Construction of a Nanosensor for Non-Invasive Imaging of Hydrogen Peroxide Levels in Living Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals, Vectros and Strains
2.2. Plasmid Construction
2.3. Protein Expression and Purification
2.4. In Vitro Characterization and Ligand Binding Affinity of Nanosensor Protein
2.5. In Vivo Characterization in Bacterial Cells
2.6. H2O2 Dynamics in Yeast under Normal and Stress Conditions
2.7. Monitoring of H2O2 Changes in Mammalian Cells
3. Results and Discussion
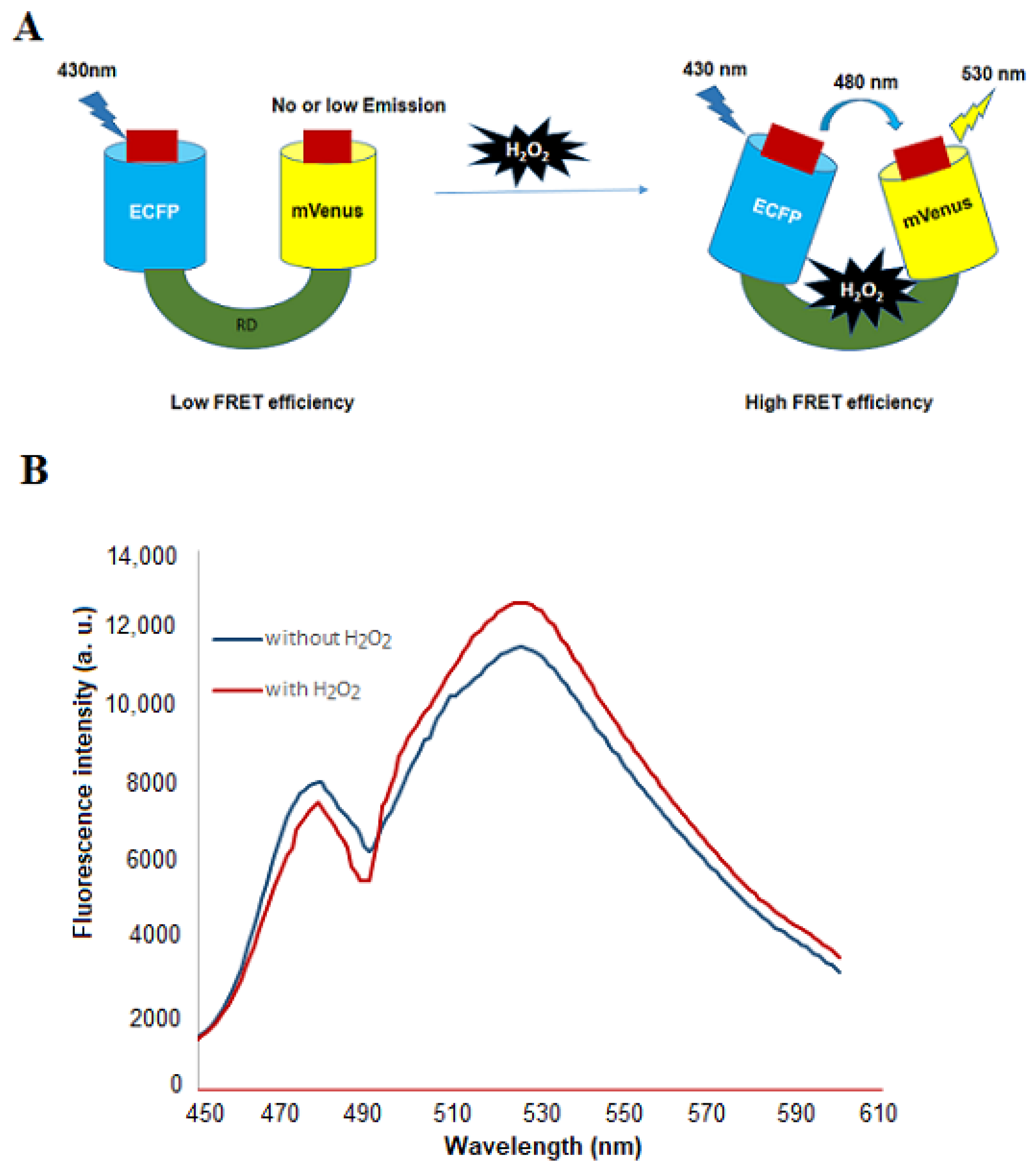
3.1. Designing, Construction and Spectral Analysis of FLIP-H2O2
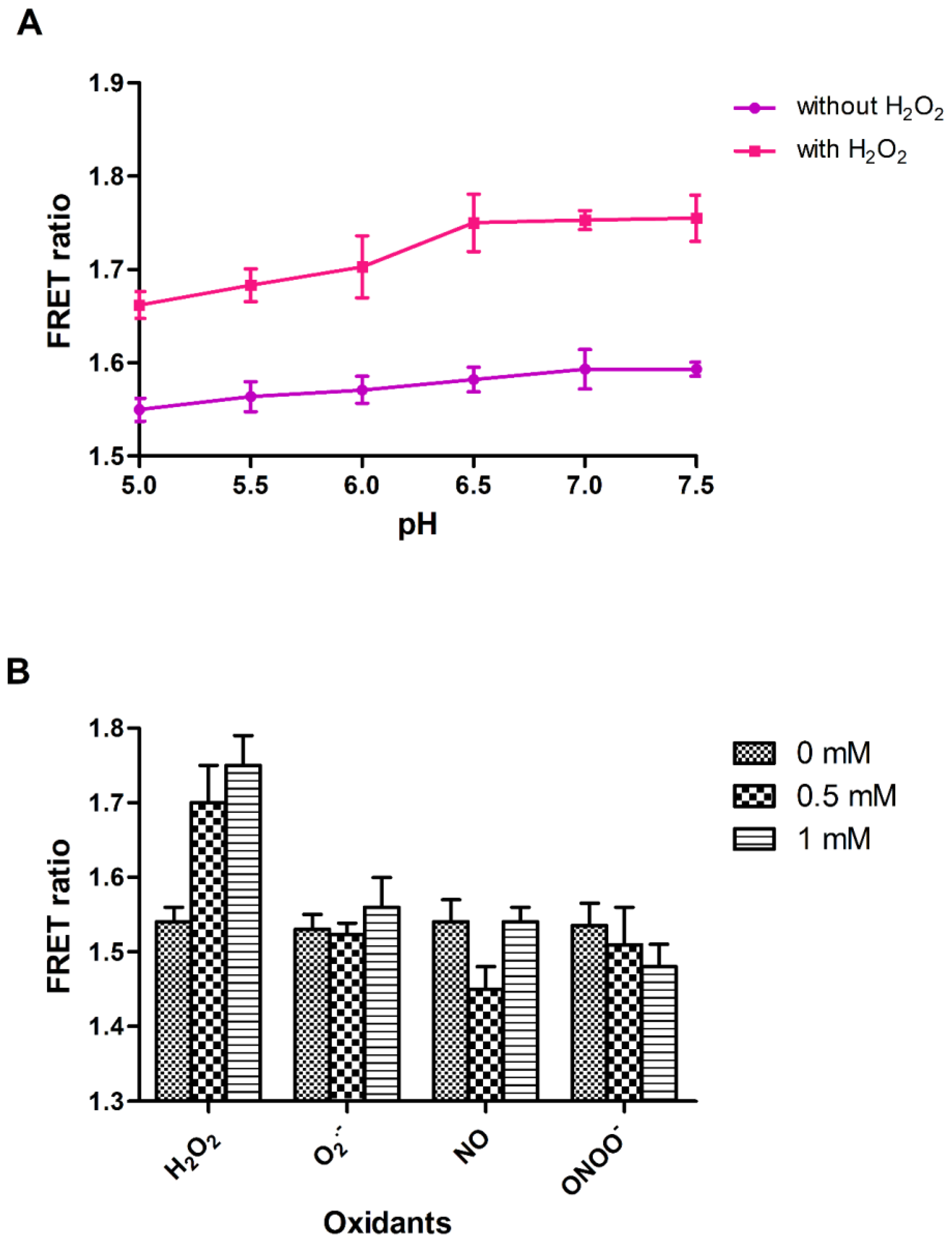
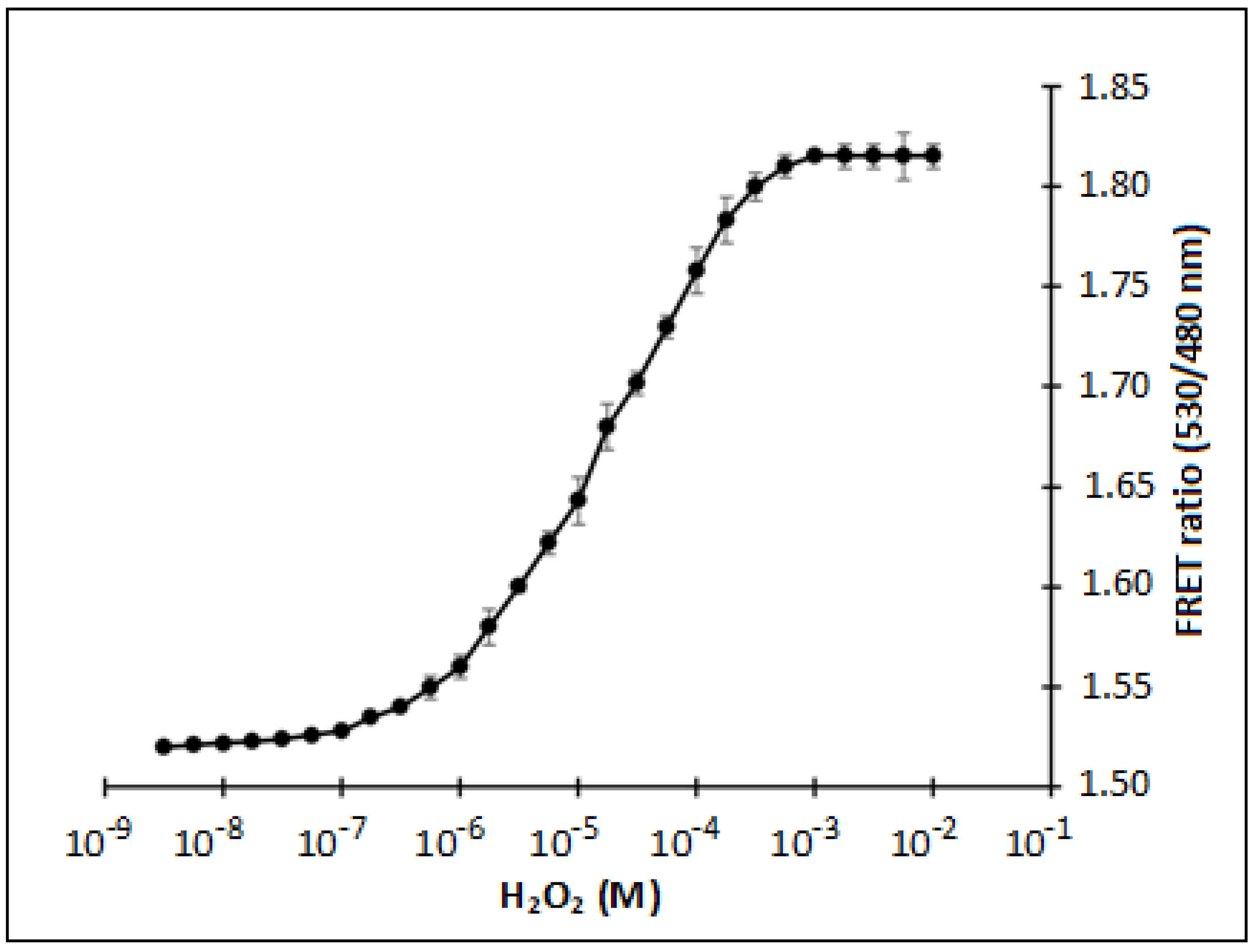
3.2. In Vitro Characterization of FLIP-H2O2 Nanosensor
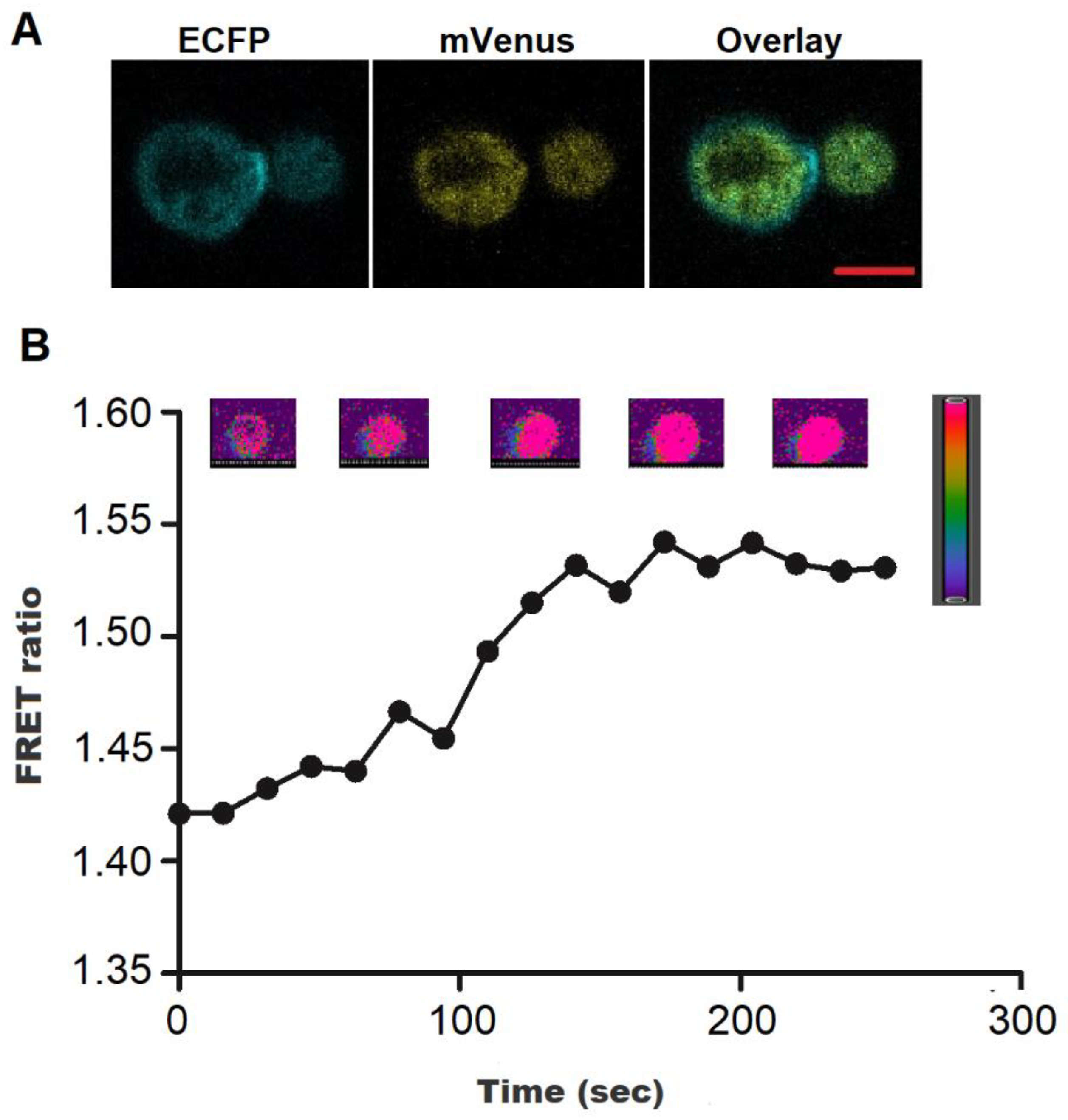
3.3. Intracellular H2O2 Flux Monitoring in Bacterial Cells
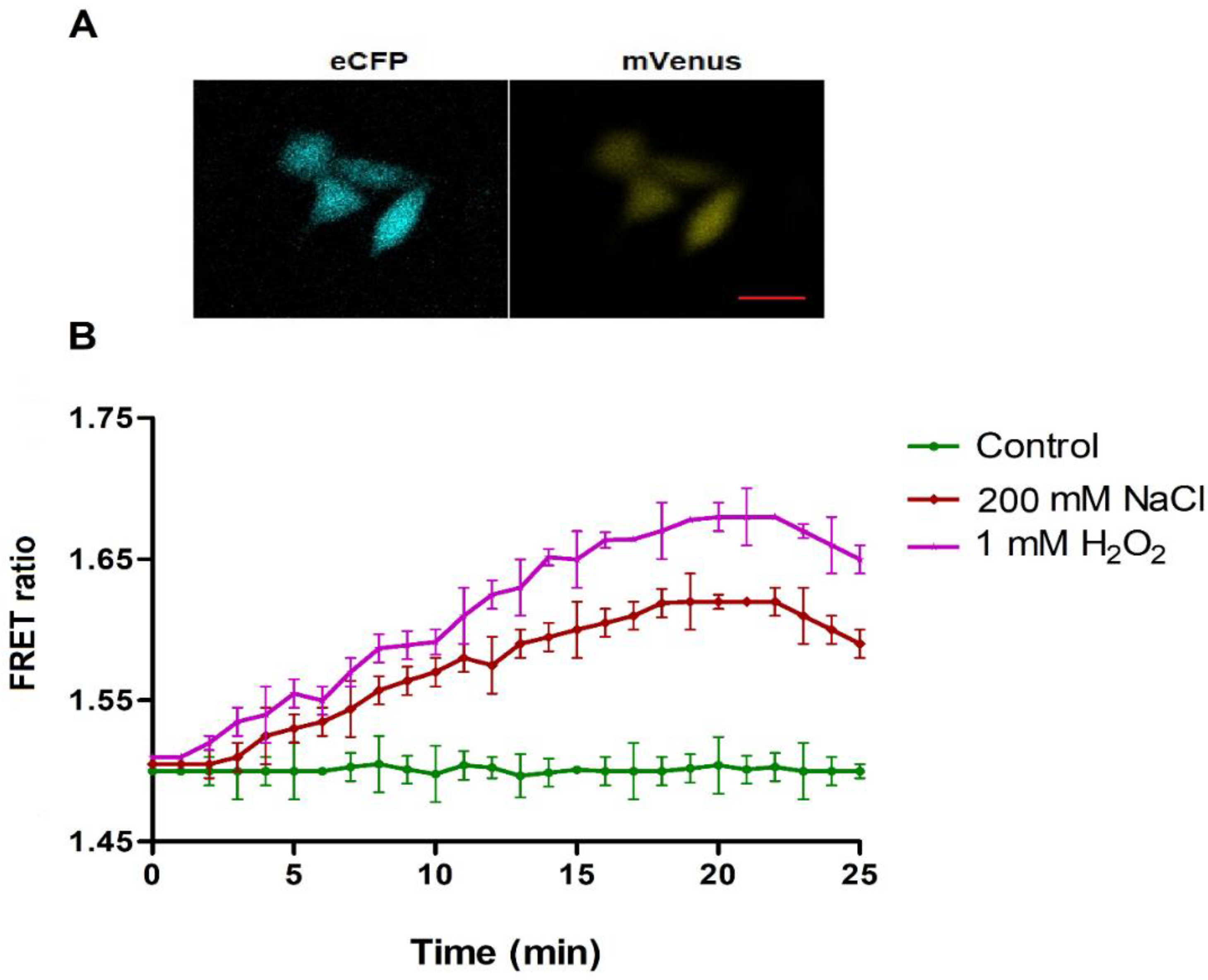
3.4. Monitoring of H2O2 in Yeast and Mammalian Cells
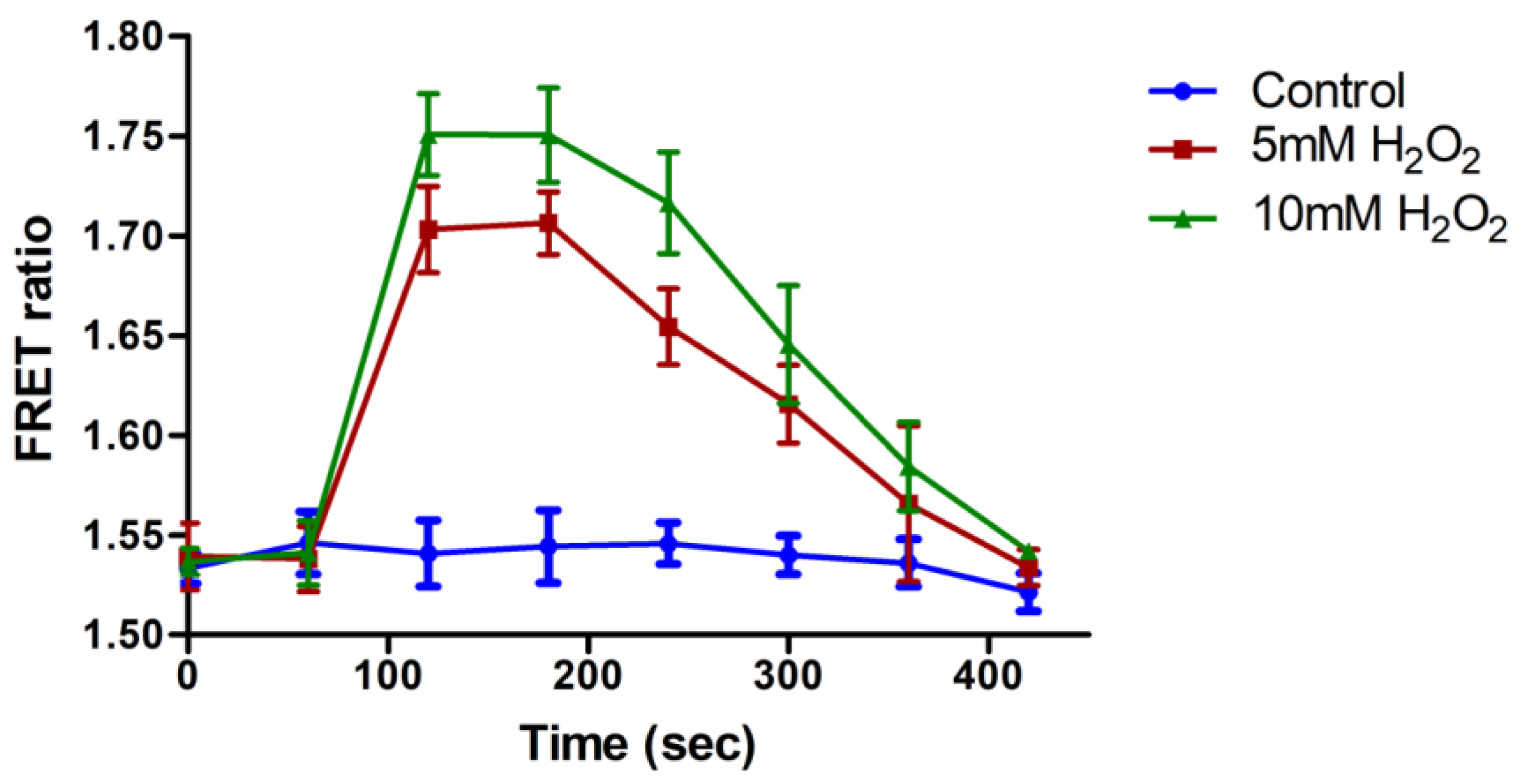
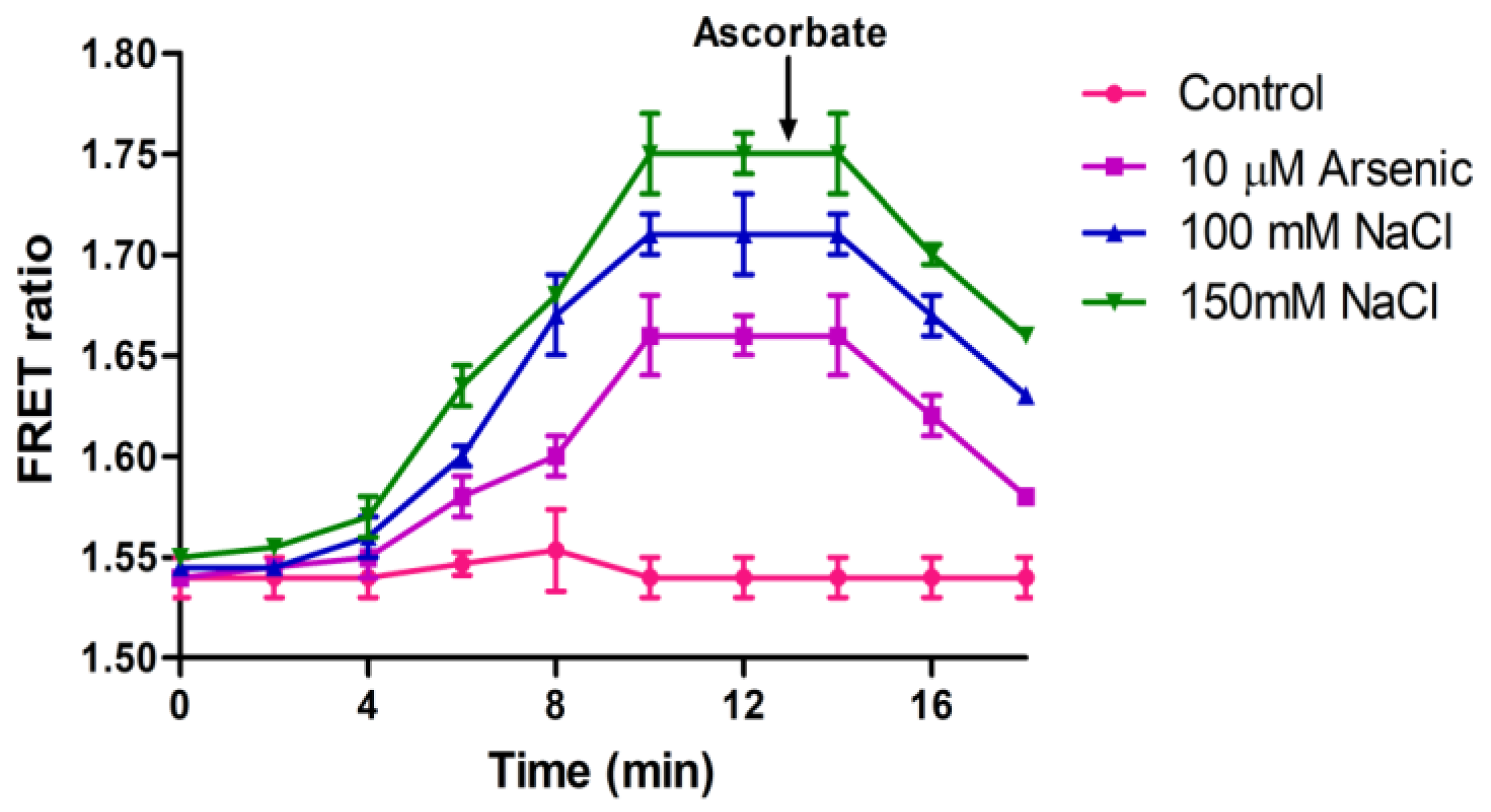
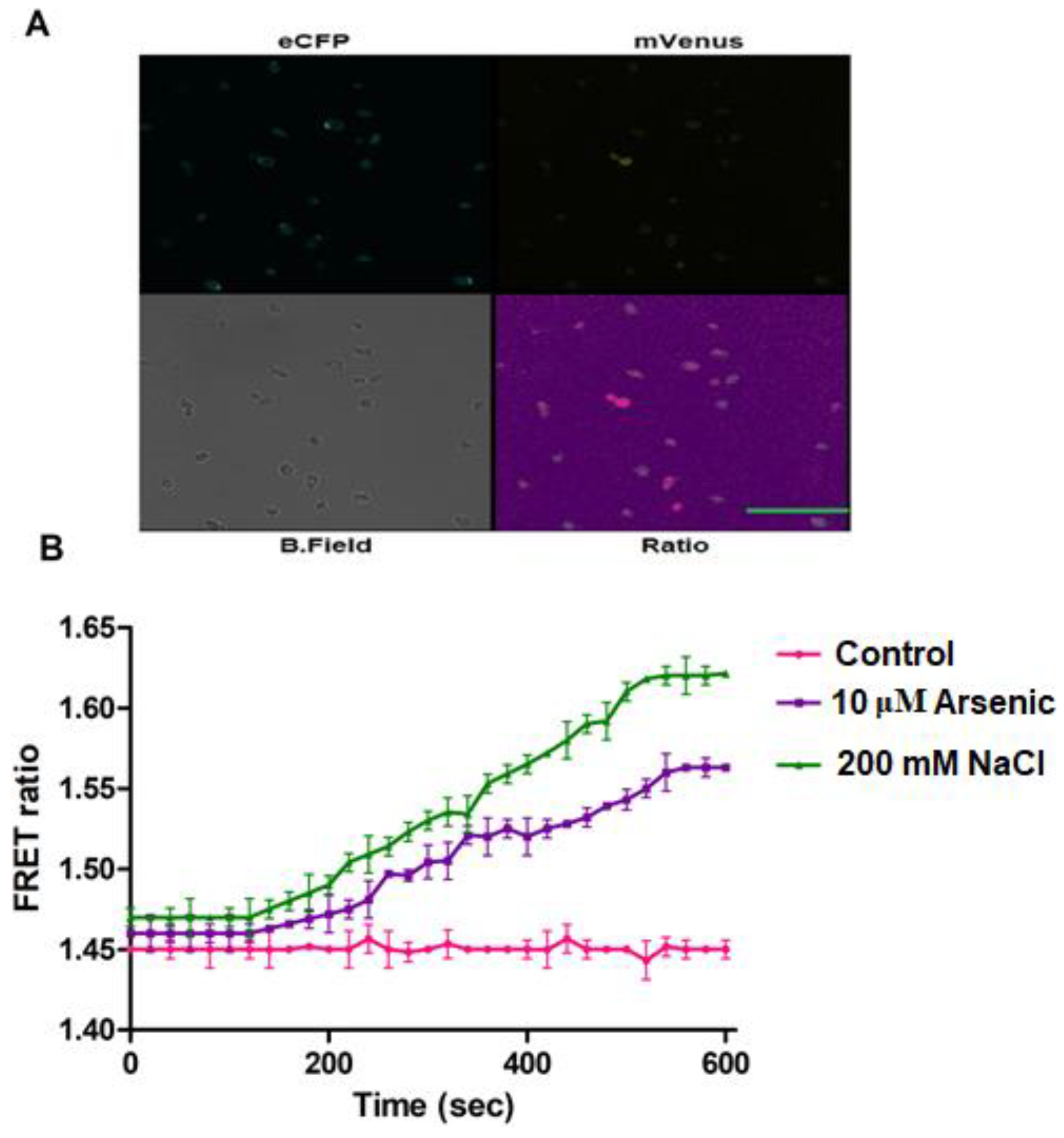
