
A Mn (II) Quinoline Complex (4QMn) Mitigates Oxidative Damage Induced by Ultraviolet Radiation and Protein Aggregation

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
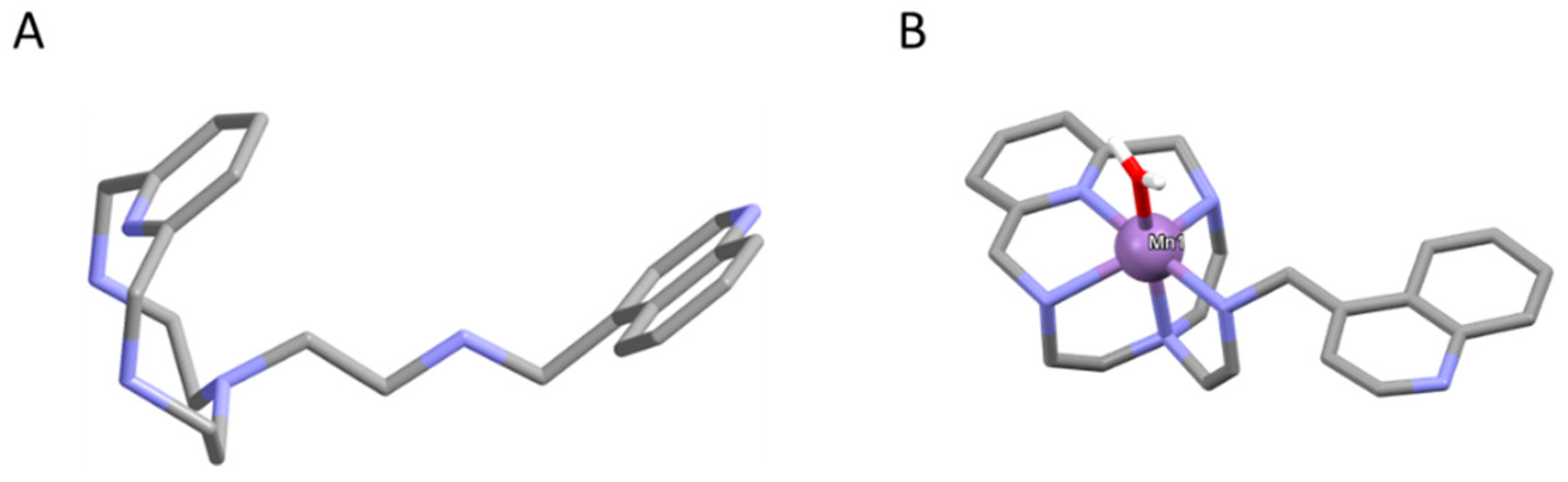
2.1. 4QMn Compound
2.2. Cell Culture
2.3. Cell Transfection
2.4. Human Skin Explants
2.5. MTT Assay
2.6. UVA Irradiation
2.7. ROS Quantification
2.8. Gene Expression Analysis (qRT-PCR)
