
Disturbances in the Skin Homeostasis: Wound Healing, an Undefined Process




Abstract
1. Introduction
2. General Morphological Characteristics of the Skin
3. Functions of the Skin
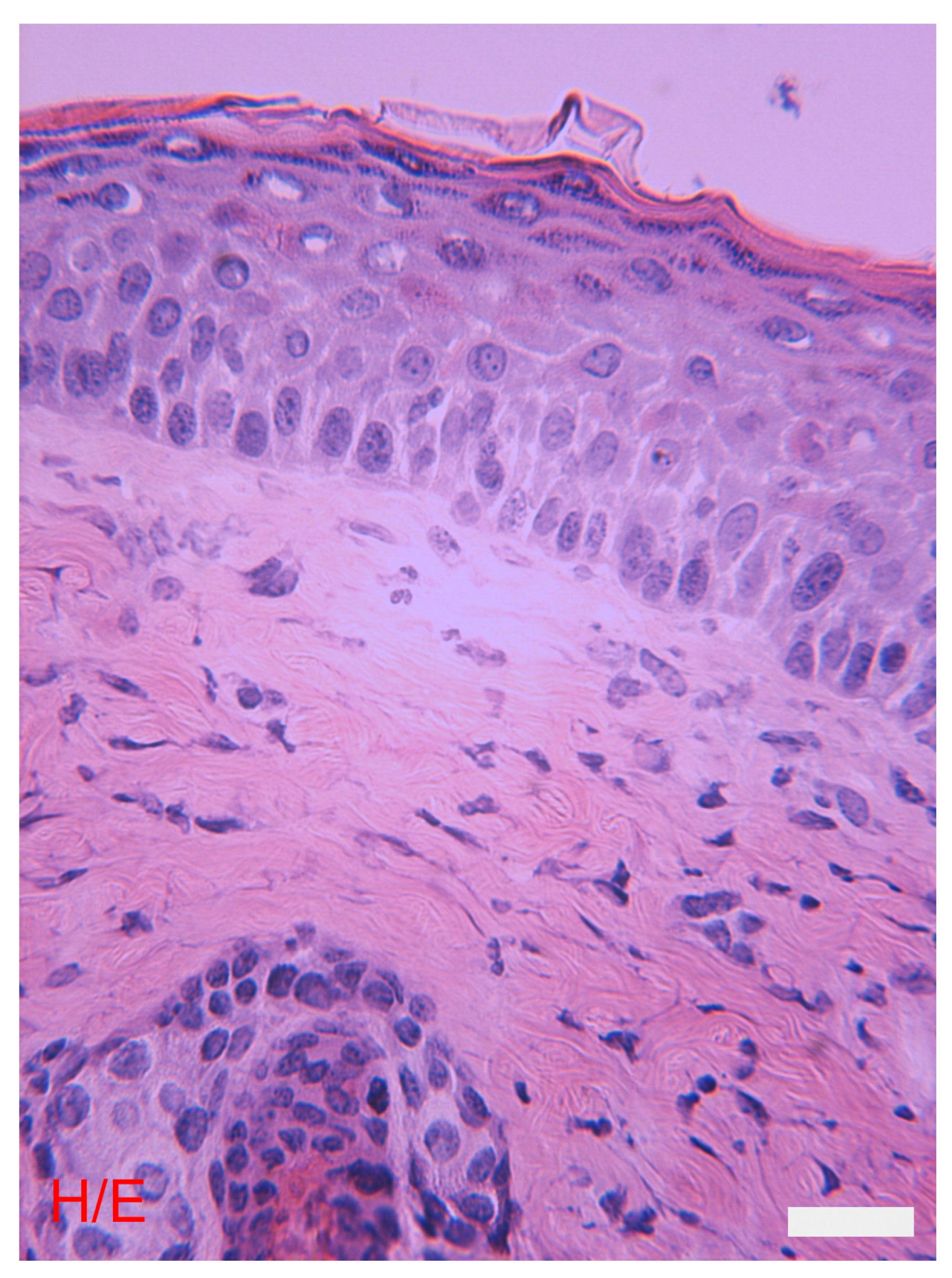
4. The Epidermis
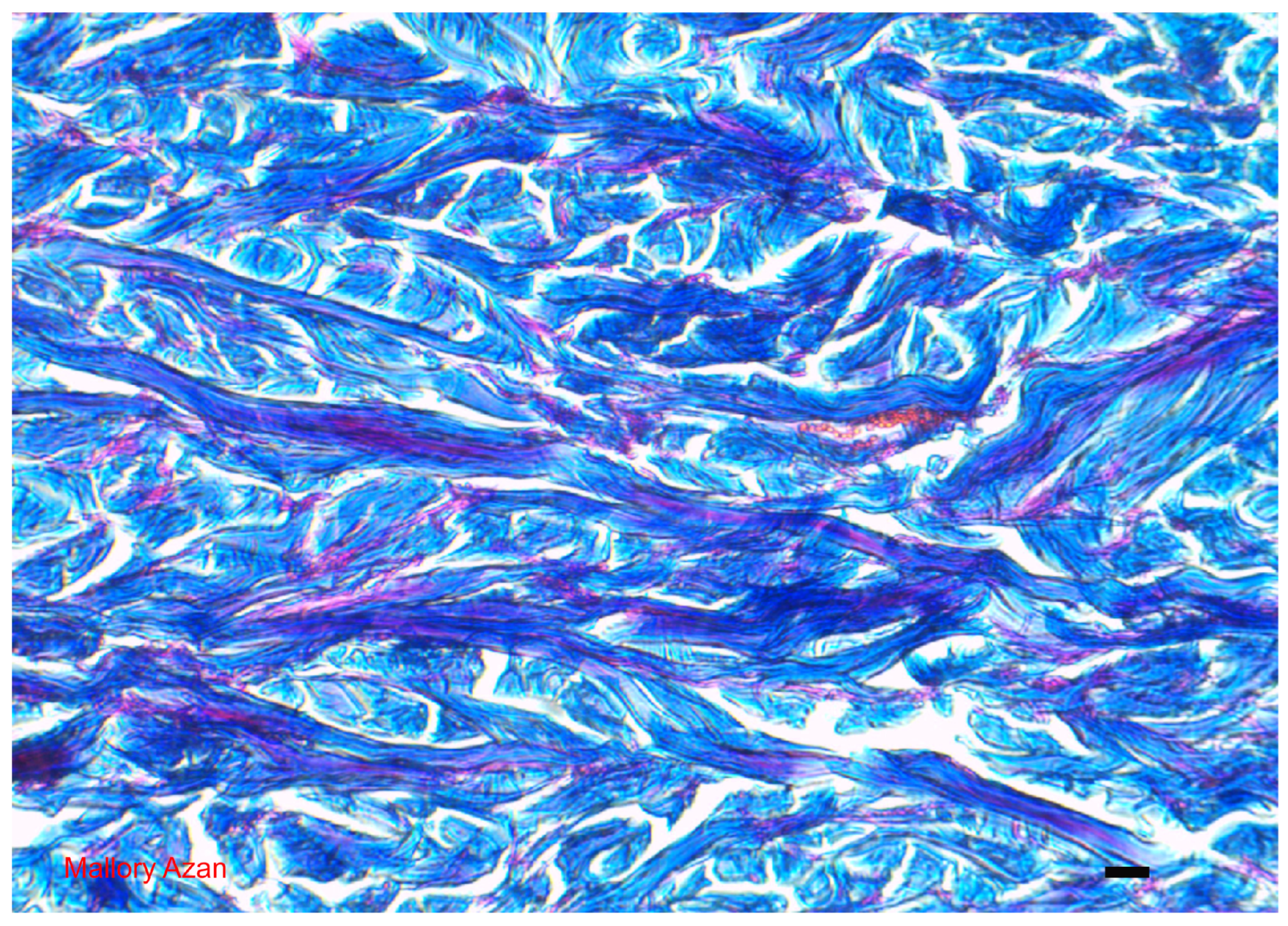
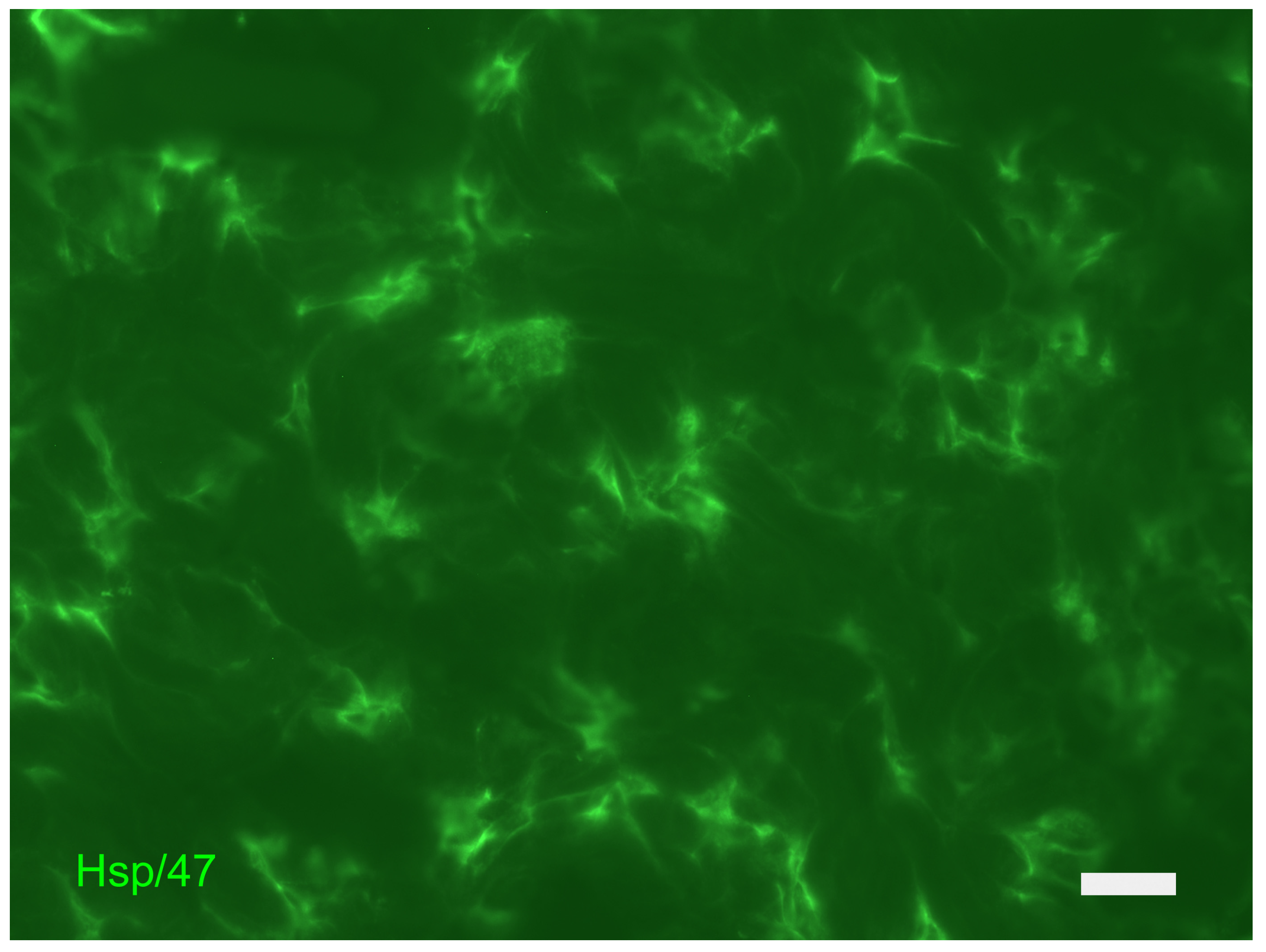
5. The Dermis
6. Subcutaneous
7. Vascularization and Innervation of the Skin
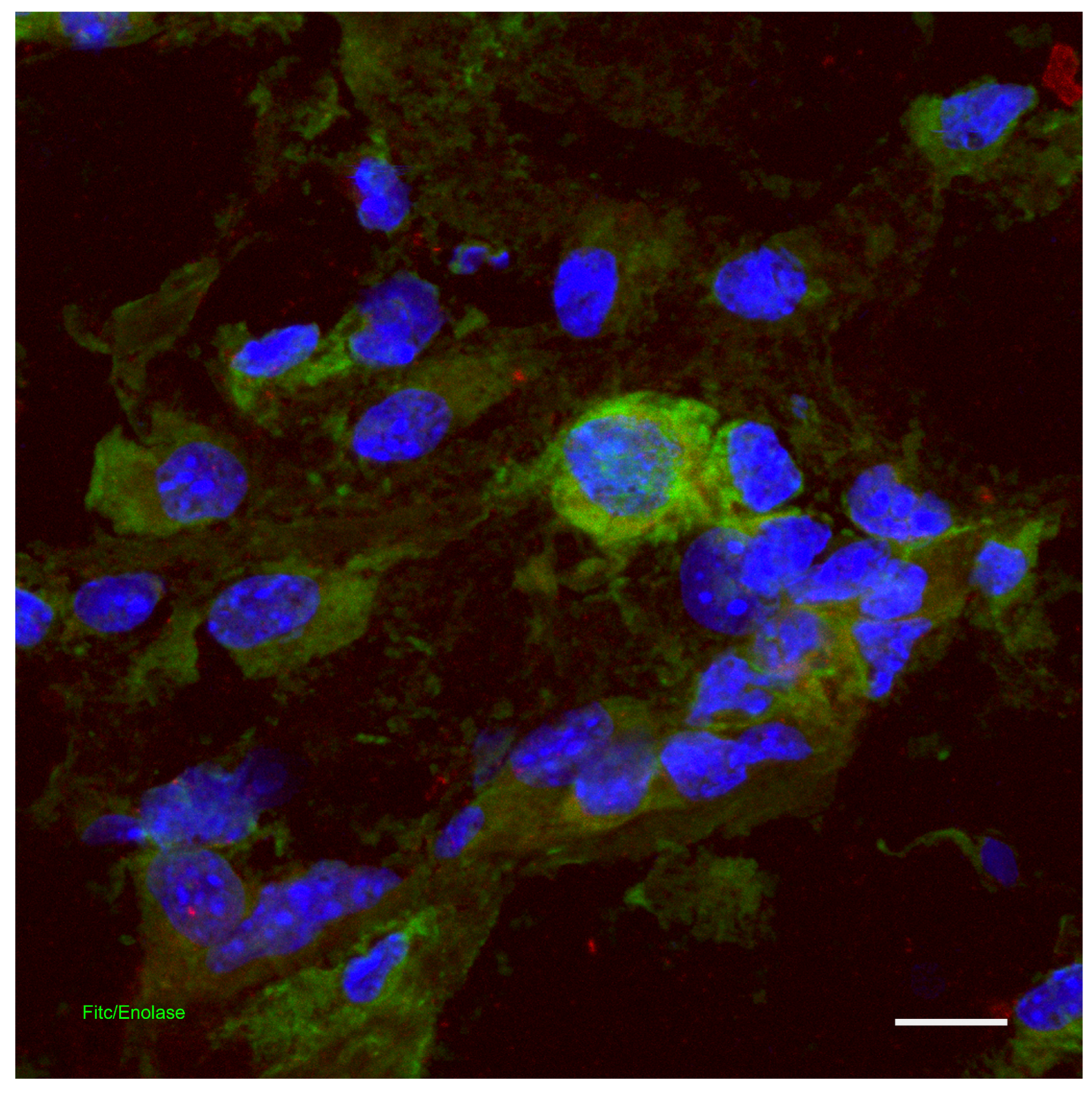
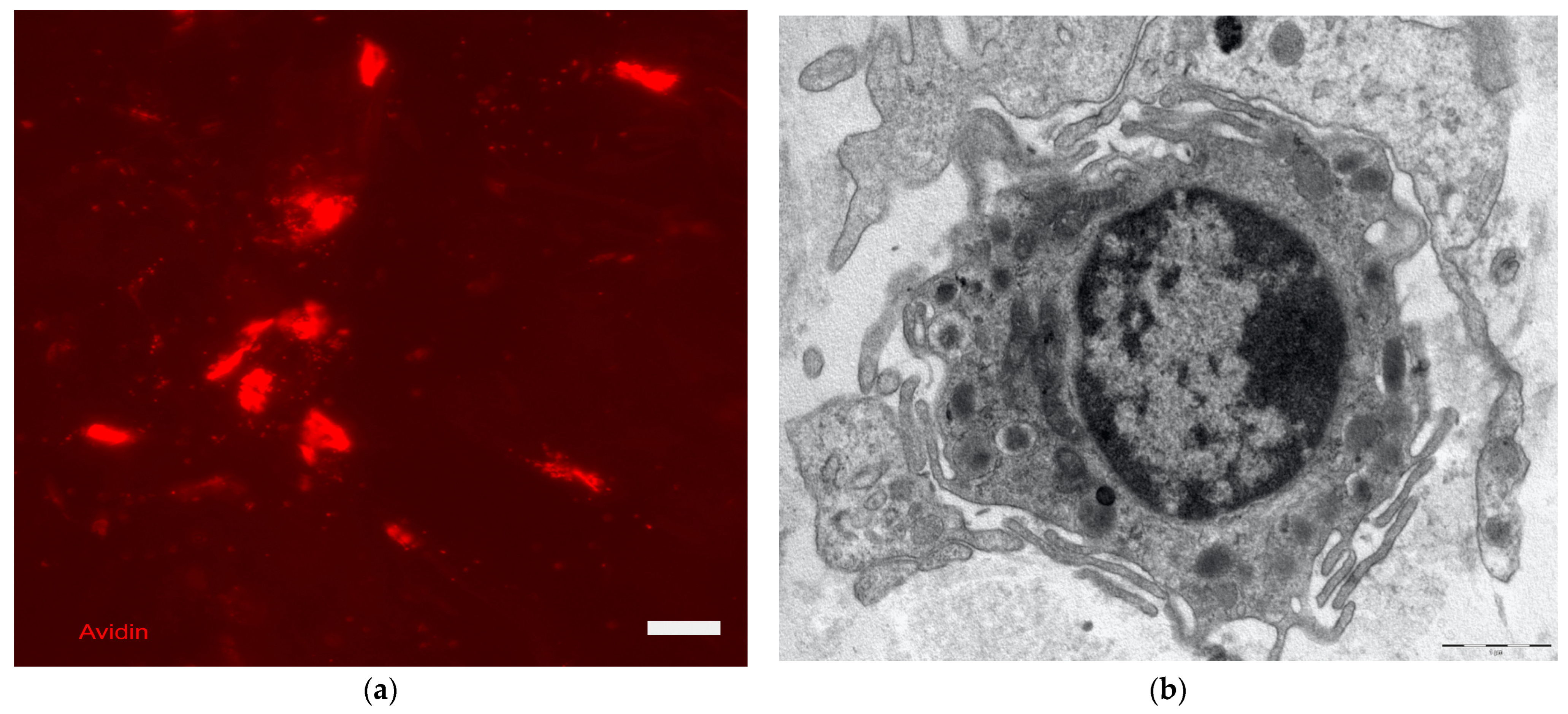
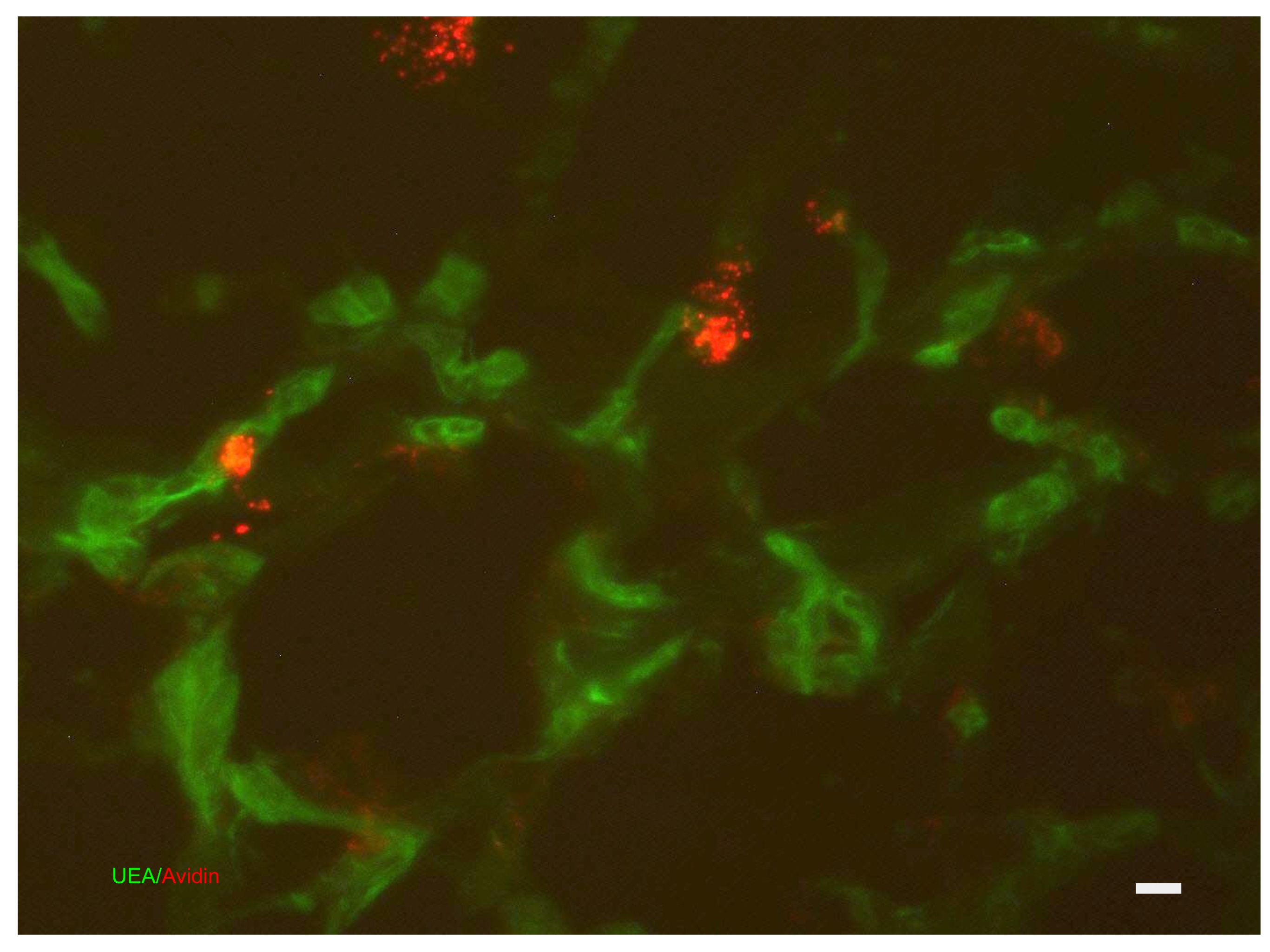
8. Skin Immune System
9. Skin pH
10. Skin Development
11. Disturbances in the Skin Homeostasis: The History of Wound Healing
11.1. Historical Aspect of a Wound
11.2. Issues Related to the Study and Management of Wound Healing
11.3. Vertebrates and Wound Healing
11.4. Wound Healing Modalities
11.5. Macroscopic Evidence of Wound Healing
- (a)
- One to three days following the injury: this phase is characterized by the formation of a primary clot (blood clot), activation of the epidermal boundaries, and an early inflammatory response.
- (b)
- Four to seven days following the injury: this stage is characterized morphologically by the development of crusts. Histological examination shows epidermal edge migration, selective proliferation of early granulation tissue, and an inflammatory response with a high concentration of macrophages and lymphocytes.
- (c)
- Eight–twelve days after the injury: morphologic investigations show that the crust has detached. Histological results reveal the development of a new epidermis, which by day 12 begins to differentiate. Furthermore, the initiation of cutaneous closure occurs simultaneously with the creation of granulation tissue. The attenuation of the inflammatory response coincides with this phase.
- (d)
11.6. Acute Inflammatory Reaction
11.7. Summary of Principal Events in Wound Healing
11.8. The Scar
11.9. Differences with Fetal Wound Healing
12. Alterations in the Normal Wound Healing Process
12.1. Chronic Skin Lesions
12.2. Keloids
12.3. Hypertrophic Scars
12.4. Wound Dressings
13. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Denomination | Acronym |
| Mast Cell (s) | MC |
| Dendritic cell (s) | DC |
| Langerhans cell (s) | LC |
| Wound Healing | WH |
| Chronic Wounds | CW |
| Matrix Metalloproteinases | MMPs |
| Tissue Inhibitors of Metalloproteinases | TIMP |
