbiome reveals region-specific differences in the bacteriome profile. BMC Res. Notes 2023, 16, 29. [Google Scholar] [CrossRef]
- Ogai, K.; Nagase, S.; Mukai, K.; Iuchi, T.; Mori, Y.; Matsue, M.; Sugitani, K.; Sugama, J.; Okamoto, S. A comparison of techniques for collecting skin microbiome samples: Swabbing versus tape-stripping. Front. Microbiol. 2018, 9, 2362. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuji, T.; Chiang, H.; Jiang, S.B.; Nagarajan, H.; Zenglar, K.; Gallo, R.L. The microbiome extends to subepidermal compartments of normal skin. Nat. Commun. 2013, 4, 1431. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, F.; Pinto, D.; Borsani, E.; Castrezzati, S.; Amedei, A.; Rezzani, R. The first evidence of bacterial foci in the hair part and dermal papilla of scalp hair follicles: A pilot comparative study in alopecia areata. Int. J. Mol. Sci. 2022, 23, 11956. [Google Scholar] [CrossRef]
- Schembri, K.; Scerri, C.; Ayers, D. Plucked human hair shafts and biomolecular medical research. Sci. World J. 2013, 2013, 620531. [Google Scholar] [CrossRef]
- Matard, B.; Meylheuc, T.; Briandet, R.; Casin, I.; Assouly, P.; Cavelier-balloy, B.; Reygagne, P. First evidence of bacterial biofilms in the anaerobe part of scalp hair follicles: A pilot comparative study in folliculitis decalvans. J. Eur. Acad. Dermatol. Venereol. 2012, 27, 853–860. [Google Scholar] [CrossRef]
- Paus, R.; Nickoloff, B.J.; Ito, T. A ‘hairy’ privilege. Trends Immunol. 2005, 26, 32–40. [Google Scholar] [CrossRef]
- Meyer, K.C.; Klatte, J.E.; Dinh, H.V.; Harries, M.J.; Reithmayer, K.; Meyer, W.; Sinclair, R.; Paus, R. Evidence that the bulge region is a site of relative immune privilege in human hair follicles. Br. J. Dermatol. 2008, 159, 1077–1085. [Google Scholar] [CrossRef]
- Cong, T.; Hao, D.; Wen, X.; Li, X.; He, G.; Jiang, X. From pathogenesis of acne vulgaris to anti-acne. Arch. Dermatol. Res. 2019, 311, 337–349. [Google Scholar] [CrossRef]
- Sabat, R.; Jemec, G.B.E.; Matusiak, Ł.; Kimball, A.B.; Prens, E.; Wolk, K. Hidradenitis suppurativa. Nat. Rev. Dis. Primers 2020, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Wilhem, K.P.; Cua, A.B.; Maibach, H.I. Skin aging. Effect on transepidermal water loss, stratum corneum hydration, skin surface pH, and causal sebum content. Arch. Dermatol. 1991, 127, 1806–1809. [Google Scholar] [CrossRef]
- Marrakchi, S.; Maibach, H.I. Biophysical parameters of skin: Map of human face, regional, and age-related differences. Contact Dermat. 2007, 57, 28–34. [Google Scholar] [CrossRef]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Functional and physiological characteristics of the aging skin. Aging Clin. Exp. Res. 2008, 20, 195–200. [Google Scholar] [CrossRef]
- Chng, K.C.; Tay, A.; Li, C.; Ng, A.; Wang, J.; Suri, B.K.; Matta, S.A.; McGovern, N.; Janela, B.; Wong, C.; et al. Whole metagenome profiling reveals skin microbiome-dependent susceptibility to atopic dermatitis flare. Nat. Microbiol. 2016, 1, 16106. [Google Scholar] [CrossRef]
- Ma, L.; Guichard, A.; Cheng, Y.; Li, J.; Qin, O.; Wang, X.; Liu, W.; Tan, Y. Sensitive scalp is associated with excessive sebum and perturbed microbiome. J. Cosmet. Dermatol. 2018, 18, 922–928. [Google Scholar] [CrossRef] [PubMed]
- Papaccio, F.; D’Arino, A.; Caputo, S.; Bellei, B. Focus on the contribution of oxidative stress in skin aging. Antioxidants 2022, 11, 1121. [Google Scholar] [CrossRef] [PubMed]
- Azzimonti, B.; Ballacchino, C.; Zanetta, P.; Cucci, M.A.; Monge, C.; Margherita, G.; Dianzani, C.; Barrera, G.; Pizzimenti, S. Microbiota, oxidative stress, and skin cancer: An unexpected triangle. Antioxidants 2023, 12, 546. [Google Scholar] [CrossRef] [PubMed]
