Abstract
Over the last fifty years there has been a continual reduction in horticultural and agricultural biodiversity of nutritionally important plants, including those of the Solanaceae family. To add to this, the broad range of traditional crops, previously grown on a sustainable scale in some parts of the world, has been replaced by a narrow range of major crops grown as large-scale monocultures. In order to counteract this trend, and to help maintain a broad wealth of genetic resources, conservation is essential. This, in turn, helps to safeguard food security. A taxonomic inventory, covering the diversity of species in a plant group, is an important first step in conservation. The Solanaceae is one of the major plant families providing food species. A survey of the biodiversity, ethnobotany and taxonomy of subfamily Solanoideae was undertaken and is presented here as an inventory of food species. Fifteen genera provide species that are utilised for food across the world. Of these, only four genera contain economically significant cultivated food cropspecies. The majority of these are in the genus Solanum, whilst Capsicum, Physalis and Lycium contribute the remainder of cultivated crop species. These genera and others also comprise species that are semi-cultivated, tolerated as useful weeds, or gathered from the wild.
1. Introduction
1.1. Conservation of Food Species
Around 30,000 of the total of approximately 250,000 flowering plant species in existence are edible [1], with the vast majority of the edible species being “minor crops.” These may be culturally important cultigens, semi-wild species, wild species, or weeds; in turn they may be locally cultivated on a small-scale, semi-cultivated in and around settlements, gathered from the wild, or tolerated in cultivated areas, respectively. According to Bemejo and Léon [2] as few as 150 plant species are actually cultivated, 12 of which provide around 75% of the world’s food. The broad range of traditional crops, which were previously grown on a sustainable scale in some parts of the world, has been replaced by a narrow range of major crops, grown as large-scale monocultures.This has consequently narrowed the number of species upon which global food security depends [3]. Furthermore, over the last 50 years, dependence upon commercial hybrids and advanced cultivars, as well as the neglect of traditional landraces, has led to a vast reduction in horticultural and agricultural biodiversity within the most popular species. In addition, habitat loss and climatic phenomena, as well as cultural changes, have caused a reduction in the range of non-commercial crops that are routinely used by humans. Vegetable genetic resourcesare being lost globally at a rate of 1%–2% per year [4]; this is largely as a result of the changes in the way the human population utilises the edible plants of the world.
1.2. Expanding the Food Species Repertoire
In recent times, the intrinsic value of the consumption of a broad repertoire of edible plant species has been pinpointed, and international recognition of the potential of minor crops has been encouraged by several plans of action involving worldwide participation, e.g., the FAO’s “Global Plan of Action for the Conservation and Sustainable Utilization of Plant Genetic Resources” and “Rome Declaration on World Food Security” [5,6]. Further interest in neglected, underutilised, or “forgotten” species has been demonstrated in various local, regional, and general accounts, as well as in coverage of individual species [7,8,9,10,11,12,13,14,15,16,17]. There has thus been a growing awareness of the vital role of a diversity of wild, semi-domesticated and underdeveloped species in food and livelihood security and their potential for further development and wider use.However, it is generally anticipated that changes in land use will limit the area available for agriculture and increase the pressure on populations of crop wild relatives (CWR) and wild food plants [18]. In this light, the conservation of such species is vital to the maintenance of plant biodiversity, as well as to the preservation of the much-needed genetic diversity (plant genetic resources), which is essential for traditional plant breeding.Recent Convention on Biological Diversity guidelines [19] take this into account, whist paying due regard to the importance of the associated indigenous and local knowledge.
1.3. The Solanaceae Family
The Solanaceae (nightshades) is one of the most important plant families and has extensive economic importance [20]. Its greatest concentration of diversity is to be found in South America and it is believed that the family originated here [21]. Spread from the neotropical region has allowed the family to be represented on every vegetated continent. Apart from the broad range of other uses (e.g., traditional medicine, traditional culture, pharmacology, ornamental horticulture) Solanaceae species have enormous importance as food plants the world over [22]. For example, in 2010 alone, 28 million hectares of solanaceous food crops were cultivated globally, producing around 540 million tonnes [23]. However, this relates to just four staple crop species (potato, tomato, aubergine, and capsicums) and does not take into account the lesser-known cultivated species, of which there are many. Nor does it account for the many species that are semi-cultivated or gathered from the wild. The majority of these are considered to be underutilised species, and a recent assessment of the most common of these [24] earmarked 17 underutilised species of Solanum L. alone.
1.4. Solanaceae Classification
Solanaceae classification presents several challenges. It is typical of that of many large families of angiosperms, in having an accepted core of genera assigned to the family, and a number of allied genera that are alternatively treated as belonging to the family, belonging to other, related families, or segregated into their own families [25]. Certain genera may or may not be regarded as synonymous, according to the rationale of the researchers. An example of this is found in some of the genera of Physalis allies (tribe Physaleae) which, over the years, have fluctuated in and out of synonymy. Many accepted species also have synonyms (ranging from several to many) which can complicate nomenclature and identification. Furthermore, new species (mostly from the tropics) are being described on a regular basis; these must then be incorporated into existing classifications. Recent taxonomic studies of the Solanaceae have generated phylogenetic systems based on DNA analyses; four main clades at subfamily level, 14 main clades at tribe level, around 100 genera and 2500 species are currently recognised [26].
1.5. The Taxonomic Inventory
Surveys, taxonomic identification and analysis of the extent and distribution of plant genetic diversity are priorities for neglected and underutilised plant species [27]. For large, diverse plant families such as the Solanaceae the construction of a taxonomic inventory is the first step in the assessment of the biodiversity of its food species; this preliminary task may be particularly important in the protected areas of the world where few floristic studies have been undertaken to assess the naturally-occurring CWRs and traditional landraces found there [28]. One such study recently demonstrated that, on the basis of taxonomic diversity, the Solanaceae was placed in the top three families containing CWRs of Venezuelan crops [29].Once adequate information has been amassed, the inventory may then become the starting point for subsequent conservation efforts.
1.6. Previous Studies
Some general checklists of useful and edible plants in different parts of the world (e.g., [30,31,32,33,34,35,36]) have demonstrated the importance of plant families, such as the Solanaceae, as sources of food. In addition, there have been several studies which focus solely on solanaceous species in localised areas, (e.g., [37,38,39,40,41,42,43,44,45,46]), or brief accounts of solanaceous crop use (e.g., [20,47]). These have provided excellent starting points for a worldwide study such as the present work, which is the first detailed inventory of its kind to be constructed for the Solanaceae.
2. Materials and Methods
2.1. Literature and Data Survey
Commencing in 2011, a survey of literature and online sources of data was undertaken. These were primarily regional and local floras, and studies relating to biodiversity and ethnobotany of Solanaceae taxa with recorded use as food plants (e.g., [8,30,35,48,49,50,51,52,53]).
2.2. Examination of Herbarium Specimens
This was made possible via visits to various herbaria in Europe, Asia, and South America: BM (Natural History Museum, London, UK); K (Royal Botanic Gardens, Kew, UK); L/U (Leiden, Utrecht; Naturalis Biodiversity Centre, Leiden, The Netherlands); MA (Real Jardín Botánico, Madrid, Spain); QCA (Pontificia Universidad Católica del Ecuador, Quito, Ecuador); RO (Università degli Studi di Roma La Sapienza, Rome, Italy); SING (Singapore Botanic Gardens, Singapore). Data from herbarium specimen sheet labels and annotations were recorded.
2.3. Access to Online Digitalised Herbarium Specimens/Flowering Plant Databases
Examination of high-resolution digital herbarium specimen images and their labels and annotations, as well as distribution and ethnobotanical information, was mediated via online access to: JSTOR Global Plants digitalised plant specimen database (Ithaka, NY, USA), Radboud University Solanaceae Database (Experimental Garden and Genebank, Radboud University, Nijmegen, Netherlands), Solanaceae Source (Natural History Museum, London, UK), Tropicos (Missouri Botanic Garden, St Louis, MO, USA), as well as several virtual herbaria holding Solanaceae specimen images and data. Data from herbarium sheet labels and annotations, as well as those from online databases, were recorded.
2.4. Collation of Data and Compilation of Taxonomic Inventory
Information on the various food species was collated in order to generate an inventory based on the taxonomic status of the taxa, down to sectional level where appropriate. The original inventory [47] was considerably expanded in order to provide the account below, whilst also being limited to coverage of subfamily Solanoideae Kostel, as this group contains by far the majority of food species; coverage of other, lesser-known subfamilies may be the subject of future studies. Care was taken to incorporate contemporary taxonomic viewpoints (from recent taxonomic monographs, reviews and revisions). Subfamily, tribe and subtribe names that are used below are mostly based on the scheme of [25], although these have yet to be formally published; for the purposes of clarity, the present work follows their system. The subgenera of Solanum recognised here follow the scheme of Child and Lester [54]; subgenera Archaesolanum Marzell, Brevantherum (Seithe) D’Arcy and Leptostemonum (Dunal) Bitter approximate to the equivalent clades of Weese and Bohs [55], whereas subgenera Solanum L. and Potatoe (G. Don) D’Arcy are further broken down into distinctly separate clades by the latter authors. Sections comprising the subgenera of Solanum follow Child and Lester [54] and Lester et al. [56], except that Cyphomandra (Cav.) Sendtn. is incorporated into Solanum [57] section Pachyphyllum Dunal. Total numbers of species for each genus are based on D’Arcy, Olmstead and Bohs, and Olmstead et al. [25,26,58,59], unless indicated otherwise. The author names of taxa follow the recommendations of IPNI [60] and, where relevant, common synonyms are listed.
3. Taxonomic Inventory of Subfamily Solanoideae Kostel
3.1. Tribe Capsiceae D’Arcy
Capsicum L.
Is a genus of approximately 32 species [61]; however, with new species being named on a regular basis, this number is likely to exceed 40 in the near future. Comprising small to robust shrubs, some with a climber-like habit. Capsicum is largely known for its edible berries, although in Asia and elsewhere the leaves of some species are sometimes eaten [62]. All of the truly wild Capsicum species are found solely in Latin America. There are five domesticated Capsicum species, two of which also have wild or semi-domesticated varieties, which are found in the human diet. In addition, there are up to another 18 wild species (crop wild relatives of the domesticated species); it is likely that these all have food value. Another 18or so species appear to be non-pungent, and studies on this group [63,64,65] suggest that it represents a separate branch of Capsicum evolution. In the following account of Capsicum an informal classification scheme has been adopted; synonymy and priority of names largely follow [64,66].
Domesticated Species (2n = 24)
C. annuum L. var. annuum L. comprises several thousand cultivars, including pungent “chilli peppers” and non-pungent forms, including the “bell peppers”, which are cultivated all over the world. There are many specialised regional forms, differing in fruit form, colour, flavour and pungency.
There are several hundred cultivars of C. baccatum L. var. pendulum (Willd.) Eshbaugh (Figure 1) which are grown across Colombia, Ecuador, Peru, Chile, Bolivia, Argentina and Brazil.This variety has also been introduced into Costa Rica, California, Nevada, Hawaii and India. It is generally known in South America as ají; fruit morphology may vary from pod-like to irregular, Scotch bonnet-shaped.

Figure 1.
C. baccatum var. pendulum: long pod form, cultivated in UK.
Figure 1.
C. baccatum var. pendulum: long pod form, cultivated in UK.

The several hundred cultivars of C. chinense Jacq. produce diverse fruit forms, such as jolokia (Assam),the Scotch Bonnet (Caribbean), and the Habañero (Mexico; Figure 2). This species is mostly grown across lowland South America, East of the Andes, Central America, Southern Mexico, Florida, Texas, the Carribean, tropical Africa and North-East India. This species is well-known for its fruity flavour and high pungency. The jolokia cultivars are amongst the world’s hottest known chilli peppers, although evidence suggests that these types may be of hybrid origin [67], originating from C. chinense x C. frutescens L. crosses. Wild C. chinense forms may be found in eastern lowland South America, e.g., Yellow Bird’s Eye from Brazil, and elsewhere.

Figure 2.
C. chinense cv. Orange Habañero.
Figure 2.
C. chinense cv. Orange Habañero.

C. frutescens L. shows a narrow range of morphological diversity and has few well-known cultivars or forms. The best-known are the Tabasco types (originally from Mexico) and the “bird peppers”. C. frutescens is grown in Ecuador and Louisiana for Tabasco sauce production, as well as in other parts of the Americas, the Pacific region, Eurasia, India, the Far East, Southeast Asia, and Africa.It may also be found wild or naturalised in regions where it is cultivated; some have colloquial names, e.g. melegueta, a popular form from Brazil.
C. pubescens Ruiz and Pav. (syn. C. guatemalense Bitter) fruit pods show a narrow range of morphological diversity, with five main forms: spherical, blocky (bell pepper-shaped), conical (Figure 3), ovoid and cylindrical-pointed [68]. This species bears black seeds [22], unlike the other cultivated species, whose seeds are generally a light brownish-grey colour. It is grown throughout the Andes from western Venezuela to Chile, and to a lesser extent in Costa Rica, Honduras, Guatemala, and Southern Mexico [16]. A recent survey has also revealed a history of cultivation in Java, Bali, and the Karimunjawa Islands, Indonesia [69]. It is known in much of the Andes as rocoto (Spanish) and rócot-uchu (Kichwa), or in Bolivia as locoto. It is also known as the “tree chilli” [70], on account of its robust growth, whereby perennial plants may reach 4 m in height. There are no known wild forms of this domesticate.

Figure 3.
C. pubescens: conical form.
Figure 3.
C. pubescens: conical form.

Wild Varieties of the Domesticated Species
C. annuum var. glabriusculum (Dunal) Heiser and Pickersgill (syn. C. annuum var. aviculare (Dierb.) D’Arcy and Pickersgill), known in Mexico as chiltepin (wild/semi-cultivated) or chilipiquin (cultivated), is found in Louisiana, Texas, Arizona, New Mexico, Florida, Mexico, Central America, Caribbean, Guyana, Venezuela, Colombia, Ecuador, and Northern Peru (Figure 4). Fruits are small, spherical or bullet-shaped, and red when mature. This is believed to be the most ancient form of C. annuum from which present-day cultivated C. annuum var. annuum evolved.

Figure 4.
C. annuum var. glabriusculum: cultivated piquin form. Courtesy of Clifford Ambers (Sweet Briar College, Sweet Briar, VA, USA).
Figure 4.
C. annuum var. glabriusculum: cultivated piquin form. Courtesy of Clifford Ambers (Sweet Briar College, Sweet Briar, VA, USA).

C. baccatum var. baccatum sensu Eshbaugh is found in Peru, Argentina, Bolivia, Paraguay, and Brazil [71], in localised areas. Fruits are relatively small (smallest forms may only reach 0.3 cm. diameter) ovoid and red when mature.
Wild Species (Mostly 2n = 24, Pungent)
These produce mostly spherical fruits, some ovoid, ripening to red, orange or yellow. The better known of these capsicums are: C. cardenasii Heiser and P.G. Sm. (Bolivia; Figure 5), C. eshbaughii Barboza (syn. C. eximium Hunz. var. tomentosum Eshbaugh and P.G. Sm., Bolivia [61]), and C. eximium Hunz. (Argentina and Bolivia), all known as ulupica; C. chacoense Hunz. (ají del monte, quitucho) from Argentina, Bolivia, and Paraguay; C. galapagoense Hunz., from the Galapagos Islands, Ecuador; C. tovarii Eshbaugh, P.G. Sm. and Nickrent, from Peru. Some wild species are regarded as semi-domesticated, as they may be encouraged to grow in the vicinity of settlements and habitations, e.g., C. praetermissum Heiser and P.G. Sm. (syn. C. baccatum var. praetermissum (Heiser and P.G. Sm.) Hunz. From Southern Brazil [65].