2.9. MitoROS Quantification
2.10. 8-Hydroxy-2′-deoxyguanosine (8-OHdG) Quantification
2.11. Statistical Analysis
3. Results
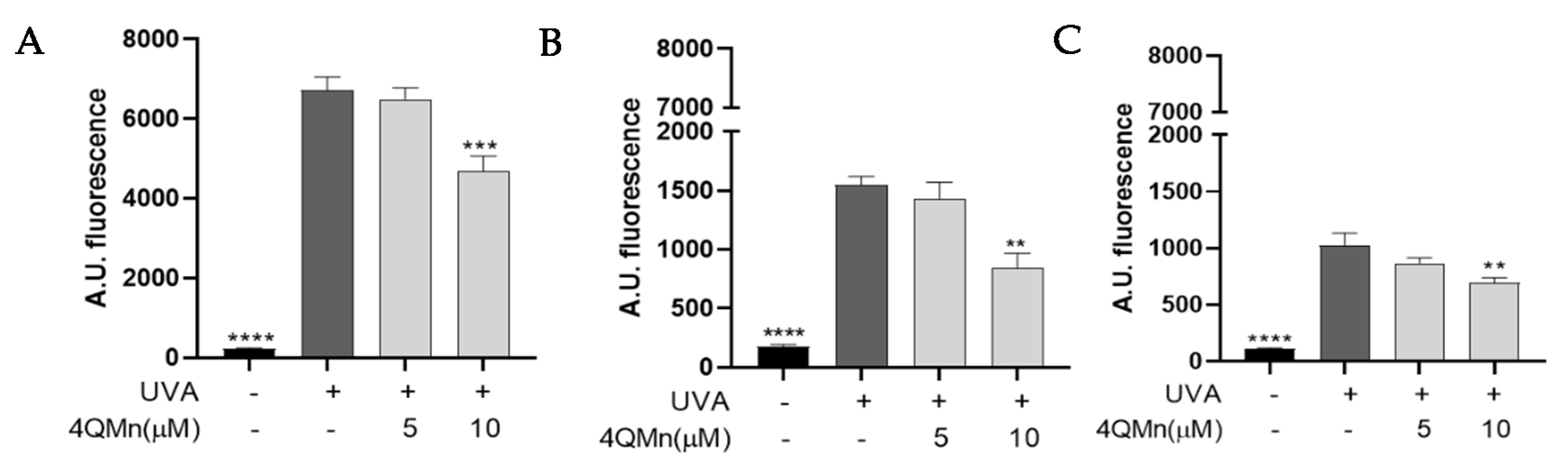
3.1. Evaluation of 4QMn Antioxidant Capacity by Quantification of UVA-Induced ROS in Human Skin Cells and Skin Explants
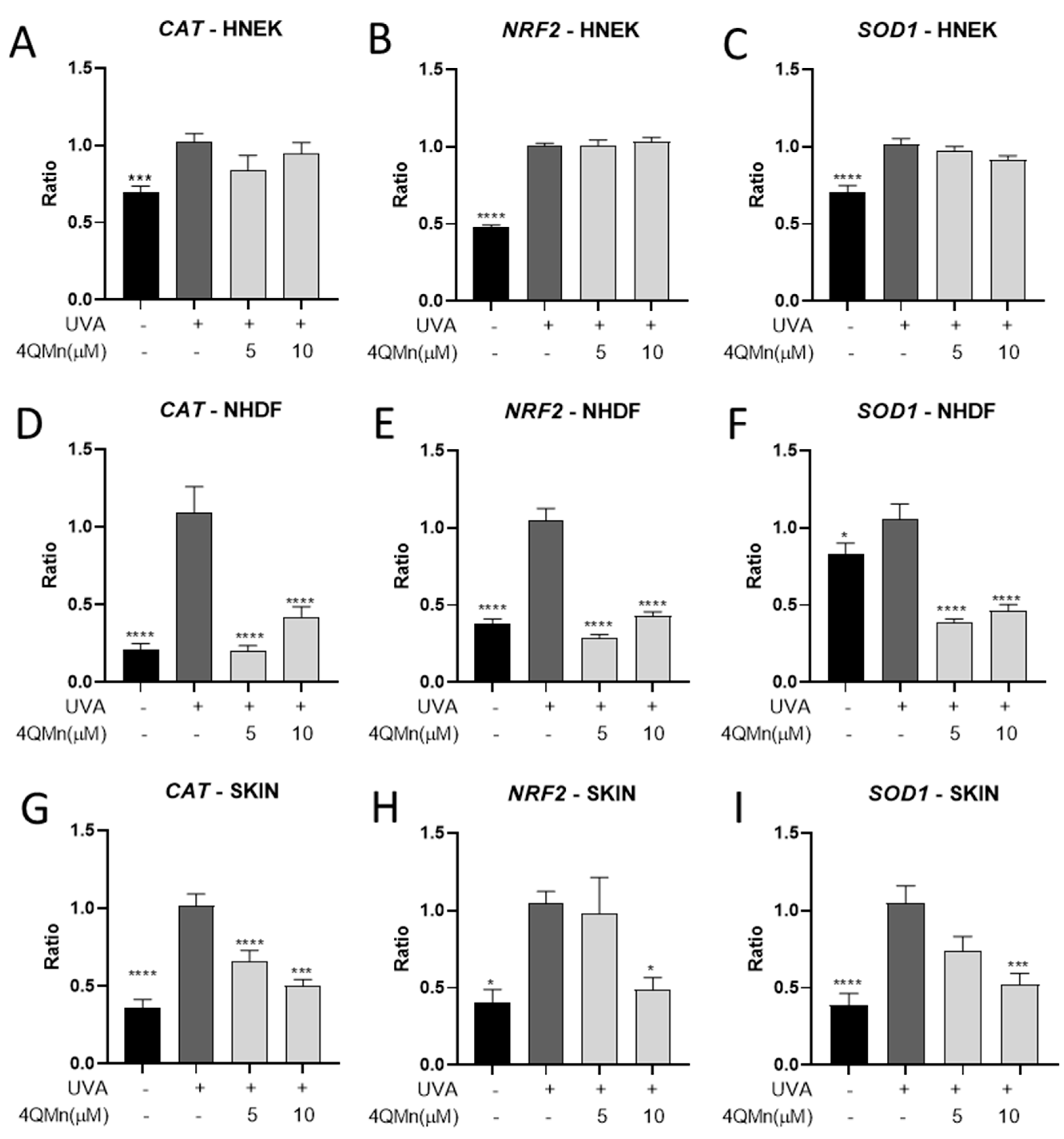
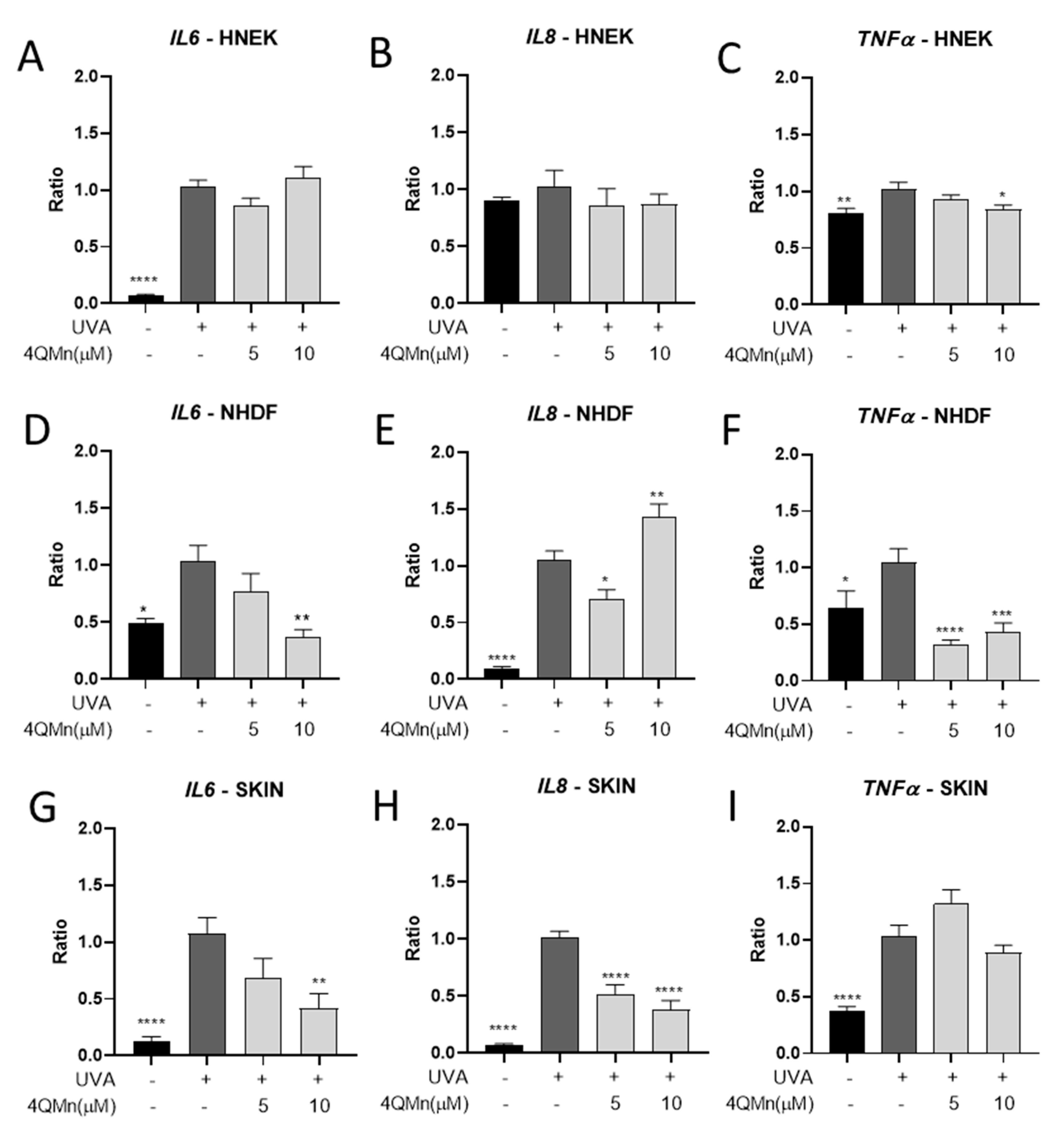