| Platelet Derived Growth Factor | PDGF |
| Interleukin | IL |
| Tumor Necrosis Factor | TNF |
| Fibroblast Growth Factor | FGF |
| Granulocyte-Macrophage Colony Stimulating Factor | GM-CSF |
| Nitric Oxide | NO |
| Transforming Growth Factor | TGF |
| Glycosaminoglycans | GAGs |
| Extracellular Matrix | ECM |
| Nerve Growth Factor | NGF |
| Hepatocyte Growth Factor | HGF |
| Heparin-Binding Epidermal Growth Factor | HB-EGF |
| Vascular Endothelial Growth Factor | VEGF |
| Calcitonin Gene Related Peptide | CGRP |
| Neurokinin A | NKA |
| Neuropeptide Y | NPY |
| Substance P | SP |
| Protein Gene Product 9.5 | PGP 9.5 |
| Vaso Active Intestinal Peptide | VIP |
References
- Joey, E.; Cheong, L.; McGrath, J.A. Structure and function of skin, hair and nails. Medicine 2021, 49, 337–342. [Google Scholar] [CrossRef]
- Fore, J. A review of skin and the effects of aging on skin structure and function. Ostomy/Wound Manag. 2006, 52, 24–37. [Google Scholar]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Structural characteristics of the aging skin: A review. Cutan. Ocul. Toxicol. 2007, 26, 343–357. [Google Scholar] [CrossRef]
- Sorg, H.; Sorg, C.G.G. Skin wound healing: Of players, patterns, and processes. Eur. Surg. Res. 2023, 64, 141–157. [Google Scholar] [CrossRef]
- Wong, R.; Geyer, S.; Weninger, W.; Guimberteau, J.C.; Wong, J.K. The dynamic anatomy and patterning of skin. Exp. Dermatol. 2016, 25, 92–98. [Google Scholar] [CrossRef]
- Kolarsick, P.A.J.; Kolarsick, M.A.; Goodwin, C. Anatomy and physiology of the skin. J. Dermatol. Nurses Assoc. 2011, 3, 203–213. [Google Scholar] [CrossRef]
- Furuse, M.; Hata, M.; Furuse, K.; Yoshida, Y.; Haratake, A.; Sugitani, Y.; Noda, T.; Kubo, A.; Tsukita, S. Claudin-based tight junctions are crucial for the mammalian epidermal barrier: A lesson from claudin-1-deficient mice. J. Cell Biol. 2002, 156, 1099–1111. [Google Scholar] [CrossRef]
- Madison, K.C. Barrier function of the skin: “la raison d’être” of the epidermis. J. Investig. Dermatol. 2003, 121, 231–241. [Google Scholar] [CrossRef]
- Johansson, J.A.; Headon, D.J. Regionalisation of the skin. Semin. Cell Dev. Biol. 2014, 26, 3–10. [Google Scholar] [CrossRef]
- Mc Grath, J.A.; Llitto, J. Anatomy and organization of human skin. In Rook’s Textbook of Dermatology, 8th ed.; Burns, T., Breathnach, S., Cox, N., Griffiths, C., Eds.; Wiley Blachwell: Hoboken, NY, USA, 2010; Chapter 3. [Google Scholar]
