Sources and Fates of Carbamyl Phosphate: A Labile Energy-Rich Molecule with Multiple Facets



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
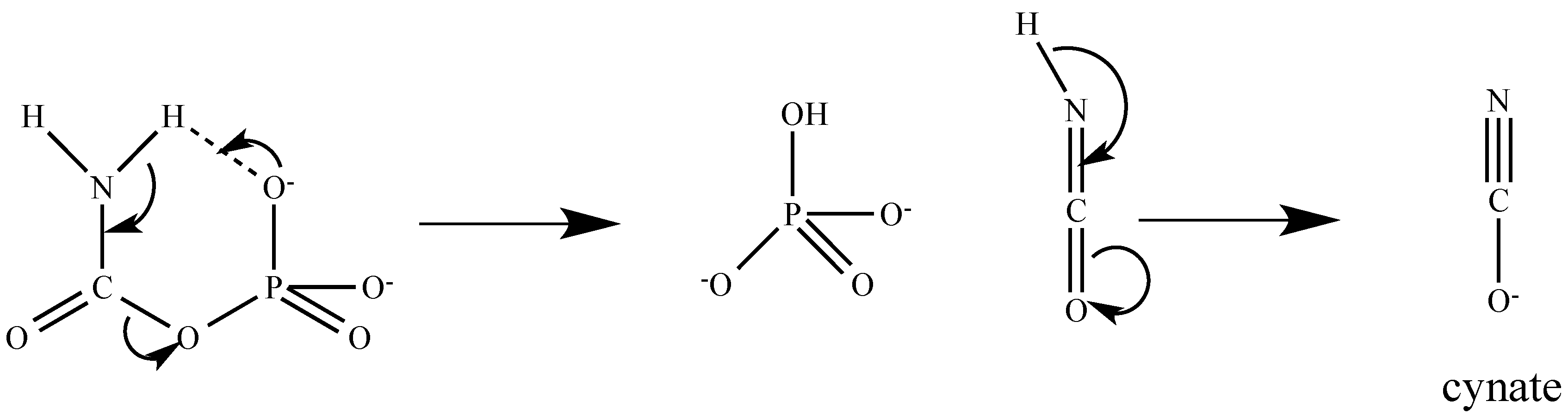
2. Production of CP Chemically
3. Production of CP in Biological Systems
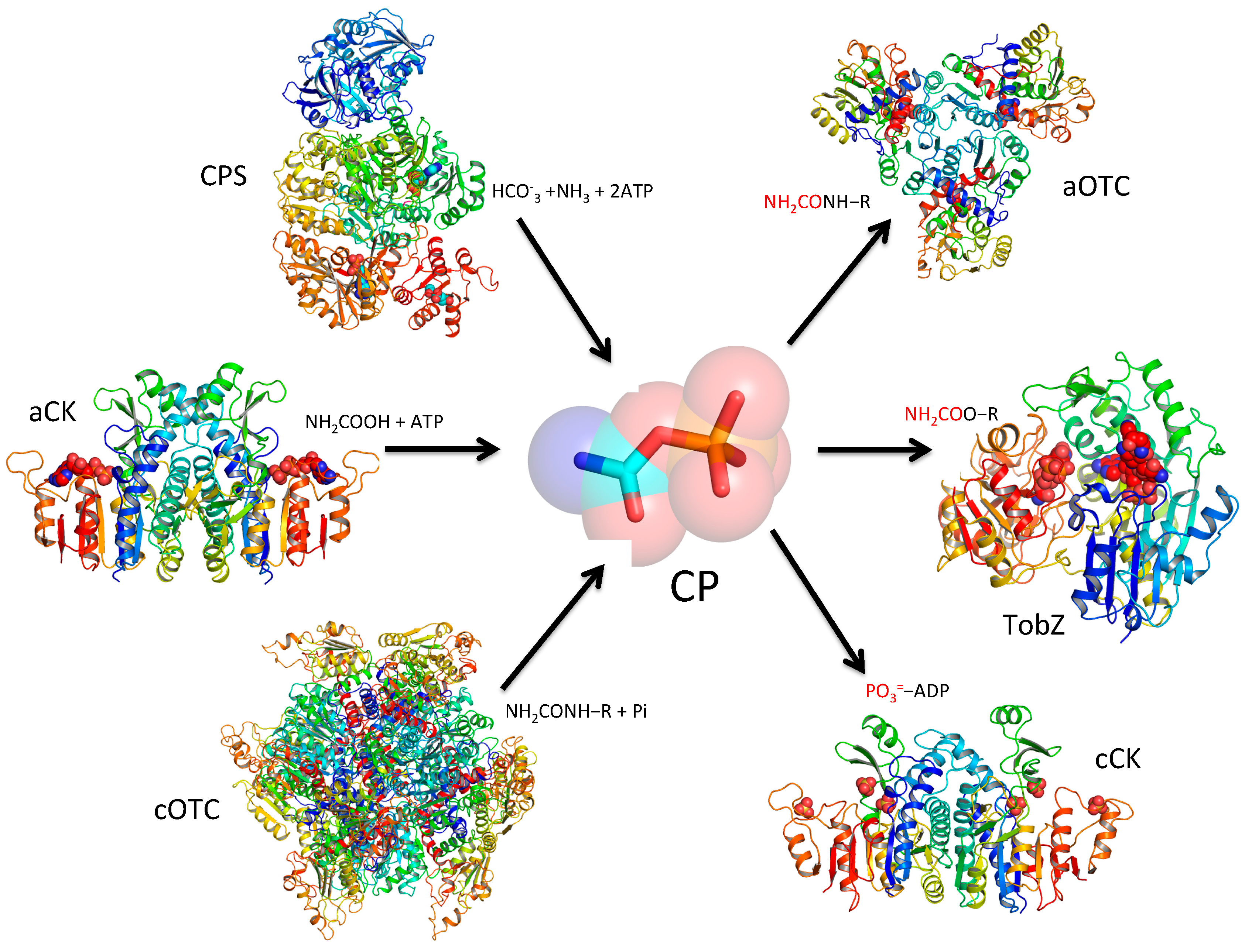
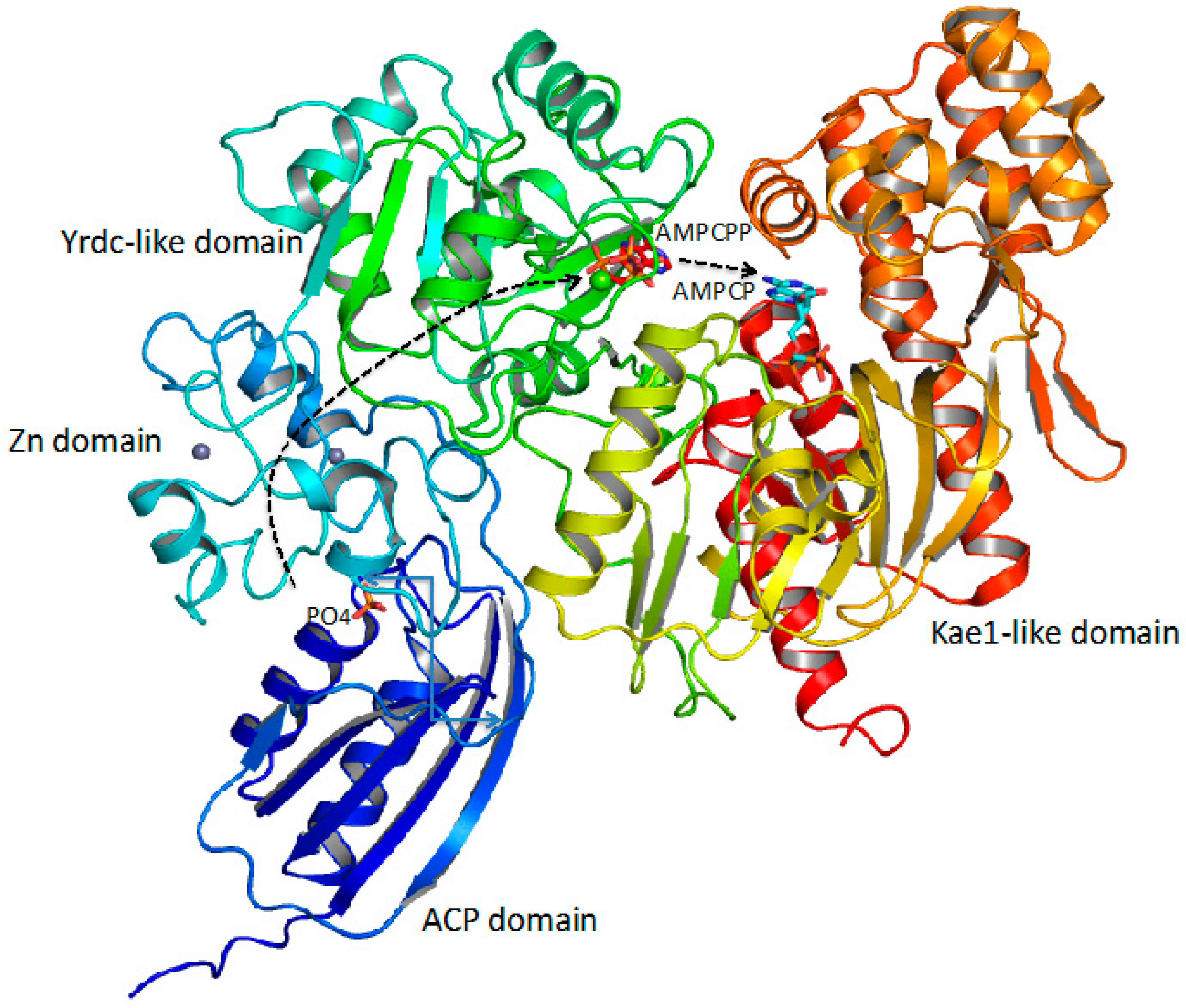