3.2. Gene Expression Analysis after 4QMn Treatment in Human Skin Cells and Skin Explants following UVA Radiation
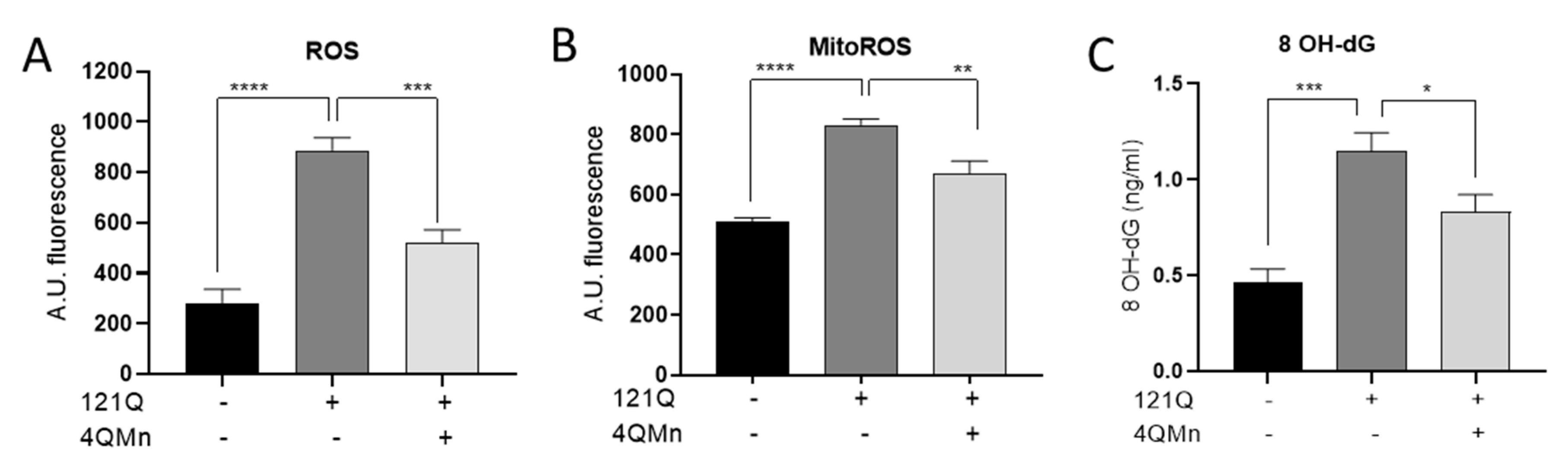
3.3. Oxidative Damage Assessment after 4QMn Treatment in an In Vitro Model of Protein Aggregation
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prost-Squarcioni, C. Histology of skin and hair follicle. Med. Sci. 2006, 22, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Arda, O.; Göksügür, N.; Tüzün, Y. Basic histological structure and functions of facial skin. Clin. Dermatol. 2014, 32, 3–13. [Google Scholar] [CrossRef]
- Lopez-Ojeda, W.; Pandey, A.; Alhajj, M.; Oakley, A.M. Anatomy, Skin (Integument). In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK441980/ (accessed on 4 January 2024).