- Fernandez-Flores, A. Regional variations in the histology of the skin. Am. J. Dermatopathol. 2015, 37, 737–754. [Google Scholar] [CrossRef]
- Khavkin, J.; Ellis, D.A. Aging skin: Histology, physiology, and pathology. Facial Plast. Surg. Clin. N. Am. 2015, 19, 229–234. [Google Scholar] [CrossRef]
- Laverdet, B.; Danigo, A.; Girard, D.; Magy, L.; Demiot, C.; Desmoulière, A. Skin innervation: Important roles during normal and pathological cutaneous repair. Histol. Histopathol. 2015, 30, 875–892. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, M.; Baguneid, M.; Bayat, A. The role of neuromediators and innervation in cutaneous wound healing. Acta Derm.-Venereol. 2016, 96, 587–594. [Google Scholar] [CrossRef]
- Nguyen, A.V.; Soulika, A.M. The dynamics of the skin’s immune system. Int. J. Mol. Sci. 2019, 20, 1811. [Google Scholar] [CrossRef] [PubMed]
- Quaresma, J.A.S. Organization of the skin immune system and compartmentalized immune responses in infectious diseases. Clin. Microbiol. Rev. 2019, 10, 1128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Merana, G.R.; Harris-Tryon, T.; Scharschmidt, T.C. Skin immunity: Dissecting the complex biology of our body’s outer barrier. Mucosal Immunol. 2022, 15, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Zanna, M.Y.; Yasmin, A.R.; Omar, A.R.; Arshad, S.S.; Mariatulqabtiah, A.R.; Nur-Fazila, S.H.; Mahiza, M.I.N. Review of dendritic cells, their role in clinical immunology, and distribution in various animal species. Int. J. Mol. Sci. 2021, 22, 8044. [Google Scholar] [CrossRef] [PubMed]
- Focardi, M.; Puliti, E.; Grifoni, R.; Palandri, M.; Bugelli, V.; Pinchi, V.; Norelli, G.A.; Bacci, S. Immunohistochemical localization of Langerhans cells as a tool for vitality in hanging mark wounds: A pilot study. Aust. J. Forensic Sci. 2020, 52, 393–405. [Google Scholar] [CrossRef]
- Bacci, S.; Defraia, B.; Cinci, L.; Calosi, L.; Guasti, D.; Pieri, L.; Lotti, V.; Bonelli, A.; Romagnoli, P. Immunohistochemical analysis of dendritic cells in skin lesions: Correlations with survival time. Forensic Sci. Int. 2014, 244, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Bacci, S. Fine regulation during wound healing by mast cells, a physiological role not yet clarified. Int. J. Mol. Sci. 2022, 23, 1820. [Google Scholar] [CrossRef]