3.5. In Vivo H2O2 Flux Monitoring under Stress Conditions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- D’Autréaux, B.; Toledano, M.B. ROS as signalling molecules: Mechanisms that generate specificity in ROS homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef]
- Halliwell, B. Free radicals and antioxidants–quo vadis? Trends Pharm. Sci. 2011, 32, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Zentgraf, U. Oxidative stress and leaf senescence. Annu. Plant Rev. Senescence Process. Plants 2007, 26, 69–86. [Google Scholar]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef]
- Costa, A.; Drago, I.; Behera, S.; Zottini, M.; Pizzo, P.; Schroeder, J.I.; Pozzan, T.; Schiavo, F.L. H2O2 in plant peroxisomes: An in vivo analysis uncovers a Ca2+-dependent scavenging system. Plant J. 2010, 62, 760–772. [Google Scholar] [CrossRef]
- Doke, N. Generation of superoxide anion by potato tuber protoplasts during the hypersensitive response to hyphal wall components of Phytophthora infestans and specific inhibition of the reaction by suppressors of hypersensitivity. Physiol. Plant Pathol. 1983, 23, 359–367. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley—Powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Dikalov, S.I.; Harrison, D.G. Methods for detection of mitochondrial and cellular reactive oxygen species. Antioxid. Redox Signal. 2014, 20, 372–382. [Google Scholar] [CrossRef]