Figure 5.
C. cardenasii cv. “CAP 1530”: typical spherical fruits. Courtesy of Juppa Kilpinen (http://www.fatalii.net).
Figure 5.
C. cardenasii cv. “CAP 1530”: typical spherical fruits. Courtesy of Juppa Kilpinen (http://www.fatalii.net).

Wild Species (2n = 26, Probably all Non-Pungent)
These species have a diploid number of 2n = 26 [63,64,65] and seem to represent a separate evolutionary line in the genus. Pungency in some of these species has not been documented, but it is likely that the majority, at least, will turn out to be non-pungent; they may have food value, however. All have spherical fruits, ripening to red, orange or yellow. Species include C. rhomboideum (Humb., Bonpl. and Kunth) Kuntze (syn. C. ciliatum (Kunth) Kuntze), distributed through Southern Mexico to Northern Peru (Figure 6); C. lanceolatum (Greenm.) C.V. Morton and Standl., found in Mexico and Guatemala; C. schottianum Sendtn., found in Argentina, Brazil, and Paraguay.

Figure 6.
C. rhomboideum: yellow flowers and non-pungent fruits. Courtesy of Juppa Kilpinen (http://www.fatalii.net).
Figure 6.
C. rhomboideum: yellow flowers and non-pungent fruits. Courtesy of Juppa Kilpinen (http://www.fatalii.net).

New species of all branches of the genus are being named on a regular basis, e.g., C. caatingae Barboza and Agra, and C. longidentatum Agra and Barboza [72], both from Eastern Brazil. The ethnobotany of many new taxa has yet to be documented, but it is likely that those with pungent fruits will have significant food value as part of the diet of localised ethnic groups.
Lycianthes (Dunal) Hassl.
Is a genus of around 150–200 species of mostly herbs, shrubs, or vine-like plants distributed in tropical America and Eastern Asia.Fruits are typically brightly-coloured berries. Examples with edible fruits are: L. asarifolia (Kunth and Bouché) Bitter, common from Bolivia to Venezuela [40]; L. lycioides (L.) Hassl. (chucluma), in the Lima region of Perú [45]. In addition, L. moziniana (Dunal) Bitter is a semi-cultivated species which is endemic to the Mexican highland plateau and Oaxaca [73]. It is employed for domestic consumption, small-scale commercial use and ceremonial purposes. Other species, known for edible leaves, are: L. laevis (Dunal) Bitter (kattumudunga), in Kerala, Southern India [74] and L. synanthera (Sendtn.) Bitter (Figure 7) eaten in Guatemala as chomte [75]. Thirty-eightspecies of Lycianthes are endemic to Mesoamerica [76] and it is here that there are undoubtedly more examples of this genus being employed as food.

Figure 7.
L. synanthera: panama. Copyright: Thomas Croat, used with permission (http://www.tropicos.org).
Figure 7.
L. synanthera: panama. Copyright: Thomas Croat, used with permission (http://www.tropicos.org).

3.2. Tribe Solandreae Miers
Subtribe Juanulloinae sensu Olmstead et al. [25].
Markea Rich.
Is a genus of around 10 species of shrubs, some of which are epiphytes or vines, distributed through Central and South America. The fruit of Markea is distinctive [77]in being a berry with a thin, papery pericarp that dries to be translucent.At least one (unidentified) species found in Southern Ecuador has edible berries [36], whichis probably M. fosbergii Hunz. Recent investigations of this genus are limited and more plant collections are urgently needed in countries, such as Peru [78].
3.3. Tribe Lycieae Hunz.
Lycium L.
Is a genus of between 75 and 85 species of shrubs with a worldwide distribution. Two species, L. barbarum L., ning xia gou qi, and L. chinense Mill., gou qi, are widely consumed in the Far East, particularly China and parts of the Himalayas. The majority of commercial cultivation takes place in Northern China [79]. The fleshy berries (Figure 8) of both species are eaten; those of L. barbarum [53,79] and L. chinense [53] are believed to have tonic properties. L. barbarum fruits are also used to make wine, and the seeds from L. chinense used for making cooking oil. The leaves of both species are also consumed in China-those of L. barbarum for tea and the young leaves of L. chinense as a vegetable. The dried fruits of both species have also become popular in Europe, where they are known as goji berries, and are considered to be “superfruits” on account of their high vitamin and mineral content.

Figure 8.
L. barbarum: typical fruits on naturalised bush, east coast of Jutland, Denmark. Courtesy of Sten Porse (http://en.wikipedia.org).
Figure 8.
L. barbarum: typical fruits on naturalised bush, east coast of Jutland, Denmark. Courtesy of Sten Porse (http://en.wikipedia.org).

Several other species are employed as “famine foods,”i.e., an additional nutritional source when regular dietary food is scarce. Examples include: L. pallidum Miers, whose fruits are traditionally eaten by the Hopi indigenous people of Northeastern Arizona [80] and L. ferocissimum Miers, the African boxthorn, whose fruits are consumed in Southern Africa. In addition, the fruits of L. europaeum L. and L. intricatum Boiss. are eaten by desert people in Morocco, who also consume the seedlings of both species as vegetables [81]. The leaves of L. acutifolium E. Mey. are consumed in South Africa [82] and its roots may be used to provide extra starch for extending bread flour [83].
3.4. Tribe Physaleae D’Arcy
The Physaleae is a major tribe comprising around 25 genera with about 200 species [26]. Historically, opinions have differed over the taxonomic positions of various genera and their synonymies. The genera recognised below follow the scheme of Olmstead et al. [26,59].
3.4.1. Subtribe Iochrominae (Miers) Hunz.
Acnistus Schott
Is a monotypic genus consisting of A. arborescens Schltdl. [84], which is a tree found across Southern Mexico, Central America, Northern South America and Eastern Brazil [85]. Its fruits are orange and edible when ripe (Figure 9), and are recorded as being consumed in Nicaragua (guitite; [86]), Colombia (palo de gallina; [87]), Southern Ecuador (sabaluco; [36]), and Brazil (marianeira).

Figure 9.
A. arborescens: fruiting branch, Brazil. Courtesy of Eugênio de Melo (http://www.arvores.brasil.nom.br).
Figure 9.
A. arborescens: fruiting branch, Brazil. Courtesy of Eugênio de Melo (http://www.arvores.brasil.nom.br).

Iochroma Benth.
Is a genus of around around 25 species of shrubs and small treesdistributed atmid-elevation from Colombia to Peru, with one species in the Galapagos Islands [85]. Information on edibility is scant, but I. fuchsioides Miers (pepinillo) is semi-cultivated in hedgerows in Ecuador, where its fruits are eaten [88]. Iochroma fruits are also eaten in Peru (Figure 10), and possibly elsewhere. The serminuela, I. australis Griseb., grows in Southern Bolivia, where its yellow fruits are eaten [89]; However, this species was recommended for exclusion from the genus Iochroma [90].

Figure 10.
Iochroma sp: wild, edible fruits, Peru. Courtesy of University of Nebraska-Lincoln, Lincoln, NE, USA (http://newsroom.unl.edu/).
Figure 10.
Iochroma sp: wild, edible fruits, Peru. Courtesy of University of Nebraska-Lincoln, Lincoln, NE, USA (http://newsroom.unl.edu/).

Vassobia Rusby
Is a genus of twospecies of shrubs and small trees [85].Information on the food value of this genus is scarce.V. breviflora (Sendtn.) Hunz. is a spiny shrub whose fruit (fruta de sabia; Figure 11) is eaten in Bolivia, Southern Brazil, Paraguay, and North Argentina [91]. The other species, V. dichotoma (Rusby) Bitter is a tree of the cloud forests of Bolivia [92] and is likely to provide valuable forage food for nearby human communities.

Figure 11.
V. breviflora: fruiting branch. Copyright: Missouri Botanical Garden, used with permission (http://tropicos.org).
Figure 11.
V. breviflora: fruiting branch. Copyright: Missouri Botanical Garden, used with permission (http://tropicos.org).

3.4.2. Subtribe Physalinae Hunz.
Chamaesaracha (A. Gray) A. Gray
Is a genus consisting of 10 perennial herbs found in the Southern USA, Mexico, and Central America. There is record of historical use of the fruits of C. coronopus (Dunal) A.Gray (the small ground-cherry; Figure 12) in the diet of the Hopi and Navajo peoples of the Southwestern USA [93]. It is likely that this species and possibly up to another fourrelated species are still employed as forage foods in the arid regions of North (and possibly Central) America.

Figure 12.
C. coronopus: habit, Arizona. Courtesy of Sue Carnahan (swbiodiversity.org).
Figure 12.
C. coronopus: habit, Arizona. Courtesy of Sue Carnahan (swbiodiversity.org).

Physalis L.
Is a genus of about 90 species [94]. They are mostly scrambling or semi-erect, herbaceous or semi-woody plants, primarily centred in the New World. Physalis has its centre of diversity in Mexico, with over 70 species, many of which are endemic [94]. Distribution radiates from here to the USA, Central America, South America, and the Caribbean [21].
Of the threecultivated species P. peruviana L., the uchuva or uvilla del monte, is widely cultivated in the Andes, and has been introduced into Asia, Australia, and South Africa [32]. It is well-known for its juicy, yellow or orange fruits which are enclosed in a “husk,” formed from the persistent, inflated calyx at maturity (Figure 13). This is a typical feature of this genus. P. philadelphica Lam. is grown across Mexico and Guatemala, at altitudes of around sea level to 2600m, and is known as tomatillo or miltomate [2]. It is popular for its unripe fruits, which may be eaten raw or cooked, whilst the husk may also be used in the preparation of dough to give extra stretchiness. Most cultivars are green-fruited, but some have purple fruits (Figure 14). P. alkekengi L. (Chinese lantern plant, sometimes called Alkekengi officinarum Moench), is believed to be the only native Physalis to China. Apart from being cultivated worldwide for its ornamental value, some cultivars of this species are grown in Cuba for their edible fruits [95] and encouraged to survive elsewhereas a forage fruit.

Figure 13.
P. peruviana: detail of fruiting bush, UK.
Figure 13.
P. peruviana: detail of fruiting bush, UK.


Figure 14.
P. philadelphica: purple-fruited form, cv. Purple Tomatillo.
Figure 14.
P. philadelphica: purple-fruited form, cv. Purple Tomatillo.

The fruits ofnumerous wild species of Physalis are consumed in Mexico and tropical America. Examples given in the survey of Martinez [94] include: P. chenopodifolia Lam., P. coztomatl Moc. and Sessé ex Dunal, P. leptophylla B.L. Rob. and Greenm. (tomate del monte, Northwest Mexico), P. nicandroides Schltdl. (tomate zope, Southern Mexico and Guatemala), P. pruinosa L. (tomate de culebra, Mexico) and P. pubescens L. (uva de perro, Mexico, El Salvador, Colombia, Ecuador, Peru, and Bolivia). P. pubescens has also become naturalised inCentral and Southern USA [93].
Other wild species include P. sulphurea (Fernald) Waterf., consumed in Southern Mexico [96], as well as P. pringleri Greenm. and P. stapelioides (Decne. ex Regel) Bitter, both known as tomatillo, and consumed in Northeastern Mexico. P. cordata Mill. (popita; [97]) and P. minuta Griggs produce edible fruits, and are found from Mexico to Panama, whilst P. victoriana J.M. Toledo, whose fruits are also eaten, is endemic to Northern Argentina[98]. The tropical American P. viscosa L. (uvilla del campo), is consumed in Argentina [91] and elsewhere, and has become naturalised amongst cultivations east of the Mississippi to the Gulf of Mexico [99]. P. longifolia Nutt. is distributed from Nebraska to Texas, USA, and was formerly cultivated by the Zuñi indigenous people [99]; it is likely to be a contemporary dietary component in that region.
P. angulata L. (popa) and P. minima L. (vejiga de perro) are widespread weeds in the Old World, but originate from sub-tropical or tropical America. Both have edible fruits, whilst the leaves of P. angulata may also be eaten as a raw vegetable in Southeast Asia [100]. P.grisea (Waterf.) M. Martinez (dwarf groundcherry) is distributed across Southern Canada and throughout the USA, where it may be encouraged to grow. It is eaten fresh, or in preserves [94].Several other Physalis species above are also semi-cultivated or tolerated as weedsin areas around settlements. In addition, it is likely that several more Physalis species have, as yet, unrecorded food use, particularly in Mexico.
Quincula Raf.
Is a monotypic genus consisting of the herbaceous perennial Q. lobata (Torr.) Raf., the purple ground-cherry, from the Southwest USA and Northern Mexico (Figure 15). Similarly to Physalis, the berries are edible and enclosed in a bladder consisting of the inflated calyx.

Figure 15.
Q. lobata: Texas. Courtesy of William Carr (University of Texas, Austin, Austin, TX, USA).
Figure 15.
Q. lobata: Texas. Courtesy of William Carr (University of Texas, Austin, Austin, TX, USA).

Witheringia L’Hér.
Is a genus of between 10 and 15 tropical herbs and shrubs, ranging from Mexico to Bolivia. Although fruiting plants typically produce berries (Figure 16) there is only record of the leaves being consumed. W. meiantha (Donn. Sm.) Hunz. (ashenté) occurs in Mexico and Central America, and its cooked leaves are consumed in Chiapas, Mexico [101]. Similarly, the leaves of W. solanacea L’Hér. (hierbamora cimarrona), a species of coffee plantations, are consumed in the Sierra Norte de Puebla, Eastern Mexico [102].

Figure 16.
W. meiantha: Costa Rica. Courtesy of http://efg.cs.umb.edu/keys/html.
Figure 16.
W. meiantha: Costa Rica. Courtesy of http://efg.cs.umb.edu/keys/html.

3.4.3. Subtribe Withaninae Bohs and Olmstead
Withania Pauquy
Is a genus of around 10 species of perennial herbs or semi-woody, unarmed shrubswith a distribution covering the Canary Islands, Southern Spain, the Southern Mediterranean, parts of Africa, the Middle East, and South Asia [103]. Flowers have an inflated calyx which develops to surround the berry to a greater or lesser extent. W. frutescens (L.) Pauquy berries (Figure 17) are eaten by children in Morocco [81]. In parts of India (and elsewhere), the fruits of W. coagulans Dunal are used in coagulating milk, in preparation for cheese-making [104]. Similar use has been reported for W. somnifera (L.) Dunal.

Figure 17.
W. frutescens, Southwest Morocco. Courtesy of Jean-Claude Thiaudière (Plant Biodiversity of South-western Morocco, http://www.teline.fr).
Figure 17.
W. frutescens, Southwest Morocco. Courtesy of Jean-Claude Thiaudière (Plant Biodiversity of South-western Morocco, http://www.teline.fr).

3.4.4. Subtribe Salpichroinae sensu Olmstead et al. [25]
Salpichroa Miers
Consists of around 15 species of perennial herbs, sub-shrubs or shrubs of South America, largely centred in the Andes. Around sixspecies are known to have edible fruits, all of which are ovoid or conical in shape and which generally mature to become yellow, orange or purple-coloured, with a maximum length of 3.5 cm. S. glandulosa Miers (ayanata), S. microloba Keel (antarupa), S. ramosissima Miers (callalluma) and S. tristis Walp.var. lehmannii (Dammer) Keel (frutilla) are all wild shrubs or sub-shrubs whose fruits are consumed in the region around Lima, Peru [45]. In addition, S. diffusa Walp. is eaten in Northern Ecuador, where it is known as hiru lulu, as well as in Southern Ecuador [36,105]. S. origanifolia (Lam.) Thell. (huevito de gallo; Figure 18), has a broad distribution in South America and is also an adventive of the USA, Europe, Africa, Australia, etc. [21]. Its fruits are eaten in various regions, e.g., Chile [106] and elsewhere.Several other Salpichroa species may provide forage fruits for local Andean populations, most likely in Peru.