- Krueger, A.; Mohamed, A.; Kolka, C.M.; Stoll, T.; Zaugg, J.; Linedale, R.; Morrison, M.; Soer, H.P.; Hugenholtz, P.; Frazer, I.H.; et al. Skin cancer-associated S. aureus strains can induce DNA damage in human keratinocytes by downregulating DNA repair and promoting oxidative stress. Cancers 2022, 14, 2143. [Google Scholar] [CrossRef]
- Später, S.; Hipler, U.; Haustein, U.; Nenoff, P. Generation of reactive oxygen species in vitro by Malessezia yeasts. Hautarzt 2009, 60, 122–127. [Google Scholar] [CrossRef]
- Trüeb, R.M. Oxidative stress and its impact on skin, scalp and hair. Int. J. Cosmet. Sci. 2021, 43, S9–S13. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Sweren, E.; Andrews, W.; Li, Y.; Chen, J.; Xue, Y.; Wier, E.; Alphonse, M.P.; Luo, L.; Miao, Y.; et al. Commensal microbiome promotes hair follicle regeneration by inducing keratinocyte HIF-1α signalling and glutamine metabolism. Sci. Adv. 2023, 9, eabo7555. [Google Scholar] [CrossRef]
- Šebetić, K.; Masnec, I.S.; Čavka, V.; Biljan, D.; Krolo, I. UV damage of the hair. Coll. Antropol. 2008, 32, 163–165. [Google Scholar]
- Hoting, E.; Zimmerman, M.; Höcker, H. Photochemical alterations in human hair part III: Analysis of melanin. J. Soc. Cosmet. Chem. 1995, 46, 181–190. [Google Scholar]
- Lee, W. Photoaggravation of hair aging. Int. J. Trichology 2009, 1, 94–99. [Google Scholar] [CrossRef]
- Evans, T.A. How Damaged in Hair? Part I: Surface Damage. Cosmetics & Toiletries. 2017. Available online: https://www.cosmeticsandtoiletries.com/testing/sensory/article/21836754/how-damaged-is-hair-part-i-surface-damage (accessed on 4 July 2025).
- Dias, M.F.R.G. Hair cosmetics: An overview. Int. J. Trichology 2015, 7, 2–15. [Google Scholar] [CrossRef]
- Robbins, C.R. Interactions of shampoo and crème rinse ingredients with human hair. In Chemical and Physical Behaviour of Human Hair, 4th ed.; Springer: New York, NY, USA, 2013; pp. 193–310. [Google Scholar] [CrossRef]
- La Torre, C.; Bhushan, B. Nanotribological effects of silicone type, silicone deposition level and surfactant type on human hair using atomic force microscopy. J. Cosmet. Sci. 2006, 57, 37–56. [Google Scholar]
- Mysore, V.; Arghya, A. Hair oils: Indigenous knowledge revisited. Int. J. Trichology 2022, 14, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Akowuah, G.A.; Liew, K.B.; Lee, S.; Keng, J.; Lee, S.; Yon, J.; Tan, C.S.; Chew, Y. Natural alternatives from your garden for hair care: Revisiting the benefits of tropical herbs. Heliyon 2023, 9, e21876. [Google Scholar] [CrossRef] [PubMed]
- Jeanneton, O.; Kurfurst, R.; Cavagnino, A.; Notte, D.; Gourguillon, L.; LeBlanc, E.; Nizard, C.; Baraibar, M.; Pays, K. A mixture of honeys and Royal Jelly for protection and repair of Chinese hair from UVA combined with environmental pollution [Poster presentation]. In Proceedings of the European Society for Dermatological Research 50th Annual Meeting, Amsterdam, The Netherlands, 28 September–1 October 2021. [Google Scholar]
- Kurfurst, R.; Jeanneton, O.; Cavagnino, A.; Desroches, J.; Nizard, C.; Baraibar, M.; Pays, K. A combination of honey to repair Asian hair from UV-A and environmental pollution-induced damage [Poster presentation]. In Proceedings of the First International Societies for Investigative Dermatology Meeting, Tokyo, Japan, 10–13 May 2023. [Google Scholar]
- Cohen, S. What’s New in the ‘Skinification’ of Hair Care? Cosmetics Design-Europe. 2024. Available online: https://www.cosmeticsdesign-europe.com/Article/2024/02/15/Skincare-ingredients-for-hair/ (accessed on 4 July 2025).
