
Sargassum fusiforme Extract Induces Melanogenesis through the cAMP/PKA/CREB Signaling Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sargassum Fusiforme Extract
2.2. Cell Culture
2.3. Chemical Reagents and Antibodies
2.4. Cell Viability Measurement and Melanin Contents Assay
2.5. Oxidative Stress Induction with Hydrogen Peroxide
2.6. Western Blot Analysis
2.7. Tyrosinase Zymography
2.8. Measurement of Cellular Tyrosinase Activity
2.9. Reconstituted Three-Dimensional Human Skin Model
2.10. Ex Vivo Human Hair Follicles
2.11. Statistical Analysis
3. Results
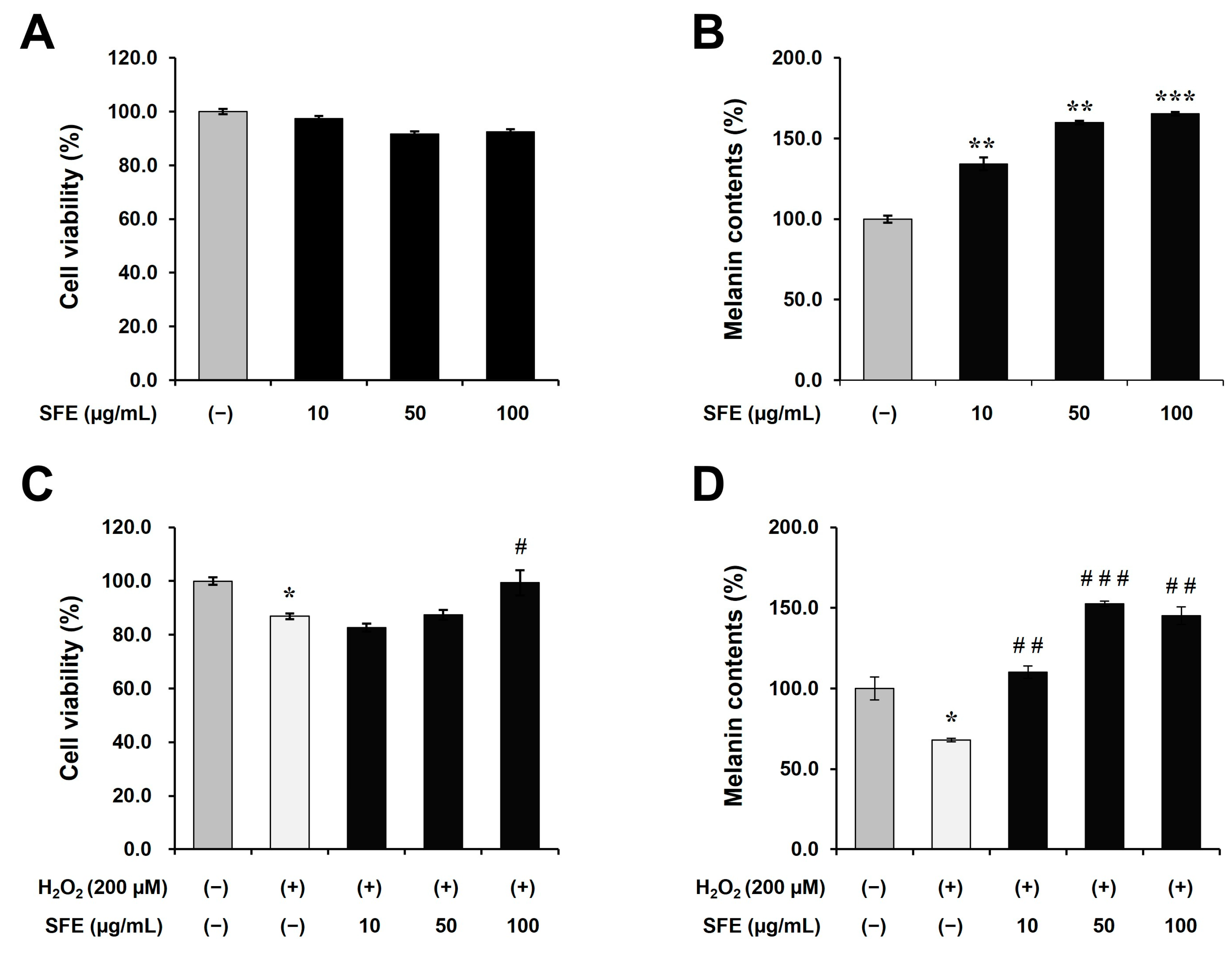
3.1. SFE Induces Melanogenesis in Human Epidermal Melanocytes
3.2. SFE Attenuates Both Cytotoxicity and Depigmentation in H2O2-Treated Human Epidermal Melanocytes
3.3. SFE Increases Expression Levels of Melanogenesis-Related Proteins in Human Epidermal Melanocytes
3.4. Effects of SFE on Cellular Tyrosinase Activity
3.5. SFE Promotes Melanogenesis by Regulating cAMP/PKA/CREB Pathway
3.6. Hyperpigmentation Effect of SFE on a Reconstituted Human Skin Model
3.7. Hyperpigmentation Effect of SFE on Ex Vivo Human Hair Follicles
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Proksch, E.; Brandner, J.M.; Jensen, J.M. The skin: An indispensable barrier. Exp. Dermatol. 2008, 17, 106–112. [Google Scholar] [CrossRef]
- Elias, P.M.; Feingold, K.R. Skin barrier. In Medical Biochemistry; Baron, J., Ed.; Elsevier: Philadelphia, PA, USA, 2018; Chapter 60. [Google Scholar]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Wakamatsu, K.; Ozeki, H. Chemical analysis of melanins and its application to the study of the regulation of melanogenesis. Pigment. Cell Res. 2000, 13, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Hearing, V.J. Determination of melanin synthetic pathways. J. Investig. Dermatol. 2011, 131, E8–E11. [Google Scholar] [CrossRef] [PubMed]
- Taieb, A.; Picardo, M.; VETF Members. The definition and assessment of vitiligo: A consensus report of the Vitiligo European Task Force. Pigment. Cell Res. 2007, 20, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Gloster, H.M., Jr.; Neal, K. Skin cancer in skin of color. J. Am. Acad. Dermatol. 2006, 55, 741–764. [Google Scholar] [CrossRef]
- Nishimura, E.K.; Granter, S.R. Mechanisms of hair graying: Incomplete melanocyte stem cell maintenance in the niche. Science 2005, 307, 720–724. [Google Scholar] [CrossRef]
- Arck, P.C.; Overall, R.; Spatz, K.; Liezman, C.; Handjiski, B.; Klapp, B.F.; Birch-Machin, M.A. Towards a “free radical theory of graying”: Melanocyte apoptosis in the aging human hair follicle is an indicator of oxidative stress induced tissue damage. FASEB J. 2006, 20, 1567–1569. [Google Scholar] [CrossRef]
- Stacey, A.N.D.; Graeme, J.F.; Bruce, C.B.; Marjan, E.A. Signaling pathways in melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar]
