Storage Compound Accumulation in Diatoms as Response to Elevated CO2 Concentration


Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Biomass Determination
2.3. Lipid Extraction and Measurement
2.4. Chrysolaminarin Measurement
2.5. Protein Extraction
2.6. Enzyme Activity Measurements
2.7. Statistical Analysis
3. Results
3.1. Growth and Biomass as Response to High CO2
3.2. Storage Compounds (TAGs)
3.3. Storage Compounds (Chrysolaminarin)
3.4. Activities of Key Enzymes from Carbon Metabolism
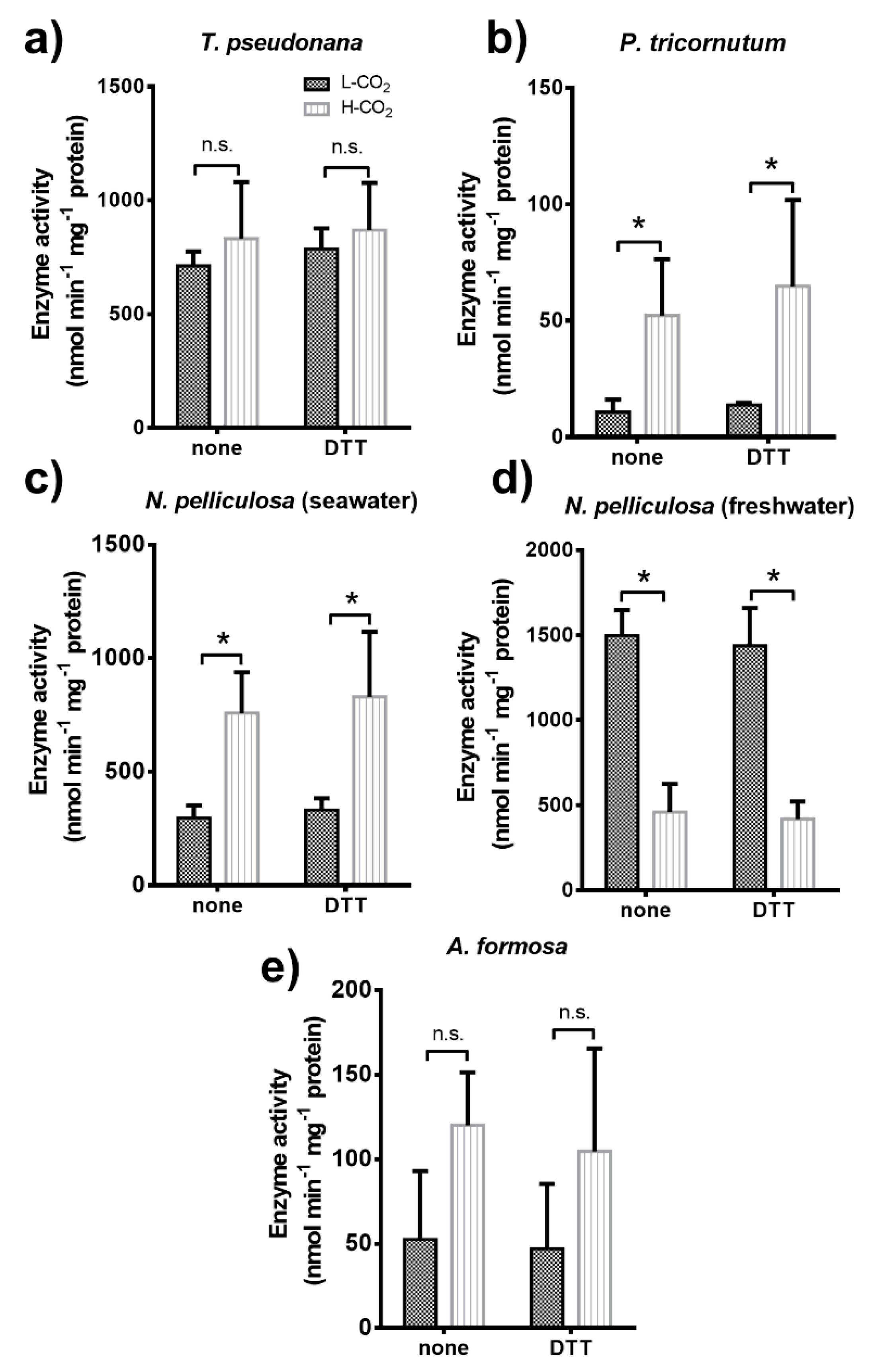
3.4.1. Glyceraldehyde 3-Phosphate Dehydrogenase (GAPDH)
3.4.2. Phosphoglycerate Kinase (PGK)
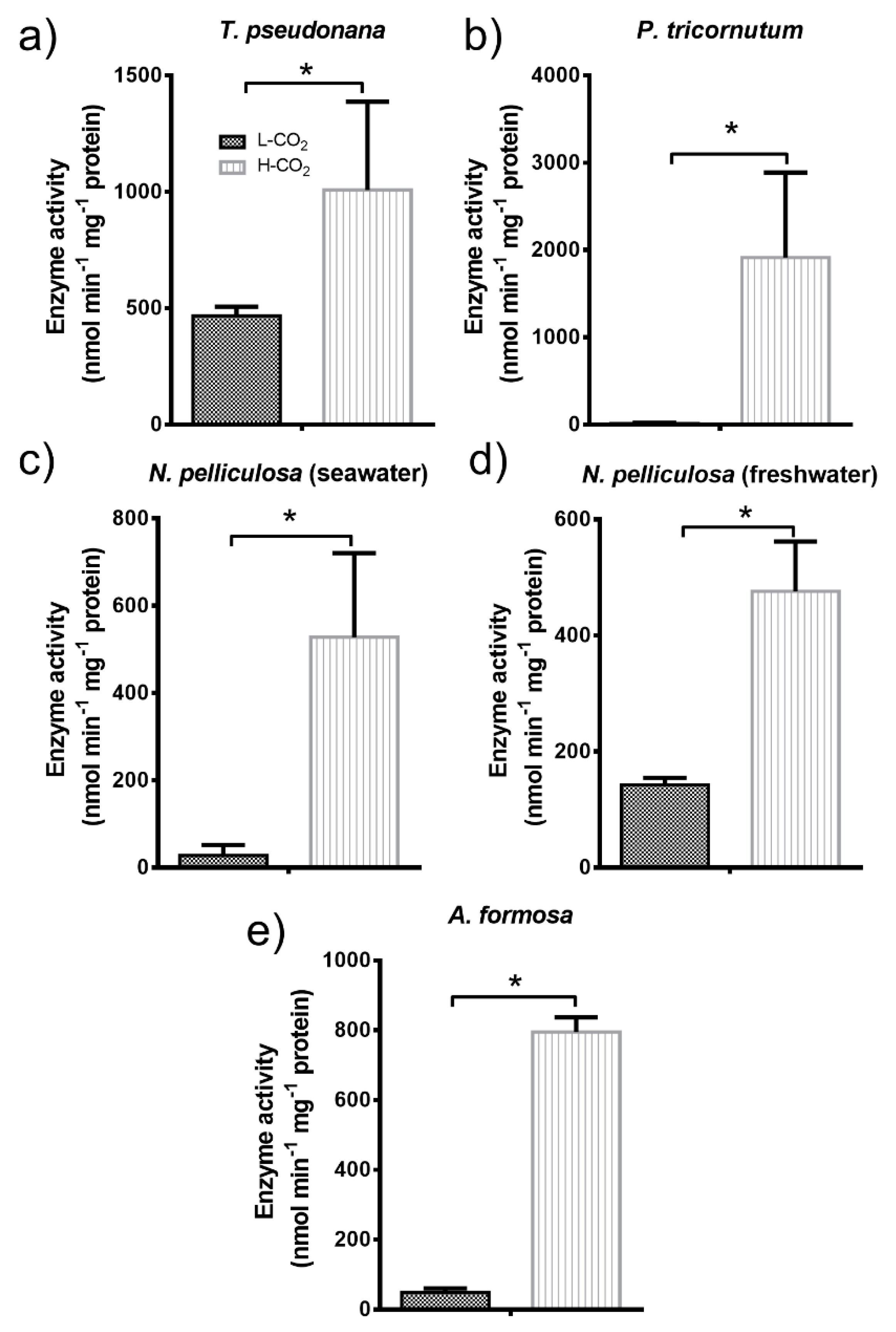
3.4.3. Pyruvate Kinase (PK)
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Armbrust, E.V. The life of diatoms in the world’s oceans. Nature 2009, 459, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Jakob, T.; Fanesi, A.; Wilhelm, C. Towards an understanding of the molecular regulation of carbon allocation in diatoms: The interaction of energy and carbon allocation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chiovitti, A.; Molino, P.; Crawford, S.A.; Teng, R.; Wetherbee, R.; Chiovitti, A.; Molino, P.; Crawford, S.A.; Teng, R. The glucans extracted with warm water from diatoms are mainly derived from intracellular chrysolaminaran and not extracellular polysaccharides. Eur. J. Phycol. 2004, 39, 117–128. [Google Scholar] [CrossRef]
- Maeda, Y.; Nojima, D.; Yoshino, T.; Tanaka, T. Structure and properties of oil bodies in diatoms. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160408. [Google Scholar] [CrossRef] [PubMed]
- Cuellar-Bermudez, S.P.; Romero-Ogawa, M.A.; Rittmann, B.E.; Parra-Saldivar, R. Algae Biofuels Production Processes, Carbon Dioxide Fixation and Biorefinery Concept. J. Pet. Environ. Biotechnol. 2014, 5. [Google Scholar] [CrossRef]
- Hildebrand, M.; Davis, A.K.; Smith, S.R.; Traller, J.C.; Abbriano, R. The place of diatoms in the biofuels industry. Biofuels 2012, 3, 221–240. [Google Scholar] [CrossRef]
- Hildebrand, M.; Abbriano, R.M.; Polle, J.E.W.; Traller, J.C.; Trentacoste, E.M.; Smith, S.R.; Davis, A.K. Metabolic and cellular organization in evolutionarily diverse microalgae as related to biofuels production. Curr. Opin. Chem. Biol. 2013, 17, 506–514. [Google Scholar] [CrossRef]
- Yi, Z.; Xu, M.; Di, X.; Brynjolfsson, S.; Fu, W. Exploring Valuable Lipids in Diatoms. Front. Mar. Sci. 2017, 4, 1–10. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Adams, C.; Bugbee, B. Enhancing lipid production of the marine diatom Chaetoceros gracilis: Synergistic interactions of sodium chloride and silicon. J. Appl. Phycol. 2013. [Google Scholar] [CrossRef]