- Westgate, G.E.; Grohmann, D.; Moya, M.M. Hair Longevity—Evidence for a multifactorial holistic approach to managing hair aging changes. J. Clin. Med. 2025, 14, 1894. [Google Scholar] [CrossRef] [PubMed]
- Szendzielorz, E.; Spiewak, R. Caffeine as an active molecule in cosmetic products for hair loss: Its mechanisms of action in the context of hair physiology and pathology. Molecules 2025, 30, 167. [Google Scholar] [CrossRef]
- Szendzielorz, E.; Spiewak, R. Adenosine as an active ingredient in topical preparations against hair loss: A systematic review and meta-analysis of published clinical trials. Biomolecules 2025, 15, 1093. [Google Scholar] [CrossRef]
- Habeebuddin, M.; Karnati, R.K.; Shiroorkar, P.N.; Nagaraja, S.; Asdaq, S.M.B.; Anwer, M.K.; Fattepur, S. Topical probiotics: More than a skin deep. Pharmaceutics 2022, 14, 557–579. [Google Scholar] [CrossRef]
- Herbert, S.; Haughton, R.; Nava, J.; Ji-Xu, A.; Le, S.T.; Maverakis, E. A review of topical probiotic therapy for atopic dermatitis. Clin. Exp. Dermatol. 2023, 48, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Castiel, I.; Gueniche, A. Cosmetic and Dermatological Use of Probiotic Lactobacillus Paracasei Microorganisms for the Treatment of Greasy Scalp Disorders. European Patent Office EP2149368B1, 30 May 2018. Available online: https://patents.google.com/patent/EP2149368B1/en (accessed on 11 August 2025).
- Yin, C.; Nguyen, T.T.M.; Yi, E.; Zheng, S.; Bellere, A.D.; Zheng, Q.; Jin, X.; Kim, M.; Park, S.; Oh, S.; et al. Efficacy of probiotics in hair growth and dandruff control: A systematic review and meta-analysis. Heliyon 2024, 10, e29539. [Google Scholar] [CrossRef]
- Woo, Y.M.; Seo, J.Y.; Kim, S.; Cha, J.H.; Cho, H.D.; Cha, Y.K.; Jeong, J.T.; Park, S.M.; Ryu, H.S.; Kim, J.M.; et al. Hair growth effect of TS-SCLF from Schisandra chinensis extract fermented with Lactobacillus plantarum. Microbiol. Biotechnol. Lett. 2022, 50, 533–547. [Google Scholar] [CrossRef]
- de Jesus, G.F.A.; Rossetto, M.P.; Voytena, A.; Feder, B.; Borges, H.; da Costa Borges, G.; Feuser, Z.P.; Dal-Bó, S.; Michels, M. Clinical evaluation of paraprobiotic-associated Bifidobacterium lactis CCT 7858 anti-dandruff shampoo efficacy: A randomized placebo-controlled clinical trial. Int. J. Cosmet. Sci. 2023, 45, 572–580. [Google Scholar] [CrossRef]
- Tsai, W.; Fang, Y.; Huang, T.; Chiang, Y.; Lin, C.; Chang, W. Heat-killed Lacticaseibacillus paracasei GMNL-653 ameliorates human scalp health by regulating scalp microbiome. BMC Microbiol. 2023, 23, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Mori-Ichioka, A.; Sunada, Y.; Koikeda, T.; Matsuda, H.; Matsuo, S. Effect of applying Lactiplantibacillus plantarum subsp. plantarum N793 to the scalps of men and women with thinning hair: A randomized, double-blind, placebo-controlled, parallel-group study. Biosci. Microbiota Food Health 2024, 43, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Kesika, P.; Sivamaruthi, B.S.; Thangaleela, S.; Bharathi, M.; Chaiyasut, C. Role and mechanisms of phytochemicals in hair growth and health. Pharmaceuticals 2023, 16, 206–228. [Google Scholar] [CrossRef]
- Babich, O.; Ivanova, S.; Bakhtiyarova, A.; Kalashnikova, O.; Sukhikh, S. Medicinal plants are the basis of natural cosmetics. Process Biochem. 2025, 154, 35–51. [Google Scholar] [CrossRef]
- Park, S.; Lee, J. Modulation of hair growth promoting effect by natural products. Pharmaceutics 2021, 13, 2163. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Boo, M.Y.; Boo, Y.C. Can plant extracts help prevent hair loss or promote hair growth? A review comparing their therapeutic efficacies, phytochemical components, and modulatory targets. Molecules 2024, 29, 2288. [Google Scholar] [CrossRef]
- Sennett, R.; Wang, Z.; Rezza, A.; Grisanti, L.; Roitershtein, N.; Sicchio, C.; Mok, K.W.; Heitman, N.J.; Clavel, C.; Ma’ayan, A.; et al. An integrated transcriptome atlas of embryonic hair follicle progenitors, their niche, and the developing skin. Dev. Cell 2015, 34, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Liang, A.; Fang, Y.; Ye, L.; Meng, J.; Wang, X.; Chen, J.; Xu, X. Signaling pathways in hair aging. Front. Cell Dev. Biol. 2023, 11, 1278278. [Google Scholar] [CrossRef] [PubMed]
- Forraz, N.; Kurfurst, R.; Jeanneton, O.; Desroches, A.; Payen, P.; Milet, C.; Vialle, A.; Choisy, P.; Nizard, C.; Pays, K.; et al. Stimulation of the KLF4 pathway by bee products modulates the progression of hair anagen to telogen molecular switch [Paper presentation]. In Proceedings of the International Federation of Societies of Cosmetic Scientists 2022 Congress, London, UK, 19–22 September 2022. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).