- Miller, E.W.; Albers, A.E.; Pralle, A.; Isacoff, E.Y.; Chang, C.J. Boronate-based fluorescent probes for imaging cellular hydrogen peroxide. J. Am. Chem. Soc. 2005, 127, 16652–16659. [Google Scholar] [CrossRef]
- Dickinson, B.C.; Huynh, C.; Chang, C.J. A palette of fluorescent probes with varying emission colors for imaging hydrogen peroxide signaling in living cells. J. Am. Chem. Soc. 2010, 132, 5906–5915. [Google Scholar] [CrossRef] [PubMed]
- Albers, A.E.; Okreglak, V.S.; Chang, C.J. A FRET-based approach to ratiometric fluorescence detection of hydrogen peroxide. J. Am. Chem. Soc. 2006, 128, 9640–9641. [Google Scholar] [CrossRef] [PubMed]
- Nietzel, T.; Elsässer, M.; Ruberti, C.; Steinbeck, J.; Ugalde, J.M.; Fuchs, P.; Wagner, S.; Ostermann, L.; Moseler, A.; Lemke, P.; et al. The fluorescent protein sensor ro GFP 2-Orp1 monitors in vivo H2O2 and thiol redox integration and elucidates intracellular H2O2 dynamics during elicitor-induced oxidative burst in Arabidopsis. New Phytol. 2019, 221, 1649–1664. [Google Scholar] [CrossRef] [PubMed]