- Graham, H.K.; Eckersley, A.; Ozols, M.; Mellody, K.T.; Sherratt, M.J. Human Skin: Composition, Structure and Visualisation Methods. In Skin Biophysics: From Experimental Characterisation to Advanced Modelling; Limbert, G., Ed.; Studies in Mechanobiology, Tissue Engineering and Biomaterials; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–18. ISBN 978-3-030-13279-8. [Google Scholar]
- Yousef, H.; Alhajj, M.; Sharma, S. Anatomy, Skin (Integument), Epidermis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Amaro-Ortiz, A.; Yan, B.; D’Orazio, J.A. Ultraviolet Radiation, Aging and the Skin: Prevention of Damage by Topical cAMP Manipulation. Molecules 2014, 19, 6202–6219. [Google Scholar] [CrossRef]
- Polefka, T.G.; Meyer, T.A.; Agin, P.P.; Bianchini, R.J. Effects of solar radiation on the skin. J. Cosmet. Dermatol. 2012, 11, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Ichihashi, M.; Ando, H.; Yoshida, M.; Niki, Y.; Matsui, M. Photoaging of the skin. Anti-Aging Med. 2009, 6, 46–59. [Google Scholar] [CrossRef]
- Diffey, B.L. Human exposure to solar ultraviolet radiation. J. Cosmet. Dermatol. 2002, 1, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Juzeniene, A.; Moan, J. Beneficial effects of UV radiation other than via vitamin D production. Dermato-Endocrinology 2012, 4, 109–117. [Google Scholar] [CrossRef]
- Shin, D.W. Various biological effects of solar radiation on skin and their mechanisms: Implications for phototherapy. Anim. Cells Syst. 2020, 24, 181–188. [Google Scholar] [CrossRef]
- Vierkötter, A.; Krutmann, J. Environmental influences on skin aging and ethnic-specific manifestations. Dermato-Endocrinology 2012, 4, 227–231. [Google Scholar] [CrossRef]
- Neale, R.E.; Lucas, R.M.; Byrne, S.N.; Hollestein, L.; Rhodes, L.E.; Yazar, S.; Young, A.R.; Berwick, M.; Ireland, R.A.; Olsen, C.M. The effects of exposure to solar radiation on human health. Photochem. Photobiol. Sci. 2023, 22, 1011–1047. [Google Scholar] [CrossRef]
- Merino, M.; Mullor, J.L.; Sánchez-Sánchez, A.V. Medaka (Oryzias latipes) Embryo as a Model for the Screening of Compounds That Counteract the Damage Induced by Ultraviolet and High-Energy Visible Light. Int. J. Mol. Sci. 2020, 21, 5769. [Google Scholar] [CrossRef] [PubMed]
- Squier, T.C. Oxidative stress and protein aggregation during biological aging. Exp. Gerontol. 2001, 36, 1539–1550. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, K.; Dewan, R.; Kumar, J.V.; Dewan, A. Primary Cutaneous Amyloidosis: A Clinical, Histopathological and Immunofluorescence Study. J. Clin. Diagn. Res. 2017, 11, WC01–WC05. [Google Scholar] [CrossRef] [PubMed]
- Kowalska-Kępczyńska, A. Systemic Scleroderma—Definition, Clinical Picture and Laboratory Diagnostics. J. Clin. Med. 2022, 11, 2299. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.N.; Kim, S.-J. Dermatitis Herpetiformis: An Update on Diagnosis, Disease Monitoring, and Management. Medicina 2021, 57, 843. [Google Scholar] [CrossRef] [PubMed]
- Svobodová, A.; Vostálová, J. Solar radiation induced skin damage: Review of protective and preventive options. Int. J. Radiat. Biol. 2010, 86, 999–1030. [Google Scholar] [CrossRef] [PubMed]
- Biskanaki, F.; Kefala, V.; Lazaris, A.C.; Rallis, E. Aging and the Impact of Solar Ultraviolet Radiation on the Expression of Type I and Type VI Collagen. Cosmetics 2023, 10, 48. [Google Scholar] [CrossRef]