- Martel, J.L.; Miao, J.H.; Badri, T. Anatomy, Hair Follicle. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2024. Available online: https://www.ncbi.nlm.nih.gov/books/NBK470321/ (accessed on 29 May 2024).
- Lousada, M.B.; Lachnit, T.; Edelkamp, J.; Rouillé, T.; Ajdic, D.; Uchida, Y.; Di Nardo, A.; Bosch, T.C.G.; Paus, R. Exploring the human hair follicle microbiome. Br. J. Dermatol. 2021, 184, 802–815. [Google Scholar] [CrossRef] [PubMed]
- De Berker, D. Nail anatomy. Clin. Dermatol. 2013, 31, 509–515. [Google Scholar] [CrossRef]
- Okoro, O.E.; Camera, E.; Flori, E.; Ottaviani, M. Insulin and the sebaceous gland function. Front. Physiol. 2023, 14, 1252972. [Google Scholar] [CrossRef] [PubMed]
- Makrantonaki, E.; Ganceviciene, R.; Zouboulis, C. An update on the role of the sebaceous gland in the pathogenesis of acne. Dermato-Endocrinol. 2011, 3, 41–49. [Google Scholar] [CrossRef]
- Baker, L.B. Physiology of sweat gland function: The roles of sweating and sweat composition in human health. Temperature 2019, 17, 211–259. [Google Scholar] [CrossRef] [PubMed]
- Kurata, R.; Futaki, S.; Nakano, I.; Fujita, F.; Tanemura, A.; Murota, H.; Katayama, I.; Okada, F.; Sekiguchi, K. Three-dimensional cell shapes and arrangements in human sweat glands as revealed by whole-mount immunostaining. PLoS ONE 2017, 21, e0178709. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Zhang, L.; Chen, J.; Wang, C.; Zhao, J.; Liu, X.; Yan, Y.; Tang, Y.; Chen, Z.; Li, H. Differential antigen expression between human apocrine sweat glands and eccrine sweat glands. Eur. J. Histochem. 2023, 67, 3559. [Google Scholar] [CrossRef]
- Saga, K. Histochemical and immunohistochemical markers for human eccrine and apocrine sweat glands: An aid for histopathologic differentiation of sweat gland tumors. J. Investig. Dermatol. Symp. Proc. 2001, 6, 49–53. [Google Scholar] [CrossRef]
- Ferreira, T.; Gama, A.; Seixas, F.; Faustino-Rocha, A.I.; Lopes, C.; Gaspar, V.M.; Mano, J.F.; Medeiros, R.; Oliveira, P.A. Mammary glands of women, female dogs and female rats: Similarities and differences to be considered in breast cancer research. Vet. Sci. 2023, 10, 379. [Google Scholar] [CrossRef] [PubMed]
- Musumeci, G.; Castrogiovanni, P.; Szychlinska, M.A.; Aiello, F.C.; Vecchio, G.M.; Salvatorelli, L.; Magro, G.; Imbesi, R. Mammary gland: From embryogenesis to adult life. Acta Histochem. 2015, 117, 379–385. [Google Scholar] [CrossRef]
