The Human Respiratory System and its Microbiome at a Glimpse

 ,
,  ,
,  ,
, 
Simple Summary
Abstract
1. Introduction
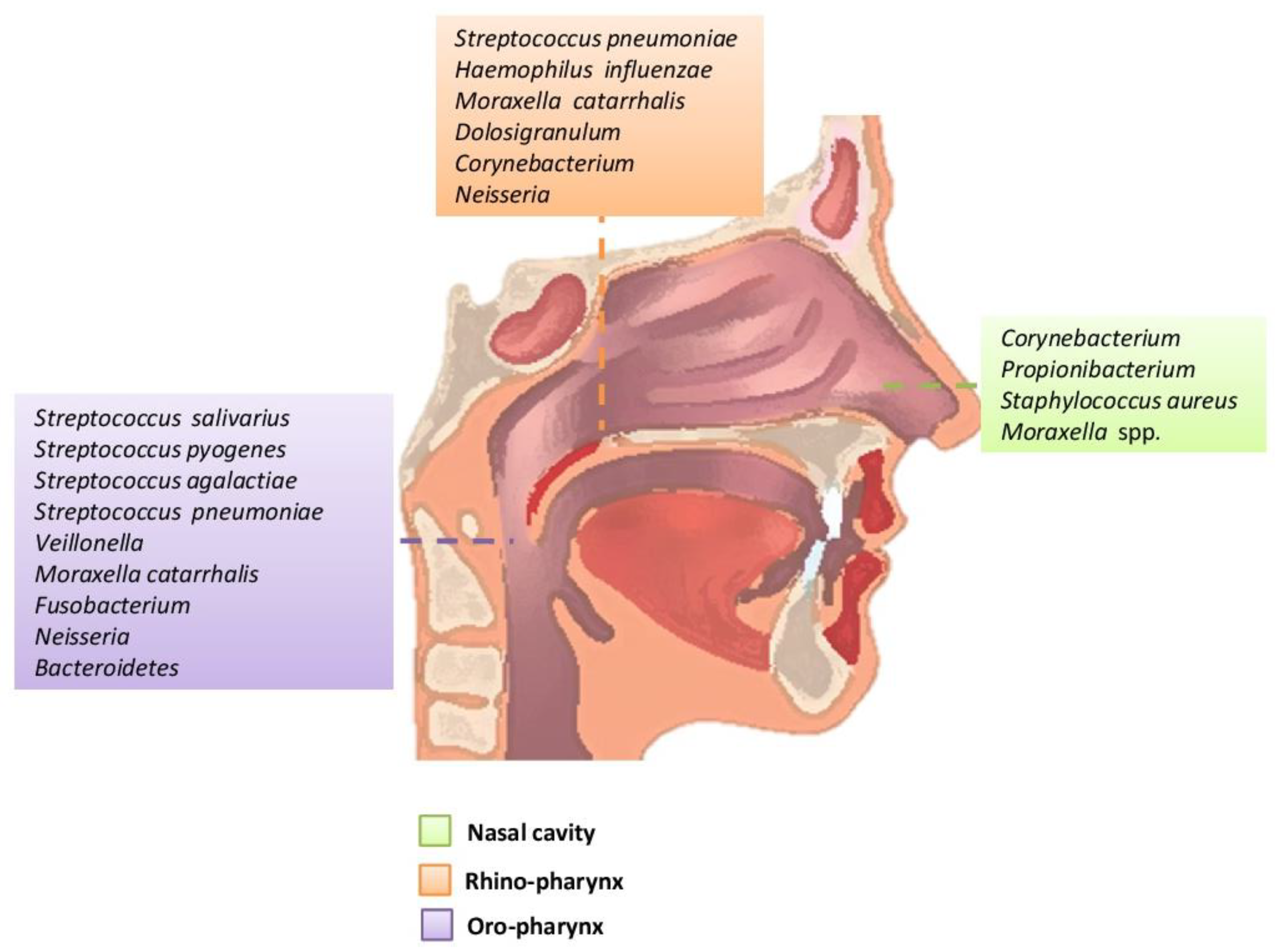
2. The Upper Respiratory Tract Microbiome (Airways Colonization and Evolution during Life)
2.1. Nostrils
2.2. Rhinopharynx
2.3. Oropharynx
3. The Lower Respiratory Tract Microbiome (Airway Colonization and Evolution during Life)
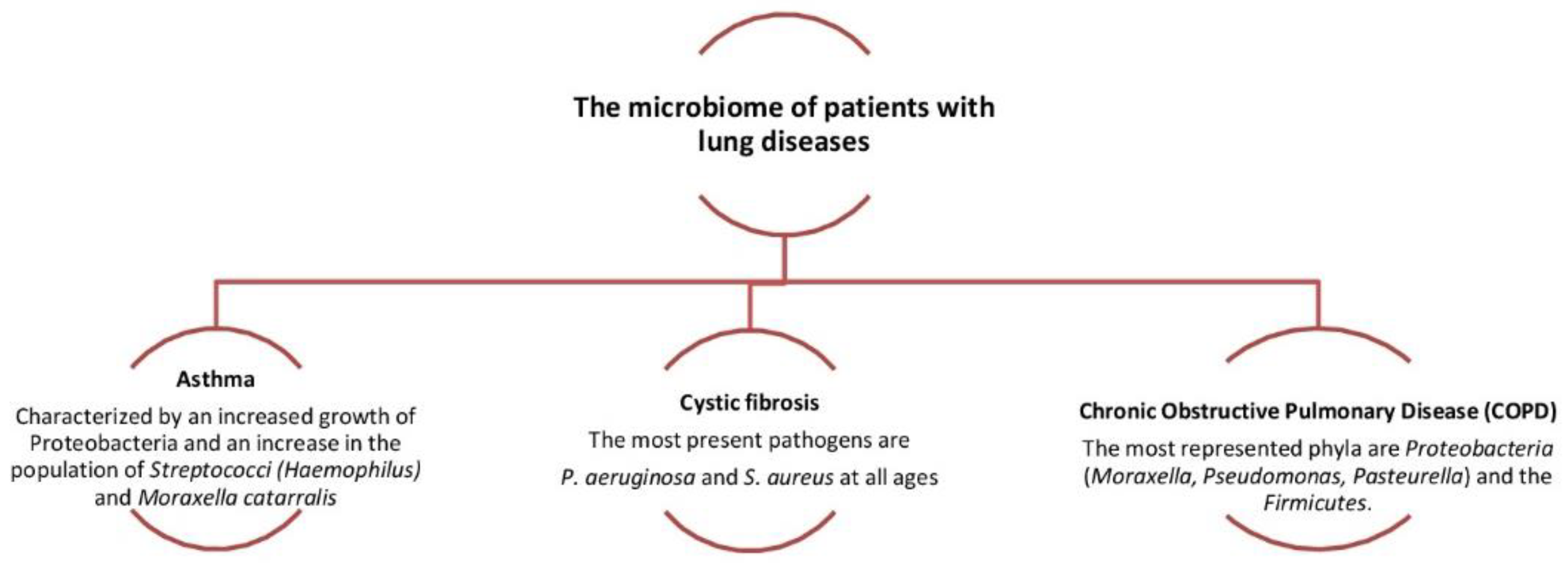
4. Pathogenesis of Respiratory Disorders
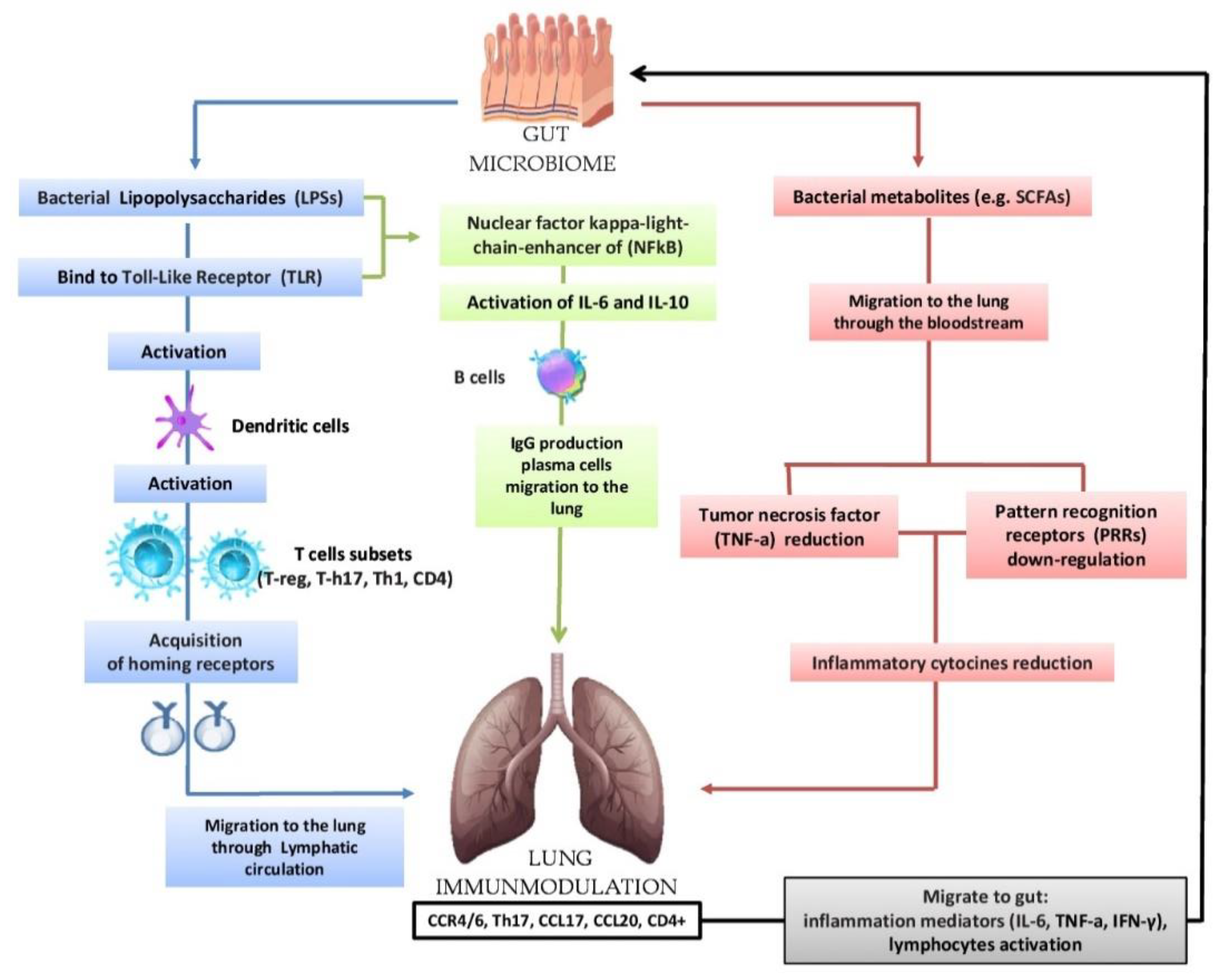
5. The Gut-Lung Axis
5.1. Biomolecular Mechanisms
5.2. Effects of Environmental Factors Influencing the Eubiosis of the Airway’s Microbiota
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Finlay, B.B.; Finlay, J.M. The Whole-Body Microbiome: How to Harness Microbes―Inside and Out―for Lifelong Health Hardcover, 1st ed.; The Experiment: New York, NY, USA, 2019; pp. 238–260. [Google Scholar]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; FitzGerald, M.G.; Fulton, R.S.; et al. The Human Microbiome Project Consortium. Structure function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar]
- Sahin-Yilmaz, A.; Naclerio, R.M. Anatomy and physiology of the upper airway. Proc. Am. Thorac. Soc. 2011, 8, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Rogers, G.B.; Shaw, D.; Marsh, R.L.; Carroll, M.P.; Serisier, D.J.; Bruce, K.D. Respiratory microbiota: Addressing clinical questions, informing clinical practice. Thorax 2015, 70, 74–81. [Google Scholar] [CrossRef]
- Mitchell, A.B.; Oliver, B.G.; Glanville, A.R. Translational aspects of the human respiratory virome. Am. J. Respir. Crit. Care Med. 2016, 194, 1458–1464. [Google Scholar] [CrossRef]
- Campanella, V.; Syed, J.; Santacroce, L.; Saini, R.; Ballini, A.; Inchingolo, F. Oral probiotics influence oral and respiratory tract infections in pediatric population: A randomized double-blinded placebo-controlled pilot study. Eur Rev Med Pharmacol Sci. 2018, 22, 8034–8041. [Google Scholar]
- Biesbroek, G.; Tsivtsivadze, E.; Sanders, E.A.; Montijn, R.; Veenhoven, R.H.; Keijser, B.J.; Bogaert, D. Early respiratory microbiota composition determines bacterial succession patterns and respiratory health in children. Am. J. Respir. Crit. Care Med. 2014, 190, 1283–1292. [Google Scholar] [CrossRef]
- Huffnagle, G.; Dickson, R.; Lukacs, N. The respiratory tract microbiome and lung inflammation: A two-way street. Mucosal Immunol. 2017, 10, 299–306. [Google Scholar] [CrossRef]
- Kumpitsch, C.; Koskinen, K.; Schöpf, V.; Moissl-Eichinger, C. The microbiome of the upper respiratory tract in health and disease. BMC Biol. 2019, 17, 87. [Google Scholar] [CrossRef]
- Schenck, L.P.; Surette, M.G.; Bowdish, D.M.E. Composition and immunological significance of the upper respiratory tract microbiota. FEBS Lett. 2016, 21, 3705–3720. [Google Scholar] [CrossRef]