- Garnacho Saucedo, G.M.; Salido Vallejo, R.; Moreno Giménez, J.C. Effects of solar radiation and an update on photoprotection. An. Pediatría 2020, 92, 377.e1–377.e9. [Google Scholar] [CrossRef]
- Clares, M.P.; Blasco, S.; Inclán, M.; del Castillo Agudo, L.; Verdejo, B.; Soriano, C.; Doménech, A.; Latorre, J.; García-España, E. Manganese(II) complexes of scorpiand-like azamacrocycles as MnSOD mimics. Chem. Commun. 2011, 47, 5988–5990. [Google Scholar] [CrossRef]
- Clares, M.P.; Serena, C.; Blasco, S.; Nebot, A.; del Castillo, L.; Soriano, C.; Domènech, A.; Sánchez-Sánchez, A.V.; Soler-Calero, L.; Mullor, J.L.; et al. Mn(II) complexes of scorpiand-like ligands. A model for the MnSOD active centre with high in vitro and in vivo activity. J. Inorg. Biochem. 2015, 143, 1–8. [Google Scholar] [CrossRef]
- Serena, C.; Calvo, E.; Clares, M.P.; Diaz, M.L.; Chicote, J.U.; Beltrán-Debon, R.; Fontova, R.; Rodriguez, A.; García-España, E.; García-España, A. Significant in vivo anti-inflammatory activity of Pytren4Q-Mn a superoxide dismutase 2 (SOD2) mimetic scorpiand-like Mn (II) complex. PLoS ONE 2015, 10, e0119102. [Google Scholar] [CrossRef] [PubMed]
- Merino, M.; Sequedo, M.D.; Sánchez-Sánchez, A.V.; Clares, M.P.; García-España, E.; Vázquez-Manrique, R.P.; Mullor, J.L. Mn(II) Quinoline Complex (4QMn) Restores Proteostasis and Reduces Toxicity in Experimental Models of Huntington’s Disease. Int. J. Mol. Sci. 2022, 23, 8936. [Google Scholar] [CrossRef] [PubMed]
- Ryšavá, A.; Čížková, K.; Franková, J.; Roubalová, L.; Ulrichová, J.; Vostálová, J.; Vrba, J.; Zálešák, B.; Rajnochová Svobodová, A. Effect of UVA radiation on the Nrf2 signalling pathway in human skin cells. J. Photochem. Photobiol. B 2020, 209, 111948. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Ansary, T.M.; Hossain, M.d.R.; Kamiya, K.; Komine, M.; Ohtsuki, M. Inflammatory Molecules Associated with Ultraviolet Radiation-Mediated Skin Aging. Int. J. Mol. Sci. 2021, 22, 3974. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.-C.; Xiao, T.; Chen, Y.-J. Ultraviolet Induced Skin Inflammation. Int. J. Dermatol. Venereol. 2021, 4, 229–235. [Google Scholar] [CrossRef]
- Hruza, L.L.; Pentland, A.P. Mechanisms of UV-Induced Inflammation. J. Investig. Dermatol. 1993, 100, S35–S41. [Google Scholar] [CrossRef]
- Singh, A.; Willems, E.; Singh, A.; Hafeez, B.B.; Ong, I.M.; Mehta, S.L.; Verma, A.K. Ultraviolet radiation-induced tumor necrosis factor alpha, which is linked to the development of cutaneous SCC, modulates differential epidermal microRNAs expression. Oncotarget 2016, 7, 17945–17956. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Gao, J.; Dinh, Q.T.; Chen, C.; Fimmel, S. IL-8 production and AP-1 transactivation induced by UVA in human keratinocytes: Roles of d-α-tocopherol. Mol. Immunol. 2008, 45, 2288–2296. [Google Scholar] [CrossRef]
- Schneider, L.A.; Raizner, K.; Wlaschek, M.; Brenneisen, P.; Gethöffer, K.; Scharffetter-Kochanek, K. UVA-1 exposure in vivo leads to an IL-6 surge within the skin. Exp. Dermatol. 2017, 26, 830–832. [Google Scholar] [CrossRef]