- Hanson, G.T.; Aggeler, R.; Oglesbee, D.; Cannon, M.; Capaldi, R.A.; Tsien, R.Y.; Remington, S.J. Investigating mitochondrial redox potential with redox-sensitive green fluorescent protein indicators. J. Biol. Chem. 2004, 279, 13044–13053. [Google Scholar] [CrossRef]
- Belousov, V.V.; Fradkov, A.F.; Lukyanov, K.A.; Staroverov, D.B.; Shakhbazov, K.S.; Terskikh, A.V.; Lukyanov, S. Genetically encoded fluorescent indicator for intracellular hydrogen peroxide. Nat. Methods 2006, 3, 281–286. [Google Scholar] [CrossRef]
- Okumoto, S.; Jones, A.; Frommer, W.B. Quantitative imaging with fluorescent biosensors. Annu. Rev. Plant Biol. 2012, 63, 663–706. [Google Scholar] [CrossRef]
- Hochreiter, B.; Pardo-Garcia, A.; Schmid, J.A. Fluorescent proteins as genetically encoded FRET biosensors in life sciences. Sensors 2015, 15, 26281–26314. [Google Scholar] [CrossRef]
- Vinkenborg, J.L.; Nicolson, T.J.; Bellomo, E.A.; Koay, M.S.; Rutter, G.A.; Merkx, M. Genetically encoded FRET sensors to monitor intracellular Zn 2+ homeostasis. Nat. Methods 2009, 6, 737. [Google Scholar] [CrossRef]
