Role of Rhodopsins as Circadian Photoreceptors in the Drosophila melanogaster

 , ,
, ,
Abstract
1. Introduction
2. The Role of the Eyes in Circadian Entrainment and in the Direct Light Effects on Activity
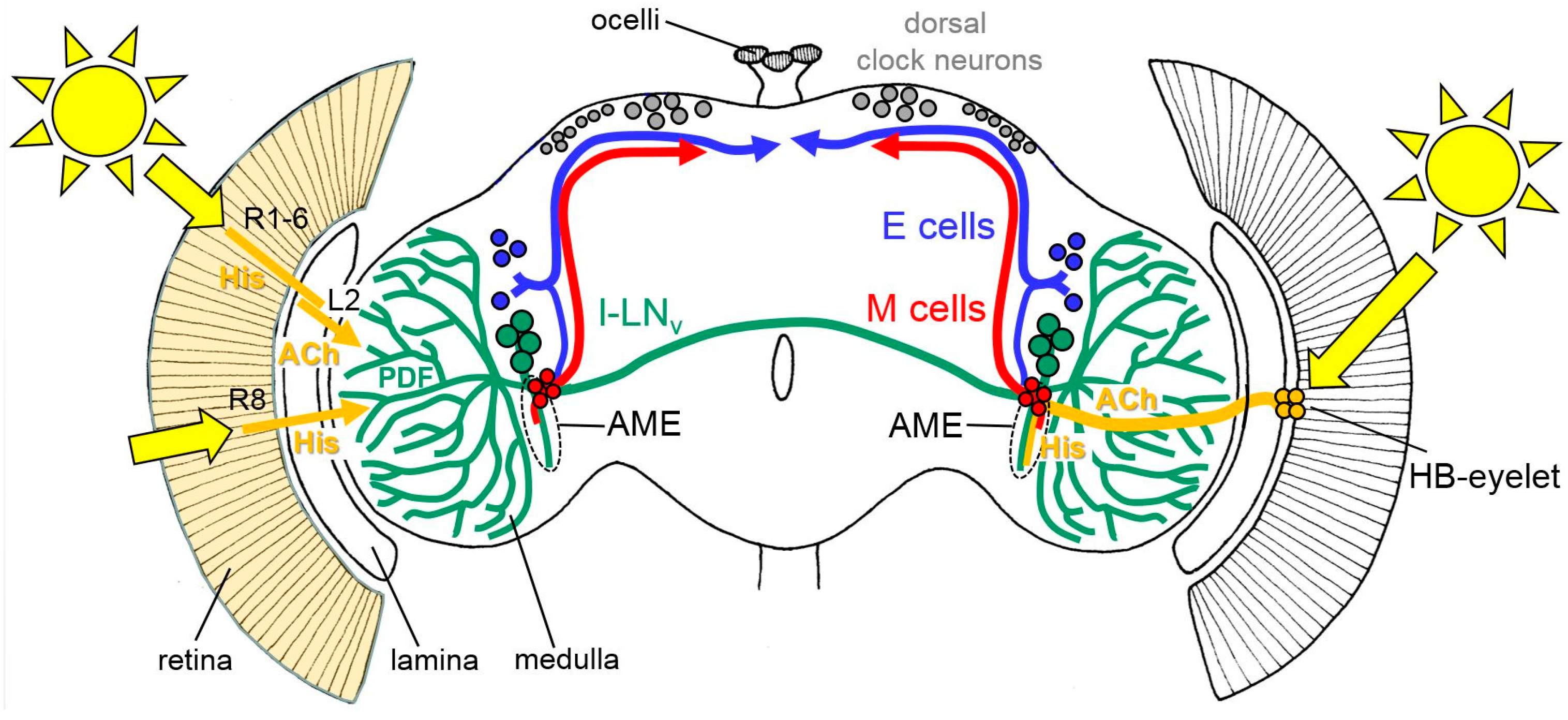
3. Input Pathways from the Eyes to the Clock Neurons
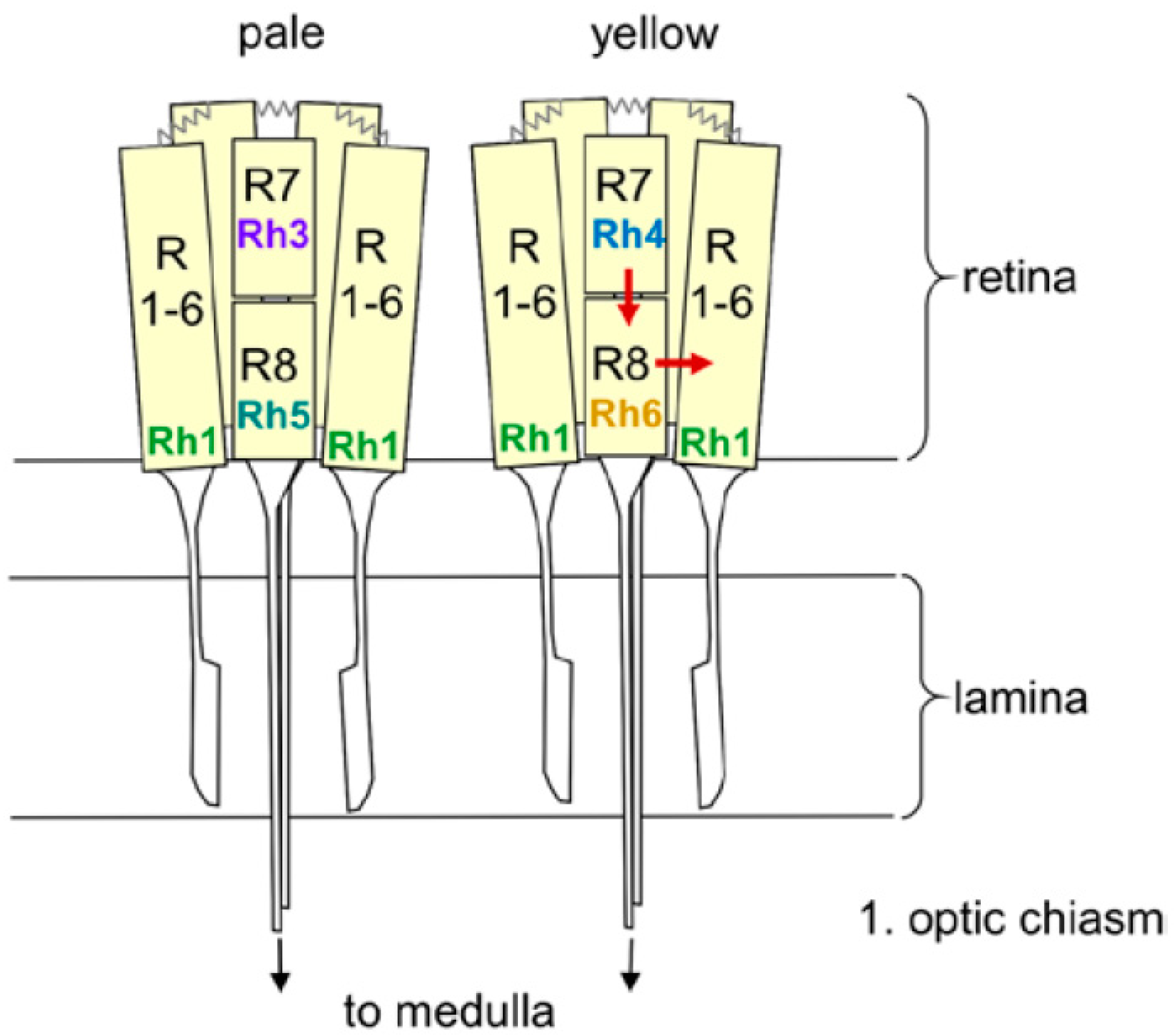
4. Roles of Rhodopsins 1 to 6 in Entrainment and Direct Light Effects
5. Diverging Hypotheses Concerning the Role of Rh7
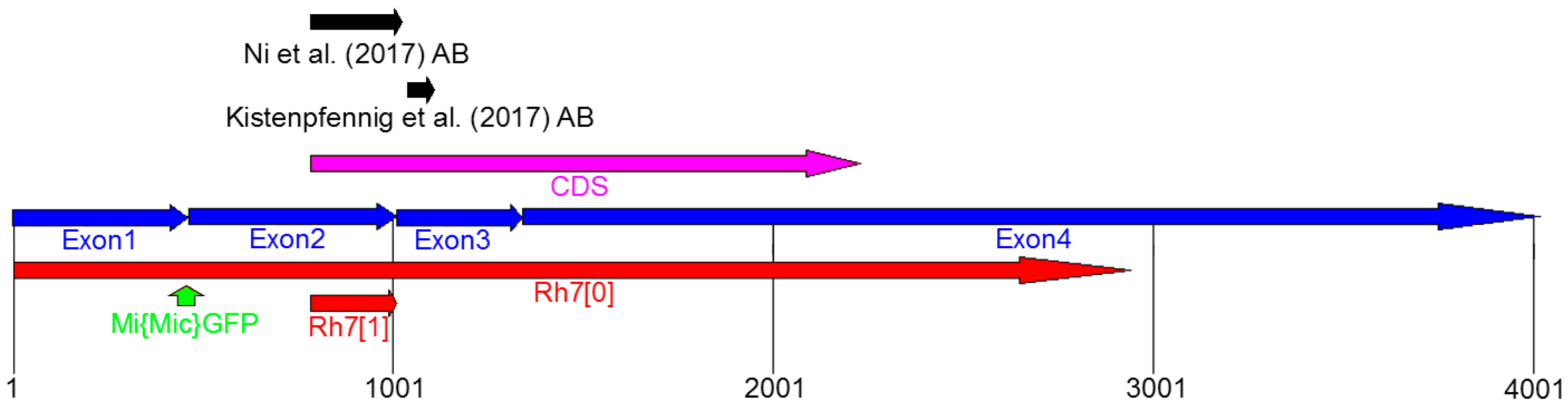
5.1. Testing the Function of Rh7 as a Photopigment
5.2. Place of the Rh7 Action
5.3. Effects of the Rh7 on the Entrainment of the Circadian Clock
6. New Results Reveal a Role of the Rh7 in Compound Eyes
6.1. Materials and Methods
6.2. Results
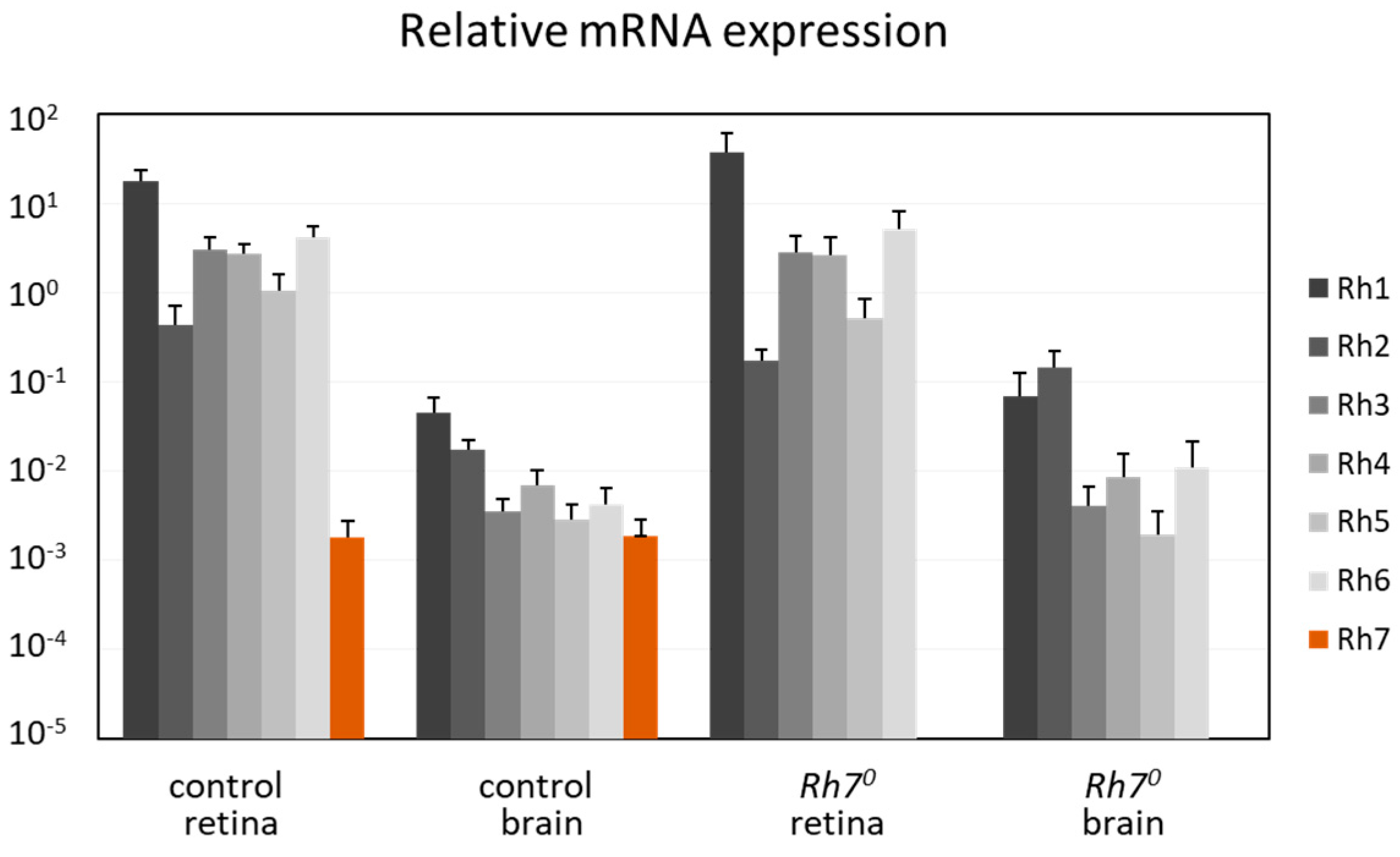
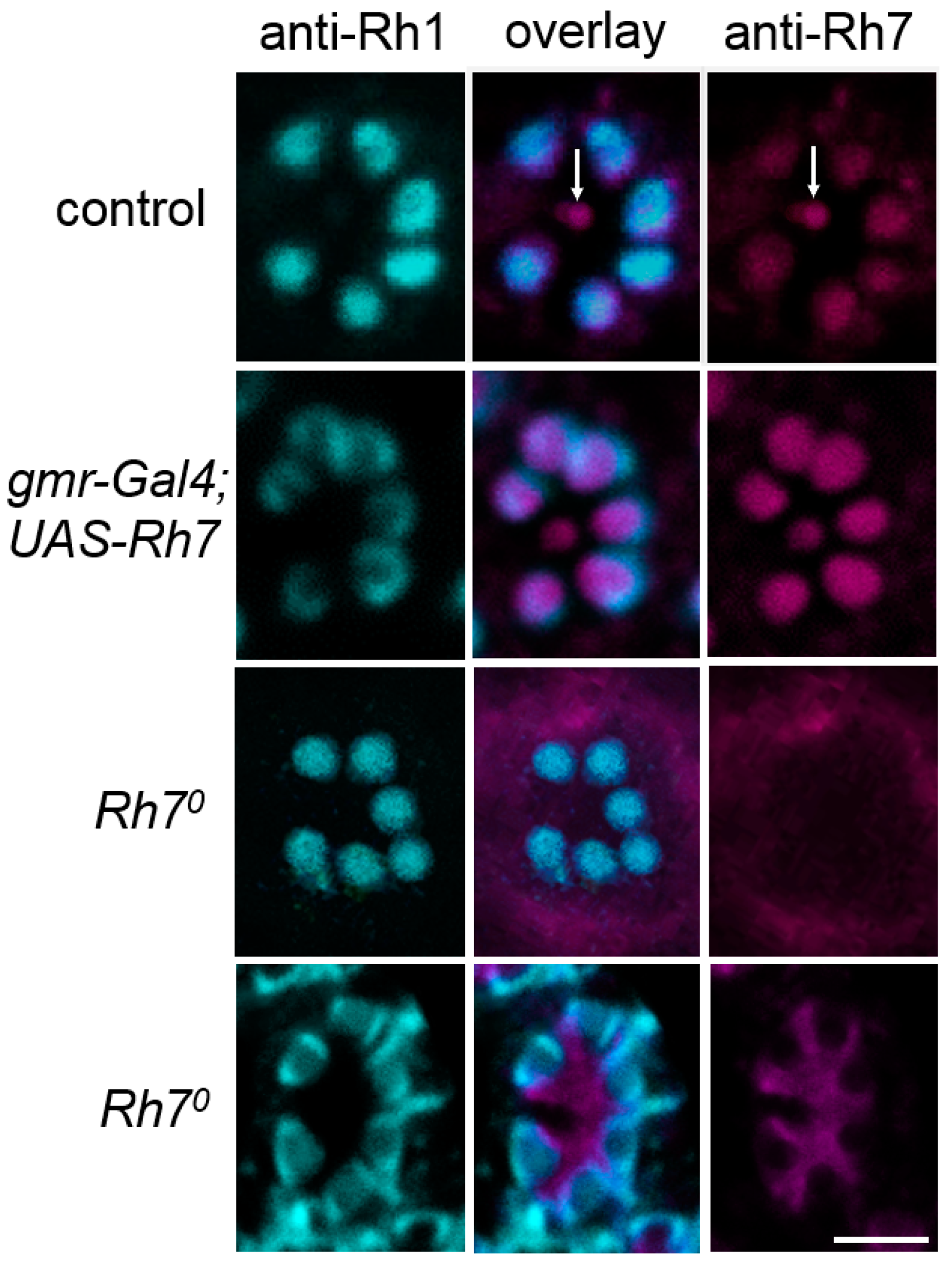
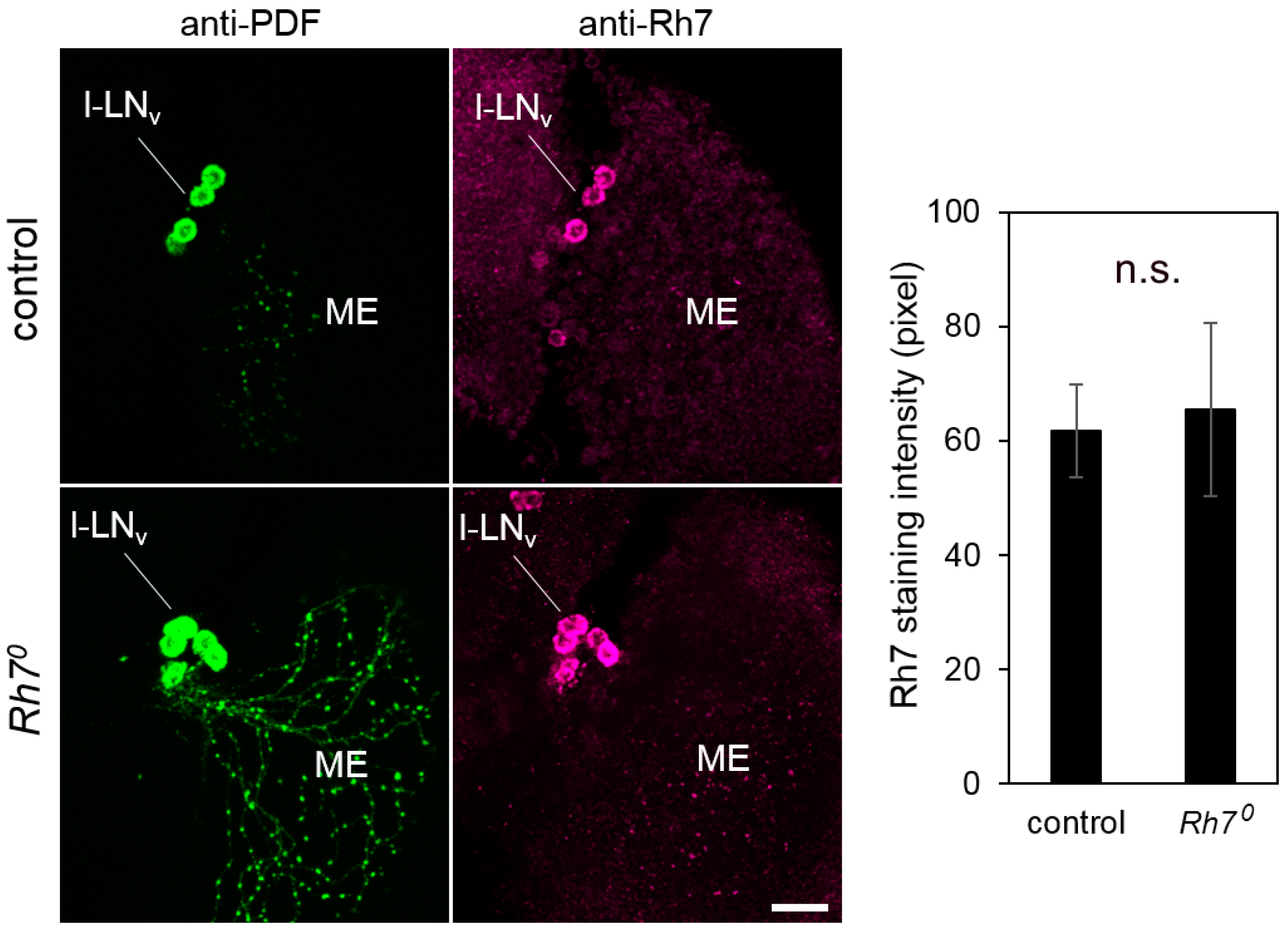
6.2.1. Rh7 is Expressed in the Compound Eyes and in the Brain, but Not in the l-LNv
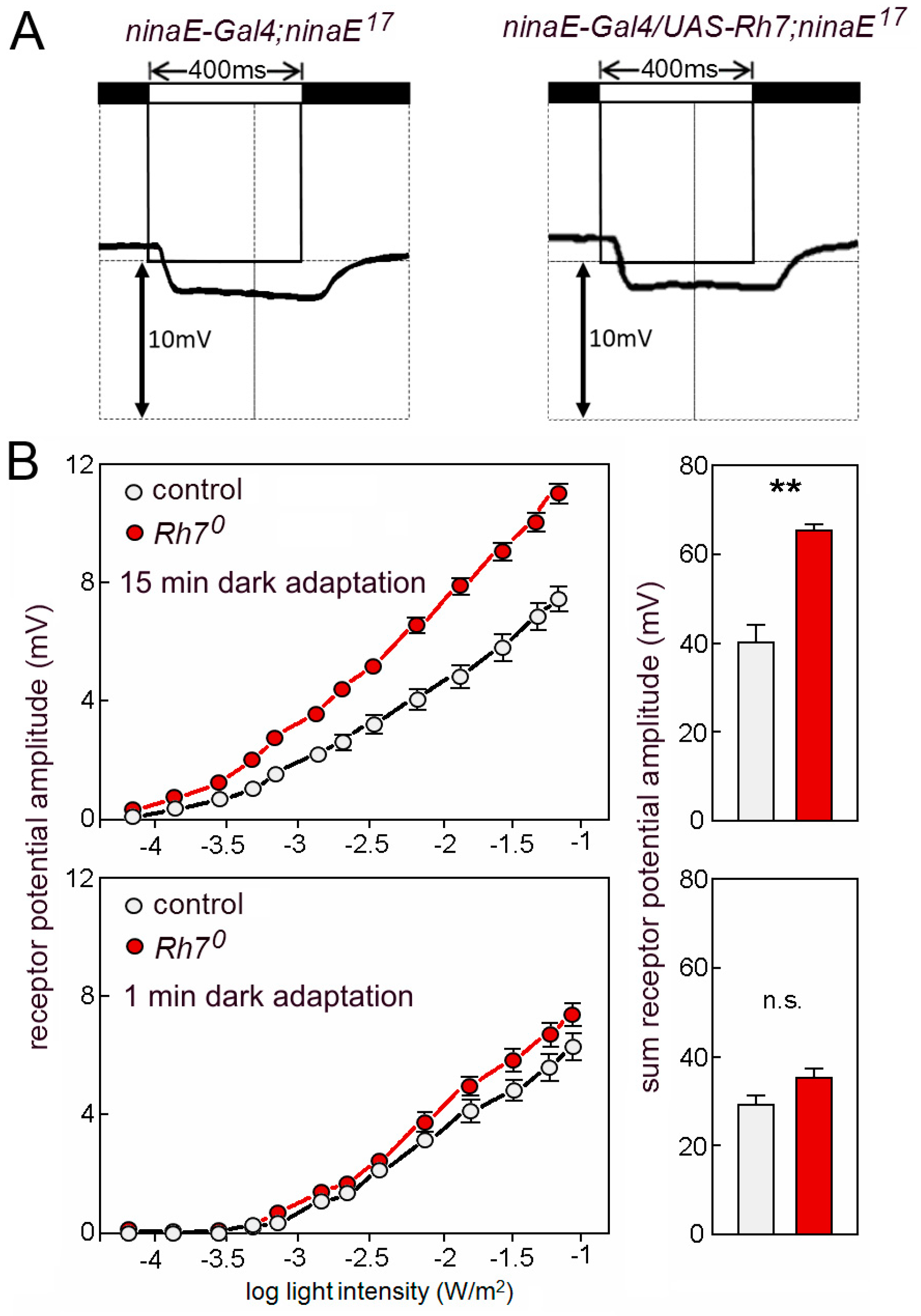
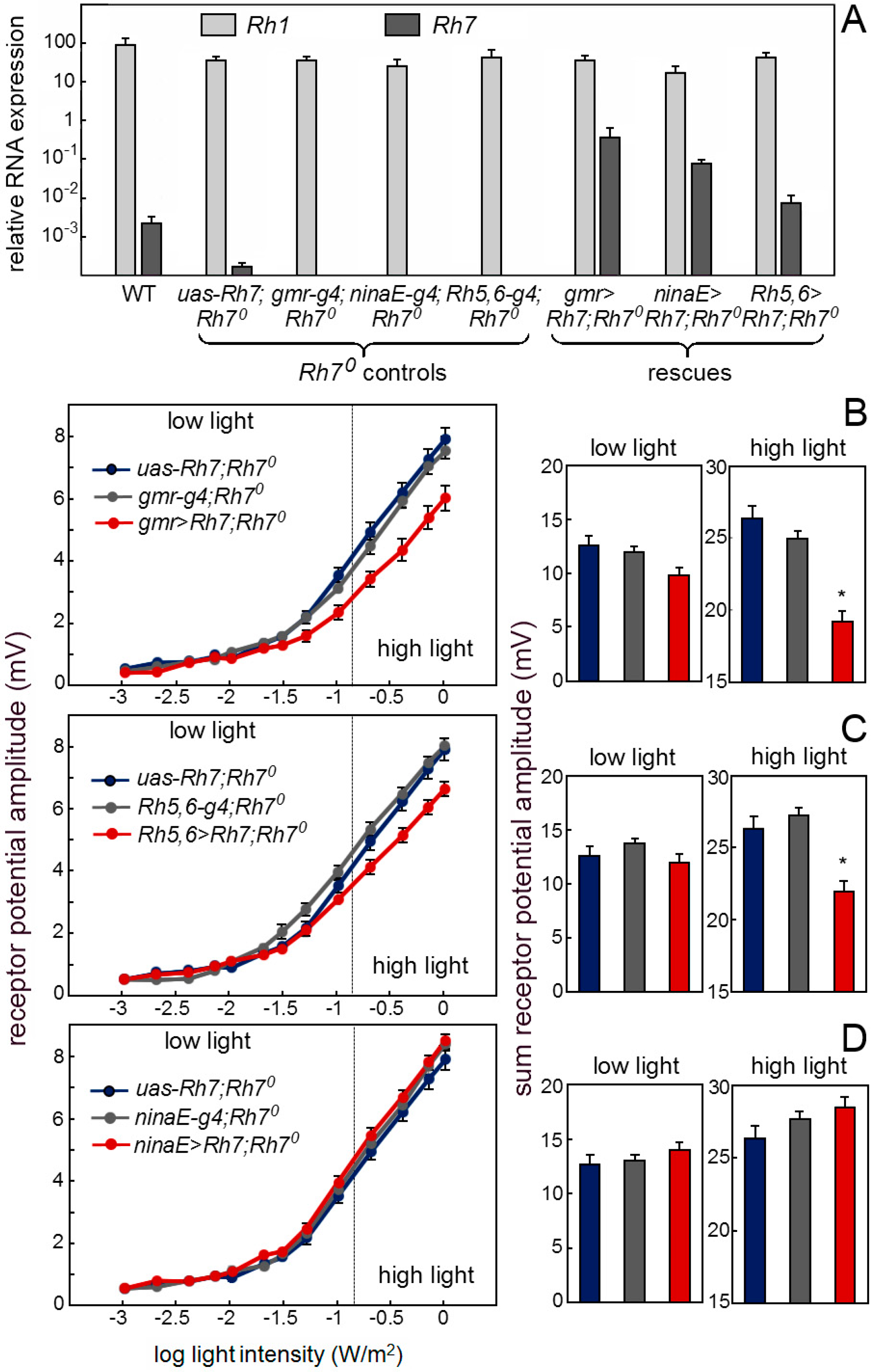
6.2.2. Rh7 Reduces the Magnitude of the ERG
6.3. Discussion
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roenneberg, T.; Foster, R.G. Twilight times: Light and the circadian system. Photochem. Photobiol. 1997, 66, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.G.; Helfrich-Förster, C. The regulation of circadian clocks by light in fruitflies and mice. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 1779–1789. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.G.; Provencio, I.; Hudson, D.; Fiske, S.; De Grip, W.; Menaker, M. Circadian photoreception in the retinally degenerate mouse (rd/rd). J. Comp. Physiol. A 1991, 169, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Provencio, I.; Rodriguez, I.R.; Jiang, G.; Hayes, W.P.; Moreira, E.F.; Rollag, M.D. A novel human opsin in the inner retina. J. Neurosci. Off. J. Soc. Neurosci. 2000, 20, 600–605. [Google Scholar] [CrossRef]
- Berson, E.L.; Rosner, B.; Weigel-DiFranco, C.; Dryja, T.P.; Sandberg, M.A. Disease progression in patients with dominant retinitis pigmentosa and rhodopsin mutations. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3027–3036. [Google Scholar]