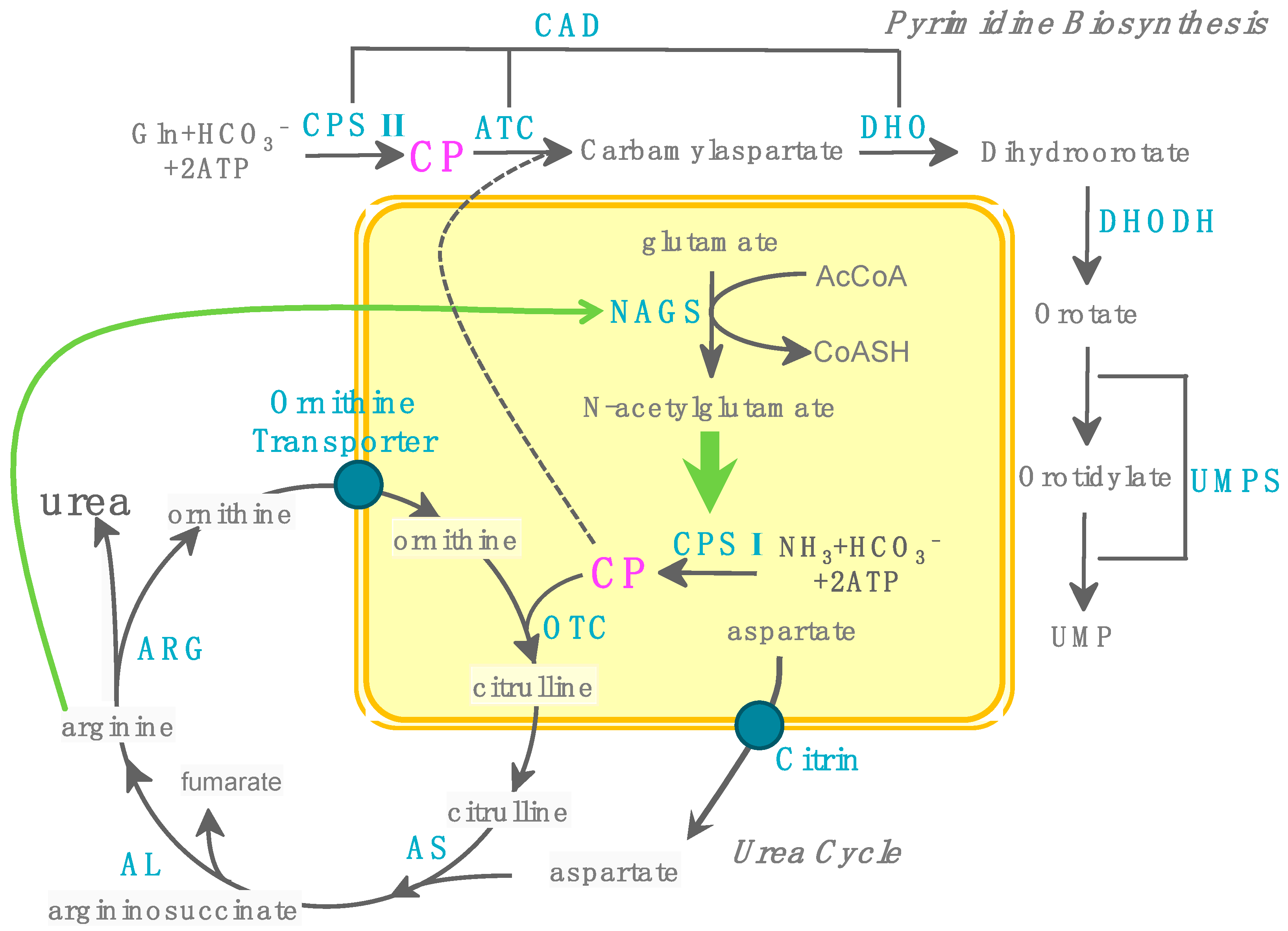
3.1. Production of CP Using CPS
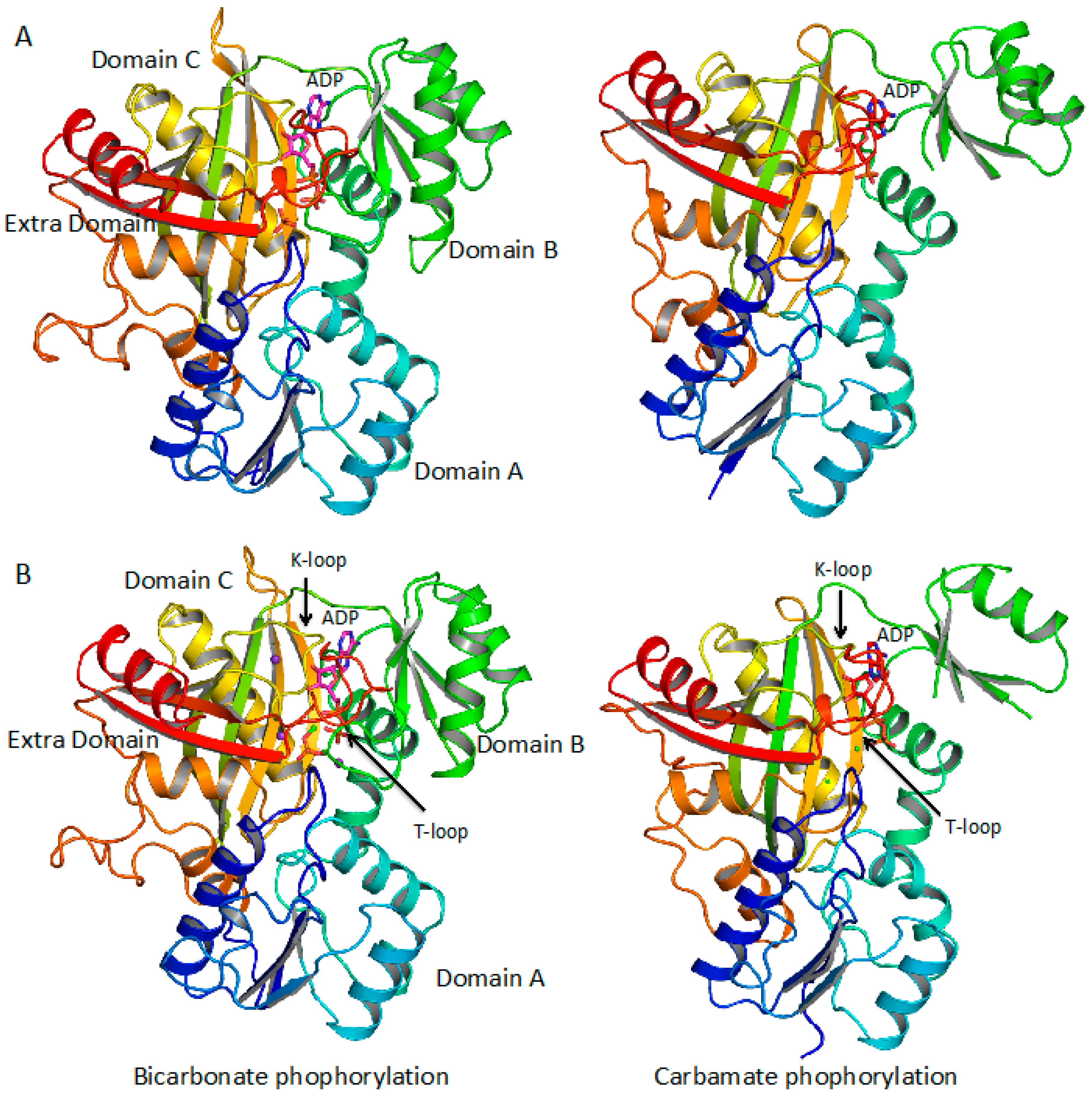
3.2. Production of CP Using Carbamate Kinase
3.3. Production of CP Using Catabolic Transcarbamylases
4. CP as Carbamyl Group Donor
4.1. Amino Nitrogen as a Carbamyl Group Acceptor
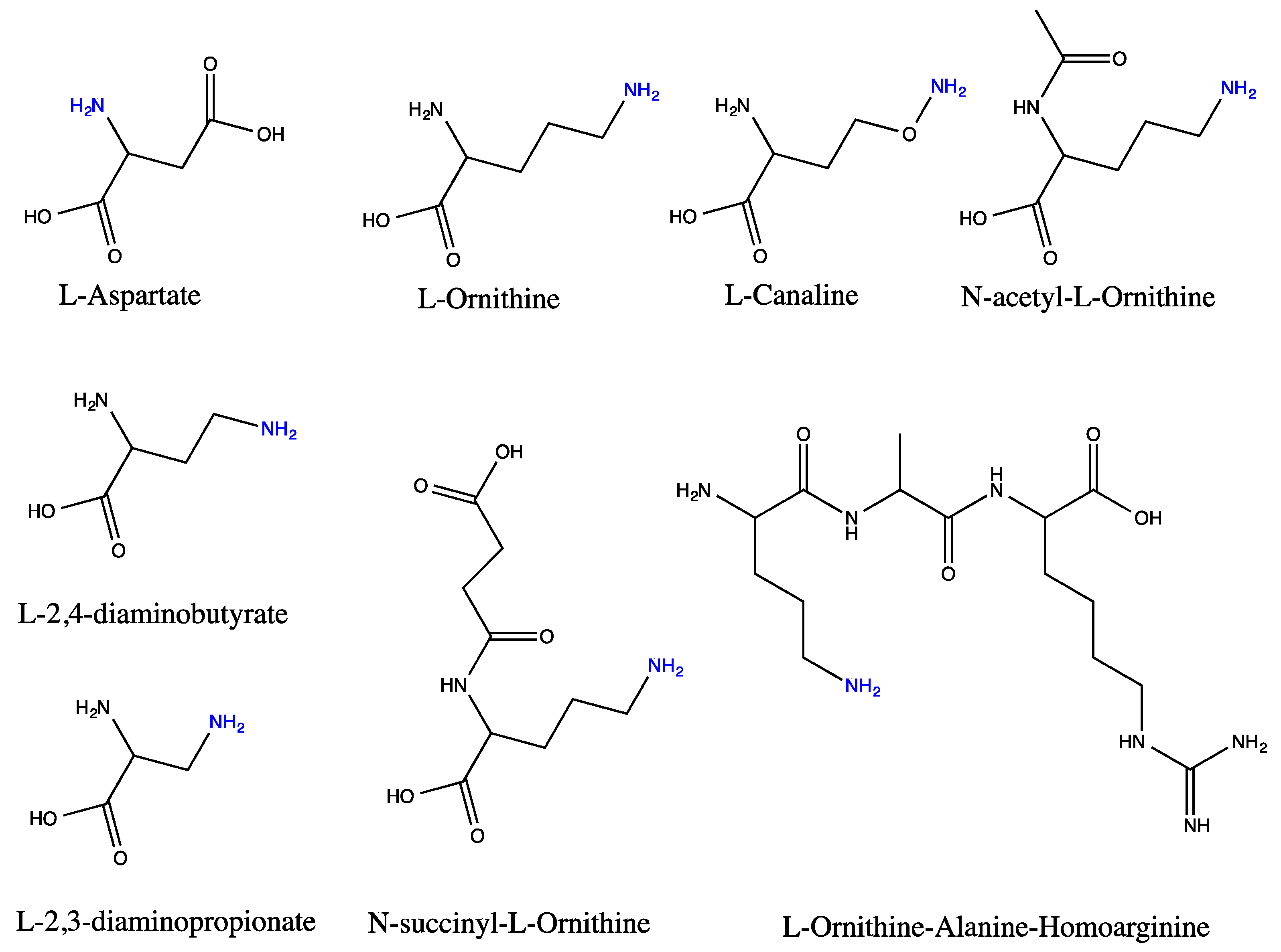