- Wang, X.-W.; Liang, J.-R.; Luo, C.-S.; Chen, C.-P.; Gao, Y.-H. Biomass, total lipid production, and fatty acid composition of the marine diatom Chaetoceros muelleri in response to different CO2 levels. Bioresour. Technol. 2014, 161, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Lu, Y.; Peng, K.; Huang, T.; Niu, Y.; Xie, W.; Yang, W.; Liu, J. Glycerol and neutral lipid production in the oleaginous marine diatom Phaeodactylum tricornutum promoted by overexpression of glycerol-3-phosphate dehydrogenase. Biotechnol. Biofuels 2014, 7, 1–9. [Google Scholar] [CrossRef]
- Mekhalfi, M.; Amara, S.; Robert, S.; Carriere, F.; Gontero, B. Effect of environmental conditions on various enzyme activities and triacylglycerol contents in cultures of the freshwater diatom, Asterionella formosa (Bacillariophyceae). Biochimie 2013, 101, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Levitan, O.; Dinamarca, J.; Hochman, G.; Falkowski, P.G. Diatoms: A fossil fuel of the future. Trends Biotechnol. 2014, 32, 117–124. [Google Scholar] [CrossRef]
- Breuer, G.; Lamers, P.P.; Martens, D.E.; Draaisma, R.B.; Wijffels, R.H. The impact of nitrogen starvation on the dynamics of triacylglycerol accumulation in nine microalgae strains. Bioresour. Technol. 2012, 124, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Yodsuwan, N.; Sawayama, S. Effect of nitrogen concentration on growth, lipid production and fatty acid pro fi les of the marine diatom Phaeodactylum tricornutum. Agric. Nat. Resour. 2017, 51, 190–197. [Google Scholar] [CrossRef]
- Lin, Q.; Zhuo, W.-H.; Wang, X.; Chen, C.-P.; Gao, Y.; Liang, J.-R. Effects of fundamental nutrient stresses on the lipid accumulation profiles in two diatom species Thalass. Weissflogii Chaetoceros Muelleri. Bioprocess Biosyst. Eng. 2018, 41, 1213–1224. [Google Scholar] [CrossRef]
- Zulu, N.N.; Zienkiewicz, K.; Vollheyde, K.; Feussner, I. Current trends to comprehend lipid metabolism in diatoms. Prog. Lipid Res. 2018, 70, 1–16. [Google Scholar] [CrossRef]
- Mekhalfi, M.; Avilan, L.; Lebrun, R.; Botebol, H.; Gontero, B. Consequences of the presence of 24-epibrassinolide, on cultures of a diatom, Asterionella Formosa. Biochimie 2012, 94, 1213–1220. [Google Scholar] [CrossRef]
- Comte, M.; Lupette, J.; Seddiki, K.; Mei, C.; Dolch, L.-J.; Gros, V.; Reveille, F.; Jouhet, J.; Marechal, E. Screening for biologically annotated drugs that trigger triacylglycerol accumulation in the diatom. Phaeodactylum Plant Physiol. 2018, 177, 532–552. [Google Scholar] [CrossRef]
- Prioretti, L.; Avilan, L.; Carrière, F.; Montané, M.H.; Field, B.; Grégori, G.; Menand, B.; Gontero, B. The inhibition of TOR in the model diatom Phaeodactylum tricornutum promotes a get-fat growth regime. Algal. Res. 2017, 26, 265–274. [Google Scholar] [CrossRef]
- Xia, S.; Gao, B.; Li, A.; Xiong, J.; Ao, Z.; Zhang, C. Preliminary characterization, antioxidant properties and production of chrysolaminarin from marine diatom Odontella aurita. Mar. Drugs 2014, 12, 4883–4897. [Google Scholar] [CrossRef] [PubMed]