- Ranucci, G.; Buccigrossi, V.; de Freitas, M.B.; Guarino, A.; Giannattasio, A. Early-Life Intestine Microbiota and Lung Health in Children. J. Immunol. Res. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, S.; Wang, N.; Tan, H.Y.; Zhang, Z.; Feng, Y. The Cross-Talk between Gut Microbiota and Lungs in Common Lung Diseases. Front. Microbiol. 2020, 25, 301. [Google Scholar] [CrossRef] [PubMed]
- Gallacher, D.J.; Kotecha, S. Respiratory Microbiome of New-Born Infants. Front. Pediatr. 2016, 23, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Cicinelli, E.; Ballini, A.; Marinaccio, M.; Poliseno, A.; Coscia, M.F.; Monno, R.; De Vito, D. Microbiological findings in endometrial specimen: Our experience. Arch. Gynecol. Obstet. 2012, 285, 1325–1329. [Google Scholar] [CrossRef]
- DiGiulio, D.B. Diversity of microbes in amniotic fluid. Semin. Fetal Neonatal Med. 2012, 17, 2–11. [Google Scholar] [CrossRef]
- Krzych-Fałta, E.; Furmańczyk, K.; Lisiecka-Biełanowicz, M.; Sybilski, A.; Tomaszewska, A.; Raciborski, F.; Wojas, O.; Walkiewicz, A.; Samel-Kowalik, P.; Samoliński, B. The effect of selected risk factors, including the mode of delivery, on the development of allergic rhinitis and bronchial asthma. Adv. Dermatol. Allergol. 2018, 35, 267–273. [Google Scholar] [CrossRef]
- David, W.; Cleary, S.C. Clarke the nasopharyngeal microbiome Emerging Topics in Life. Sciences 2017, 1, 297–312. [Google Scholar]
- Bosch, A.A.T.M.; Levin, E.; van Houten, M.A.; Hasrat, R.; Kalkman, G.; Biesbroek, G.; de Steenhuijsen Piters, W.A.A.; de Groot, P.C.M.; Pernet, P.; Keijser, B.J.F.; et al. Development of Upper Respiratory Tract Microbiota in Infancy is Affected by Mode of Delivery. EBioMedicine 2016, 9, 336–345. [Google Scholar] [CrossRef]
- Ballini, A.; Cantore, S.; Farronato, D.; Cirulli, N.; Inchingolo, F.; Papa, F.; Malcangi, G.; Inchingolo, A.D.; Dipalma, G.; Sardaro, N.; et al. Periodontal disease and bone pathogenesis: The crosstalk between cytokines and porphyromonas gingivalis. J. Biol. Regul. Homeost. Agents 2015, 29, 273–281. [Google Scholar]
- Gnoni, A.; De Nitto, E.; Scacco, S.; Santacroce, L.; Palese, L.L. A New Look at the Structures of Old Sepsis Actors by Exploratory Data Analysis Tools. Antibiotics 2019, 8, 225. [Google Scholar] [CrossRef]
- Di Serio, F.; Lovero, R.; D’Agostino, D.; Nisi, L.; Miragliotta, G.; Contino, R.; Man, A.; Ciccone, M.M.; Santacroce, L. Evaluation of procalcitonin, Vitamin D and C-reactive protein levels in septic patients with positive emocoltures. Our preliminary experience. Acta Med. Mediterr. 2016, 32, 1911–1914. [Google Scholar]
- Ballini, A.; Dipalma, G.; Isacco, C.G.; Boccellino, M.; Di Domenico, M.; Santacroce, L.; Nguyễn, K.C.D.; Scacco, S.; Calvani, M.; Boddi, A.; et al. Oral Microbiota and Immune System Crosstalk: A Translational Research. Biology 2020, 9, 131. [Google Scholar] [CrossRef] [PubMed]
- Pulvirenti, G.; Parisi, G.F.; Giallongo, A.; Papale, M.; Manti, S.; Savasta, S.; Licari, A.; Marseglia, G.L.; Leonardi, S. Lower Airway Microbiota. Front. Pediatr. 2019, 7, 393. [Google Scholar] [CrossRef] [PubMed]