4.1.1. Aspartate as Acceptor
4.1.2. Ornithine and Other Ornithine Derivatives as Acceptors
4.1.3. l-2,3-diaminopropionate and l-2,4-diaminobutyrate as Acceptor
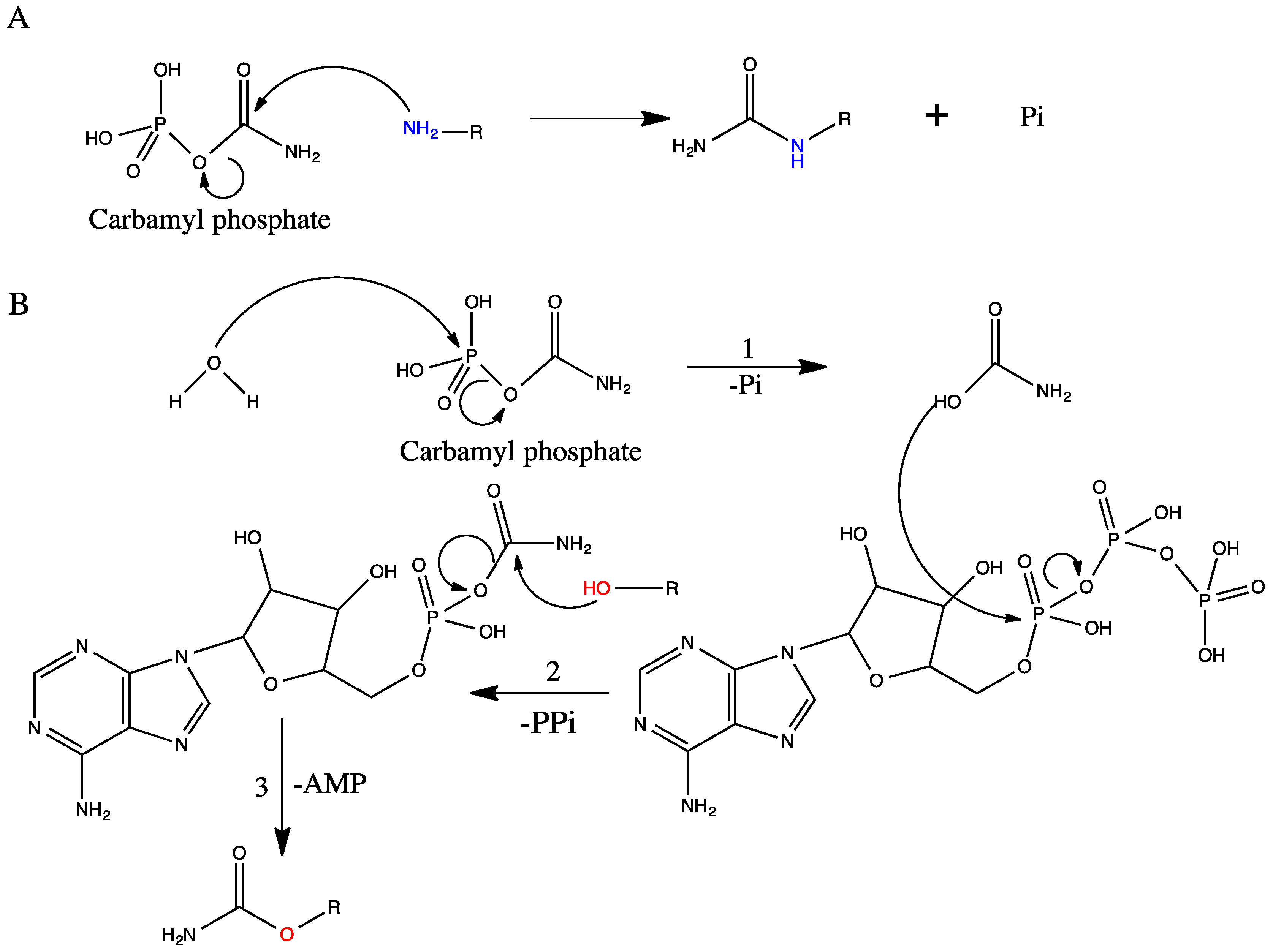
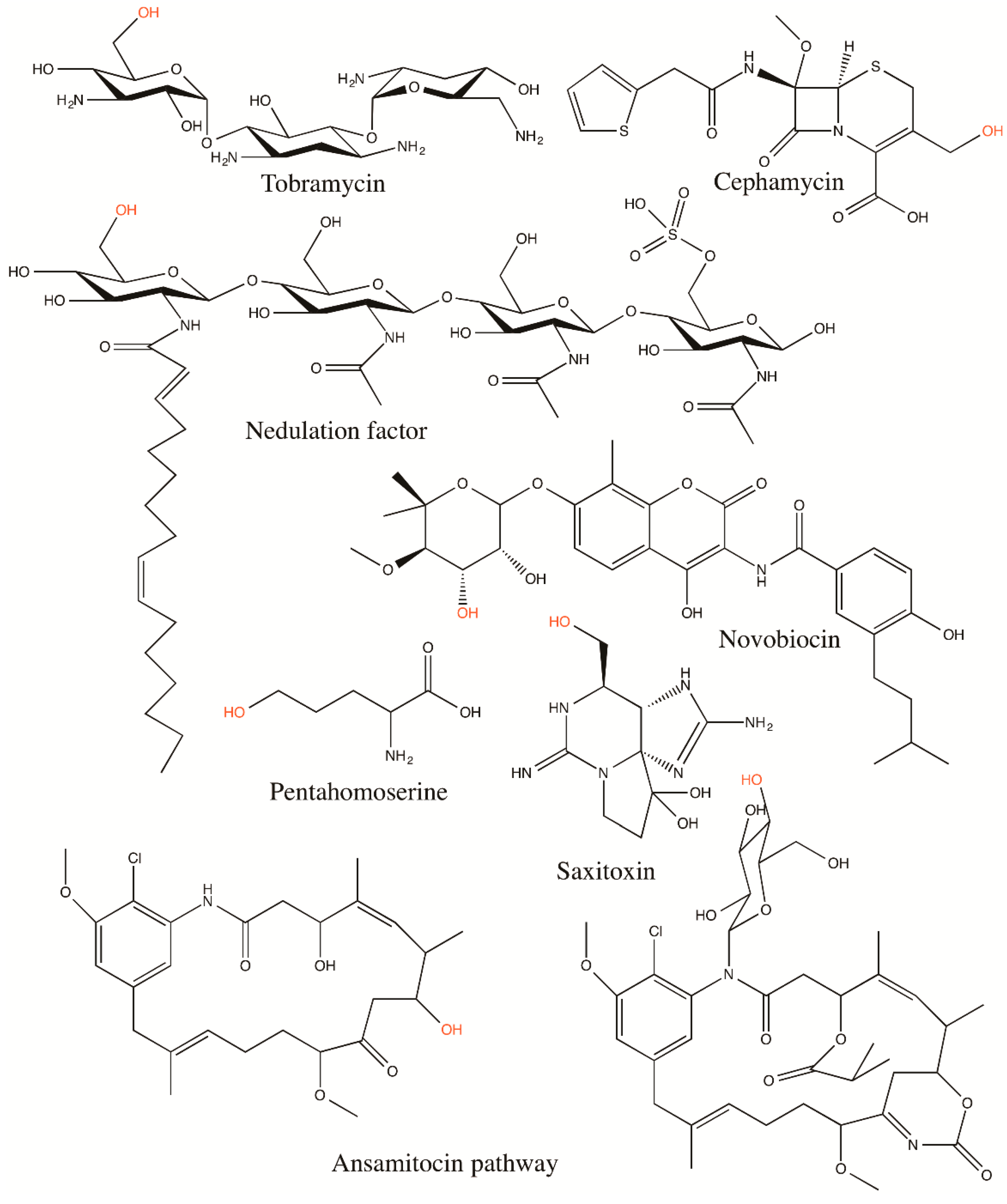
4.2. Hydroxyl Oxygen as a Carbamyl Group Acceptor
4.3. Sulfur Group as an Acceptor