- Carballo, C.; Chronopoulou, E.G.; Maya, C.; Labrou, N.E.; Infante, C.; Power, D.M.; Manchado, M. Antioxidant capacity and immunomodulatory effects of a chrysolaminarin-enriched extract in Senegalese sole. Fish Shellfish. Immunol. 2018, 82, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kusaikin, M.I.; Ermakova, S.P.; Shevchenko, N.M. Structurel characteristics and antitumor activity of a new chrysolaminarin from the diatom Synedra acus. Chem. Nat. Compd. 2010, 46, 5–8. [Google Scholar] [CrossRef]
- Varum, K.M.; Myklestad, S. Effects of light, salinity and nutrient limitation of β-1,3-D-glucan and exo-D-glucanase activity in Skeletonema costatum (Grev.) Cleve. J. Exp. Mar. Biol. Ecol. 1984, 83, 13–25. [Google Scholar] [CrossRef]
- Botte, P.; D’Ippolito, G.; Gallo, C.; Sardo, A.; Fontana, A. Combined exploitation of CO2 and nutrient replenishment for increasing biomass and lipid productivity of the marine diatoms Thalassiosira weissflogii and Cyclotella cryptica. J. Appl. Phycol. 2018, 30, 243–251. [Google Scholar] [CrossRef]
- Sabia, A.; Clavero, E.; Pancaldi, S.; Salvadó Rovira, J. Effect of different CO2 concentrations on biomass, pigment content, and lipid production of the marine diatom Thalassiosira pseudonana. Appl. Microbiol. Biotechnol. 2018, 102, 1945–1954. [Google Scholar] [CrossRef]
- Ishida, Y.; Hiragushi, N.; Kitaguchi, H.; Mitsutani, A. A highly CO2 -tolerant diatom, Thalassiosira weissflogii H1, enriched from coastal sea, and its fatty acid composition. Fish. Sci. 2000, 66, 655–659. [Google Scholar] [CrossRef]
- Hopkinson, B.M.; Dupont, C.L.; Allen, A.E.; Morel, F.M.M. Efficiency of the CO2-concentrating mechanism of diatoms. Proc. Natl. Acad. Sci. USA 2011, 108, 3830–3837. [Google Scholar] [CrossRef]
- Nakajima, K.; Tanaka, A.; Matsuda, Y. SLC4 family transporters in a marine diatom directly pump bicarbonate from seawater. Proc. Natl. Acad. Sci. USA 2013, 110, 1767–1772. [Google Scholar] [CrossRef]
- Kikutani, S.; Nakajima, K.; Nagasato, C.; Tsuji, Y.; Miyatake, A.; Matsuda, Y. Thylakoid luminal θ-carbonic anhydrase critical for growth and photosynthesis in the marine diatom Phaeodactylum tricornutum. Proc. Natl. Acad. Sci. USA 2016, 113, 9828–9833. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.L.; Clement, R.; Kosta, A.; Maberly, S.C.; Gontero, B. A new widespread subclass of carbonic anhydrase in marine phytoplankton. ISME J. 2019. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.L.; Clément, R.; Maberly, S.C.; Gontero, B.; Clement, R.; Maberly, S.C.; Gontero, B. Regulation of the Calvin – Benson – Bassham cycle in the enigmatic diatoms: Biochemical and evolutionary variations on an original theme. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef] [PubMed]
- Clement, R.; Dimnet, L.; Maberly, S.C.; Gontero, B. The nature of the CO2 -concentrating mechanisms in a marine diatom, Thalassiosira pseudonana. New Phytol. 2016, 209, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- Hempel, N.; Petrick, I.; Behrendt, F. Biomass productivity and productivity of fatty acids and amino acids of microalgae strains as key characteristics of suitability for biodiesel production. J. Appl. Phycol. 2012, 24, 1407–1418. [Google Scholar] [CrossRef]