Figure 18.
S. origanifolia: detail of immature fruit (approx. 1.5 cm long), cultivated, Italy. Courtesy of Franz Neidl (http://www.actaplantarum.org).
Figure 18.
S. origanifolia: detail of immature fruit (approx. 1.5 cm long), cultivated, Italy. Courtesy of Franz Neidl (http://www.actaplantarum.org).

3.5. Tribe Solaneae Miers
Jaltomata Schltdl.
Is a genus of approximately 60 perennial herbs or shrubs, distributed from Southeastern Arizona, USA, to Southern Bolivia, as well as three of the Galapagos islands, Cuba, Jamaica, Haiti, Dominican Republic, and Puerto Rico [107]. The majority of species are Andean, with a high incidence of species in Peru (Figure 19). One of the most common, edible species, J. repandidentata (Dunal) Hunz., is distributed throughout the continental range, from Mexico to Bolivia [108]. Jaltomata fruits are globose berries and may be green, white, orange, red, or blackish-purple, often with a typically reflexed calyx. Those of approximately 21 species are eaten in Mexico, Guatemala, Colombia, Ecuador, Nicaragua, Peru, and Bolivia [32,36,45,86,105,107,109].Examples include: J. aspera (Ruiz and Pav.) Mione (chucrume), eaten in Peru; J. chihuahuensis (Bitter) Mione & Bye, in North-Eastern Mexico; J. bicolor (Ruiz andPav.) Mione (antarupa), in Peru; J. herrerae (C.V. Morton) Mione (ahuaimantu), in Bolivia and Peru; J. dentata (Ruiz andPav.) Benitez (shucullmay), in Peru; J. lojae Mione (uvilla), in Southern Ecuador and Peru; J. procumbens (Cav.) J.L. Gentry (uva de monte, sometimes found in gardens, or wild) in Mexico, Central America, Columbia, and Ecuador (Figure 20); J. propinqua (Miers) Mione and M. Nee (shucrume)in Peru; J. repandidentata (Dunal) Hunz., in Guatemala; J. viridiflora (Kunth) M. Nee and Mione (uvilla), in Ecuador. The young leaves of J. repandidentata in Guatemala, Nicaragua, and Mexico, and probably those of J. procumbens in Mexico, are cooked and consumed like spinach.

Figure 19.
J. cajamarca: Cajamarca, Peru. (scale = 1 mm. markings). Courtesy of Thomas Mione (http://web.ccsu.edu/faculty/mione/jaltomat.htm).
Figure 19.
J. cajamarca: Cajamarca, Peru. (scale = 1 mm. markings). Courtesy of Thomas Mione (http://web.ccsu.edu/faculty/mione/jaltomat.htm).


Figure 20.
J. procumbens. Courtesy of Thomas Mione (http://web.ccsu.edu/faculty/mione/jaltomat.htm).
Figure 20.
J. procumbens. Courtesy of Thomas Mione (http://web.ccsu.edu/faculty/mione/jaltomat.htm).

Solanum L. (including Cyphomandra Sendtn. and Lycopersicon Mill.)
Is the largest genus in the Solanaceae and one of the 10 largest plant genera [110]. Its exact size is difficult to establish, but there are at least 1350 species. Solanum species may be herbaceous annuals, herbaceous perennials, tuber-bearing perennials, climbing vines, semi-epiphytes, semi-aquatics, subshrubs, shrubs, or trees. They are distributed across much of the world, with the largest concentration of species in South America.
As the genus Solanum is so large and diverse, the infrageneric framework presented below can only cover subgenera and sections. These follow the synopsis of Child andLester [54], but with the incorporation of Cyphomandra into Solanum subgenus Solanum (rather than standing alone as a distinct genus).The vast diversity of the genus means that the following information should be regarded as anoverview (and not an exhaustive account) of the range of edible species of Solanum across the world.
Subgenus Archaesolanum Marzell.
This subgenus comprises eightspecies of herbaceous to soft-wooded shrubs, found in Australia, Melanesia, and New Zealand [111]. S. aviculare G. Forst. (poroporo; Australia, Papua New Guinea and nearby islands), S. laciniatum Aiton (poroporo; Australia and New Zealand; Figure 21), S. simile F. Muell. (oondoroo; Australian endemic) and S. vescum F. Muell. (gunyang; Australian endemic) are known as kangaroo apples, and consumed by the indigenous peoples of Australia [44] and elsewhere. A further species, S. symonii H. Eichler, is closely related to S. simile and S. vescum [111] and may also be edible.

Figure 21.
S. laciniatum: immature fruits with foliage, Royal Botanic Gardens, Kew.
Figure 21.
S. laciniatum: immature fruits with foliage, Royal Botanic Gardens, Kew.

Subgenus Solanum L.
This subgenus comprises about 170 species of diverse habit and life form, with a cosmopolitan distribution, but with the largest centre of diversity in the neotropics.
Section Holophyllum (G. Don) Walp.
S. spirale Roxb. is a species with a range covering Southern China, and South Asia to Northeast Australia. The orange-yellow fruits and the leaves are eaten in Assam, North-East India, where it is called titakuchi [31], as well as in Thailand and Laos.Additionally, in section Holophyllum is S. diphyllum L., a species distributed from lowland Mexico through Guatemala to Costa Rica [112]. In Mexico, the Pima people of the mountains of Chihuahua sometimes consume the ripe, red fruits [113].
Section Pachyphyllum Dunal
The only domesticated species in this section is S. betaceum Cav. (=Cyphomandra betacea [Cav.] Sendtn.) the tomate de árbol (Figure 22). It is a small, short-lived tree, cultivated in the Andean region for its fleshy fruits [114], which have a flavour of tomato/mango [115]. It is grown both for subsistence use and for its horticultural value. S. betaceum is grown commercially in New Zealand, where it is known as the “tamarillo”, and is also cultivated on a smaller scale in parts of the Mediterranean and elsewhere. Diversity within this cultigen can be assessed by the designation of five cultivar groups based on fruit shape and colour: orange, orange/pointed, purple, red, and red/conical [116]. In addition to S. betaceum, section Pachyphyllum contains over 30 wild relatives, several of which are edible, such as S. cajanumense Kunth (casana; Colombia, Ecuador, N. Peru), S. circinatum Bohs (=Cyphomandra hartwegii (Miers) Walp.; Southern Mexico to Bolivia, Brazil, Venezuela, Guianas), S. paralum Bohs (=Cyphomandra heterophylla Taub.; eastern Brazil). Some newly-described species in section Pachyphylla have edible fruits, for example S. zumbense Bohs, from Southern Ecuador [117]. It is likely that several more species of this section and of the closely-related sections Cyphomandropsis Bitter, Allophylla (Child) Bohs, and Pteroidea Dunal are eaten by local populations in the Andes.

Figure 22.
S. betaceum: plantation, Northern Andes.
Figure 22.
S. betaceum: plantation, Northern Andes.

Section Solanum Seithe
This section is known as the black nightshade complex. The black nightshades (Solanum nigrum L. and related species) are worldwideweeds of arable land, gardens, rubbish tips, and soils rich in nitrogen, in moderately light, warm situations [118]. They are widely used as leafy vegetables, as a source of fruit, and for various medicinal purposes; their consumption is widespread, particularly in Africa and Southeast Asia [118].
The11black nightshade species listed below have well-documented food use, including several which are cultivated for small-scale, local consumption. S. americanum Miller, the American nightshade (Figure 23) is found throughout lowland tropics and subtropics. It is often cultivated locally and the fruits and leaves may be consumed, e.g., in Brazil, where it is known as yerba morro. S. nigrum, the black nightshade, occurs across the world as a weed; forms with reduced bitterness and toxicity are also cultivated for their shoots and fruits. In Kerala, Southern India, where the wild fruits are eaten by indigenous people, it is known as mudungachapu [74] and, in El Salvador, the cultivated form is known as hierba mora. The garden huckleberry, S. scabrum Mill. is widely cultivated for its fruits and shoots in Africa; it has been introduced into North America, Europe, Australia, and New Zealand as a small-scale fruit crop. S. retroflexum Dunal was introduced commercially into the USA at the beginning of the twentieth century, when it became known as the “sunberry;” fruits and leaves are also consumed in many parts of Africa, where it is occasionally cultivated. S. memphiticum Gmel. is a weed of cultivation whose leaves are eaten as a vegetable in Southern Kenya [119]. S. grossidentatum A. Rich. is cultivated for its leaves in Kenya, where it is known as sujet; fruits and leaves are consumed in other parts of Africa. S. chenopodioides Lam. (velvety nightshade; leaves), S. florulentum Bitter (enswiga; leaves), S. physalifolium Rusby var. nitibaccatum (Bitter) Edmonds (Argentinian nightshade; fruits), S. tarderemotum Bitter (soko; leaves) and S. villosum Bitter (red-fruited nightshade; fruits and leaves) are consumed in various parts of Africa; S. villosumleaves are also consumed in Greece [118].

Figure 23.
S. americanum: Florida. Courtesy of Rufino Osorio (http://rufino-osorio.blogspot.co.uk).
Figure 23.
S. americanum: Florida. Courtesy of Rufino Osorio (http://rufino-osorio.blogspot.co.uk).

Subgenus Potatoe (G. Don) D’Arcy
This group consists of around 330 species of sprawling, creeping, erect, or climbing herbs, herbaceous perennials with underground stem tubers, subshrubs, shrubs, or vines, distributed in the USA, Mexico, Central and South America, Madeira, some of the Canary Islands and Eurasia [54].
Section Anarrhichomenum Bitter
S. brevifolium Dunal is found in Venezuela, Colombia, Ecuador and Peru [112]. It is a slender vine whose fruits are consumed in Southern Ecuador, where it is known as uchuchi by the Shuar ethnic group [36].
Section Basarthrum Bitter
The cultivated crop S. muricatum Aiton, known as cachum or pepino dulce, as well as a dozen or so wild relatives, belong in this section [54]. The pepino dulce is a scrambling subshrub producing fruits with a flavour of melon/cucumber. Fruit colour may be white, cream, yellow, maroon, or purplish, sometimes with purple stripes at maturity, whilst the shape may be spherical, conical, heart-shaped or horn-shaped (Figure 24). It is a well-known cultigen of the Andes, with Southern Colombia/Northern Ecuador as the main centre of diversity; this region has been suggested as the centre of origin of the crop [120]. It is often grown for local consumption, but also grown commercially in Ecuador, Peru, and Chile, primarily for export to the USA and Europe. Commercial production has also been attempted in New Zealand, with most exports reaching the Japanese market [121]. Cultivation is possible in temperate European conditions [22,122] as long as winter protection is provided, or the crop is grown as an annual.
There is no clearly-defined wild ancestral form of the pepino [123]. However, at least one Andean wild relative of S. muricatum is known to be edible-the cimbalo, S. caripense Dunal, which has edible fruits with an acid-sweet taste [42]. The two other wild relatives S. basendopogon Bitter and S. tabanoense Correll, both of Peru, may also be picked from the wild and consumed. All three wild species represent an important genetic resource for the breeding of pepino.

Figure 24.
S. muricatum: fruit diversity, UK.
Figure 24.
S. muricatum: fruit diversity, UK.

Section Lycopersicum (Tourn.) Wettst.
There are 13 or so species in this group [124], the best-known of which is the cultivated tomato-S.lycopersicum L. (=Lycopersicon esculentum Mill.), a popular, worldwide crop (Figure 25). This species is only known in cultivation, apart from plants which have become naturalized after escape. The remaining 12 wild species are found in the Galapagos Islands, Central Ecuador through to Peru, and Northern Bolivia to Central Chile.

Figure 25.
S. lycopersicum: commercial cultivar. Courtesy of Friedrich Bӧhringer (http://commons.wikimedia.org).
Figure 25.
S. lycopersicum: commercial cultivar. Courtesy of Friedrich Bӧhringer (http://commons.wikimedia.org).

Several species produce fruits of differing colours at maturity-these are notably: S. peruvianumL. (greenish and purple-suffused/purple-striped), S. chilense (Dunal) Reiche (green to whitish-green, with purple stripes), S. pimpinellifolium L. (red), and S. habrochaites S. Knapp and D.M. Spooner (green with dark green stripes). Only S. pimpinellifolium is known to be gathered from the wild and consumed. Little information on the edibility of the tomato wild relatives S. juglandifolium Dunal, S. lycopersicoides Dunal, S. ochranthum Dunal and S. sitiens I.M. Johnst. is available yet.
Section Petota Dumort.
This sectioncontains one of the most commercially important crops of the world, S. tuberosum L. (=S. tuberosum subsp. tuberosum), which seems to have been derived from S. tuberosum subsp. Andigena (Juz. and Bukasov) Hawkes. In section Petota belongs a number of species of mostly herbaceous perennials, producing edible stem tubers, distributed from the Southern USA to Chile and Argentina. Several Andean species may be found above 4000m altitude, e.g., S. megistacrolobum Bitter. The Centro Internacional del Papa (Peru) has gathered together over 4300 morphologically unique accessions of native potatoes, many of which are uncultivatedandcollectedfrom the wild (Figure 26). These include: S. jamesii Torr. (Southwestern USA and Northwest Mexico), S. stoloniferum Schltdl. and Bouché (Central Mexico), S. cardiophyllum Lindl. (Central Mexico) and S. ehrenbergii (Bitter) Rydb. (Central Mexico). The tubers of most species need to be cooked, but some may be eaten raw. Some bitter-tasting tubers are traditionally processed by freezing, washing and dehydrating into chuño, moraya or tunta to make them edible and storable [125]. Seven species are regionally cultivated [126]: S. ajanhuiri Juz. and Bukasov (Southern Peru, Northern Bolivia), S. chaucha Juz. and Bukasov (Central Peru to central Bolivia), S. curtilobum Juz. and Bukasov (Central Peru to Central Bolivia), S. juzepczukii Bukasov (Central Peru to Northern Bolivia), S. phureja Juz. and Bukasov (Venezuela, Colombia, Ecuador, Peru, Northern Bolivia), S. stenotomum Juz. and Bukasov (Peru to Central Bolivia), and S. tuberosum L. subsp. andigena (Southern Peru to Northern Bolivia), and are suited to altitudes of between 3000 and 4300 m [112,125].

Figure 26.
Selection of Andean wild potatoes. Courtesy of Centro Internacional de la Papa, Peru (http://www.cipotato.org).
Figure 26.
Selection of Andean wild potatoes. Courtesy of Centro Internacional de la Papa, Peru (http://www.cipotato.org).