- Hattar, S.; Liao, H.W.; Takao, M.; Berson, D.M.; Yau, K.W. Melanopsin-containing retinal ganglion cells: Architecture, projections, and intrinsic photosensitivity. Science 2002, 295, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhong, H.; Wang, M.-H.H.; Luo, D.-G.; Liao, H.-W.; Maeda, H.; Hattar, S.; Frishman, L.J.; Yau, K.-W. Intrinsically photosensitive retinal ganglion cells detect light with a vitamin A-based photopigment, melanopsin. Proc. Natl. Acad. Sci. USA 2005, 102, 10339–10344. [Google Scholar] [CrossRef]
- Hattar, S.; Lucas, R.J.; Mrosovsky, N.; Thompson, S.; Douglas, R.H.; Hankins, M.W.; Lem, J.; Biel, M.; Hofmann, F.; Foster, R.G.; et al. Melanopsin and rod-cone photoreceptive systems account for all major accessory visual functions in mice. Nature 2003, 424, 76–81. [Google Scholar] [CrossRef]
- Ota, W.; Nakane, Y.; Hattar, S.; Yoshimura, T. Impaired Circadian Photoentrainment in Opn5-Null Mice. Science 2018, 6, 299–305. [Google Scholar] [CrossRef]
- Rieger, D.; Stanewsky, R.; Helfrich-Förster, C. Cryptochrome, compound eyes, Hofbauer-Buchner eyelets, and ocelli play different roles in the entrainment and masking pathway of the locomotor activity rhythm in the fruit fly Drosophila melanogaster. J. Biol. Rhythms 2003, 18, 377–391. [Google Scholar] [CrossRef]
- Yoshii, T.; Hermann-Luibl, C.; Helfrich-Förster, C. Circadian light-input pathways in Drosophila. Commun. Integr. Biol. 2016, 9, e1102805. [Google Scholar] [CrossRef] [PubMed]
- Egan, E.S.; Franklin, T.M.; Hilderbrand-Chae, M.J.; McNeil, G.P.; Roberts, M.A.; Schroeder, A.J.; Zhang, X.; Jackson, F.R. An extraretinally expressed insect cryptochrome with similarity to the blue light photoreceptors of mammals and plants. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 3665–3673. [Google Scholar] [CrossRef]
- Emery, P.; So, W.V.; Kaneko, M.; Hall, J.C.; Rosbash, M. CRY, a Drosophila clock and light-regulated cryptochrome, is a major contributor to circadian rhythm resetting and photosensitivity. Cell 1998, 95, 669–679. [Google Scholar] [CrossRef]
- Emery, P.; Stanewsky, R.; Hall, J.C.; Rosbash, M. A unique circadian-rhythm photoreceptor. Nature 2000, 404, 456–457. [Google Scholar] [CrossRef] [PubMed]
- Emery, P.; Stanewsky, R.; Helfrich-Förster, C.; Emery-Le, M.; Hall, J.C.; Rosbash, M. Drosophila CRY is a deep brain circadian photoreceptor. Neuron 2000, 26, 493–504. [Google Scholar] [CrossRef]