- Mohsin, M.; Ahmad, A. Genetically-encoded nanosensor for quantitative monitoring of methionine in bacterial and yeast cells. Biosens. Bioelectron. 2014, 59, 358–364. [Google Scholar] [CrossRef]
- Ballerstadt, R.; Gowda, A.; McNichols, R. Fluorescence resonance energy transfer-based near-infrared fluorescence sensor for glucose monitoring. Diabetes Technol. Ther. 2004, 6, 191–200. [Google Scholar] [CrossRef]
- Waadt, R.; Hitomi, K.; Nishimura, N.; Hitomi, C.; Adams, S.R.; Getzoff, E.D.; Schroeder, J.I. FRET-based reporters for the direct visualization of abscisic acid concentration changes and distribution in Arabidopsis. Elife 2014, 3, e01739. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Mohsin, M.; Iqrar, S.; Manzoor, O.; Siddiqi, T.O.; Ahmad, A. Live cell imaging of vitamin B12 dynamics by genetically encoded fluorescent nanosensor. Sens. Actuator B Chem. 2018, 257, 866–874. [Google Scholar] [CrossRef]
- Zheng, M.; Åslund, F.; Storz, G. Activation of the OxyR transcription factor by reversible disulfide bond formation. Science 1998, 279, 1718–1722. [Google Scholar] [CrossRef] [PubMed]