Subgenus Brevantherum (Seithe) D’Arcy
This small subgenus comprises around 80 species of shrubs, trees and climbers of the Neotropics, often naturalised elsewhere.
Section Brevantherum Seithe
S. erianthum D. Don (Figure 27) is a common species of Southern Florida, Mexico to Panama, Greater Antilles, and Colombia, and has been introduced into South and Southeast Asia, tropical Africa and the Western Pacific. In Southern India the unripe fruits, known as kattuchundai, are eaten by the Palliyar ethnic group [62]. It seems that they are consumed in advance of the development of any potentially toxic compounds known in this species.
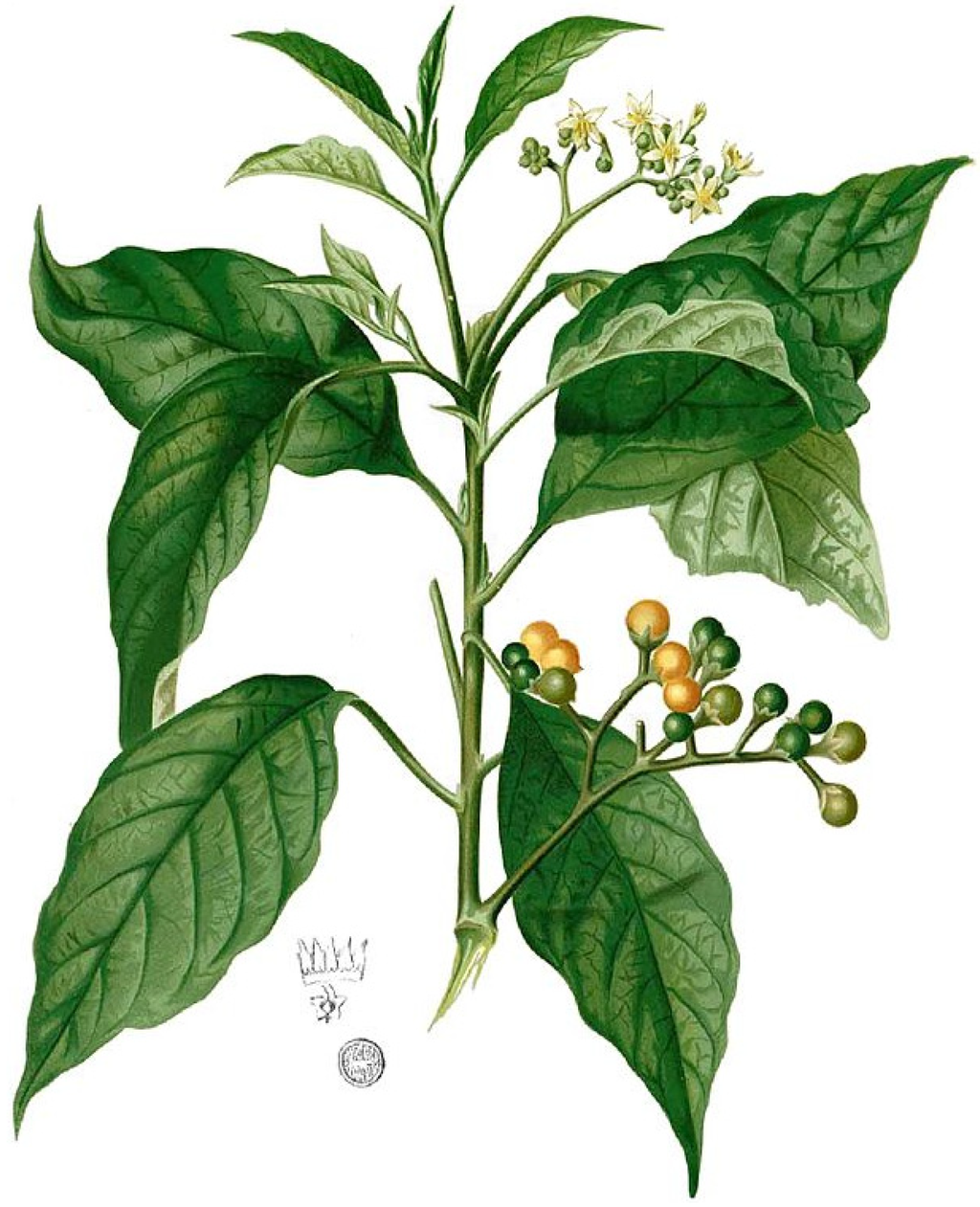
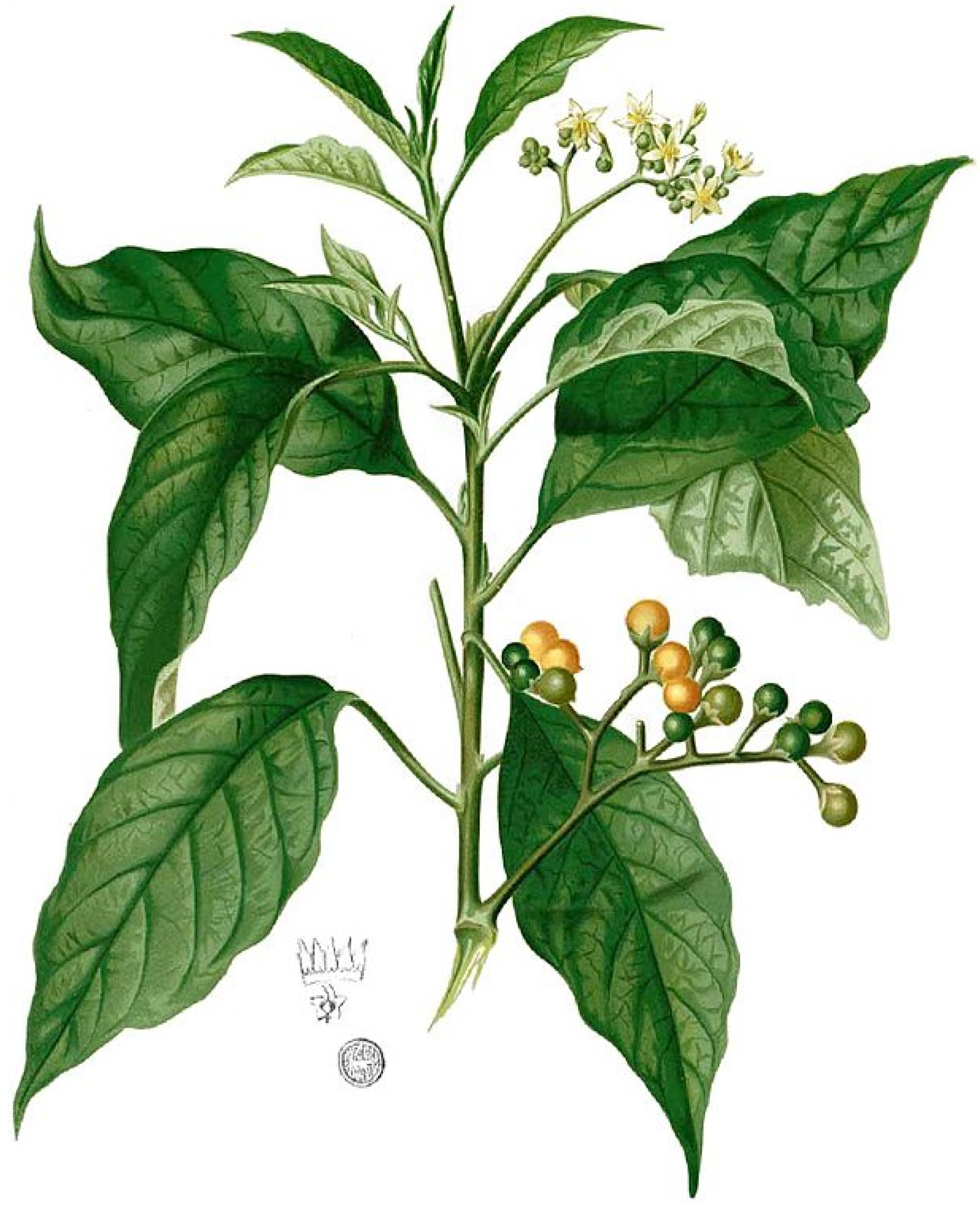
Figure 27.
S. erianthum (as S. mauritianum Blanco): Flora de Filipinas [127].
Figure 27.
S. erianthum (as S. mauritianum Blanco): Flora de Filipinas [127].
Subgenus Leptostemonum (Dunal) Bitter
This group contains around 450 species [55] of diverse habit and life-form, of warm temperate to tropical, cosmopolitan distribution, with about 200 species distributed across the Neotropical region [43]. They are commonly called the “spiny solanums”.
Section Acanthophora Dunal
S.aculeatissimum Jacq. (syn. S. khasianum C.B. Clarke), known as arrebenta-cavalo in Brazil, is probably native to South America, but now common in many tropical and sub-tropical regions. Its roasted fruits are eaten in Assam, and considered useful in deterring internal parasites [31]. S. viarum Dunal (syn. S. khasianum var. chaterjeeanum Sengupta and Sengupta), the tropical soda-apple, also originates from South America and is now found as an adventive in the Southern USA and the Old World [112]. Its leaves and fruits are eaten as vegetables in parts of South Asia.
Section Crinitum Whalen ex Child
S. grandiflorum Ruiz and Pav. is a forest tree of up to 15m of Western Amazonia and the Andean foothills of Colombia, Ecuador, and Peru. It produces edible, pulpy fruits, known as kukush by the Shuar people of Ecuador [33]. S. lycocarpum A. St.-Hil. is a robust shrub of Paraguay and Brazil (Figure 28). It is known in Brazil as the lobeira (wolf’s plant), as it produces fruits of up to 20 cm which are prominent in the diet of the maned wolf. They are also used for making jams and preserves for human consumption. Other large-fruited species in section Crinitum, such as the newly-described S. falciforme Farruggia [128] may have, as yet, unrecognised nutritional value.

Figure 28.
S. lycocarpum: fruits (approx. 15 cm diameter) and flowering branch, Brazil. Courtesy of Eugênio de Melo (http://www.arvores.brasil.nom.br).
Figure 28.
S. lycocarpum: fruits (approx. 15 cm diameter) and flowering branch, Brazil. Courtesy of Eugênio de Melo (http://www.arvores.brasil.nom.br).

Section Cryptocarpum Dunal
S. sisymbriifolium Lam., the litchi tomato, produces small edible fruits in a persistent spiny calyx. It is native to Southern Central South America, but now occurs as a weed in many tropical and subtropical regions. It is also cultivated in gardens across the world as a curiosity fruit crop (Figure 29). S. tridynamum Dunal is endemic to Mexico, where it is known as palohusi pusira. The Guarijios of the Sierra Madre use its unripe fruits which are mashed and added to milk to aid coagulation during cheese-making [129], as with several other spiny Solanum species.

Figure 29.
S. sisymbriifolium: plant cultivated in France. Courtesy of http://www.forum-jardins.com.
Figure 29.
S. sisymbriifolium: plant cultivated in France. Courtesy of http://www.forum-jardins.com.

Section Giganteiformia (Bitter) Child
The aptly-named S. giganteum Jacq., the red bitter berry, is a tall shrub which reaches 6m.It is distributed across sub-saharan Africa, and has been introduced into Australia, Sri Lanka, and India. In Uganda, where it is known as omutarabusa, its small, red or orange berries are gathered from the wild, cooked and consumed in soups [37]. The berries are collected from naturalised plants by people of the Western Ghats, India, who also use them as a vegetable they call chichurdi [130]. S. goetzei Dammer is a medium-sized shrub of Eastern and Southeastern Africa. Its leaves are consumed in Northeast Tanzania [119]. S. schumannianum Dammer is a wild, East African species which may be cultivated near dwellings. Its fruits are consumed raw or cooked in Tanzania, and juice from the fruits may be added to a vinegar substitute [131]. S. kwebense N.E. Brown is a shrub of Southern Africa; its small red or yellow berries are consumed [56].
Section Herposolanum Bitter
S. alternatopinnatum Steud. (syn. S. juciri Sendtn.) is a species of Southern Brazil and Paraguay whose fruit is consumed in Sao Paulo [132] and probably elsewhere.
Section Lasiocarpa D’Arcy
In section Lasiocarpa, the “woolly-fruited” solanums, there are three domesticated species, at least five semi-cultivated taxa, and six wild species with edible fruits. S. lasiocarpum Dunal (=S. ferox auct. non L.) is found across tropical Asia to New Guinea and Northeastern Australia. It is often cultivated in Asia where it is known as the “hairy eggplant” on account of its fruits, which develop a hairy pericarp. There seem to be forms (S. lasiocarpum var. domesticum Heiser), which are cultivated in gardens, and which are non-prickly and have green-fleshed fruits of more than 3 cm in diameter. Semi-cultivated or wild forms (S. lasiocarpum var. lasiocarpum) are from secondary forests or occur as tolerated weeds, and are prickly, with orange-fleshed fruits of around 2 cm in diameter [133].
Similarly, for S. stramonifolium Jacq., the coconilla, (Figure 30) there are semi-cultivated forms, but these may be spiny or non-spiny [134] known as S. stramonifolium var. stramonifolium and S. stramonifolium var. inerme (Dunal) Whalen, respectively [135].These are grown in dooryards by indigenous Amazonians in the Upper Orinoco Basin, in Venezuela [134] and elsewhere in Venezuela, Colombia, Ecuador, Peru, and Brazil.
S. sessiliflorum Dunal, the cocona, is a semi-woody shrub known for its juicy fruits, which may reach 12 cm in diameter, and have a taste somewhere between a peach and a tomato.It is widely grown in and around the Amazon region [136] and cultivation has also been attempted in Puerto Rico, Trinidad, and Florida. Again, two main forms exist–the spiny var. georgicum (R.E. Schult.) Whalen and the spineless var. sessiliflorum. Indigenous groups may plant these forms in their swiddens, or gatherfruits from semi-wild plants growing near their settlements. Var. sessiliflorum may also be cultivated on a small scale and its fruits sold locally, or grown on a larger scale, as in Peru. S. repandum G. Forst., found in the islands of the Western Pacific, is semi-cultivated for its fruits.
S. quitoense Lam., the naranjilla, produces hairy fruits with a characteristic, orange-lime taste (Figure 31). It is only known in cultivation-most commonly in upland Southern Colombia, Ecuador and Peru, but also in the Andes of Venezuela and the interior mountain ranges of Costa Rica. Cultivation has also been attempted in Panama, Guatemala, and Africa [137]. In addition, contemporary research into sustainable Central European glasshouse production of the naranjilla is under way in Germany [138,139]. Some cultivated populations may consist of spiny forms, which may be classified as S. quitoense var. septentrionale R.E. Schult. and Cuatrec., whilst the unarmed form is S. quitoense var. quitoense. Some of the commercial naranjilla production in South America depends (up to 90%) on S. quitoense x S. sessiliflorum F1 hybrids [140].
Species of section Lasiocarpa whose edible fruits are collected solely from the wild are: S. candidum Lindl. (syn. S. tequilense A. Gray; Mexico, Guatemala, Belise, El Salvador, Costa Rica, Colombia, Ecuador), S. hirtum Vahl (huevo de gato; eastern Central Mexico, Guatemala to Colombia, Venezuela, Trinidad, Tobago), S. hyporhodium A. Braun and C.D. Bouché (NorthernVenezuela), S. pectinatum Dunal (tumo; Southeastern Mexico, Costa Rica, Panama, Colombia, Peru), S. pseudolulo Heiser (falso lulo; Colombia; Figure 32), S. vestissimum Dunal (syn. S. tumo Heiser; toronja; Colombia, Venezuela).

Figure 30.
S. stramonifolium: fruiting branch. Courtesy of Jim West (http://www.guaycuyacu.net).
Figure 30.
S. stramonifolium: fruiting branch. Courtesy of Jim West (http://www.guaycuyacu.net).


Figure 31.
S. quitoense: picked at around 75% ripeness; hairs removed for market, Ecuador.
Figure 31.
S. quitoense: picked at around 75% ripeness; hairs removed for market, Ecuador.