- Proksch, E. pH in nature, humans and skin. J. Dermatol. 2018, 45, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Blaak, J.; Staib, P. The relation of pH and skin cleansing. Curr. Probl. Dermatol. 2018, 54, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Visscher, M.O.; Adam, R.; Brink, S.; Odio, M. Newborn infant skin: Physiology, development, and care. Clin. Dermatol. 2015, 33, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Veltri, A.; Lang, C.; Lien, W.H. Concise review: Wnt signaling pathways in skin development and epidermal stem cells. Stem Cells 2018, 36, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Pierzak, M.; Kręcisz, B.; Suliga, E. Bioactive compounds for skin health: A review. Nutrients 2021, 113, 203. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J.; Veysey, E.C.; Finlay, A.Y. Aging and the Skin. In Brocklehurst’s Textbook of Geriatric Medicine and Gerontology, 8th ed.; Fillit, H.M., Rockwood, K., Young, J., Eds.; Elsevier: Philadelphia, PA, USA, 2017; Chapter 25. [Google Scholar]
- Chhabra, S.; Chhabra, N.; Kaur, A.; Gupta, N. Wound healing concepts in clinical practice of OMFS. J. Maxillofac. Oral Surg. 2017, 16, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Andersen, C.; Black, J.; de Leon, J.; Fife, C.; Lantis Ii, J.C.; Niezgoda, J.; Snyder, R.; Sumpio, B.; Tettelbach, W.; et al. Management of chronic wounds: Diagnosis, preparation, treatment, and follow-up. Wounds 2017, 29, S19–S36. [Google Scholar] [PubMed]
- Babalska, Z.L.; Korbecka-Paczkowska, M.; Karpiński, T.M. Wound antiseptics and european guidelines for antiseptic application in wound treatment. Pharmaceuticals 2021, 14, 1253. [Google Scholar] [CrossRef] [PubMed]
- Nuutila, K.; Katayama, S.; Vuola, J.; Kankuri, E. Human wound-healing research: Issues and perspectives for studies using wide-scale analytic platforms. Adv. Wound Care New Rochelle 2014, 3, 264–271. [Google Scholar] [CrossRef]
- Monika, P.; Chandraprabha, M.N.; Rangarajan, A.; Waiker, P.V.; Chidambara Murthy, K.N. Challenges in healing wound: Role of complementary and alternative medicine. Front. Nutr. 2022, 8, 791899. [Google Scholar] [CrossRef]
- Mahmoud, M.; Gould, L.J. Opportunities and challenges of the management of chronic wounds: A multidisciplinary viewpoint. Chronic Wound Care Manag. Res. 2020, 7, 27–36. [Google Scholar] [CrossRef]