- Helfrich-Förster, C.; Winter, C.; Hofbauer, A.; Hall, J.C.; Stanewsky, R. The circadian clock of fruit flies is blind after elimination of all known photoreceptors. Neuron 2001, 30, 249–261. [Google Scholar] [CrossRef]
- Senthilan, P.R.; Helfrich-Förster, C. Rhodopsin 7-The unusual Rhodopsin in Drosophila. PeerJ 2016, 4, e2427. [Google Scholar] [CrossRef]
- Grebler, R.; Kistenpfennig, C.; Rieger, D.; Bentrop, J.; Schneuwly, S.; Senthilan, P.R.; Helfrich-Förster, C. Drosophila Rhodopsin 7 can partially replace the structural role of Rhodopsin 1, but not its physiological function. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2017, 203, 649–659. [Google Scholar] [CrossRef]
- Kistenpfennig, C.; Grebler, R.; Ogueta, M.; Hermann-Luibl, C.; Schlichting, M.; Stanewsky, R.; Senthilan, P.R.; Helfrich-Förster, C. A New Rhodopsin Influences Light-dependent Daily Activity Patterns of Fruit Flies. J. Biol. Rhythms 2017, 32, 406–422. [Google Scholar] [CrossRef]
- Ni, J.D.; Baik, L.S.; Holmes, T.C.; Montell, C. A rhodopsin in the brain functions in circadian photoentrainment in Drosophila. Nature 2017, 545, 340–344. [Google Scholar] [CrossRef]
- Mrosovsky, N. Further experiments on the relationship between the period of circadian rhythms and locomotor activity levels in hamsters. Physiol. Behav. 1999, 66, 797–801. [Google Scholar] [CrossRef]
- Wheeler, D.A.; Hamblen-Coyle, M.J.; Dushay, M.S.; Hall, J.C. Behavior in light-dark cycles of Drosophila mutants that are arrhythmic, blind, or both. J. Biol. Rhythms 1993, 8, 67–94. [Google Scholar] [CrossRef] [PubMed]
- Redlin, U.; Mrosovsky, N. Masking by light in hamsters with SCN lesions. J. Comp. Physiol. A 1999, 184, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Hamblen-Coyle, M.J.; Wheeler, D.A.; Rutila, J.E.; Rosbash, M.; Hall, J.C. Behavior of period-altered circadian rhythm mutants of Drosophila in light: Dark cycles (Diptera: Drosophilidae). J. Insect Behav. 1992, 5, 417–446. [Google Scholar] [CrossRef]
- Vanin, S.; Bhutani, S.; Montelli, S.; Menegazzi, P.; Green, E.W.; Pegoraro, M.; Sandrelli, F.; Costa, R.; Kyriacou, C.P. Unexpected features of Drosophila circadian behavioural rhythms under natural conditions. Nature 2012, 484, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Menegazzi, P.; Yoshii, T.; Helfrich-Förster, C. Laboratory versus nature: The two sides of the Drosophila circadian clock. J. Biol. Rhythms 2012, 27, 433–442. [Google Scholar] [CrossRef]