- Fehr, M.; Frommer, W.B.; Lalonde, S. Visualization of maltose uptake in living yeast cells by fluorescent nanosensors. Proc. Natl. Acad. Sci. USA 2002, 99, 9846–9851. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Anjum, N.A.; Asif, A.; Ahmad, A. Real-time monitoring of glutathione in living cells using genetically encoded FRET-based ratiometric nanosensor. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Ahmad, M.; Ameen, S.; Siddiqi, T.O.; Khan, P.; Ahmad, A. Live cell monitoring of glycine betaine by FRET-based genetically encoded nanosensor. Biosens. Bioelectron. 2016, 86, 169–175. [Google Scholar] [CrossRef]
- Yu, D.; Wang, P.; Zhao, Y.; Fan, A. Iodophenol blue-enhanced luminol chemiluminescence and its application to hydrogen peroxide and glucose detection. Talanta 2016, 146, 655–661. [Google Scholar] [CrossRef]
- Gutscher, M.; Sobotta, M.C.; Wabnitz, G.H.; Ballikaya, S.; Meyer, A.J.; Samstag, Y.; Dick, T.P. Proximity-based protein thiol oxidation by H2O2-scavenging peroxidases. J. Biol. Chem. 2009, 284, 31532–31540. [Google Scholar] [CrossRef]
- Bischof, H.; Rehberg, M.; Stryeck, S.; Artinger, K.; Eroglu, E.; Waldeck-Weiermair, M.; Gottschalk, B.; Rost, R.; Deak, A.T.; Niedrist, T.; et al. Novel genetically encoded fluorescent probes enable real-time detection of potassium in vitro and in vivo. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef]