- Dickson, R.P.; Erb-Downward, J.R.; Freeman, C.M.; McCloskey, L.; Falkowski, N.R.; Huffnagle, G.B.; Curtis, J.L. Bacterial topography of the healthy human lower respiratory tract. MBio 2017, 8, e02287-16. [Google Scholar] [CrossRef]
- West, J.B. Regional differences in the lung. Chest 1978, 74, 426–437. [Google Scholar]
- Dickson, R.P.; Erb-Downward, J.R.; Freeman, C.M.; McCloskey, L.; Beck, J.M.; Huffnagle, G.B.; Curtis, J.L. Spatial Variation in the Healthy Human Lung Microbiome and the Adapted Island Model of Lung Biogeography. Ann. Am. Thorac. Soc. 2015, 12, 821–830. [Google Scholar] [CrossRef]
- Basis, C.M.; Tang, A.L.; Young, V.B.; Pynnonen, M.A. The nasal cavity microbiota of healthy adults. Microbiome 2014, 2, 1–5. [Google Scholar] [CrossRef]
- Huang, Y.J.; Boushey, H.A. The microbiome and asthma. Ann. Am. Thorac. Soc. 2014, 11, S48–S51. [Google Scholar] [CrossRef]
- Wouter, A.A.; de Steenhuijsen, P.; Huijskens, E.G.W.; Wyllie, A.L.; Biesbroek, G.; van den Bergh, M.R.; Veenhoven, R.H.; Wang, X.; Trzciński, K.; Bonten, M.J.; et al. Dysbiosis of upper respiratory tract microbiota in elderly pneumonia patients. ISME J. 2016, 10, 97–108. [Google Scholar]
- Teo, S.M.; Mok, D.; Pham, K.; Kusel, M.; Serralha, M.; Troy, N.; Holt, B.J.; Hales, B.J.; Walker, M.L.; Hollams, E.; et al. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe 2015, 17, 704–715. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Chung, J.; Battaglia, T.; Henderson, N.; Jay, M.; Li, H.; Lieber, A.; Wu, F.; Perez-Perez, G.I.; Chen, Y.; et al. Antibiotics, birth mode, and diet shape microbiome maturation during early life. Sci. Transl. Med. 2016, 8, 343ra82. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Williams, D.; Lemas, D.J.; Spiryda, L.; Patel, K.; Carney, O.O.; Neu, J.; Carson, T.L. The Neonatal Microbiome and Its Partial Role in Mediating the Association between Birth by Cesarean Section and Adverse Pediatric Outcomes. Neonatology 2018, 114, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Giudice, G.; Cutrignelli, D.A.; Sportelli, P.; Limongelli, L.; Tempesta, A.; Gioia, G.D.; Santacroce, L.; Maiorano, E.; Favia, G. Rhinocerebral Mucormycosis with Orosinusal Involvement: Diagnostic and Surgical Treatment Guidelines. Endocr. Metab. Immune Disord. Drug Targets 2016, 16, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T.; Frank, D.N.; Ramakrishnan, V. Microbiome of the paranasal sinuses: Update and literature review. Am. J. Rhinol. Allergy 2016, 30, 3–16. [Google Scholar] [CrossRef]
- Kellner, J.D.; Vanderkooi, O.G.; MacDonald, J.; Church, D.L.; Tyrrell, G.J.; Scheifele, D.W. Changing epidemiology of invasive pneumococcal disease in Canada, 1998–2007: Update from the Calgary-area Streptococcus pneumoniae research (CASPER) study. Clin. Infect. Dis. 2009, 49, 205–212. [Google Scholar] [CrossRef]
- Marsland, B.J.; Gollwitzer, E.S. Host-microorganism interactions in pulmonary diseases. Nat. Rev. Immunol. 2014, 14, 827–835. [Google Scholar] [CrossRef]