- Rieger, D.; Fraunholz, C.; Popp, J.; Bichler, D.; Dittmann, R.; Helfrich-Förster, C. The fruit fly Drosophila melanogaster favors dim light and times its activity peaks to early dawn and late dusk. J. Biol. Rhythms 2007, 22, 387–399. [Google Scholar] [CrossRef]
- Schlichting, M.; Grebler, R.; Menegazzi, P.; Helfrich-Förster, C. Twilight dominates over moonlight in adjusting Drosophila’s activity pattern. J. Biol. Rhythms 2015, 30, 117–128. [Google Scholar] [CrossRef]
- Schlichting, M.; Menegazzi, P.; Helfrich-Förster, C. Normal vision can compensate for the loss of the circadian clock. Proc. Biol. Sci. 2015, 282. [Google Scholar] [CrossRef]
- Bachleitner, W.; Kempinger, L.; Wülbeck, C.; Rieger, D.; Helfrich-Förster, C. Moonlight shifts the endogenous clock of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2007, 104, 3538–3543. [Google Scholar] [CrossRef]
- Kempinger, L.; Dittmann, R.; Rieger, D.; Helfrich-Förster, C. The nocturnal activity of fruit flies exposed to artificial moonlight is partly caused by direct light effects on the activity level that bypass the endogenous clock. Chronobiol. Int. 2009, 26, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Saint-Charles, A.; Michard-Vanhée, C.; Alejevski, F.; Chélot, E.; Boivin, A.; Rouyer, F. Four of the six Drosophila rhodopsin-expressing photoreceptors can mediate circadian entrainment in low light. J. Comp. Neurol. 2016, 524, 2828–2844. [Google Scholar] [CrossRef] [PubMed]
- Kistenpfennig, C.; Nakayama, M.; Nihara, R.; Tomioka, K.; Helfrich-Förster, C.; Yoshii, T. A Tug-of-War between Cryptochrome and the Visual System Allows the Adaptation of Evening Activity to Long Photoperiods in Drosophila melanogaster. J. Biol. Rhythms 2018, 33, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Wardill, T.J.; List, O.; Li, X.; Dongre, S.; McCulloch, M.; Ting, C.-Y.; O’Kane, C.J.; Tang, S.; Lee, C.-H.; Hardie, R.C.; et al. Multiple spectral inputs improve motion discrimination in the Drosophila visual system. Science 2012, 336, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Schnaitmann, C.; Haikala, V.; Abraham, E.; Oberhauser, V.; Thestrup, T.; Griesbeck, O.; Reiff, D.F. Color Processing in the Early Visual System of Drosophila. Cell 2018, 172, 318–330.e18. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, A.; Buchner, E. Does Drosophila have seven eyes? Naturwissenschaften 1989, 76, 335–336. [Google Scholar] [CrossRef]