- Cavalier, J.F.; Lafont, D.; Boullanger, P.; Houisse, D.; Giallo, J.; Ballester, J.M.; Carrière, F. Validation of lipolysis product extraction from aqueous/biological samples, separation and quantification by thin-layer chromatography with flame ionization detection analysis using O-cholesteryl ethylene glycol as a new internal standard. J. Chromatogr. A 2009, 1216, 6543–6548. [Google Scholar] [CrossRef]
- Granum, E.; Myklestad, S.M. A simple combined method for determination of β-1,3-glucan and cell wall polysaccharides in diatoms. Sci. Technol. 2002, 477, 155–161. [Google Scholar]
- Bosco, M.B.; Aleanzi, M.C.; Iglesias, A.Á. Plastidic phosphoglycerate kinase from Phaeodactylum tricornutum: On the critical role of cysteine residues for the enzyme function. Protist 2012, 163, 188–203. [Google Scholar] [CrossRef]
- Mccarthy, A.; Rogers, S.P.; Duffy, S.J.; Campbell, D.A. Elevated carbon dioxide differentially alters the photophysiology of Thalassiosira pseudonana (Bacillariophyceae) and Emiliania huxleyi (Haptophyta). J. Phycol. 2012, 48, 635–646. [Google Scholar] [CrossRef]
- Li, G.; Campbell, D.A. Rising CO2 Interacts with Growth Light and Growth Rate to Alter Photosystem II Photoinactivation of the Coastal Diatom Thalassiosira pseudonana. PLoS ONE 2013, 8, e55562. [Google Scholar] [CrossRef]
- Gao, K.; Xu, J.; Gao, G.; Li, Y.; Hutchins, D.A.; Huang, B.; Wang, L.; Zheng, Y.; Jin, P.; Cai, X.; et al. Rising CO2 and increased light exposure synergistically reduce marine primary productivity. Nat. Clim. Chang. 2012, 2, 519. [Google Scholar] [CrossRef]
- Wu, S.; Huang, A.; Zhang, B.; Huan, L.; Zhao, P.; Lin, A.; Wang, G. Enzyme activity highlights the importance of the oxidative pentose phosphate pathway in lipid accumulation and growth of Phaeodactylum tricornutum under CO2 concentration. Biotechnol. Biofuels 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Clement, R.; Jensen, E.; Prioretti, L.; Maberly, S.C.; Gontero, B. Diversity of CO2 concentrating mechanisms and responses to CO2 concentration in marine and freshwater diatoms. J. Exp. Bot. 2017, 68, 3925–3935. [Google Scholar] [CrossRef] [PubMed]
- Coombs, J.; Darley, W.M.; Volcani, B.E. Studies on the Biochemistry and Fine Structure of Silica Shell Formation in Diatoms. Chemical Composition of Navicula pelliculosa during Silicon-Starvation Synchrony. Plant Physiol. 1967, 42, 1601–1606. [Google Scholar] [CrossRef]
- Tadros, M.G. Screening and Characterizing Oleaginous Microalgal Species from the Southeastern United States. Final Subcontract Report; Alabama A&M University: Huntsville, AL, USA, 1985. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Sayanova, O.; Mimouni, V.; Ulmann, L.; Morant, A.; Napier, J.A.; Pasquet, V. Modulation of lipid biosynthesis by stress in diatoms. Philos. Trans. R. Soc. B Biol. Sci. 2017. [Google Scholar] [CrossRef]