- Santacroce, L.; Bottalico, L.; Charitos, I.A. The Impact of COVID-19 on Italy: A Lesson for the Future. Int. J. Occup. Environ. Med. 2020, 11, 151–152. [Google Scholar] [CrossRef]
- Passarelli, P.C.; Santacroce, L.; D’Addona, A.; Garcia-Godoy, F. The Coronavirus Disease-19 Infection and the Oral Mucosa. Open Access Maced. J. Med. Sci. 2020, (in press).
- Passarelli, P.C.; Passarelli, G.; Charitos, I.A.; Rella, E.; Santacroce, L.; D’Addona, A. COVID-19 and Oral Diseases: How can we Manage Hospitalized and Quarantined Patients while Reducing Risks? Electron. J. Gen. Med. 2020, 17. [Google Scholar] [CrossRef]
- Cazzolla, A.P.; Lovero, R.; Lo Muzio, L.; Testa, N.F.; Schirinzi, A.L.; Palmieri, G.; Pozzessere, P.; Procacci, V.; Di Comite, M.; Ciavarella, D.; et al. Taste and smell disorders in COVID-19 patients: Role of Interleukin-6. ACS Chem. Neurosci. 2020, 11, 2774–2781. [Google Scholar] [CrossRef]
- Santacroce, L.; Charitos, I.A.; Del Prete, R. COVID-19 in Italy: An Overview from the First Case to Date. Electron. J. Gen. Med. 2020, 17, em235. [Google Scholar] [CrossRef]
- Johnston, J.J.; Douglas, R. Adenotonsillar microbiome: An update. Postgrad. Med. J. 2018, 94, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Faner, R.; Sibila, O.; Agustí, A.; Bernasconi, E.; Chalmers, J.D.; Huffnagle, G.B.; Manichanh, C.; Molyneaux, P.L.; Paredes, R.; Pérez Brocal, V.; et al. The microbiome in respiratory medicine: Current challenges and future perspectives. Eur. Respir. J. 2017, 49, 1602086. [Google Scholar] [CrossRef] [PubMed]
- Biswas, K.; Chang, A.; Hoggard, M.; Radcliff, F.J.; Jiang, Y.; Taylor, M.W.; Darveau, R.; Douglas, R.G. Toll-like receptor activation by sino-nasal mucus in chronic rhinosinusitis. Rhinology 2017, 55, 59–69. [Google Scholar] [CrossRef]
- Dickson, R.P.; Erb-Downward, J.R.; Martinez, F.J.; Huffnagle, G.B. The Microbiome and the Respiratory Tract. Ann. Rev. Physiol. 2016, 78, 481–504. [Google Scholar] [CrossRef]
- Yadava, K.; Pattaroni, C.; Sichelstiel, A.K.; Trompette, A.; Gollwitzer, E.S.; Salami, O.; von Garnier, C.; Nicod, L.P.; Marsland, B.J. Microbiota Promotes Chronic Pulmonary Inflammation by Enhancing IL-17A and Autoantibodies. Am. J. Respir. Crit. Care Med. 2016, 193, 975–987. [Google Scholar] [CrossRef]
- Bisgaard, H.; Hermansen, M.N.; Buchvald, F.; Loland, L.; Halkjaer, L.B.; Bønnelykke, K.; Brasholt, M.; Heltberg, A.; Vissing, N.H.; Thorsen, S.V.; et al. Childhood asthma after bacterial colonization of the airway in neonates. N. Engl. J. Med. 2007, 357, 1487–1495. [Google Scholar] [CrossRef]
- Hasegawa, K.; Mansbach, J.M.; Ajami, N.J.; Petrosino, J.F.; Freishtat, R.J.; Teach, S.J.; Piedra, P.A.; Camargo, C.A., Jr. The relationship between nasopharyngeal CCL5 and microbiota on disease severity among infants with bronchiolitis. Allergy 2017, 72, 1796–1800. [Google Scholar] [CrossRef]
- Wenzel, S.E. Asthma phenotypes: The evolution from clinical to molecular approaches. Nat. Med. 2012, 18, 716. [Google Scholar] [CrossRef]