- Helfrich-Förster, C.; Edwards, T.; Yasuyama, K.; Wisotzki, B.; Schneuwly, S.; Stanewsky, R.; Meinertzhagen, I.A.; Hofbauer, A. The extraretinal eyelet of Drosophila: Development, ultrastructure, and putative circadian function. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 9255–9266. [Google Scholar] [CrossRef]
- Malpel, S.; Klarsfeld, A.; Rouyer, F. Larval optic nerve and adult extra-retinal photoreceptors sequentially associate with clock neurons during Drosophila brain development. Dev. Camb. Engl. 2002, 129, 1443–1453. [Google Scholar]
- Li, M.-T.; Cao, L.-H.; Xiao, N.; Tang, M.; Deng, B.; Yang, T.; Yoshii, T.; Luo, D.-G. Hub-organized parallel circuits of central circadian pacemaker neurons for visual photoentrainment in Drosophila. Nat. Commun. 2018, 9, 4247. [Google Scholar] [CrossRef]
- Schlichting, M.; Grebler, R.; Peschel, N.; Yoshii, T.; Helfrich-Förster, C. Moonlight detection by Drosophila’s endogenous clock depends on multiple photopigments in the compound eyes. J. Biol. Rhythms 2014, 29, 75–86. [Google Scholar] [CrossRef]
- Schubert, F.K.; Hagedorn, N.; Yoshii, T.; Helfrich-Förster, C.; Rieger, D. Neuroanatomical details of the lateral neurons of Drosophila melanogaster support their functional role in the circadian system. J. Comp. Neurol. 2018, 526, 1209–1231. [Google Scholar] [CrossRef] [PubMed]
- Helfrich-Förster, C. The Drosophila clock system. In Biological Timekeeping: Clocks, Rhythms and Behaviour; Springer: New Delhi, India; pp. 133–176.
- Parisky, K.M.; Agosto, J.; Pulver, S.R.; Shang, Y.; Kuklin, E.; Hodge, J.J.L.; Kang, K.; Kang, K.; Liu, X.; Garrity, P.A.; et al. PDF cells are a GABA-responsive wake-promoting component of the Drosophila sleep circuit. Neuron 2008, 60, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Griffith, L.C.; Rosbash, M. Light-arousal and circadian photoreception circuits intersect at the large PDF cells of the Drosophila brain. Proc. Natl. Acad. Sci. USA 2008, 105, 19587–19594. [Google Scholar] [CrossRef]