- Han, G.; Ceilley, R. Chronic wound healing: A review of current management and treatments. Adv. Ther. 2017, 34, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Bacci, S. Cellular mechanisms and therapies in wound healing: Looking toward the future. Biomedicines 2021, 9, 1611. [Google Scholar] [CrossRef] [PubMed]
- Takeo, M.; Lee, W.; Ito, M. Wound healing and skin regeneration. Cold Spring Harb. Perspect. Med. 2015, 5, a023267. [Google Scholar] [CrossRef] [PubMed]
- Arenas Gómez, C.M.; Sabin, K.Z.; Echeverri, K. Wound healing across the animal kingdom: Crosstalk between the immune system and the extracellular matrix. Dev. Dyn. 2020, 249, 834–846. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.J. Parallels between vertebrate cardiac and cutaneous wound healing and regeneration. npj Regen. Med. 2018, 3, 21. [Google Scholar] [CrossRef] [PubMed]
- Belacortu, Y.; Paricio, N. Drosophila as a model of wound healing and tissue regeneration in vertebrates. Dev. Dyn. 2011, 240, 2379–2404. [Google Scholar] [CrossRef]
- Seifert, A.W.; Maden, M. New insights into vertebrate skin regeneration. Int. Rev. Cell Mol. Biol. 2014, 310, 129–169. [Google Scholar] [CrossRef] [PubMed]
- Ozgok Kangal, M.K.; Regan, J.P. Wound Healing 2023. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK535406/ (accessed on 29 May 2024).
- Healing by intention. Adv. Ski. Wound Care 2017, 30, 246–247. [CrossRef]
- Childs, D.R.; Murthy, A.S. Overview of wound healing and management. Surg. Clin. N. Am. 2017, 97, 189–207. [Google Scholar] [CrossRef]
- Braiman-Wiksman, L.; Solomonik, I.; Spira, R.; Tennenbaum, T. Novel insights into wound healing sequence of events. Toxicol. Pathol. 2007, 35, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Grey, J.E.; Enoch, S.; Harding, K.G. Wound assessment. BMJ 2006, 332, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Madkour, F.F.; Hassan, M.M.; Abdo, W.; Khalil, W.F. Wound healing activity of brown algae plus polyherbal extract in normal and alloxan-induced diabetic rats. J. Adv. Vet. Res. 2013, 3, 102–108. [Google Scholar]
- Annoodee, S.; Nasuruddin, D.N. Acute Inflammatory Response. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2024. Available online: https://www.ncbi.nlm.nih.gov/books/NBK556083/ (accessed on 29 May 2024).