- Hilty, M.; Burke, C.; Pedro, H.; Cardenas, P.; Bush, A.; Bossley, C.; Davies, J.; Ervine, A.; Poulter, L.; Pachter, L.; et al. Disordered microbial communities in asthmatic airways. PLoS ONE 2010, 5, e8578. [Google Scholar] [CrossRef]
- Huang, Y.J.; Nariya, S.; Harris, J.M.; Lynch, S.V.; Choy, D.F.; Arron, J.R.; Boushey, H. The airway microbiome in patients with severe asthma: Associations with disease features and severity. J. Allergy Clin. Immunol. 2015, 136, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Durack, J.; Boushey, H.A.; Huang, Y.J. Incorporating the airway microbiome into asthma phenotyping: Moving toward personalized medicine for noneosinophilic asthma. J. Allergy Clin. Immunol. 2018, 141, 82–83. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.L.; Daly, J.; Baines, K.J.; Yang, I.A.; Upham, J.W.; Reynolds, P.N.; Hodge, S.; James, A.L.; Hugenholtz, P.; Willner, D.; et al. Airway dysbiosis: Haemophilus influenzae and Tropheryma in poorly controlled asthma. Eur. Respir. J. 2016, 47, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Renwick, J.; McNally, P.; John, B.; DeSantis, T.; Linnane, B.; Murphy, P. The microbial community of the cystic fibrosis airway is disrupted in early life. PLoS ONE 2014, 9, e109798. [Google Scholar] [CrossRef] [PubMed]
- Pragman, A.A.; Kim, H.B.; Reilly, C.S.; Wendt, C.; Isaacson, R.E. The lung microbiome in moderate and severe chronic obstructive pulmonary disease. PLoS ONE 2012, 7, e47305. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.L.; Baines, K.J.; Horvat, J.C.; Essilfie, A.T.; Brown, A.C.; Tooze, M.; McDonald, V.M.; Gibson, P.G.; Hansbro, P.M. COPD is characterized by increased detection of Haemophilus influenzae, Streptococcus pneumoniae and a deficiency of Bacillus species. Respirology 2016, 21, 697–704. [Google Scholar] [CrossRef]
- Mosca, A.; Carucci, A.; Santacroce, L.; Schettini, F.; De Mattia, D.; Miragliotta, G. Streptococcus pneumoniae nasopharyngeal colonization in young healthy children: Rate of carriage, serotype distribution, and antibiotic resistance. New Microbiol. 2003, 26, 187–192. [Google Scholar]
- Basis, C.M.; Erb-Downward, J.R.; Dickson, R.P.; Freeman, C.M.; Schmidt, T.M.; Young, V.B.; Beck, J.M.; Curtis, J.L.; Huffnagle, G.B. Analysis of the upper respiratory tract microbiotas as the source of the lung and gastric microbiotas in healthy individuals. MBio 2015, 6, e00037-15. [Google Scholar] [CrossRef]
- Samuelson, D.R.; Welsh, D.A.; Shellito, J.E. Regulation of lung immunity and host defense by the intestinal microbiota. Front. Microbiol. 2015, 7, 1085. [Google Scholar] [CrossRef]
- Bottalico, L.; Castellaneta, F.; Charitos, I.A. From Hydrotherapy to the Discovery of The Gut Microbiota: The Historical Gastrointestinal Health Concept. Pharmacophore 2020, 11, 82–90. [Google Scholar]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef]
- Gozdz, J.; Ober, C.; Vercelli, D. Innate Immunity and Asthma Risk. N. Engl. J. Med. 2016, 375, 1898–1899. [Google Scholar] [PubMed]
- Oddy, W.H. Breastfeeding, Childhood asthma, and allergic disease. Ann. Nutr. Metab. 2017, 70 (Suppl. 2), 26–36. [Google Scholar] [CrossRef] [PubMed]