- Carlos, I.R.; Vijayasaradhi, S. Cyclic AMP (cAMP) signaling in melanocytes and melanoma. Arch. Biochem. Biophys. 2014, 563, 22–27. [Google Scholar]
- Buscà, R.; Ballotti, R. Cyclic AMP a key messenger in the regulation of skin pigmentation. Pigment. Cell Res. 2000, 13, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Bhakti, T.; Avinash, M. Nutraceutical Potential of Seaweed Polysaccharides: Structure, Bioactivity, Safety, and Toxicity. Compr. Rev. Food Sci. Food Saf. 2019, 18, 817–831. [Google Scholar]
- Jian, L.; Sibusiso, L.; Yue, Y.; Yang, C.; Ya, Z.; Mingjiang, W.; Jong-il, C.; Haibin, T. Therapeutic and nutraceutical potentials of a brown seaweed Sargassum fusiforme. Food Sci. Nutr. 2020, 8, 5195–5205. [Google Scholar]
- Zhang, R.; Zhang, X.; Tang, Y.; Mao, J. Composition, isolation, purification and biological activities of Sargassum fusiforme polysaccharides: A review. Carbohydr. Polym. 2020, 228, 115381. [Google Scholar] [CrossRef] [PubMed]
- Subhash, R.Y.; Uday, N.H.; Bhupal, B.C. Therapeutic potential and health benefits of Sargassum species. Pharmacogn. Rev. 2014, 8, 1–7. [Google Scholar]
- Wang, L.; Cui, Y.R.; Lee, H.-G.; Fu, X.; Wang, K.; Xu, J.; Gao, X.; Jeon, Y.-J. Fucoidan isolated from fermented Sargassum fusiforme suppresses oxidative stress through stimulating the expression of superoxidase dismutase and catalase by regulating Nrf2 signaling pathway. Int. J. Biol. Macromol. 2022, 209 Pt A, 935–941. [Google Scholar] [CrossRef]
- Maria, E.R.D.; Marc, A.C.; Miguel, D.N.; Alberto, S.C. Structural studies on fucoidans from the brown seaweed Sargassum stenophyllum. Carbohydr. Res. 2001, 333, 281–293. [Google Scholar]
- Marcelo, D.C.; Rita, S.R.; Amina, C.; Sónia, S.; Susana, S.B.; Artur, M.S.S.; Susana, M.C. Applications of antioxidant secondary metabolites of Sargassium. Mar. Drugs 2023, 21, 172. [Google Scholar]
- Antia, G.P.; Maria, F.C.; Paula, G.O.; Catarina, L.L.; Maria, C.; Miguel, A.P.; Jesus, S.G. The use of invasive algae species as a source of secondary metabolites and biological activities: Spain as case study. Mar. Drugs 2021, 19, 178. [Google Scholar]
- Wang, L.; Kim, H.-S.; Je, J.-G.; Fu, X.; Huang, C.; Ahn, G.; Oh, J.-Y.; Sanjeewa, A.; Xu, J.; Gao, X.; et al. In vitro and in vivo photoprotective effects of (-)-Loliode isolated from the Brown seaweed Sargassum horneri. Molecules 2021, 26, 6898. [Google Scholar] [CrossRef]
- Hui-Min, D.W.; Ching-Chun, C.; Pauline, H.; Jo-Shu, C. Exploring the potential of using algae in cosmetics. Bioresour. Technol. 2014, 184, 355–362. [Google Scholar]
- Thouria, B.; Myra, O.V.; François, C.; Abdellatif, H.; Hiroko, I.; Chemseddoha, G. Melanogenesis Promoting Effect, Antioxidant Activity, and UPLC-ESI-HRMS Characterization of Phenolic Compounds of Argan Leaves Extract. Molecules 2021, 26, 371. [Google Scholar]
- Nuramina, M.; Xue, Y.L.; Maidina, K.; Haji, A.A. Potential anti-vitiligo properties of cynarine extracted from Vernonia anthelmintica (L.) Willd. Int. J. Mol. Med. 2018, 42, 2665–2675. [Google Scholar]