Figure 32.
S. pseudolulo: mature fruit. Courtesy of Anesto Mezzomo (http://www.crfg.org/g2/main.php?g2_itemId=29).
Figure 32.
S. pseudolulo: mature fruit. Courtesy of Anesto Mezzomo (http://www.crfg.org/g2/main.php?g2_itemId=29).

Section Melongena (Mill.) Dunal
This section includes S. melongena L., the brinjal eggplant, which is a worldwide crop with thousands of localised landraces and many commercial cultivars. Its greatest genetic diversity is to be found in South and Southeast Asia. The edible fruitsare fleshy berries, eaten when immature. They range from 4 to 45 cm long, from 3 to 12 cm in diameter, and may be green, white, yellow, pink, violet, or purple in colour [141] (Figure 33).

Figure 33.
S. melongena: cv. Très Hâtive de Barbentane.
Figure 33.
S. melongena: cv. Très Hâtive de Barbentane.

The brinjal eggplant is believed to have been domesticated from S. incanum L., the bitter tomato, a prickly shrub native to Africa, the Middle East and Northwestern India. As for some other wild, spiny solanums, the fruits of S. incanum are too bitter to eat, but may be employed in coagulating milk during cheese-making.
Bitter brinjal, S. virginianum L., is a close wild relative of S. melongena found in South and Southeast Asia. Its fruits are yellow and around 2.5 cm across when ready for cooking and consumption in various parts of India [31,142] and Bangladesh. The fruits of other close wild relatives of the brinjal eggplant in this section may be gathered and consumed, e.g., S. melongena subspecies cumingii (Dunal) J. Samuels of Southeast Asia, often found in disturbed habitats near human settlements.
The Gboma eggplant, S. macrocarpon L. subsp. macrocarpon sensu Lester et al. [56], is cultivated widely throughout tropical Africa [143]. Many cultivars are robust perennials, and all are non-prickly. The edible, glabrous leaves arecooked like spinach, whilst the fruits are cooked in stews. Fruitsare 3–12 cm in diameter, spherical, green, whitish or purplish, sometimes with lighter markings when immature, but ready for eating[143] (Figure 34).Leaf cultivars are grown in Central and West Africa, whilst fruit cultivars found in West Africa, as well as Suriname and parts of the Caribbean. Many landraces have their own designated names; for example, in Ghana, akwaseho, gboma, kade, and mankessim are grown for both fruits and leaves, whilst bui and sarpeiman are cultivated for their fruits only [131]. S. macrocarpon subsp. dasyphyllum (Schumach. and Thonn.) sensu Lester et al. [56] is believed to be the nearest wild relative of S. macrocarpon. It is a spiny, weedy undershrub, and has fruits, which reach 7 cm in diameter. In Uganda [144] and Southern Nigeria it is consumed in a similar way to S. macrocarpon.Semi-wild forms in Uganda are known as kitulatula.

Figure 34.
S. macrocarpon: Rotterdam market.
Figure 34.
S. macrocarpon: Rotterdam market.

The goat apple, S. aculeastrum Dunal subsp. aculeastrum sensu Lester et al. [56] is a wildplant of much of sub-Saharan Africa; some forms found in Kenya produce edible fruits.
S. chippendalei Symon (ngaru), S. diversiflorum F. Muell. and S. dioicum W. Fitzg. (gilu) were considered by Symon [145] to be part of section Melongena. This was called into question by Martine et al. [146], who showed that the first two species were part of the section Melongena “Andromonoecious Bush Tomato” clade, whilst the latter species was placed in the “Dioicum Complex” clade. The precise relationships between these clades and section Melongena sensu stricto have yet to be determined satisfactorily. All threespecies provide fruits, which are gathered from the wild and consumed by the indigenous peoples of Australia. They are often eaten in the dry season, when little fresh vegetable matter is available.
Section Micracantha Dunal
S. barbeyanum Huber (fruto bayo; Figure 35) and S. nemorense Dunal are species of the western lowland Amazon rainforest [112]. Both produce edible fruits which are gathered from the wild. S. jamaicense Mill., Jamaican nightshade, also provides edible fruits and is a common humid lowland weed of Southern Mexico to Bolivia, Brazil, Venezuela, Trinidad, and the Greater and Lesser Antilles [112].

Figure 35.
S. barbeyanum: mature fruits, Madre de Dios, Southern Peru. Courtesy of Piher Maceda (Andes to Amazon Biodiversity Program at the Botanical Research Institute of Texas, http://atrium.andesamazon.org).
Figure 35.
S. barbeyanum: mature fruits, Madre de Dios, Southern Peru. Courtesy of Piher Maceda (Andes to Amazon Biodiversity Program at the Botanical Research Institute of Texas, http://atrium.andesamazon.org).

Section Monodolichopus Bitter
S. coagulans Forssk. is a prickly, perennial herb, distributed across Northeast Africa and Egypt, and has berries of around 1 cm diameter [147]. As the species name implies, it is employed in the coagulation of milk [148].
Section Oliganthes (Dunal) Bitter
The domesticated scarlet eggplant, S aethiopicum L., is placed in this section. This crop plant is grown throughout tropical Africa, as well as Brazil (known as giló), Southern France, Italy, etc. According to Lester and Daunay and Lester and Niakan [143,149] there are four cultivar groups comprising the vast diversity of this crop, as listed below:
S. aethiopicum Gilo Group is typified by the production of fruits which may be more or less spherical to ellipsoid, and from 2 to 8 cm in diameter. The leaves are hairy and are not eaten. Shum Group plants have small, glabrous leaves and shoots that are plucked frequently as a green vegetable. However, the small, bitter fruits are not eaten (Figure 36). Kumba Group plants produce large, glabrousleaves that can be used as a green vegetable, as well as large, grooved fruits of 5–10 cm in diameter that are picked green, or even red, and stewed or eaten raw.The Aculeatum Group comprises purely ornamental plants, probably produced inEurope by hybridisation between S. aethiopicum Kumba Group and S. anguivi Lam.They are prickly and hairy, but with fairly large, grooved fruits.
The wild ancestor of S. aethiopicum is S. anguivi, a shrub bearing small, bitter fruits which is distributed throughout tropical Africa and its neighbouring islands.Lester and Daunay [143] noted that semi-cultivation in village gardens has produced the unarmed S. anguivi subsp. distichum (Schumach. and Thonn.) sensu Lester et al. [56]. Its orange or red fruits reach 1.5 cm in diameter and may be cooked and used as flavouring for stews, etc. (Figure 37). The prickly S. anguivi subsp. anguivi is found wild, near villages and other habitations, and is also consumed by humans [56].

Figure 36.
S. aethiopicum Shum Group berries. Courtesy of Radboud University Botanical and Experimental Garden, Netherlands.
Figure 36.
S. aethiopicum Shum Group berries. Courtesy of Radboud University Botanical and Experimental Garden, Netherlands.


Figure 37.
S. anguivi subsp. distichum: berries at different stages of maturity.
Figure 37.
S. anguivi subsp. distichum: berries at different stages of maturity.

S. kurzii Brace ex Prain is a rare species from Sikkim and Meghalaya, Northeast India.Its precise taxonomic affinity is unclear, but it may be a synonym of S. violaceum Ortega, the Indian nightshade, found across South and Southeast Asia. The fruits of the two shrubs have traditional medicine and food value [31,150,151]. As a consequence they may be encouraged to grow in the vicinities of settlements.
There are several Australian and Polynesian Solanum species which are often included in section Oliganthes (see [145]); however, their precise taxonomic position has yet to be clarified. Of these, the “bush tomatoes”: S. centrale J.M. Black (yakatjiri), S. cleistogamum Symon (yipuntri), S. coactiliferum J.M. Black (warakalukalu), S. ellipticum R. Br. (tawal-tawalpa), S. esuriale Lindl. (puddaddee), S. orbiculatum Dunal, and S. petrophilum F. Muell. (ngaliyirki) are all plants of the Australian arid zone, and are known by indigenous communities for their food value. Fruits may need to be cleared of abrasive hairs, pounded or have the seeds removed before cooking or preservation [152]; the fruits of some species, e.g. S. ellipticum, are edible without preparation [44]. S. hystrix R. Br. is recorded as being eaten in South Australia, but only after removal of seeds and baking with the bark of Eucalyptus sp. [44]. S. uporo Dunal (syn. S. anthropophagorum Seem.), uporo-poro”, is a small shrub of the Polynesian islands, including Fiji and Tahiti (Figure 38). The fruit has an appearance and flavour similar to a cultivated tomato. In previous times the large leaves were allegedly used by cannibalistic tribes to wrap up human flesh in preparation for cooking-hence the taxonomic synonym (transl. “man-eater’s Solanum”) and the common name “cannibal’s tomato”.

Figure 38.
S. uporo: (as S. anthropophagorum) [153].
Figure 38.
S. uporo: (as S. anthropophagorum) [153].

Section Torva Nees
This group of shrubs and small trees is found in many parts of Latin America and the West Indies. However, S. torvum Sw. has become a widespread, naturalised species in many tropical regions of the Old World, where it is sometimes cultivated. On account of its numerous, small, pea-sized berries (Figure 39), it is known as the “pea eggplant”. The cooked, unripe fruits are eaten in parts of South and Southeast Asia, and are an important component of stews and curries.

Figure 39.
S. torvum: for sale at Agomanya market, Ghana. Courtesy of Jan Germer (http://www.virboga.de).
Figure 39.
S. torvum: for sale at Agomanya market, Ghana. Courtesy of Jan Germer (http://www.virboga.de).

The following three species have questionable affinities with section Torva but, pending availability of detailed phylogenetic data, they are retained here for convenience. S. anomalum Thonn. is a shrub of Africa, occurring wild in secondary forests in Sierra Leone to Southern Nigeria, Cameroon, Democratic Republic of Congo and Southern Angola; in Ghana it is known as nsusua twi. The cooked, ripe fruits are eaten [131]. S. barbisetum Nees is distributed across India, Bangladesh, Myanmar, Thailand, and Laos, and is often found as a weed of cultivated fields. Ripe fruits may be eaten in parts of South and Southeast Asia, e.g., in Meghalaya, India [154]. S. paniculatum L., jurubeba, (Figure 40), native to Eastern Brazil, Paraguay and Northeastern Argentina [112] provides berries which are used in the commercial production of wine [155].

Figure 40.
S. paniculatum berries, Brazil. Courtesy of http://yuhanerosangelataishan.blogspot.co.uk/.
Figure 40.
S. paniculatum berries, Brazil. Courtesy of http://yuhanerosangelataishan.blogspot.co.uk/.