- El Aidy, S.; Hooiveld, G.; Tremaroli, V.; Bäckhed, F.; Kleerebezem, M. The gut microbiota and mucosal homeostasis: Colonized at birth or at adulthood, does it matter? Gut Microbes 2013, 4, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Bermúdez-Humarán, L.G.; Langella, P. Gnotobiotic Rodents: An In Vivo Model for the Study of Microbe-Microbe Interactions. Front. Microbiol. 2016, 7, 409. [Google Scholar] [CrossRef] [PubMed]
- Ballini, A.; Santacroce, L.; Cantore, S.; Bottalico, L.; Dipalma, G.; Topi, S.; Saini, R.; De Vito, D.; Inchingolo, F. Probiotics efficacy on oxidative stress values in inflammatory bowel disease: A randomized double-blinded placebo-controlled pilot study. Endocr. Metab. Immune. Disord. Drug Targets 2019, 19, 373–381. [Google Scholar] [CrossRef]
- Arrieta, M.C.; Finlay, B. The intestinal microbiota and allergic asthma. J. Infect. 2014, 69 (Suppl. 1), S53–S55. [Google Scholar] [CrossRef]
- Abrahamsson, T.R.; Jakobsson, H.E.; Andersson, A.F.; Björkstén, B.; Engstrand, L.; Jenmalm, M.C. Low gut microbiota diversity in early infancy precedes asthma at school age. Clin. Exp. Allergy. 2014, 44, 842–850. [Google Scholar] [CrossRef]
- Fujimura, K.E.; Sitarik, A.R.; Havstad, S.; Lin, D.L.; Levan, S.; Fadrosh, D.; Panzer, A.R.; LaMere, B.; Rackaityte, E.; Lukacs, N.W.; et al. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat. Med. 2016, 22, 1187–1191. [Google Scholar] [CrossRef]
- Schuijs, M.J.; Willart, M.A.; Vergote, K.; Gras, D.; Deswarte, K.; Ege, M.J.; Madeira, F.B.; Beyaert, R.; van Loo, G.; Bracher, F.; et al. Farm dust and endotoxin protect against allergy through A20 induction in lung epithelial cells. Science 2015, 349, 1106–1110. [Google Scholar] [CrossRef]
- Lovreglio, P.; Bukvic, N.; Fustinoni, S.; Ballini, A.; Drago, I.; Foà, V.; Guanti, G.; Soleo, L. Lack of genotoxic effect in workers exposed to very low doses of 1,3-butadiene. Arch. Toxicol. 2006, 80, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Loss, G.; Depner, M.; Ulfman, L.H.; van Neerven, R.J.; Hose, A.J.; Genuneit, J.; Karvonen, A.M.; Hyvärinen, A.; Kaulek, V.; Roduit, C.; et al. PASTURE study group. Consumption of unprocessed cow’s milk protects infants from common respiratory infections. J. Allergy Clin. Immunol. 2015, 135, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Dickson, R.P.; Erb-Downward, J.R.; Huffnagle, G.B. Homeostasis and its disruption in the lung microbiome. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 309, L1047–L1055. [Google Scholar] [CrossRef] [PubMed]
- Toscano, M.; De Grandi, R.; Grossi, E.; Drago, L. Role of the Human Breast Milk-Associated Microbiota on the Newborns’ Immune System: A Mini Review. Front. Microbiol. 2017, 25, 2100. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Crincoli, V.; Ballini, A.; Di Comite, M.; Tettamanti, L.; Coscia, M.F.; Mastrangelo, F.; De Vito, D. Microbiological investigation of medication-related osteonecrosis of the jaw: Preliminary results. J. Biol. Regul. Homeost. Agents 2015, 29, 977–983. [Google Scholar]