- Huang, W.; Haferkamp, I.; Lepetit, B.; Molchanova, M.; Hou, S.; Jeblick, W.; Río Bártulos, C.; Kroth, P.G. Reduced vacuolar β-1,3-glucan synthesis affects carbohydrate metabolism as well as plastid homeostasis and structure in Phaeodactylum tricornutum. Proc. Natl. Acad. Sci. USA 2018, 115, 4791–4796. [Google Scholar] [CrossRef]
- Hildebrand, M.; Manandhar-Shrestha, K.; Abbriano, R. Effects of chrysolaminarin synthase knockdown in the diatom Thalassiosira pseudonana: Implications of reduced carbohydrate storage relative to green algae. Algal Res. 2017, 23, 66–77. [Google Scholar] [CrossRef]
- Yang, Y.; Li, D.; Chen, T.; Hao, T.; Balamurugan, S.; Yang, W.; Liu, J.; Li, H. Overproduction of Bioactive Algal Chrysolaminarin by the Critical Carbon Flux Regulator Phosphoglucomutase. Biotechnol. J. 2019, 1800220, 1–8. [Google Scholar] [CrossRef]
- Gardner, R.D.; Lohman, E.; Gerlach, R.; Cooksey, K.E.; Peyton, B.M. Comparison of CO2 and bicarbonate as inorganic carbon sources for triacylglycerol and starch accumulation in Chlamydomonas reinhardtii. Biotechnol. Bioeng. 2013, 110, 87–96. [Google Scholar] [CrossRef]
- Torres-Romero, I.; Kong, F.; Légeret, B.; Beisson, F.; Peltier, G.; Li-Beisson, Y. Chlamydomonas cell cycle mutant crcdc5 over-accumulates starch and oil. Biochimie 2019. [Google Scholar] [CrossRef] [PubMed]
- Tanadul, O.; Vandergheynst, J.S.; Beckles, D.M.; Powell, A.L.T.; Labavitch, J.M. The impact of elevated CO2 concentration on the quality of algal starch as a potential biofuel feedstock. Biotechnol. Bioeng. 2014, 111, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- Caballero, M.A.; Jallet, D.; Shi, L.; Rithner, C.; Zhang, Y.; Peers, G. Quantification of chrysolaminarin from the model diatom Phaeodactylum tricornutum. Algal Res. 2016, 20, 180–188. [Google Scholar] [CrossRef]
- Kuwata, A.; Hama, T.; Takahashi, M. Ecophysiological characterization of two life forms, resting spores and resting cells, of a marine planktonic diatom, Chaetoceros pseudocurvisetus, formed under nutrient depletion. Mar. Ecol. Prog. Ser. 1993, 102, 245–256. [Google Scholar] [CrossRef]
- Cheng, D.; Li, D.; Yuan, Y.; Zhou, L.; Li, X.; Wu, T.; Wang, L.; Zhao, Q.; Wei, W.; Sun, Y. Improving carbohydrate and starch accumulation in Chlorella sp. AE10 by a novel two-stage process with cell dilution. Biotechnol. Biofuels 2017, 10, 1–14. [Google Scholar] [CrossRef]
- Puspanadan, S.; Wong, X.J.; Lee, C.K. Optimization of freshwater microalgae, Arthrospira sp. (Spirulina) for high starch production. Int. Food Res. J. 2018, 25, 1266–1272. [Google Scholar]
- Rehman, Z.U.; Anal, A.K. Enhanced lipid and starch productivity of microalga (Chlorococcum sp. TISTR 8583) with nitrogen limitation following effective pretreatments for biofuel production. Biotechnol. Rep. 2019, 21, e00298. [Google Scholar] [CrossRef]