- Subia, J.; Rafeeq, A.K.; Shadab, A.; Sakina, F. Evaluation of anti-inflammatory and anti-oxidant potential of seed extracts of Vernonia anthelmintica. Pak. J. Pharm. Sci. 2017, 30, 755–760. [Google Scholar]
- Wood, J.M.; Decker, H.; Hartmann, H.; Chavan, B.; Rokos, H.; Spencer, J.D.; Hasse, S.; Thornton, M.J.; Shalbaf, M.; Paus, R.; et al. Senile hair graying: H2O2-mediated oxidative stress affects human hair color by blunting methionine sulfoxide repair. FASEB J. 2009, 23, 2065–2075. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Lu, Y.; Wang, H.; Feng, Y.; Jiang, S.; Gao, X.-H.; Qi, R.; Wu, Y.; Chen, H.-D. Paeoniflorin Resists H2O2-Induced Oxidative Stress in Melanocytes by JNK/Nrf2/HO-1 Pathway. Front. Pharmacol. 2020, 11, 536. [Google Scholar] [CrossRef]
- Yang, L.; Yang, F.; Teng, L.; Katayama, I. 6-Shogaol Protects Human Melanocytes against Oxidative Stress through Activation of the Nrf2-Antioxidant Response Element Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 3537. [Google Scholar] [CrossRef]
- Denat, L.; Kadekaro, A.L.; Marrot, L.; Leachman, S.; Abdel-Malek, Z.A. Melanocytes as Instigators and Victims of Oxidative Stress. J. Investig. Dermatol. 2014, 134, 1512–1518. [Google Scholar] [CrossRef]
- Pilas, P.; Sarna, T.; Kalyanaraman, B.; Swartz, H.M. The effect of melanin on iron associated decomposition of hydrogen peroxide. Free Radic. Biol. Med. 1988, 4, 285–293. [Google Scholar] [CrossRef]
- Luisa, N.; Alessandra, N.; Giuseppe, P. 5,6-Dihydroxyindoles in the Fenton Reaction: A Model Study of the Role of Melanin Precursors in Oxidative Stress and Hyperpigmentary Processes. Chem. Res. Toxicol. 1999, 12, 985–992. [Google Scholar]
- Joong-Woon, C.; Bark-Lynn, L.; Woo-Young, S. A Case of Premature Hair Graying Treated with Ferrous Sulfate. Ann. Dermatol. 2016, 28, 775–776. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Shin, S.; Jang, Y.; Cho, E.; Park, D.; Jung, E. Sargassum fusiforme Extract Induces Melanogenesis through the cAMP/PKA/CREB Signaling Pathway. Cosmetics 2023, 10, 116. https://doi.org/10.3390/cosmetics10040116
Kim H, Shin S, Jang Y, Cho E, Park D, Jung E. Sargassum fusiforme Extract Induces Melanogenesis through the cAMP/PKA/CREB Signaling Pathway. Cosmetics. 2023; 10(4):116. https://doi.org/10.3390/cosmetics10040116
Chicago/Turabian StyleKim, Hayeon, Seoungwoo Shin, Youngsu Jang, Eunae Cho, Deokhoon Park, and Eunsun Jung. 2023. "Sargassum fusiforme Extract Induces Melanogenesis through the cAMP/PKA/CREB Signaling Pathway" Cosmetics 10, no. 4: 116. https://doi.org/10.3390/cosmetics10040116
APA StyleKim, H., Shin, S., Jang, Y., Cho, E., Park, D., & Jung, E. (2023). Sargassum fusiforme Extract Induces Melanogenesis through the cAMP/PKA/CREB Signaling Pathway. Cosmetics, 10(4), 116. https://doi.org/10.3390/cosmetics10040116