- Enaud, R.; Prevel, R.; Ciarlo, E.; Beaufils, F.; Wieërs, G.; Guery, B.; Delhaes, L. The Gut-Lung Axis in Health and Respiratory Diseases: A Place for Inter-Organ and Inter-Kingdom Crosstalks. Front. Cell Infect. Microbiol. 2020, 19, 9. [Google Scholar] [CrossRef]
- Santacroce, L. Letter in response to the article “Enhancing immunity in viral infections, with special emphasis on COVID-19: A review” (Jayawardena et al.). Diabetes Metab. Syndr. 2020, 14, 927. [Google Scholar] [CrossRef]
- Charitos, I.A.; Ballini, A.; Bottalico, L.; Cantore, S.; Passarelli, P.C.; Inchingolo, F.; D’Addona, A.; Santacroce, L. Special features of SARS-CoV2 in daily practice. World J. Clin. Cases 2020, 8, 3920–3933. [Google Scholar] [CrossRef]
- Charitos, I.A.; Del Prete, R.; Mosca, A.; Inchingolo, F.; Carretta, D.; Ballini, A.; Santacroce, L. What We Have Learned for the Future About COVID-19 and Healthcare Management of it? Acta Biomed. 2020, 91. [Google Scholar] [CrossRef]
- Cantore, S.; Ballini, A. Coronavirus Disease 2019 (COVID-19) pandemic burst and its relevant consequences in dental practice. Open Dent. J. 2020, 14, 111–112. [Google Scholar] [CrossRef]
- Pham, V.H.; Gargiulo, I.C.; Nguyen, K.C.D.; Le, S.H.; Tran, D.K.; Nguyen, Q.V.; Pham, H.T.; Aityan, S.; Pham, S.T.; Cantore, S.; et al. Rapid and sensitive diagnostic procedure for multiple detection of pandemic Coronaviridae family members SARS-CoV-2, SARS-CoV, MERS-CoV and HCoV: A translational research and cooperation between the Phan Chau Trinh University in Vietnam and University of Bari “Aldo Moro” in Italy. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7173–7191. [Google Scholar] [PubMed]
- Santacroce, L.; Charitos, I.A.; Bottalico, L. A successful history: Probiotics and their potential as antimicrobials. Expert Rev. Anti-Infect. Ther. 2019, 17, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Isacco, C.G.; Ballini, A.; De Vito, D.; Nguyen, K.C.D.; Cantore, S.; Bottalico, L.; Quagliuolo, L.; Boccellino, M.; Di Domenico, M.; Santacroce, L.; et al. Rebalance the oral microbiota as efficacy tool in endocrine, metabolic, and immune disorders. Endocr. Metab. Immune Disord. Drug Targets 2020. [Google Scholar] [CrossRef]
- Ballini, A.; Cantore, S.; Scacco, S.; Coletti, D.; Tatullo, M. Mesenchymal stem cells as promoters, enhancers, and playmakers of the translational regenerative medicine 2018. Stem Cells Int. 2018, 2018, 6927401. [Google Scholar] [CrossRef]
- Ballini, A.; Gnoni, A.; De Vito, D.; Dipalma, G.; Cantore, S.; Gargiulo, I.C.; Saini, R.; Santacroce, L.; Topi, S.; Scarano, A.; et al. Effect of probiotics on the occurrence of nutrition absorption capacities in healthy children: A randomized double-blinded placebo-controlled pilot study. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8645–8657. [Google Scholar]