- Avilan, L.; Puppo, C.; Erales, J.; Woudstra, M.; Lebrun, R.; Gontero, B. CP12 residues involved in the formation and regulation of the glyceraldehyde-3-phosphate dehydrogenase-CP12-phosphoribulokinase complex in Chlamydomonas Reinhardtii. Mol. Biosyst. 2012, 8, 2994–3002. [Google Scholar] [CrossRef]
- Erales, J.; Lignon, S.; Gontero, B. CP12 from Chlamydomonas reinhardtii, a permanent specific “chaperone-like” protein of glyceraldehyde-3-phosphate dehydrogenase. J. Biol. Chem. 2009, 284, 12735–12744. [Google Scholar] [CrossRef]
- Morisse, S.; Michelet, L.; Bedhomme, M.; Marchand, C.H.; Calvaresi, M.; Trost, P.; Fermani, S.; Zaffagnini, M.; Lemaire, S.D. Thioredoxin-dependent redox regulation of chloroplastic phosphoglycerate kinase from Chlamydomonas reinhardtii. J. Biol. Chem. 2014, 289, 30012–30024. [Google Scholar] [CrossRef]
- Gontero, B.; Maberly, S.C. An intrinsically disordered protein, CP12: Jack of all trades and master of the Calvin cycle. Biochem. Soc. Trans. 2012, 40, 995–999. [Google Scholar] [CrossRef] [PubMed]
- Groben, R.; Kaloudas, D.; Raines, C.A.; Offmann, B.; Maberly, S.C.; Gontero, B. Comparative sequence analysis of CP12, a small protein involved in the formation of a Calvin cycle complex in photosynthetic organisms. Photosynth. Res. 2010, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Maberly, S.C.; Courcelle, C.; Groben, R.; Gontero, B. Phylogenetically-based variation in the regulation of the Calvin cycle enzymes, phosphoribulokinase and glyceraldehyde-3-phosphate dehydrogenase, in algae. J. Exp. Bot. 2010, 61, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Clement, R.; Lignon, S.; Mansuelle, P.; Jensen, E.; Pophillat, M.; Lebrun, R.; Denis, Y.; Puppo, C.; Maberly, S.C.; Gontero, B. Responses of the marine diatom Thalassiosira pseudonana to changes in CO2 concentration: A proteomic approach. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Kroth, P.G.; Chiovitti, A.; Gruber, A.; Martin-Jezequel, V.; Mock, T.; Parker, M.S.; Stanley, M.S.; Kaplan, A.; Caron, L.; Weber, T.; et al. A model for carbohydrate metabolism in the diatom Phaeodactylum tricornutum deduced from comparative whole genome analysis. PLoS ONE 2008, 3, e1426. [Google Scholar] [CrossRef]
- Thieulin-Pardo, G.; Remy, T.; Lignon, S.; Lebrun, R.; Gontero, B. Phosphoribulokinase from Chlamydomonas reinhardtii: A Benson-Calvin cycle enzyme enslaved to its cysteine residues. Mol. Biosyst. 2015, 11, 1134–1145. [Google Scholar] [CrossRef]
- Mekhalfi, M.; Puppo, C.; Avilan, L.; Lebrun, R.; Mansuelle, P.; Maberly, S.C.; Gontero, B. Glyceraldehyde-3-phosphate dehydrogenase is regulated by ferredoxin-NADP reductase in the diatom Asterionella formosa. New Phytol. 2014, 203, 414–423. [Google Scholar] [CrossRef]
- Troncoso-Ponce, M.A.; Kruger, N.J.; Ratcliffe, G.; Garcés, R.; Martínez-Force, E. Characterization of glycolytic initial metabolites and enzyme activities in developing sunflower (Helianthus annuus L.) seeds. Phytochemistry 2009, 70, 1117–1122. [Google Scholar] [CrossRef]