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Palmblad, J. The acute inflammatory reaction: New concepts for old cells. J. Intern. Med. 2010, 268, 1. [Google Scholar] [CrossRef] [PubMed]
- Fioranelli, M.; Roccia, M.G.; Flavin, D.; Cota, L. Regulation of inflammatory reaction in health and disease. Int. J. Mol. Sci. 2021, 332, 5277. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Guarino, M.; Hernández-Bule, M.L.; Bacci, S. Cellular and molecular processes in wound healing. Biomedicines 2023, 11, 2526. [Google Scholar] [CrossRef]
- Canedo-Dorantes, L.; Canedo-Ayala, M. Skin acute wound healing: A comprehensive review. Int. J. Inflamm. 2019, 2019, 3706315. [Google Scholar] [CrossRef]
- Tyavambiza, C.; Meyer, M.; Meyer, S. Cellular and molecular events of wound healing and the potential of silver based nanoformulations as wound healing agents. Bioengineering 2022, 9, 712. [Google Scholar] [CrossRef]
- Raziyeva, K.; Kim, Y.; Zharkinbekov, Z.; Kassymbek, K.; Jimi, S.; Saparov, A. Immunology of acute and chronic wound healing. Biomolecules 2021, 11, 700. [Google Scholar] [CrossRef]
- Wilkinson, H.N.; Hardman, M.J. Wound healing: Cellular mechanisms and pathological outcomes. Open Biol. 2020, 10, 200223. [Google Scholar] [CrossRef] [PubMed]
- Banu, S.A.; Sharun, M.K.; Mamachan, L.; Abualigah, R.; Kumar, A.M.; Pawde, K.; Dhama, S.K.; Maiti, A. Wound healing and skin regeneration: Present status and future directions. J. Exp. Biol. Agric. Sci. 2023, 11, 871–883. [Google Scholar] [CrossRef]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound Healing: A cellular perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef] [PubMed]
- Wilgus, T.A. Fetal wound healing. In Wound Healing, Tissue Repair, and Regeneration in Diabetes; Bagchi, D., Das, A., Roy, S., Eds.; Academic Press: Cambridge, MA, USA, 2020; Chapter 27; pp. 579–591. [Google Scholar]
- Yagi, L.H.; Watanuki, L.M.; Isaac, C.; Gemperli, R.; Nakamura, M.Y.; Ladeira, P.R.S. Human fetal wound healing a review of molecular and cellular aspects. Eur. J. Plast. Surg. 2016, 39, 239–246. [Google Scholar] [CrossRef]
- Harding, K.G.; Morris, H.L.; Patel, G.K. Science, medicine and the future, Healing chronic wounds. Br. Med. J. 2002, 324, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Toporcer, T.; Lakyová, L.; Radonak, J. Venous ulcer-present view on aetiology, diagnostics and therapy. Cas. Lek. Cesk. 2008, 147, 199–205. [Google Scholar] [PubMed]
- Günter, C.I.; Machens, H.G. New strategies in clinical care of skin wound healing. Eur. Surg. Res. 2012, 49, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Sen, C.K. Human wounds and its burden: An updated compendium of estimates. Adv. Wound Care 2019, 8, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Kyaw, B.M.; Järbrink, K.; Martinengo, L.; Car, J.; Harding, K.; Schmidtchen, A. Need for improved definition of chronic wounds in clinical studies. Acta Derm. Venereol. 2018, 12, 157–158. [Google Scholar] [CrossRef]
- Fernández-Guarino, M.; Bacci, S.; Pérez González, L.A.; Bermejo-Martínez, M.; Cecilia-Matilla, A.; Hernández-Bule, M.L. The role of physical therapies in wound healing and assisted scarring. Int. J. Mol. Sci. 2023, 19, 7487. [Google Scholar] [CrossRef]
- Martin, P.; Nunan, R. Cellular and molecular mechanisms of repair in acute and chronic wound healing. Br. J. Dermatol. 2015, 173, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Grandi, V.; Corsi, A.; Pimpinelli, N.; Bacci, S. Cellular mechanisms in acute and chronic wounds after PDT Therapy: An Update. Biomedicines 2022, 10, 1624. [Google Scholar] [CrossRef] [PubMed]
- Falanga, V.; Isseroff, R.R.; Soulika, A.M.; Romanelli, M.; Margolis, D.; Kapp, S.; Granick, M.; Harding, K. Chronic wounds. Nat. Rev. Dis. Prim. 2022, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.C.; Costa, T.F.; Andrade, Z.A.; Medrado, A.R. Wound healing—A literature review. An. Bras. Dermatol. 2016, 91, 614–620. [Google Scholar] [CrossRef]
- Tottoli, E.M.; Dorati, R.; Genta, I.; Chiesa, E.; Pisani, S.; Conti, B. Skin wound healing process and new emerging technologies for skin wound care and regeneration. Pharmaceutics 2020, 12, 735. [Google Scholar] [CrossRef] [PubMed]
- Steinman, L. Elaborate interactions between the immune and nervous systems. Nat. Immunol. 2004, 5, 575–581. [Google Scholar] [CrossRef]
- Grandi, V.; Paroli, G.; Puliti, E.; Bacci, S.; Pimpinelli, N. Single ALA-PDT irradiation induces increase in MC degranulation and neuropeptide acute response in chronic venous ulcers: A pilot study. Photodiagn. Photodyn. Ther. 2021, 34, 102222. [Google Scholar] [CrossRef] [PubMed]
- Nardini, P.; Notari, L.; Magazzini, M.; Mariani, B.; Rossi, F.; Rossi, S.; Van Aardt, E.; Marszalek, K.; Grandi, V.; Corsi, A.; et al. Neuroimmunomodulatory Effect of NO on chronic wound healing after photodynamic therapy. Photodiagnosis Photodyn. Ther. 2024, 47, 104078. [Google Scholar] [CrossRef] [PubMed]
- Corsi, A.; Lecci, P.P.; Bacci, S.; Cappugi, P.; Pimpinelli, N. Early activation of fibroblasts during PDT treatment in leg ulcers. G. Ital. Dermatol. Venereol. 2016, 151, 223–229. [Google Scholar]
- Grandi, V.; Bacci, S.; Corsi, A.; Sessa, M.; Puliti, E.; Murciano, N.; Scavone, F.; Cappugi, P.; Pimpinelli, N. ALA-PDT exerts beneficial effects on chronic venous ulcers by inducing changes in inflammatory microenvironment, especially through increased TGF-beta release: A pilot clinical and translational study. Photodiagnosis Photodyn. Ther. 2018, 21, 252–256. [Google Scholar] [CrossRef]
- Kadam, S.; Nadkarni, S.; Lele, J.; Sakhalkar, S.; Mokashi, P.; Kaushik, K.S. Bioengineered platforms for chronic wound infection studies: How can we make them more human-relevant? Front. Bioeng. Biotechnol. 2019, 7, 418. [Google Scholar] [CrossRef]