- Inchingolo, F.; Dipalma, G.; Cirulli, N.; Cantore, S.; Saini, R.S.; Altini, V.; Santacroce, L.; Ballini, A.; Saini, R. Microbiological results of improvement in periodontal condition by administration of oral probiotics. J. Biol. Regul. Homeost. Agents 2018, 32, 1323–1328. [Google Scholar]
- Ballini, A.; Santacroce, L.; Cantore, S.; Bottalico, L.; Dipalma, G.; Vito, D.; Saini, R.; Inchingolo, F. Probiotics Improve Urogenital Health in Women. Maced. J. Med. Sci. 2018, 6, 1845–1850. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Fungi |
|---|---|
| Prevotella | Aspergillus |
| Sphingomonas | Cladosporium |
| Pseudomonas | Penicillum |
| Acinetobacter | Eurotium |
| Fusobacterium | Candida |
| Megasphaera | Malassezia |
| Veillonella | Neosartorya |
| Staphylococcus | Saccharomyces |
| Streptococcus |
| Factors Influencing the Gut Microbiota | |
|---|---|
| Intrinsic | Extrinsic |
| Gastric secretion | Age |
| Oxigen presence/concentration | Delivery (cesarean section, natural) |
| Motility | Diet (breast milk, formula milk, pre- and probiotic foods) |
| Mucus and Gastro-Intestinal (GI) secretions | Environmental stress |
| Antimicrobiale peptides | Infections |
| Immunity state | Drugs (Proton pump inhibitors, H2 Blockers, Prokinetics, Antibiotics, Laxatives, Opioids, Nonsteroidal anti-inflammatory drugs: NSAIDs) |
| Risk of Respiratory Diseases | |
|---|---|
| Alteration of Lung Microbiota | Alteration of Gut Microbiota |
| Bisgaard H. et al. (N Engl J Med 2007): Pneumonia and bronchiolitis in the first 3 years | Abrahamsson T.R. et al. (Clin Exp Allergy. 2014): Asthma at 7 years old |
| Huang YJ and Boushey HA (Am Thorac Soc. 2014): Pneumonia and bronchiolitis in the first 3 years | Fujimura K.E. et al. (Nat Med 2016): Asthma at 6–7 years old |
| Teo S.M. et al. (Cell Host Microbe 2015): Asthma at 5 years old | Arrieta MC and Finlay B. (J Infect. 2014): Asthma at 5 years old |
| Hasegawa K. et al. (Allergy 2017): Recurrent respiratory infection in the first 3 years | Stokholm J. et al. (Nat Commun. 2018): Asthma at age 5 years old |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santacroce, L.; Charitos, I.A.; Ballini, A.; Inchingolo, F.; Luperto, P.; De Nitto, E.; Topi, S. The Human Respiratory System and its Microbiome at a Glimpse. Biology 2020, 9, 318. https://doi.org/10.3390/biology9100318
Santacroce L, Charitos IA, Ballini A, Inchingolo F, Luperto P, De Nitto E, Topi S. The Human Respiratory System and its Microbiome at a Glimpse. Biology. 2020; 9(10):318. https://doi.org/10.3390/biology9100318
Chicago/Turabian StyleSantacroce, Luigi, Ioannis Alexandros Charitos, Andrea Ballini, Francesco Inchingolo, Paolo Luperto, Emanuele De Nitto, and Skender Topi. 2020. "The Human Respiratory System and its Microbiome at a Glimpse" Biology 9, no. 10: 318. https://doi.org/10.3390/biology9100318
APA StyleSantacroce, L., Charitos, I. A., Ballini, A., Inchingolo, F., Luperto, P., De Nitto, E., & Topi, S. (2020). The Human Respiratory System and its Microbiome at a Glimpse. Biology, 9(10), 318. https://doi.org/10.3390/biology9100318