- Widmer, R.; Ziaja, I.; Grune, T. Protein oxidation and degradation during aging: Role in skin aging and neurodegeneration. Free Radic. Res. 2006, 40, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Lopes, C.; Ferreira, I.L.; Maranga, C.; Beatriz, M.; Mota, S.I.; Sereno, J.; Castelhano, J.; Abrunhosa, A.; Oliveira, F.; Rosa, M.D.; et al. Mitochondrial and redox modifications in early stages of Huntington’s disease. Redox Biol. 2022, 56, 102424. [Google Scholar] [CrossRef]
- Goyeneche, A.A.; Garcia de Alba Graue, P.; Mastromonaco, C.; McDonald, M.; Burnier, J.V.; Burnier, M.N. Distinctive responses of keratinocytes and fibroblasts to sunlight-induced DNA damage. Investig. Ophthalmol. Vis. Sci. 2020, 61, 4671. [Google Scholar]
- Izykowska, I.; Cegielski, M.; Gebarowska, E.; Podhorska-Okolow, M.; Piotrowska, A.; Zabel, M.; Dziegiel, P. Effect of Melatonin on Human Keratinocytes and Fibroblasts Subjected to UVA and UVB Radiation In Vitro. In Vivo 2009, 23, 739–745. [Google Scholar] [PubMed]
- Tuite, M.F.; Melki, R. Protein Misfolding and Aggregation in Ageing and Disease. Prion 2007, 1, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Jones, R. Protein Aggregation Increases with Age. PLOS Biol. 2010, 8, e1000449. [Google Scholar] [CrossRef] [PubMed]
- Ando, Y.; Nyhlin, N.; Suhr, O.; Holmgren, G.; Uchida, K.; el Sahly, M.; Yamashita, T.; Terasaki, H.; Nakamura, M.; Uchino, M.; et al. Oxidative stress is found in amyloid deposits in systemic amyloidosis. Biochem. Biophys. Res. Commun. 1997, 232, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Ando, Y. Amyloidosis and oxidative stress. Rinsho Byori 2003, 51, 140–145. [Google Scholar]
- Ando, Y.; Suhr, O.; el-Salhy, M. Oxidative stress and amyloidosis. Histol. Histopathol. 1998, 13, 845–850. [Google Scholar] [CrossRef]
- Bertino, L.; Guarneri, F.; Cannavò, S.P.; Casciaro, M.; Pioggia, G.; Gangemi, S. Oxidative Stress and Atopic Dermatitis. Antioxidants 2020, 9, 196. [Google Scholar] [CrossRef] [PubMed]
- Merino, M.; González, S.; Tronch, M.C.; Sánchez-Sánchez, A.V.; Clares, M.P.; García-España, A.; García-España, E.; Mullor, J.L. Small Molecule Pytren-4QMn Metal Complex Slows down Huntington’s Disease Progression in Male zQ175 Transgenic Mice. Int. J. Mol. Sci. 2023, 24, 15153. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merino, M.; González, S.; Clares, M.P.; García-España, E.; Mullor, J.L. A Mn (II) Quinoline Complex (4QMn) Mitigates Oxidative Damage Induced by Ultraviolet Radiation and Protein Aggregation. Cosmetics 2024, 11, 95. https://doi.org/10.3390/cosmetics11030095
Merino M, González S, Clares MP, García-España E, Mullor JL. A Mn (II) Quinoline Complex (4QMn) Mitigates Oxidative Damage Induced by Ultraviolet Radiation and Protein Aggregation. Cosmetics. 2024; 11(3):95. https://doi.org/10.3390/cosmetics11030095
Chicago/Turabian StyleMerino, Marián, Sonia González, Mª Paz Clares, Enrique García-España, and José L. Mullor. 2024. "A Mn (II) Quinoline Complex (4QMn) Mitigates Oxidative Damage Induced by Ultraviolet Radiation and Protein Aggregation" Cosmetics 11, no. 3: 95. https://doi.org/10.3390/cosmetics11030095
APA StyleMerino, M., González, S., Clares, M. P., García-España, E., & Mullor, J. L. (2024). A Mn (II) Quinoline Complex (4QMn) Mitigates Oxidative Damage Induced by Ultraviolet Radiation and Protein Aggregation. Cosmetics, 11(3), 95. https://doi.org/10.3390/cosmetics11030095