4. Conclusions
4.1. Overall Summary
The preliminary findings recorded above indicate that 15 genera in subfamily Solanoideae provide around 180 species that are utilised for food across the world. Based on a family total of 2500 species [26], approximately 7% of Solanaceae species are edible species of subfamily Solanoideae. The regular description of new Capsicum, Jaltomata, Physalis and Solanum species (e.g., [98,156,157,158]), as well as others, suggests that there are undoubtedly numerous other specieswhich do not yet appear in the inventory above. These are likely to be those that are largely gathered from the wild and form part of the dietary repertoire of localised indigenous groups. The majority of new species of this nature is likely to originate in the tropics and, with regard to Solanum species, probably belongs to subgenus Leptostemonum, in which the majority of food species is found. As a realistic figure, then, the overall proportion of Solanaceae species represented by edible members of subfamily Solanoideae may be in the vicinityof 10%. The number of known edible plant species across the world is approximately 30,000 [1], and the number of cultivated species is about 7000 [159]; thus, the Solanoideae provides approximately 0.6% and 2.6%, respectively, of these global totals.
4.2. The Food Species and Genera
The inventory above demonstrates a variety of sources of food species; these may becommercially cultivated, produced on a subsistence scale, semi-cultivated, tolerated as useful weeds, or gathered from the wild. The majority (150) of food species of the Solanoideae belongs tofourgenera: Solanum, Capsicum, Jaltomata and Physalis (Box 1).A considerable number of economically significant food crops are to be found amongst the 30 speciesor so of domesticated crops in the genus Solanum. These include the well-known Solanum tuberosum (Irish potato), S. lycopersicum (tomato), and S. melongena (brinjal eggplant), all favoured on a worldwide scale. Regionally-important domesticated solanums include S. quitoense (naranjilla), S. sessiliflorum (cocona), S. muricatum (pepino dulce), S. betaceum (tomate de árbol), S. aethiopicum (scarlet eggplant), and S. macrocarpon (Gboma eggplant). Numerous other solanums are semi-cultivated or collected from the wild from many parts of the world. In addition, it is likely that the consumption of many more semi-cultivated species, crop wild relative species, and distantly-related wild solanums across the world has yet to be recorded. Fivespecies of Capsicum are cultivated: C. annuum (bell pepper/chilli), C. chinense (Habañero/Scotch bonnet), C. frutescens (bird pepper/Tabasco pepper), C. baccatum (aji), and C. pubescens (rocoto).Around 20 other Capsicum species are semi-cultivated or collected from the wild. All Jaltomata species, which are recorded as edible, e.g., J. procumbens (uva de monte), are collected from the wild. The majority of Physalis food species are also gathered wild, whilst P. peruviana (uchuva) and P. philadelphica (tomatillo) are well-known cultigens in Latin America. Around 30 species from the other 11 genera provide the remainder of species with known food value.
Box 1. List of food species in subfamily Solanoideae (in text order; cultivated crop species = underscored).
Subfamily SOLANOIDEAE Kostel
1. Tribe Capsiceae D’Arcy
Capsicum L.
C. annuum L. var. annuum L.,C. baccatum L. var. pendulum (Willd.) Eshbaugh, C. chinense Jacq., C. frutescens L., C. pubescens Ruiz & Pav., C. annuum var. glabriusculum (Dunal) Heiser & Pickersgill, C. baccatum var. baccatum, C. cardenasii Heiser & P.G. Sm., C. eshbaughii Barboza, C. eximium Hunz., C. chacoense Hunz., C. galapagoense Hunz., C. tovarii Eshbaugh, P.G. Sm. & Nickrent, C. praetermissum Heiser & P.G. Sm., C. rhomboideum (Humb., Bonpl. & Kunth) Kuntze, C. lanceolatum (Greenm.) C.V. Morton & Standl., C. schottianum Sendtn., C. caatingae Barboza & Agra, C .longidentatum Agra & Barboza
Lycianthes (Dunal) Hassl.
L. asarifolia (Kunth & Bouché) Bitter, L. lycioides (L.) Hassl., L. moziniana (Dunal) Bitter, L. laevis (Dunal) Bitter, L. synanthera (Sendtn.) Bitter
2. Tribe Solandreae Miers
2.1. Subtribe Juanulloinae sensu Olmstead et al.
Markea Rich.
M. fosbergii Hunz.
3. Tribe Lycieae Hunz.
Lycium L.
L. barbarum L., L. chinense Mill., L. pallidum Miers, L. ferocissimum Miers, L. europaeum L., L. intrincatum Boiss., L. acutifolium E. Mey.
4. Tribe Physaleae D’Arcy
4.1. Subtribe Iochrominae(Miers) Hunz.
Acnistus Schott
A. arborescens Schltdl.
Iochroma Benth.
I. fuchsioides Miers
Vassobia Rusby
V. breviflora (Sendtn.) Hunz.
4.2. Subtribe Physalinae Hunz.
Chamaesaracha (A. Gray) A. Gray
C. coronopus (Dunal) A.Gray
Physalis L.
P. peruviana L., P. philadelphica Lam., P. alkekengi L., P. chenopodifolia Lam., P. coztomatl Moc. & Sessé ex Dunal, P. leptophylla B.L. Rob. & Greenm., P. nicandroides Schltdl., P. pruinosa L., P. pubescens L., P. sulphurea (Fernald) Waterf., P. pringleri Greenm., P. stapelioides (Decne. ex Regel) Bitter, P. cordata Mill., P. minuta Griggs, P. victoriana J.M. Toledo, P. viscosa L., P. longifolia Nutt., P. angulata L., P. minima L., P. grisea (Waterf.) M. Martinez
Quincula Raf.
Q. lobata (Torr.) Raf.
Witheringia L’Hér.
W. meiantha (Donn. Sm.) Hunz., W. solanacea L’Hér.
4.3. Subtribe Withaninae Bohs & Olmstead
Withania Pauquy
W. frutescens (L.) Pauquy, W. coagulans Dunal, W. somnifera (L.) Dunal
4.4. Subtribe Salpichroinae sensu Olmstead et al.
Salpichroa Miers
S. glandulosa Miers, S. microloba Keel, S. ramosissima Miers, S. tristis Walp. var. lehmannii (Dammer) Keel, S. diffusa Walp., S. origanifolia (Lam.) Thell.
5. Tribe Solaneae Dumort.
Jaltomata Schltdl.
J. repandidentata (Dunal) Hunz., J. aspera (Ruiz & Pav.) Mione, J. chihuahuensis (Bitter) Mione & Bye, J. bicolor (Ruiz & Pav.) Mione, J. herrerae (C.V. Morton) Mione, J. dentata (Ruiz & Pav.) Benitez, J. lojae Mione, J. procumbens (Cav.) J.L. Gentry, J. propinqua (Miers) Mione & M. Nee, J. repandidentata (Dunal) Hunz., J. viridiflora (Kunth) M. Nee & Mione
Solanum L.
Subgenus Archaesolanum Marzell
S. aviculare G. Forst., S. laciniatum Aiton, S. simile F. Muell., S. vescum F.Muell.
Subgenus Solanum L.
Section Holophyllum (G. Don) Walp.
S. spirale Roxb., S. diphyllum L.
Section Pachyphyllum Dunal
S. betaceum Cav., S. cajanumense Kunth, S. circinatum Bohs, S. paralum Bohs, S. zumbense Bohs
Section Solanum Seithe
S. americanum Miller, S. nigrum L., S. scabrum Mill., S. retroflexum Dunal, S. memphiticum Gmel., S. grossidentatum A. Rich., S. chenopodioides Lam., S. florulentum Bitter, S. physalifolium Rusby var. nitibaccatum (Bitter) Edmonds, S. tarderemotum Bitter, S. villosum Bitter
Subgenus Potatoe (G. Don) D’Arcy
Section Anarrhichomenum Bitter
S. brevifolium Dunal
Section Basarthrum Bitter
S. muricatum Aiton, S. caripense Dunal, S. basendopogon Bitter, S. tabanoense Correll
Section Lycopersicum (Tourn.) Wettst.
S. lycopersicum L., S. pimpinellifolium L.
Section Petota Dumort.
S. tuberosum subsp. tuberosum, S. tuberosum L. subsp. andigena (Juz. & Bukasov) Hawkes, S. jamesii Torr., S. stoloniferum Schltdl. & Bouché, S. cardiophyllum Lindl., S. ehrenbergii (Bitter) Rydb., S. ajanhuiri Juz. & Bukasov, S. chaucha Juz. & Bukasov, S. curtilobum Juz. & Bukasov, S. juzepczukii Bukasov, S. phureja Juz. & Bukasov, S. stenotomum Juz. & Bukasov
Subgenus Brevantherum (Seithe) D’Arcy
Section Brevantherum Seithe
S. erianthum D. Don
Subgenus Leptostemonum (Dunal) Bitter
Section Acanthophora Dunal
S. aculeatissimum Jacq., S. viarum Dunal
Section Crinitum Whalen ex Child
S. grandiflorum Ruiz & Pav., S. lycocarpum A. St.-Hil.
Section Cryptocarpum Dunal
S. sisymbriifolium Lam., S. tridynamum Dunal
Section Giganteiformia (Bitter) Child
S. giganteum Jacq., S. goetzei Dammer, S. schumannianum Dammer, S. kwebense N.E. Brown
Section Herposolanum Bitter
S. alternatopinnatum Steud.
Section Lasiocarpa D’Arcy
S. lasiocarpum Dunal var. lasiocarpum, S. lasiocarpum var. domesticum Heiser, S. stramonifolium Jacq. var. stramonifolium, S. stramonifolium var. inerme (Dunal) Whalen, S. sessiliflorum Dunal var. georgicum (R.E. Schult.) Whalen, S. sessiliflorum var. sessiliflorum, S. repandum G. Forst., S. quitoense Lam. var. septentrionale R.E. Schult. & Cuatrec., S. quitoense var. quitoense, S. candidum Lindl., S. hirtum Vahl, S. hyporhodium A. Braun & C.D. Bouché, S. pectinatum Dunal, S. pseudolulo Heiser, S. vestissimum Dunal
Section Melongena (Mill.) Dunal
S. melongena L., S. incanum L., S. virginianum L., S. melongena subspecies cumingii (Dunal) J. Samuels, S. macrocarpon L. subsp. macrocarpon, S. macrocarpon subsp. dasyphyllum (Schumach. & Thonn.) sensu Lester et al., S. aculeastrum Dunal subsp. aculeastrum, S. chippendalei Symon, S. diversiflorum F. Muell., S. dioicum W. Fitzg.
Section Micracantha Dunal
S. barbeyanum Huber, S. nemorense Dunal, S. jamaicense Mill.
Section Monodolichopus Bitter
S. coagulans Forssk.
Section Oliganthes (Dunal) Bitter
S. aethiopicum L., S. anguivi Lam. subsp. anguivi, S. anguivi subsp. distichum (Schumach. & Thonn.) sensu Lester et al., S. kurzii Brace ex Prain, S. violaceum Ortega, S. centrale J.M. Black, S. cleistogamum Symon, S. coactiliferum J.M. Black, S. ellipticum R. Br., S. esuriale Lindl., S. orbiculatum Dunal, S. petrophilum F. Muell., S. hystrix R. Br., S. uporo Dunal
Section Torva Nees
S. torvum Sw., S. anomalum Thonn., S. barbisetum Nees, S. paniculatum L.
4.3. Future Studies and Potential
The inventory above is to be regarded as a “snapshot” of the wide range of edible nightshade species of subfamily Solanoideae across the world. It is a starting point from which further studies may progress to determine a more precise assessment of the extent to which the Solanaceae provides the human race with sources of food. The conservation status of the majority of species listed in the inventory above has not yet been assessed. There is an urgent need to broach this so that the International Union for Conservation of Nature Red List status of all species can be combined with the other details for each entry. A number of solanaceous crop species (particularly in Solanum, Capsicum, and Physalis) are cultivated in regions outside of their centre of origin. Considerable interest has recently been directed towards novel Solanaceae crops for cultivation in Europe (e.g., [14,22,68,122,138,139]), and this kind of potential for many of the lesser-known species in the inventory above merits careful further investigation. With the advent of climate change it may be possible for several of these crops to be grown outside of the climatic zones to which they would normally be limited.
Acknowledgments
This paper is dedicated to Charles Bixler Heiser (1920–2010), author of many important works on nightshadespecies. His latter research on species of Solanum section Lasiocarpa became formative to our understanding of the taxonomy and production of the “woolly-fruited”Solanum crops, such as the naranjilla.
I wish to express my sincere thanks to the following people in Ecuador: Karla Aguaguiña Méndez, Organising Committee of the Primer Congreso Internacional de Biotecnología y Biodiversidad (CIBB CIBE-ESPOL/AEBE) 2012, Guayaquil; Pablo Acosta-Quezada, Universidad Técnica Particular de Loja; Katya Romoleroux, Herbario QCA, Quito; Omar Vacas-Cruz, Herbario QCA, Quito; Wilson Vasquez, Instituto Nacional de Investigaciones Agropecuarias, Quito.
In the UK, the following deserve my grateful recognition: the late Alan Child, Keith Fowler, Richard Lester, John Moschopoulos, and Jim Tennant. Their taxonomic, botanic and ethnobotanic knowledge has proven to be a source of inspiration and encouragement. Many thanks also to John Page, Clive Simms and Owen Smith for plant material and cultivation information.
I would also like to thank the Kent Horticultural Bursary Group (East Malling Research, Kent, UK), the Stanley Smith Horticultural Trust (UK), the Department of Education, Cornwall Council, UK, as well as Alfred Jones and Margot Young, for contributions towards various aspects of a research trip to investigate novel solanaceous crops in Ecuador in 2012.
I gratefully acknowledge the kind assistance of staff at the herbaria visited: Roxali Bijmoer, Naturalis Biodiversity Centre herbarium, Leiden; Anja Buijsen, Naturalis Biodiversity Centre botanical library, Leiden; Esther García Guillén, Real Jardín Botánico herbarium, Madrid; Eugenia Insúa, Real Jardín Botánico library, Madrid; David Goyder, Royal Botanic Gardens, Kew herbarium; Anna Millozza, Università degli Studi di Roma La Sapienza herbarium, Rome; Tan Wee Kiat, Singapore Botanic Gardens herbarium.
Disclaimer
It should be noted that, in many species of the Solanaceae family, various parts including fruits, leaves and roots may contain potentially harmful secondary metabolites, such as alkaloids. Various parts may be edible at certain stages, but toxic at others. Although every attempt has been made to ensure the accuracy of the preceding account, no responsibility can be taken by the author or the publishers for the actions of any third party relating to the information presented above.
Conflicts of Interest
The author declares no conflict of interest.
References
- FAO. The State of the World’s Plant Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 1997. [Google Scholar]
- Bemejo, J.E.H.; Léon, J. Neglected Crops: 1492 from a Different Perspective; FAO Plant Production and Protection Series; FAO: Rome, Italy, 1994. [Google Scholar]
- Thies, E. Promising and Underutilized Species, Crops and Breeds; Deutsche Gesellschaft für Technische Zusammenarbeit (GTZ): Eschborn, Germany, 2000. [Google Scholar]
- Da Silva Dias, J.C. Impact of improved vegetable cultivars in overcoming food insecurity. Euphytica 2010, 176, 125–136. [Google Scholar]
- FAO. Global Plan of Action for the Conservation and Sustainable Utilization of Plant Genetic Resources for Food and Agriculture and the Leipzig Declaration; FAO: Rome, Italy, 1996. [Google Scholar]
- FAO. Rome Declaration on World Food Security and World Food Summit Plan of Action; FAO: Rome, Italy, 1996. [Google Scholar]
- Arce, R.G. De Flores, brotes y palmitos: Alimentos olvidados. Agron. Costarric. 2008, 32, 183–192. [Google Scholar]
- Heiser, C.B. Nightshades: The Paradoxical Plants; Freeman: San Francisco, CA, USA, 1969. [Google Scholar]