- Zhang, C.; Ye, B.C. A single fluorescent protein-based sensor for in vivo 2-oxogluatarate detection in cell. Biosens. Bioelectron. 2014, 54, 15–19. [Google Scholar] [CrossRef]
- Ambrin, G.; Ahmad, M.; Alqarawi, A.A.; Hashem, A.; Abd_Allah, E.F.; Ahmad, A. Conversion of cytochrome P450 2D6 of human into a FRET-based tool for real-time monitoring of Ajmalicine in living cells. Front. Bioeng. Biotechnol. 2019, 7, 375. [Google Scholar] [CrossRef] [PubMed]
- Ying, W.Z.; Aaron, K.; Sanders, P.W. Mechanism of dietary salt-mediated increase in intravascular production of TGF-β1. Am. J. Physiol. Renal Physiol. 2008, 295, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Thannickal, V.J.; Hassoun, P.M.; White, A.C.; Fanburg, B.L. Enhanced rate of H2O2 release from bovine pulmonary artery endothelial cells induced by TGF-beta 1. Am. J. Physiol. Lung Cell Mol. Physiol. 1993, 265, 622–626. [Google Scholar] [CrossRef] [PubMed]
- Nisimoto, Y.; Diebold, B.A.; Cosentino-Gomes, D.; Lambeth, J.D. Nox4: A hydrogen peroxide-generating oxygen sensor. Biochemistry 2014, 53, 5111–5120. [Google Scholar] [CrossRef]