- Limandjaja, G.C.; Niessen, F.B.; Scheper, R.J. Gibbs S: The keloid disorder: Heterogeneity, histopathology, mechanisms and models. Front. Cell Dev. Biol. 2020, 8, 360. [Google Scholar] [CrossRef]
- Huang, C.; Ogawa, R. Keloidal pathophysiology: Current notions. Scars Burn. Health 2021, 7, 2059513120980320. [Google Scholar] [CrossRef]
- Magni, G.; Banchelli, M.; Cherchi, F.; Coppi, E.; Fraccalvieri, M.; Rossi, M.; Tatini, F.; Pugliese, A.M.; Rossi Degl’Innocenti, D.; Alfieri, D.; et al. Experimental study on blue light interaction with human keloid-derived fibroblasts. Biomedicines 2020, 8, 573. [Google Scholar] [CrossRef]
- Boyce, D.E.; Ciampolini, J.; Ruge, F.; Murison, M.S.; Harding, K.G. Inflammatory-cell subpopulations in keloid scars. Br. J. Plast. Surg. 2001, 54, 511–516. [Google Scholar] [CrossRef]
- Bagabir, R.; Byers, R.J.; Chaudhry, I.H.; Muller, W.; Paus, R.; Bayat, A. Site-specific immunophenotyping of keloid disease demonstrates immune upregulation and the presence of lymphoid aggregates. Br. J. Dermatol. 2012, 167, 1053–1066. [Google Scholar] [CrossRef]
- Van der Veer, W.M.; Bloemen, M.C.; Ulrich, M.M.; Molema, G.; van Zuijlen, P.P.; Middelkoop, E.; Niessen, F.B. Potential cellular and molecular causes of hypertrophic scar formation. Burns 2009, 35, 15–29. [Google Scholar] [CrossRef]
- Mony, M.P.; Harmon, K.A.; Hess, R.; Dorafshar, A.H.; Shafikhani, S.H. An updated review of hypertrophic scarring. Cells 2023, 12, 678. [Google Scholar] [CrossRef]
- Dhivya, S.; Padma, V.V.; Santhini, E. Wound dressings—A review. BioMedicine 2015, 5, 22. [Google Scholar] [CrossRef]
- Rezvani Ghomi, E.; Khalili, S.; Nouri Khorasani, S.; Esmaeely Neisiany, R.; Ramakrishna, S. Wound dressings: Current advances and future directions. J. Appl. Polym. Sci. 2019, 136, 47738. [Google Scholar] [CrossRef]
- Obagi, Z.; Damiani, G.; Grada, A.; Falanga, V. Principles of Wound Dressings: A Review. Surg. Technol. Int. 2019, 35, 50–57. [Google Scholar]
- Dong, R.; Guo, B. Smart wound dressings for wound healing. Nanotoday 2021, 41, 101290. [Google Scholar] [CrossRef]
- Rossi, F.; Tatini, F.; Pini, R.; Bacci, S.; De Siena, G.; Cicchi, R.; Pavone, S.; Alfieri, D. Improved wound healing in blue LED treated superficial abrasions. In Proceedings of the European Conferences on Biomedical Optics, Munich, Germany, 12–16 May 2013; p. 88030S. [Google Scholar] [CrossRef]
- Cicchi, R.; Rossi, F.; Alfieri, D.; Bacci, S.; Tatini, F.; De Siena, G.; Paroli, G.; Pini, R.; Pavone, F.S. Observation of an improved healing process in superficial skin wounds after irradiation with a blue-LED haemostatic device. J. Biophotonics 2016, 9, 645–655. [Google Scholar] [CrossRef]
- Magni, G.; Tatini, F.; Siena, G.; Pavone, F.S.; Alfieri, D.; Cicchi, R.; Rossi, M.; Murciano, N.; Paroli, G.; Vannucci, C.; et al. Blue-LED-light photobiomodulation of inflammatory responses and new tissue formation in mouse-skin wounds. Life 2022, 12, 1564. [Google Scholar] [CrossRef]
- Magni, G.; Tatini, F.; Bacci, S.; Paroli, G.; De Siena, G.; Cicchi, R.; Pavone, F.S.; Pini, R.; Rossi, F. Blue LED light modulates inflammatory infiltrate and improves the healing of superficial wounds. Photodermatol. Photoimmunol. Photomed. 2020, 36, 166–168. [Google Scholar] [CrossRef]
- Hernández-Bule, M.L.; Naharro-Rodríguez, J.; Bacci, S.; Fernández-Guarino, M. Unlocking the Power of Light on the Skin: A Comprehensive Review on Photobiomodulation. Int. J. Mol. Sci. 2024, 25, 4483. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Processes | Associated Visible Changes and Events | Cells | Cell Adhesion Molecules | ECM Components | Key Mediators/Regulators |
|---|---|---|---|---|---|---|
| 0–15 min | Hemostasis | Whitening, formation of fibrin clot | Endothelial cells, platelets | Fibrin, fibronectin | Epinephrine, prostaglandins, thromboxanes, thrombin | |
| 15 min–6 days | Inflammation | Redness, swelling, heat of the inflamed part, pain, functional alteration, vasodilation, diapedesis, inflammatory response, phagocytosis | Endothelial cells, MCs, macrophages, T lymphocytes | Temporary matrix formed by fibrin, complement proteins, PDGF, IL8, IL1 alpha, IL1 beta, IL6 and TNF-alpha | Sympathetic nervous system, histamine, kinins, leukotrienes, thrombin | |
| 24–48 h–7 days | Proliferation | Angiogenesis detachment of the eschar, epithelialization, granulation tissue | Endothelial cells, fibroblasts keratinocytes | Alphabeta3, Beta1-integrins, integrins | Collagen, fibronectin, GAGs, proteoglycans, Tenascin, Vitronectin (temporary ECM) | Angiopoietin, MMPs/TIMP, FGF2, FGF-7, FGF-10, GM-CSF, NO, TGF-beta, NGF, HGF, HB-EGF, IL-6, Leptin, PDGF, VEGF |
| From 2 days to several weeks | Maturation | Approaching the edges of the wound (wound contraction), scar deposition of the collagen network | Fibroblasts, myofibroblasts | Integrins | Collagen | EGF, IGF, FGF-2, NGF, PDGF, TGF-beta |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Férnandez-Guarino, M.; Naharro-Rodriguez, J.; Bacci, S. Disturbances in the Skin Homeostasis: Wound Healing, an Undefined Process. Cosmetics 2024, 11, 90. https://doi.org/10.3390/cosmetics11030090
Férnandez-Guarino M, Naharro-Rodriguez J, Bacci S. Disturbances in the Skin Homeostasis: Wound Healing, an Undefined Process. Cosmetics. 2024; 11(3):90. https://doi.org/10.3390/cosmetics11030090
Chicago/Turabian StyleFérnandez-Guarino, Montserrat, Jorge Naharro-Rodriguez, and Stefano Bacci. 2024. "Disturbances in the Skin Homeostasis: Wound Healing, an Undefined Process" Cosmetics 11, no. 3: 90. https://doi.org/10.3390/cosmetics11030090
APA StyleFérnandez-Guarino, M., Naharro-Rodriguez, J., & Bacci, S. (2024). Disturbances in the Skin Homeostasis: Wound Healing, an Undefined Process. Cosmetics, 11(3), 90. https://doi.org/10.3390/cosmetics11030090