- Bourgis, F.; Kilaru, A.; Cao, X.; Ngando-Ebongue, G.-F.; Drira, N.; Ohlrogge, J.B.; Arondel, V. Comparative transcriptome and metabolite analysis of oil palm and date palm mesocarp that differ dramatically in carbon partitioning. Proc. Natl. Acad. Sci. USA 2011, 108, 12527–12532. [Google Scholar] [CrossRef]
- Tan, K.W.M.; Lee, Y.K. The dilemma for lipid productivity in green microalgae: Importance of substrate provision in improving oil yield without sacrificing growth. Biotechnol. Biofuels 2016, 9, 1–14. [Google Scholar] [CrossRef]
- Cai, Y.; Zhang, W.; Jin, J.; Yang, X.; You, X.; Yan, H.; Wang, L.; Chen, J.; Xu, J.; Chen, W.; et al. OsPKpα1 encodes a plastidic pyruvate kinase that affects starch biosynthesis in the rice endosperm. J. Integr. Plant Biol. 2018, 60, 1097–1118. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Hao, G.; Wang, L.; Wang, H.; Gu, Z.; Liu, L.; Zhang, H.; Chen, W.; Chen, Y.Q. Identification of a critical determinant that enables efficient fatty acid synthesis in oleaginous fungi. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rajala, A.; Wang, Y.; Brush, R.S.; Tsantilas, K.; Jankowski, C.S.R.; Lindsay, K.J.; Linton, J.D.; Hurley, J.B.; Anderson, R.E.; Rajala, R.V.S. Pyruvate kinase M2 regulates photoreceptor structure, function, and viability article. Cell Death Dis. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.; Wu, G.; Deshpande, R.R.; Vieler, A.; Gartner, K.; Li, X.; Moellering, E.R.; Zauner, S.; Cornish, A.J.; Liu, B.; et al. Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen Deprivation Predict Diversion of Metabolism. Plant Physiol. 2010, 154, 1737–1752. [Google Scholar] [CrossRef]
- Longworth, J.; Wu, D.; Huete-Ortega, M.; Wright, P.C.; Vaidyanathan, S. Proteome response of Phaeodactylum tricornutum, during lipid accumulation induced by nitrogen depletion. Algal Res. 2016, 18, 213–224. [Google Scholar] [CrossRef]
- Kooistra, W.H.C.F.; Gersonde, R.; Medlin, L.K.; Mann, D.G. The origin and evolution of the diatoms. Their adaptation to a planktonic existence. In Evolution of Primary Producers in the Sea; Elsevier Academic Press: Cambridge, MA, USA, 2007; pp. 207–249. [Google Scholar] [CrossRef]
- Tortell, P.D. Evolutionary and ecological perspectives on carbon acquisition in phytoplankton. Limnol. Oceanogr. 2000, 45, 744–750. [Google Scholar] [CrossRef]
- Schippers, P.; Lürling, M.; Scheffer, M. Increase of atmospheric CO2 promotes phytoplankton productivity. Ecol. Lett. 2004, 7, 446–451. [Google Scholar] [CrossRef]
- Gao, K.; Campbell, D.A. Photophysiological responses of marine diatoms to elevated CO2 and decreased pH: A review. Funct. Plant Biol. 2014, 41, 449–459. [Google Scholar] [CrossRef]
- Bowler, C.; Vardi, A.; Allen, A.E. Oceanographic and biogeochemical insights from diatom genomes. Ann. Rev. Mar. Sci. 2009, 2, 333–365. [Google Scholar] [CrossRef]