- NRC (National Research Council). Lost Crops of the Incas: Little-known Plants of the Andes with Promise for Worldwide Cultivation; National Academy Press: Washington, DC, USA, 1989.
- NRC (National Research Council). Lost Crops of Africa. Volume I: Grains; National Academies Press: Washington, DC, USA, 1996.
- NRC (National Research Council). Lost Crops of Africa Volumn II: Vegetables; National Academies Press: Washington, DC, USA, 2006.
- NRC (National Research Council). Lost Crops of Africa. Volume III: Fruits; National Academies Press: Washington, DC, USA, 2008.
- Padulosi, S.; Amaya, K.; Jäger, M.; Gotor, E.; Rojas, W.; Valdivia, R. A holistic approach to enhance the use of neglected and underutilized species: The case of Andean grains in Bolivia and Peru. Sustainability 2014, 6, 1283–1312. [Google Scholar] [CrossRef]
- Prohens, J.; Rodriguez-Burruezo, A.; Nuez, F. New crops: An alternative for the development of horticulture. Food Agric. Env. 2003, 1, 75–79. [Google Scholar]
- Rudebjer, P.; Meldrum, G.; Padulosi, S.; Hall, R.; Hermanowicz, E. Realizing the Promise of Neglected and Underutilized Species; Bioversity International: Rome, Italy, 2014. [Google Scholar]
- Samuels, J. Capsicum pubescens: An underutilized species of hot pepper. Sol Newsl. 2009, 25, 5–8. [Google Scholar]
- Schelderman, X.; Rojas, W.; Valdivia, R.; Peralta, E.; Padulosi, S. Retos y posibildades del uso de species olvidadas y subutilizadas en un desarollo sostenible. In Proceedings of the Memorias del XII Congreso Internacional de Cultivos Andinos, Cochabamba, Bolivia, 15–19 October 2003; pp. 1–8.
- FAO. Second Global Plan of Action for Plant Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 2012. [Google Scholar]
- Convention on Biological Diversity (CBD). Global Strategy for Plant Conservation, 2011–2020. Available online: www.cbd.int/gspc/targets.shtml (accessed on 2 August 2014).
- Hawkes, J.G. The economic importance of the family Solanaceae. In Solanaceae IV: Advances in Biology and Utilization; Nee, M., Symon, D.E., Lester, R.N., Jessop, J.P., Eds.; Royal Botanic Gardens: Kew, UK, 1999; pp. 1–8. [Google Scholar]
- Hunziker, A.T. South American Solanaceae: A synoptic survey. In The Biology and Taxonomy of the Solanaceae; Hawkes, J.G., Lester, R.N., Skelding, A.D., Eds.; Academic Press: London, UK, 1979; pp. 49–85. [Google Scholar]
- Samuels, J. The Solanaceae-novel crops with high potential. Org. Grow. 2009, 9, 32–34. [Google Scholar]
- FAO. FAOSTAT Crop Production Data for 2010. Available online: http://faostat.fao.org (accessed on 27 February 2012).
- Global Facilitation Unit for Underutilized Species (GFU). Available online: http://www.underutilized-species.org/ (accessed on 31 July 2014).
- Olmstead, R.G.; Sweere, J.A.; Spangler, R.E.; Bohs, L.; Palmer, J.D. Phylogeny and provisional classification of the Solanaceae based on chloroplast DNA. In Solanaceae IV: Advances in Biology and Utilization; Nee, M., Symon, D.E., Lester, R.N., Jessop, J.P., Eds.; Royal Botanic Gardens: Kew, UK, 1999; pp. 111–137. [Google Scholar]
- Olmstead, R.G.; Bohs, L.; Abdel Migid, H.; Santiago-Valentin, E.; Garcia, V.F.; Collier, S.M. A molecular phylogeny of the Solanaceae. Taxon 2008, 57, 1159–1181. [Google Scholar]
- International Plant Genetic Resources Institute (IPGRI). Neglected and Underutilized Plant Species: Strategic Action Plan of the International Plant Genetic Resources Institute; IPGRI: Rome, Italy, 2002. [Google Scholar]
- Stolton, S.; Maxted, N.; Ford-Lloyd, B.; Kell, S.; Dudley, N. Food Stores: Using Protected Areas to Secure Crop Genetic Diversity; Worldwide Fund for Nature: Woking, UK, 2006. [Google Scholar]
- Berlingeri, C.; Crespo, M.B. Inventory of related wild species of priority crops in Venezuela. Gen. Res. Crop Evol. 2012, 59, 655–681. [Google Scholar] [CrossRef]
- Burkill, H.M. The Useful Plants of West Tropical Africa, 2nd ed.; Royal Botanic Gardens: Kew, UK, 2000. [Google Scholar]
- Patiri, B.; Borah, A. The Wild Edible Plants of Assam; Geetakhi Publishers and Printers: Guwahati, India, 2007. [Google Scholar]
- Sanjinés, A.; Øllgaard, B.; Balslev, H. Frutos Comestibles. In Botanica Economica de los Andes Centrales; Moraes, M., Ollgaard, B., Kvist, L.P., Borchsenius, F., Balslev, H., Eds.; Universidad Mayor de San Andres: La Paz, Bolivian, 2006; pp. 329–346. [Google Scholar]
- Santin Luna, F.M. Etnobotanica de las comunidades de la zona alta del Rio Nangaritza. In Memorias del IV Congreso Ecuatoriano de Botanica; Lozano, P., Bussman, R., Navarrete, H., Eds.; Fundacion Ecuatoriana para la Investigacion y Desarollo de la Botanica (FUNBOTANICA): Loja, Ecuador, 2005; pp. 339–361. [Google Scholar]
- De la Torre, L.; Navarrete, H.; Muriel, P.; Macia, M.J.; Balslev, H. Enciclopedia de las Plantas Utiles del Ecuador; Herbario QCA de la Escuela de Ciencias Biologicas de la Pontificia Universidad Catolica del Ecuador: Quito, Ecuador; Herbario AAU del Departamento de Ciencias Biologicas de la Universidad de Aarhus: Aarhus, Denmark, 2008. [Google Scholar]
- Tindall, H.D. Vegetables in the Tropics; Macmillan Press: London, UK, 1983. [Google Scholar]
- Van den Eynden, V.; Cueva, E.; Cabrero, O. Wild foods from southern Ecuador. Econ. Bot. 2003, 57, 576–603. [Google Scholar] [CrossRef]
- Bukenya, Z.R.; Carasco, J.F. Ethnobotanical aspects of Solanum L. (Solanaceae) in Uganda. In Solanaceae IV: Advances in Biology and Utilization; Nee, M., Symon, D.E., Lester, R.N., Jessop, J.P., Eds.; Royal Botanic Gardens: Kew, UK, 1999; pp. 345–360. [Google Scholar]
- Lu, A. Solanaceae in China. In Solanaceae: Biology and Systematics; D’Arcy, W., Ed.; Columbia University Press: New York, NY, USA, 1986; pp. 79–85. [Google Scholar]
- Medina, M.A.; Mena, A.; Prohens, J.; Nuez, F. Survey of cultivated and wild edible plant species used in the Department of Chocó, Colombia. Pl. Gen. Res. Newsl. 2007, 150, 35–44. [Google Scholar]
- Nee, M. The systematics of lesser known edible Solanaceae of the New World. In Solanaceae III: Taxonomy, Chemistry, Evolution; Hawkes, J.G., Lester, R.N., Nee, M., Estrada, N., Eds.; Royal Botanic Gardens: Kew, UK, 1991; pp. 365–368. [Google Scholar]
- Nuez, F.; Morales, R.; Ruiz, J.J.; Fernández de Córdova, P.; Valdivieso, E.; Gonzalez, F.I. Recolección de species horticolas en Ecuador. Pl. Gen. Res. Newsl. 1993, 96, 29–33. [Google Scholar]
- Nuez, F.; Morales, R.; Prohens, J.; Fernández de Córdova, P.; Soler, S.; Valivieso, E.; Solórzano, V. Germplasm of Solanaceae horticultural crops in the south of Ecuador. Pl. Gen. Res. Newsl. 1999, 120, 44–47. [Google Scholar]
- Orozco, C.I.; Beltran, G.D.; Porras, N.; Nee, M. Listado de especies espinosas de Solanum L. (Leptostemonum, Solanaceae). Biota Colomb. 2008, 9, 239–249. [Google Scholar]
- Peterson, N. Aboriginal uses of Australian Solanaceae. In The Biology and Taxonomy of the Solanaceae; Hawkes, J.G., Lester, R.N., Skelding, A.D., Eds.; Academic Press: London, UK, 1979; pp. 171–189. [Google Scholar]
- Segovia, G.V. Frutos silvestres (Solanáceas) de la Cuenca del Río Chillón, Provincia de Canta, Lima-Perú. Ecol. Aplic. 2007, 6, 23–32. [Google Scholar]
- Stern, S.R.; Tepe, E.J.; Bohs, L.A. Checklist of Solanum of North-Central Peru, a hotspot of biological diversity. Arnaldoa 2008, 15, 277–284. [Google Scholar]
- Samuels, J. A Preliminary Taxonomic Inventory of Food Species of the Solanaceae Family; Oral presentation/paper given at the Primer Congreso Internacional de Biotecnología y Biodiversidad, 28–31 May 2012; CIBE-ESPOL/AEBE: Guayaquil, Ecuador.
- Hanelt, P. Mansfield’s Encyclopedia of Agricultural and Horticultural Crops; Springer-Verlag: Berlin, Heidelberg, Germany; New York, NY, USA, 2001. [Google Scholar]
- Hutchinson, J.; Dalziel, J.M.; Keay, R.W.J.; Hepper, F.N. Flora of West Tropical Africa; Crown Agents for Overseas Governments and Administrations: London, UK, 2013. Available online: https://ia601403.us.archive.org/13/items/FloraOfWestTropi00hutc/FloraOfWestTropicalAfrica-JohnHutchinson.pdf (accessed on 18 August 2014).
- Siemonsma, J.S.; Piluek, K. (Eds.) PROSEA: Plant Resources of South-East Asia No. 8 Vegetables; Scientific Publishers: Wageningen, The Netherlands, 1993.
- Usher, G. A Dictionary of Plants Used by Man; Constable and Company Ltd: London, UK, 1974. [Google Scholar]
- Zeven, A.C.; Zhukovsky, P.M. Dictionary of Cultivated Plants and Their Centres of Diversity; PUDOC Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1975. [Google Scholar]
- Zhang, Z.; Lu, A.; D’Arcy, W. Solanaceae. In Flora of ChinaVol. 17: Verbenaceae through Solanaceae; Wu, Z., Raven, P.H., Eds.; Science Press: Beijing, China; Missouri Botanical Garden: St. Louis, MI, USA, 1994; pp. 301–304. [Google Scholar]
- Child, A.; Lester, R.N. Synopsis of the genus Solanum L. and its infrageneric taxa. In Solanaceae V: Advances in Taxonomy and Utilization; van den Berg, R.G., Barendse, G.W.M., van der Weerden, G.M., Mariani, C., Eds.; Nijmegen University Press: Nijmegen, The Netherlands, 2001; pp. 39–52. [Google Scholar]
- Weese, T.L.; Bohs, L. A three-gene phylogeny of the genus Solanum (Solanaceae). Syst. Bot. 2007, 32, 445–463. [Google Scholar] [CrossRef]
- Lester, R.N.; Jaeger, P.-M.L.; Child, A. Solanum in Africa. Available online: www.ru.nl/publish/pages/677465/rnl_internet_version.pdf (accessed on 17 November 2014).
- Bohs, L. Transfer of Cyphomandra (Solanaceae) and its species to Solanum. Taxon 1995, 44, 583–587. [Google Scholar] [CrossRef]
- D’Arcy, W.G. The Solanaceae since 1976, with a review of its biogeography. In Solanaceae III: Taxonomy, Chemistry, Evolution; Hawkes, J.G., Lester, R.N., Nee, M., Estrada, N., Eds.; Royal Botanic Gardens: Kew, UK, 1991; pp. 75–137. [Google Scholar]
- Olmstead, R.G.; Bohs, L. A summary of molecular systematic research in Solanaceae: 1982–2006. In VI Solanaceae Conference: Genomics Meets Biodiversity; Spooner, D.M., Bohs, L., Giovannoni, J., Olmstead, R.G., Shibata, D., Eds.; Acta Horticulturae (ISHS): Leuven, Belgium, 2007; Volume 745, pp. 255–268. [Google Scholar]
- International Plant Names Index. Available online: http://www.ipni.org/ipni/plantnamesearchpage.do (accessedon 15 April 2012).
- Barboza, G.E. Lectotypifications, synonymy, and a new name in Capsicum (Solanoideae, Solanaceae). PhytoKeys 2011, 2, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Arinathan, V.; Mohan, V.R.; de Britto, A.J.; Murugan, C. Wild edibles used by Palliyars of the Western Ghats, Tamil Nadu. Ind. J. Trad. Knowl. 2007, 6, 163–168. [Google Scholar]
- Guzman, F.A.; Dean, E.; Bohs, L. Hot and not so hot: Phylogenetic relationships in Capsicum and Lycianthes (Solanaceae). In Proceedings of the Unpublished poster, Botany and Mycology 2009 Conference, Snowbird, Utah, 25–29 July 2009.
- Moscone, E.A.; Scaldaferro, M.A.; Grabiele, M.; Cecchini, N.M.; Sanchez Gárcia, Y.; Jarret, R.; Daviña, J.R.; Ducasse, D.A.; Barboza, G.E.; Ehrendorfer, F. The evolution of chilli peppers (Capsicum-Solanaceae): A cytogenetic perspective. Acta Hortic. 2007, 745, 137–170. [Google Scholar]
- Pozzobon, M.T.; Schifino-Wittman, M.T.; de Bem Bianchetti, L. Chromosome numbers in wild and semi-domesticated Brazilian Capsicum L. (Solanaceae) species: Do x = 12 and x = 13 represent two evolutionary lines? Bot. J. Linn. Soc. 2006, 151, 259–269. [Google Scholar]
- Baral, J.B.; Bosland, P.W. An updated synthesis of the Capsicum genus. Capsic. Eggpl. Newsl. 2002, 21, 11–21. [Google Scholar]
- Bosland, P.W.; Baral, J.B. “Bhut Jolokia”-the world’s hottest known chile pepper is a putative naturally occurring interspecific hybrid. HortScience 2007, 42, 222–224. [Google Scholar]
- Samuels, J. The rocoto pepper: A novel crop Capsicum. Org. Grow. 2014, 28, 10–12. [Google Scholar]
- Yamamoto, S.; Djarwaningsih, T.; Wiriadinata, H. Capsicum pubescens (Solanaceae) in Indonesia: Its history, taxonomy and distribution. Econ. Bot. 2013, 20, 1–10. [Google Scholar]
- Samuels, J. Extreme weather of 2007 and its effect on tree chillies in a protected cropping system. Org. Grow. 2007, 2, 18–19. [Google Scholar]
- Andrews, J. Peppers: The Domesticated Capsicums; University of Texas Press: Austin, TX, USA, 1984. [Google Scholar]
- Barboza, G.E.; Agra, M.F.; Romero, M.V.; Scaldaferro, M.A.; Moscone, E.A. New endemic species of Capsicum (Solanaceae) from the Brazilian Caatinga: Comparison with the re-circumscribed C. parvifolium. Syst. Bot. 2011, 36, 768–781. [Google Scholar] [CrossRef]
- Williams, D.E. Lycianthes moziniana (Solanaceae): An underutilized Mexican food plant with “new” crop potential. Econ. Bot. 1993, 47, 387–400. [Google Scholar] [CrossRef]
- Narayanan, M.K.; Anilkumar, N.; Balakrishnan, V.; Sivadasan, M.; Alfarhan, H.A.; Alatar, A.A. Wild edible plants used by the Kattunaikka, Paniya and Kuruma tribes of Wayanad District, Kerala, India. J. Med. Pl. Res. 2011, 5, 3520–3529. [Google Scholar]
- Salazar, J.; Velasquez, R.; Quesada, S.; Piccinelli, A.L.; Rastrelli, L. Chemical composition and antinutritional factors of Lycianthes synanthera leaves (chomte). Food Chem. 2006, 97, 343–348. [Google Scholar] [CrossRef]
- Gentry, J.L.; D’Arcy, W.G. Solanaceae of Mesoamerica. In Solanaceae: Biology and Systematics; D’Arcy, W., Ed.; Columbia University Press: New York, NY, USA, 1986; pp. 15–26. [Google Scholar]
- Knapp, S.; Persson, V.; Blackmore, S. A phylogenetic conspectus of the tribe Juanulloeae (Solanaceae). Ann. Miss. Bot. Gard. 1997, 84, 67–89. [Google Scholar] [CrossRef]
- Rodriguez, E.F. Una nueva especie de Markea (Juanulloeae: Solanaceae) para el Perú. Arnaldoa 2006, 13, 306–313. [Google Scholar]
- Wang, Y. Chinese medicinal plant-Lycium barbarum. Sol Newsl. 2007, 14, 14. [Google Scholar]
- Minnis, P.E. Famine foods of the North American Desert Borderlands in Historical Contex. In Ethnobotany—A Reader; Minnis, P.E., Ed.; University of Oklahoma Press: Norman, France, 2000; pp. 214–239. [Google Scholar]
- Nassif, F.; Tanji, A. Gathered food plants in Morocco: The long forgotten species in ethnobotanical research. Life Sci. Leafl. 2013, 3, 17–54. [Google Scholar]
- Freedman, R. Famine Foods: Solanaceae. Available online: http://www.hort.purdue.edu/newcrop/FamineFoods/ff_families/SOLANACEAE.html (accessed on 20 April 2012).
- Dhar, P.; Tayade, A.; Ballabh, B.; Chaurasia, O.P.; Bhatt, R.P.; Srivastava, R.B. Lycium ruthenicum Murray: A less-explored but high-value medicinal plant from trans-Himalayan cold deserts of Ladakh, India. Plant Archives 2011, 11, 583–586. [Google Scholar]