- Sheeba, V.; Fogle, K.J.; Kaneko, M.; Rashid, S.; Chou, Y.-T.; Sharma, V.K.; Holmes, T.C. Large ventral lateral neurons modulate arousal and sleep in Drosophila. Curr. Biol. CB 2008, 18, 1537–1545. [Google Scholar] [CrossRef]
- Sheeba, V.; Gu, H.; Sharma, V.K.; O’Dowd, D.K.; Holmes, T.C. Circadian- and light-dependent regulation of resting membrane potential and spontaneous action potential firing of Drosophila circadian pacemaker neurons. J. Neurophysiol. 2008, 99, 976–988. [Google Scholar] [CrossRef] [PubMed]
- Muraro, N.I.; Ceriani, M.F. Acetylcholine from Visual Circuits Modulates the Activity of Arousal Neurons in Drosophila. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 16315–16327. [Google Scholar] [CrossRef]
- Schlichting, M.; Menegazzi, P.; Lelito, K.R.; Yao, Z.; Buhl, E.; Dalla Benetta, E.; Bahle, A.; Denike, J.; Hodge, J.J.; Helfrich-Förster, C.; et al. A Neural Network Underlying Circadian Entrainment and Photoperiodic Adjustment of Sleep and Activity in Drosophila. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 9084–9096. [Google Scholar] [CrossRef]
- Liang, X.; Holy, T.E.; Taghert, P.H. Synchronous Drosophila circadian pacemakers display nonsynchronous Ca2+ rhythms in vivo. Science 2016, 351, 976–981. [Google Scholar] [CrossRef]
- Menegazzi, P.; Dalla Benetta, E.; Beauchamp, M.; Schlichting, M.; Steffan-Dewenter, I.; Helfrich-Förster, C. Adaptation of Circadian Neuronal Network to Photoperiod in High-Latitude European Drosophilids. Curr. Biol. CB 2017, 27, 833–839. [Google Scholar] [CrossRef]
- Eck, S.; Helfrich-Förster, C.; Rieger, D. The Timed Depolarization of Morning and Evening Oscillators Phase Shifts the Circadian Clock of Drosophila. J. Biol. Rhythms 2016, 31, 428–442. [Google Scholar] [CrossRef]
- Yoshii, T.; Hermann-Luibl, C.; Kistenpfennig, C.; Schmid, B.; Tomioka, K.; Helfrich-Förster, C. Cryptochrome-dependent and -independent circadian entrainment circuits in Drosophila. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 6131–6141. [Google Scholar] [CrossRef] [PubMed]
- Alejevski, F.; Saint-Charles, A.; Michard-Vanhée, C.; Martin, B.; Galant, S.; Vasiliauskas, D.; Rouyer, F. The HisCl1 histamine receptor acts in photoreceptors to synchronize Drosophila behavioral rhythms with light-dark cycles. Nat. Commun. 2018, in press. [Google Scholar]