- Wang, T.S.; Kuo, C.F.; Jan, K.Y.; Huang, H. Arsenite induces apoptosis in chinese hamster ovary cells by generation of reactive oxygen species. J. Cell. Physiol. 1996, 169, 256–268. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amreen; Ali, H.M.; Ahmad, M.; Salem, M.Z.M.; Ahmad, A. Construction of a Nanosensor for Non-Invasive Imaging of Hydrogen Peroxide Levels in Living Cells. Biology 2020, 9, 430. https://doi.org/10.3390/biology9120430
Amreen, Ali HM, Ahmad M, Salem MZM, Ahmad A. Construction of a Nanosensor for Non-Invasive Imaging of Hydrogen Peroxide Levels in Living Cells. Biology. 2020; 9(12):430. https://doi.org/10.3390/biology9120430
Chicago/Turabian StyleAmreen, Hayssam M. Ali, Mohammad Ahmad, Mohamed Z. M. Salem, and Altaf Ahmad. 2020. "Construction of a Nanosensor for Non-Invasive Imaging of Hydrogen Peroxide Levels in Living Cells" Biology 9, no. 12: 430. https://doi.org/10.3390/biology9120430
APA StyleAmreen, Ali, H. M., Ahmad, M., Salem, M. Z. M., & Ahmad, A. (2020). Construction of a Nanosensor for Non-Invasive Imaging of Hydrogen Peroxide Levels in Living Cells. Biology, 9(12), 430. https://doi.org/10.3390/biology9120430