- Hunziker, A.T. Estudios sobre Solanaceae XVII. Revisión sinóptica de Acnistus. Kurtziana 1982, 15, 81–102. [Google Scholar]
- Smith, S.D.; Baum, D.A. Phylogenetics of the florally diverse Andean clade Iochrominae (Solanaceae). Am. J. Bot. 2006, 93, 1140–1153. [Google Scholar] [CrossRef] [PubMed]
- Pineda, A.G. Flora Útil: Etnobotánica de Nicaragua; Gobierno de Nicaragua/Ministerio del Ambiente y los Recursos Naturales: Morena, Nicaragua, 2006. [Google Scholar]
- Martin, F.W.; Campbell, C.W.; Ruberté, R.M. Perennial Edible Fruits of the Tropics: an Inventory; USDA Agricultural Research Service Agriculture Handbook No. 642; US Government Printing Office: Washington, DC, USA, 1987.
- Shaw, J.M.H. Iochroma: A review. New Plantsman, 1998; 5, 154–191. [Google Scholar]
- Bertero, D.; Mas Serra, M.T.; Verdu, A.M.; Trillo, C. Plantas andinas y sus usos tradicionales-los recursos fitogeneticos del valle de Santa Victoria. Rev. Cienc. Hoy. 2009, 19, pp. 1–8. Available online: http://www.efn.uncor.edu/departamentos/divbioeco/divveg2/publicaciones/ (accessed on 21 April 2012).
- Smith, S.D.; Leiva, S. A new species of Iochroma (Solanaaceae) from Ecuador. Novon 2011, 21, 491–495. [Google Scholar] [CrossRef]
- Colares, M.N.; Bayón, N.D.; Stenglein, S.A.; Arambarri, A.M. Anatomía y etnobotánica de las species medicinales de la provincia pampeana: Solanaceae (excepto Grabowskia y Solanum). Acta Farm. Bonaer. 1999, 18, 171–182. [Google Scholar]
- Hunziker, A.T. Estudios sobre XIX. Sinopsis de Vassobia. Kurtziana 1984, 17, 91–118. [Google Scholar]
- Gunn, C.R.; Gaffney, F.B. Seed Characteristics of 42 Economically Important Species of Solanaceae in the United States; USDA Technical Bulletin No. 1471; Agricultural Resaerch Services, Soil Conservation Service, USDA: Washington, DC, USA, 1974. [Google Scholar]
- Martinez, M. Revision of Physalis section Epeteiorhiza (Solanaceae). An. Inst. Biol. Univ. Nac. Autón. México Ser. Bot. 1998, 69, 71–117. [Google Scholar]
- Esquivel, M.; Hammer, K. The cultivated species of the family Solanaceae in Cuba. In Solanaceae III: Taxonomy, Chemistry, Evolution; Hawkes, J.G., Lester, R.N., Nee, M., Estrada, N., Eds.; Royal Botanic Gardens: Kew, UK, 1991; pp. 357–364. [Google Scholar]
- Díaz-Betancourt, M.; Ghermandi, L.; Ladio, A.; López-Moreno, I.; Raffaele, E.; Rapoport, E.H. Weeds as a source for human consumption. A comparison between tropical and temperate Latin America. Rev. Biol. Trop. 1999, 47, 1–9. [Google Scholar]
- Williams, L.O. The Useful Plants of Central America. Ceiba 1981, 24, 3–342. [Google Scholar]
- Toledo, J.M. Physalis victoriana (Solanaceae): A new species from northern Argentina. Phytotaxa 2013, 124, 60–64. [Google Scholar] [CrossRef]
- Saunders, C.F. Edible and Useful Wild Plants of the United States and Canada; Dover Publications: Mineola, NY, USA, 1976. [Google Scholar]
- Van Valkenburg, J.L.C.H.; Bunyapraphatsara, N. (Eds.) Plant Resources of South-East Asia No. 12 (2). Medicinal and Poisonous Plants 2; PROSEA Foundation: Bogor, Indonesia, 2002; p. 782.
- Chávez, E.; Roldán, J.; Sotelo, B.E.; Ballinas, J.; López, E.J. Plantas comestibles no convencionales en Chiapas, México. Rev. Fac. Salud Publica Nutr. 2009, 10, 1–12. [Google Scholar]
- Martínez, M.A.; Evangelista, V.; Basurto, F.; Mendoza, M.; Cruz-Rivas, A. Flora útil de los cafetales en la Sierra Norte de Puebla, México. Rev. Mex. Biodiv. 2007, 78, 15–40. [Google Scholar]
- Hepper, F.N. Old World Withania (Solanaceae): A taxonomic review and key to the species. In InSolanaceae III: Taxonomy, Chemistry, Evolution; Hawkes, J.G., Lester, R.N., Nee, M., Estrada, N., Eds.; Royal Botanic Gardens: Kew, UK, 1999; pp. 211–227. [Google Scholar]
- Gupta, V.; Keshari, B.B. Withania coagulans Dunal (paneer doda): A review. Int. J. AyurvedicHerb. Med. 2013, 3, 1330–1336. [Google Scholar]
- Baez, S. Oyacachi: La Gente y La Biodiversidad; Abya-Yala: Quito, Ecuador, 2000. [Google Scholar]
- Chiarini, F.; Garboza, G.; Marticorena, A. Novedades en Solanum y Salpichroa (Solanaceae) para Sudamerica austral. Gayana Bot. 2007, 64, 46–59. [Google Scholar] [CrossRef]
- Mione, T.; Central Connecticut State University, New Britain, CT, USA. Personal communication, 2012.
- Mione, T.; Yacher, L. Jaltomata (Solanaceae) of Costa Rica. In A Festschrift for William G. D’Arcy, the Legacy of a Taxonomist; Keating, R.C., Hollowell, V.C., Croat, T.B., Eds.; Missouri Botanical Garden Press: St. Louis, MI, USA, 2005; pp. 117–130. [Google Scholar]
- Mione, T.; Bye, R. Jaltomata chihuahuensis (Solanaceae): A new combination and observations on ecology and ethnobotany. Novon 1996, 6, 78–81. [Google Scholar] [CrossRef]
- Frodin, D.G. History and concepts of big plant genera. Taxon 2004, 53, 753–776. [Google Scholar] [CrossRef]
- Symon, D.E. Kangaroo Apples: Solanum Section; Archaesolanum: Keswick, Australia, 1994. [Google Scholar]
- Nee, M. Synopsis of Solanum in the New World. In Solanaceae IV: Advances in Biology and Utilization; Nee, M., Symon, D.E., Lester, R.N., Jessop, J.P., Eds.; Royal Botanic Gardens: Kew, UK, 1999; pp. 285–333. [Google Scholar]
- Laferrière, J.E.; Weber, C.W.; Kohlhepp, E.A. Use and nutritional composition of some traditional Mountain Pima plant foods. J. Ethnobiol. 1991, 11, 93–114. [Google Scholar]
- Acosta-Quezada, P.G.; Martínez-Laborde, J.B.; Prohens, J. Variation among tree tomato (Solanum betaceum Cav.) accessions from different cultivar groups: Implications for conservation of genetic resources and breeding. Gen. Res. Crop Evol. 2011, 58, 943–960. [Google Scholar]
- Samuels, J. Incredible Crops. Smallholder 2013, June, 91. [Google Scholar]
- Acosta-Quezada, P.G.; Martínez-Laborde, J.B.; Vilanova, S.; Prohens, J. Morphological and Molecular diversity in a collection of the Andean tree tomato (Solanum betaceum Cav.). Bull. UASVM Hort. 2011, 68, 500–501. [Google Scholar]
- Stern, S.R.; Bohs, L. Two new species of Solanum from Ecuador and new combinations in Solanum section Pachyphylla (Solanaceae). J. Bot. Res. Inst. Texas 2009, 3, 503–510. [Google Scholar]
- Edmonds, J.M.; Chweya, J.A. Black Nightshades. Solanum nigrum L. and Related Species; Institute of Plant Genetics and Crop Plant Research: Gatersleben, Germany; International Plant Genetic Resources Institute: Rome, Italy, 1997. [Google Scholar]
- Vorontsova, M.V.; Knapp, S. Sections Oliganthes, Melongena and Monodolichopus. In Flora of Tropical East Africa; Beentje, H.J., Ed.; Royal Botanic Gardens: Kew, UK, 2012; pp. 77–223. [Google Scholar]
- Blanca, J.M.; Prohens, J.; Anderson, G.J.; Zuriaga, E.; Cañizares, J.; Nuez, F. AFLP and DNA sequence variation in an Andean domesticate, pepino (Solanum muricatum, Solanaceae): Implications for evolution and domestication. Am. J. Bot. 2007, 94, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Prohens, J.; Ruiz, J.J.; Nuez, F. The pepino (Solanum muricatum, Solanaceae): A “new” crop with a history. Econ. Bot. 1996, 50, 355–368. [Google Scholar] [CrossRef]
- Samuels, J. Super Fruits. Permaculture. 2013, 75, pp. 58–61. Available online: https://www.academia.edu/8142181Super_Fruits_Fruiting_Solanaceae_Species_of_Latin_America (accessed on 5 July 2013).
- Heiser, C.B.; Anderson, G. “New” Solanums. In Perspectives on New Crops and New Uses; Salick, J., Ed.; ASHS Press: Alexandria, VA, USA, 1999. [Google Scholar]
- Grandillo, S.; Chetelat, R.; Knapp, S.; Spooner, D.; Peralta, I.; Cammareri, M.; Perez, O.; Termolino, P.; Tripodi, P.; Chiusano, M.; et al. Solanum sect. Lycopersicon. In Wild Crop Relatives and Breeding Resources: Vegetables; Kole, C., Ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2011; pp. 129–215. [Google Scholar]
- Roca, W.M.; Ynouye, C.; Manrique, I.; Arbizu, C.; Gomez, R. Indigenous Andean root and tuber crops: New foods for the new millennium. Chron. Hortic. 2007, 47, 13–19. [Google Scholar]
- Hawkes, J.G. The Potato: Evolution, Biodiversity and Genetic Resources; Smithsonian Institute Press: Washington, DC, USA, 1990. [Google Scholar]
- FM Blanco. Flora de Filipinas, 1880–1883, Atlas 1, Plate 86.
- Farruggia, F.T.; Bohs, L. Two new South American species of Solanum section Crinitum (Solanaceae). Phytokeys 2010, 1, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Yetman, D. The Guarijios of the Sierra Madre: Hidden People of Northwestern Mexico; University of New Mexico Press: Albuquerqe, NM, USA; University of Arizona Southwest Center: Tucson, AZ, USA, 2002. [Google Scholar]
- Sahai, S.; Nadaf, S.M.; Nerlar, Y.S. Survey and Documentation of Wild Varieties of Crop Plants in National Park and Sanctuaries of Upper Western Ghats; Gene Campaign: New Delhi, India, 2001. [Google Scholar]
- Grubben, G.J.H.; Denton, O.A. Plant Resources of Tropical Africa 2: Vegetables; PROTA Foundation/Backhuys Publishers/CTA: Wageningen, the Netherlands, 2004. [Google Scholar]
- Centro Internacional de Agricultura Tropical. Fruits from America-Solanum. Available online: www.ciat.cgiar.org/ipgri/fruits_from_americas/frutales/species%20Solanum.htm (accessed on 7 May 2012).
- Heiser, C.B. A new domesticated variety and relationships of Solanum lasiocarpum. In Solanaceae: Biology and Systematics; D’Arcy, W.G., Ed.; Columbia University Press: New York, NY, USA, 1986; pp. 412–415. [Google Scholar]
- Volpato, G.; Marcucci, R.; Tornadore, N.; Paoletti, M.G. Domestication process of two Solanum section Lasiocarpa species among Amerindians in the Upper Orinoco, Venezuela, with special focus on Piaroa Indians. Econ. Bot. 2004, 58, 184–194. [Google Scholar] [CrossRef]
- Whalen, M.D.; Costich, D.E.; Heiser, C.B. Taxonomy of Solanum section Lasiocarpa. Gentes Herbarum 1981, 12, 4–129. [Google Scholar]
- Heiser, C.B. Ethnobotany of the naranjilla and its relatives. Econ. Bot. 1985, 39, 4–11. [Google Scholar] [CrossRef]
- Dennis, F.G.; Herner, R.C.; Camacho, S. Naranjilla: A potential cash crop for the small farmer in Latin America. Acta Hortic. ISHS 1985, 158, 475–481. [Google Scholar]
- Messinger, J.; Lauerer, M. Solanum quitoense, a new greenhouse crop for central Europe: Flowering and fruiting respond to photoperiod. Sci. Hortic. 2015, 183, 23–30. [Google Scholar] [CrossRef]
- Messinger, J.; Martini, M.M.F.; Rossi, G.; Samuels, J.; Lauerer, M. Successful pollination of the neotropical crop Solanum quitoense by Bombus terrestris: Behaviour, efficiency and yield. J. Appl. Entomol. 2015, in press. [Google Scholar]
- Heiser, C.; Soria, J.; Miller, C.; Anderson, G. A new synthetic allopolyploid naranjilla, Solanum indianense (Solanaceae). Novon 2005, 15, 290–292. [Google Scholar]
- Samuels, J. Genetically Engineered Bt Brinjal and the Implications for Plant Biodiversity-Revisited. 2012. Available online: http://www.greenpeace.org/international/Global/international/publications/agriculture/2012/GE-Bt-brinjal-revisited.pdf (accessed on 29 December 2014).
- Rai, N.; Asati, B.S.; Yadav, D.S. Conservation and genetic enhancement of underutilized vegetable crop species in North Eastern region of India. LEISA India 2004, 6, 11–12. [Google Scholar]
- Lester, R.N.; Daunay, M.-C. Diversity of African vegetable Solanum species and itsimplications for a better understanding of plant domestication. In Schriften zu Genetischen Ressourcen: Rudolf Mansfield and Plant Genetic Resources, Proceedings of a Symposium Dedicated to the 100th Birthday of Rudolf Mansfield, Gatersleben, Germany, 8–9 October 2001; Knupffer, H., Ochsmann, J., Eds.; ZADI: Bonn, Germany, 2003; pp. 137–152. [Google Scholar]
- Goode, P.M. Edible Plants of Uganda: The Value of Wild and Cultivated Plants as Food; FAO: Rome, Italy, 1989. [Google Scholar]
- Symon, D.E. A revision of the genus Solanum in Australia. J. Adelaide Bot. Gard. 1981, 4, 1–367. [Google Scholar]
- Martine, C.T.; Vanderpool, D.; Anderson, G.J.; Les, D.H. Phylogenetic relationships of andromonoecious and dioecious Australian species of Solanum subgenus Leptostemonum section Melongena: Inferences from ITS sequence data. Syst. Bot. 2006, 31, 410–420. [Google Scholar] [CrossRef]
- Jaeger, P.-M.L. Systematic Studies in the Genus Solanum in Africa. Ph.D. Thesis, University of Birmingham, Birmingham, UK, 1985. [Google Scholar]
- Schmelzer, G.H.; Gurib-Fakim, A. Plant Resources of Tropical Africa 11 (1): Medicinal Plants 1; PROTA Foundation/Backhuys Publishers/CTA: Wageningen, the Netherlands, 2008. [Google Scholar]
- Lester, R.N.; Niakan, L. Origin and domestication of the scarlet eggplant, Solanum aethiopicum, from S. anguivi in Africa. In Solanaceae: Biology and Systematics; D’Arcy, W.G., Ed.; Columbia University Press: New York, NY, USA, 1986; pp. 433–456. [Google Scholar]
- Mao, A.A.; Hynniewta, T.M.; Sanjappa, M. Plant wealth of northeast India with reference to ethnobotany. Ind. J. Trad. Knowl. 2009, 8, 96–103. [Google Scholar]
- Srivastava, R.C. Traditional knowledge of Nyishi (Daffla) tribe of Arunachal Pradesh. Ind. J. Trad. Knowl. 2010, 9, 26–37. [Google Scholar]
- Hegarty, M.P.; Hegarty, E.E.; Wills, R.B.H. Food Safety of Australian Plant Bushfoods; RIRDC Publication No. 01/28; Rural Industries Research and Development Corporation: Kingston, ACT, Australia, 2001. [Google Scholar]
- Curtis’s Botanical Magazine. Series 3, Volume 20, Plate 5424, 1864.
- Sawian, J.T.; Jeeva, S.; Lyndem, F.G.; Mishra, B.P.; Laloo, R.C. Wild edible plants of Meghalaya, North-East India. Nat. Product Radiance 2007, 6, 410–426. [Google Scholar]
- De Agra, M.F.; Bhattacharyya, J. Ethnomedicinal and Phytochemical investigation of the Solanum species in the northeast of Brazil. In Solanaceae IV: Advances in Biology and Utilization; Nee, M., Symon, D.E., Lester, R.N., Jessop, J.P., Eds.; Royal Botanic Gardens: Kew, UK, 1999; pp. 341–343. [Google Scholar]
- Barboza, G.E.; de Bem Bianchetti, L. Three new species of Capsicum (Solanaceae) and a key to the wild species from Brazil. Syst. Bot. 2005, 30, 863–871. [Google Scholar] [CrossRef]
- Mione, T.; Segundo Leiva, G.; Yacher, L. Five new species of Jaltomata (Solanaceae) from Cajamarca, Peru. Novon 2007, 17, 49–58. [Google Scholar] [CrossRef]
- Stern, S.; Bohs, L. Two new species of Solanum (Solanaceae) from the Amotape-Huancabamba Zone of southern Ecuador and northern Peru. PhytoKeys 2010, 1, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Khoshbakht, K.; Hammer, K. How many plant species are cultivated? Genet. Resour. Crop. Ev. 2008, 55, 925–928. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).