- Rister, J.; Desplan, C.; Vasiliauskas, D. Establishing and maintaining gene expression patterns: Insights from sensory receptor patterning. Dev. Camb. Engl. 2013, 140, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Schlichting, M.; Menegazzi, P.; Rosbash, M.; Helfrich-Förster, C. A distinct visual pathway mediates high light intensity adaptation of the circadian clock in Drosophila. J. Neurosci. 2018. under revision. [Google Scholar] [CrossRef]
- Provencio, I.; Rollag, M.D.; Castrucci, A.M. Photoreceptive net in the mammalian retina. This mesh of cells may explain how some blind mice can still tell day from night. Nature 2002, 415, 493. [Google Scholar] [CrossRef]
- Walmsley, L.; Hanna, L.; Mouland, J.; Martial, F.; West, A.; Smedley, A.R.; Bechtold, D.A.; Webb, A.R.; Lucas, R.J.; Brown, T.M. Colour as a signal for entraining the mammalian circadian clock. PLoS Biol. 2015, 13, e1002127. [Google Scholar] [CrossRef] [PubMed]
- Hanai, S.; Hamasaka, Y.; Ishida, N. Circadian entrainment to red light in Drosophila: Requirement of Rhodopsin 1 and Rhodopsin 6. Neuroreport 2008, 19, 1441–1444. [Google Scholar] [CrossRef]
- Hanai, S.; Ishida, N. Entrainment of Drosophila circadian clock to green and yellow light by Rh1, Rh5, Rh6 and CRY. Neuroreport 2009, 20, 755–758. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, E.; Huber, A.; Henrich, S.; Chadwell, L.V.; Chou, W.H.; Paulsen, R.; Britt, S.G. Blue- and green-absorbing visual pigments of Drosophila: Ectopic expression and physiological characterization of the R8 photoreceptor cell-specific Rh5 and Rh6 rhodopsins. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 10716–10726. [Google Scholar] [CrossRef]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef]
- Rister, J.; Razzaq, A.; Boodram, P.; Desai, N.; Tsanis, C.; Chen, H.; Jukam, D.; Desplan, C. Single-base pair differences in a shared motif determine differential Rhodopsin expression. Science 2015, 350, 1258–1261. [Google Scholar] [CrossRef] [PubMed]
- Abruzzi, K.C.; Zadina, A.; Luo, W.; Wiyanto, E.; Rahman, R.; Guo, F.; Shafer, O.; Rosbash, M. RNA-seq analysis of Drosophila clock and non-clock neurons reveals neuron-specific cycling and novel candidate neuropeptides. PLoS Genet. 2017, 13, e1006613. [Google Scholar] [CrossRef] [PubMed]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Dev. Camb. Engl. 1993, 118, 401–415. [Google Scholar]