- Hennon, G.M.M.; Ashworth, J.; Groussman, R.D.; Berthiaume, C.; Morales, R.L.; Baliga, N.S.; Orellana, M.V.; Armbrust, E.V. Diatom acclimation to elevated CO2 via cAMP signalling and coordinated gene expression. Nat. Clim. Chang. 2015, 5, 761–766. [Google Scholar] [CrossRef]
- Benoiston, A.S.; Ibarbalz, F.M.; Bittner, L.; Guidi, L.; Jahn, O.; Dutkiewicz, S.; Bowler, C. The evolution of diatoms and their biogeochemical functions. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, J.J.; López García De Lomana, A.; Lee, A.; Armbrust, E.V.; Orellana, M.V.M.V.; Baliga, N.S. Ocean acidification conditions increase resilience of marine diatoms. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diatom Species | Growth Phase | CO2 Concentration (ppm) | Biomass Productivity (mg L−1 day−1) * | TAG Productivity (µgTAG L−1 day−1) * | Percentage of TAGs (DW) * |
|---|---|---|---|---|---|
| T. pseudonana | Exponential | 400 | 61.7 ± 7.0 | 11.8 ± 3 | 2.3 ± 1.0 |
| 20,000 | 68.6 ± 9.0 | 45.5 ± 26 | 5.3 ± 4.2 | ||
| Stationary | 400 | 45.3 ± 3.0 | 13.6 ± 2 | 5.5 ± 3.3 | |
| 20,000 | 55.8 ± 2.0 b↑ | 19.0 ± 5 | 7.2 ± 5.3 | ||
| P. tricornutum | Exponential | 400 | 28.5 ± 4.0 | 3.36 ± 1 | 0.67 ± 0.65 |
| 20,000 | 36.7 ± 6.0 | 8.47 ± 7 | 1.2 ± 1.4 | ||
| Stationary | 400 | 51.0 ± 4.0 | 50.2 ± 15 | 10.1 ± 3.6 | |
| 20,000 | 56.9 ± 1.0 | 75.7 ± 9 | 13.5 ± 1.3 | ||
| N. pelliculosa (seawater) | Exponential | 400 | 40.7 ± 4.0 | 0.03 ± 1 | - |
| 20,000 | 49.3 ± 3.0 a↑ | 1.09 ± 1 | 0.22 ± 0.2 | ||
| Stationary | 400 | 48.8 ± 5.0 | 117.4 ± 12 | 25.7 ± 3.0 | |
| 20,000 | 58.6 ± 1.0 b↑ | 158.4 ± 29 b↑ | 27.0 ± 0.8 | ||
| N. pelliculosa (freshwater) | Exponential | 400 | 42.8 ± 13.0 | 44.9 ± 10 | 11.5 ± 5.0 |
| 20,000 | 47.6 ± 12.0 | 55.8 ± 6 | 12.0 ± 1.8 | ||
| Stationary | 400 | 45.1 ± 8.0 | 142.3 ± 12 | 32.0 ± 5.0 | |
| 20,000 | 38.2 ± 1.0 | 97.25 ± 5 b↓ | 25.5 ± 0.7 | ||
| A. formosa | Exponential | 400 | 6.4 ± 1.0 | 1.43 ± 0.4 | 2.2 ± 0.3 |
| 20,000 | 5.6 ± 2.0 | 1.08 ± 1.7 | 1.4 ± 2.0 | ||
| Stationary | 400 | 2.3 ± 1.0 | 0.01 ± 0.003 | 0.06 ± 0.01 | |
| 20,000 | 2.5 ± 4.0 | 0.40 ± 0.8 | 1.7 ± 0.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jensen, E.L.; Yangüez, K.; Carrière, F.; Gontero, B. Storage Compound Accumulation in Diatoms as Response to Elevated CO2 Concentration. Biology 2020, 9, 5. https://doi.org/10.3390/biology9010005
Jensen EL, Yangüez K, Carrière F, Gontero B. Storage Compound Accumulation in Diatoms as Response to Elevated CO2 Concentration. Biology. 2020; 9(1):5. https://doi.org/10.3390/biology9010005
Chicago/Turabian StyleJensen, Erik L., Karen Yangüez, Frédéric Carrière, and Brigitte Gontero. 2020. "Storage Compound Accumulation in Diatoms as Response to Elevated CO2 Concentration" Biology 9, no. 1: 5. https://doi.org/10.3390/biology9010005
APA StyleJensen, E. L., Yangüez, K., Carrière, F., & Gontero, B. (2020). Storage Compound Accumulation in Diatoms as Response to Elevated CO2 Concentration. Biology, 9(1), 5. https://doi.org/10.3390/biology9010005