- Harris, W.A.; Stark, W.S.; Walker, J.A. Genetic dissection of the photoreceptor system in the compound eye of Drosophila melanogaster. J. Physiol. 1976, 256, 415–439. [Google Scholar] [PubMed]
- White, J.F.; Grodnitzky, J.; Louis, J.M.; Trinh, L.B.; Shiloach, J.; Gutierrez, J.; Northup, J.K.; Grisshammer, R. Dimerization of the class A G protein-coupled neurotensin receptor NTS1 alters G protein interaction. Proc. Natl. Acad. Sci. USA 2007, 104, 12199–12204. [Google Scholar] [CrossRef] [PubMed]
- Bayburt, T.H.; Leitz, A.J.; Xie, G.; Oprian, D.D.; Sligar, S.G. Transducin activation by nanoscale lipid bilayers containing one and two rhodopsins. J. Biol. Chem. 2007, 282, 14875–14881. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | 5′-Primer (5′-3′) | 3′-Primer (5′-3′) |
|---|---|---|
| Rh1 Rh2 Rh3 Rh4 Rh5 Rh6 Rh7 alpha-tubulin | TGCCTACATCTGGTTCATGTCGAGC GCCATCCCAAGTACCGCATAGTTCTCA TGCGAACTCCCTCCAATATACTGGTC GAACCGCAACATGACCTTCACCAAG TTCTCCGTGCTGCCATTGTTCCAG ATATCGCTGTGGTTCTTCGCCTG CATCTGCGACTTTCTGATGCTCATC TCTGCGATTCGATGGTGCCCTTAAC | GGAGTAGAAGATCAGGTATGAGCGTG CACGATTTAAGCCTTTGAATCCGCCTC TATAAACGCATTGGTGGCACCGGC CAAACAACCGGGTGTCAAAGTTGTCC GTGAAGAGCTTGTAGTAGGACACCAGG CTGCTTGTACTTCGGGTGGCTC GGATGCACCACCACATTGTACCGATC GGATCGCACTTGACCATCTGGTTGGC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senthilan, P.R.; Grebler, R.; Reinhard, N.; Rieger, D.; Helfrich-Förster, C. Role of Rhodopsins as Circadian Photoreceptors in the Drosophila melanogaster. Biology 2019, 8, 6. https://doi.org/10.3390/biology8010006
Senthilan PR, Grebler R, Reinhard N, Rieger D, Helfrich-Förster C. Role of Rhodopsins as Circadian Photoreceptors in the Drosophila melanogaster. Biology. 2019; 8(1):6. https://doi.org/10.3390/biology8010006
Chicago/Turabian StyleSenthilan, Pingkalai R., Rudi Grebler, Nils Reinhard, Dirk Rieger, and Charlotte Helfrich-Förster. 2019. "Role of Rhodopsins as Circadian Photoreceptors in the Drosophila melanogaster" Biology 8, no. 1: 6. https://doi.org/10.3390/biology8010006
APA StyleSenthilan, P. R., Grebler, R., Reinhard, N., Rieger, D., & Helfrich-Förster, C. (2019). Role of Rhodopsins as Circadian Photoreceptors in the Drosophila melanogaster. Biology, 8(1), 6. https://doi.org/10.3390/biology8010006