5. CP as Phosphate Group Donor for ATP Production
6. Protection of CP
7. CP Accumulation and Health
8. Future Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, M.E.; Spector, L.; Lipmann, F. Carbamyl phosphate, the carbamyl donor in enzymatic citrulline synthesis. J. Am. Chem. Soc. 1955, 77, 819–820. [Google Scholar] [CrossRef]
- Keefe, A.D.; Miller, S.L. Are polyphosphates or phosphate esters prebiotic reagents? J. Mol. Evol. 1995, 41, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.E. Carbamyl phosphate: Many forms of life use this molecule to synthesize arginine, uracil, and adenosine triphosphate. Science 1963, 140, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.E.; Lipmann, F. Chemical and enzymatic synthesis of carbamyl phosphate. Proc. Natl. Acad. Sci. USA 1960, 46, 1194–1205. [Google Scholar] [CrossRef] [PubMed]
- Fawaz, M.V.; Topper, M.E.; Firestine, S.M. The ATP-grasp enzymes. Bioorg. Chem. 2011, 39, 185–191. [Google Scholar] [CrossRef] [PubMed]
- De Cima, S.; Polo, L.M.; Diez-Fernandez, C.; Martinez, A.I.; Cervera, J.; Fita, I.; Rubio, V. Structure of human carbamoyl phosphate synthetase: Deciphering the on/off switch of human ureagenesis. Sci. Rep. 2015, 5, 16950. [Google Scholar] [CrossRef] [PubMed]
- Thoden, J.B.; Holden, H.M.; Wesenberg, G.; Raushel, F.M.; Rayment, I. Structure of carbamoyl phosphate synthetase: A journey of 96 a from substrate to product. Biochemistry 1997, 36, 6305–6316. [Google Scholar] [CrossRef] [PubMed]
- Purcarea, C.; Evans, D.R.; Herve, G. Channeling of carbamoyl phosphate to the pyrimidine and arginine biosynthetic pathways in the deep sea hyperthermophilic archaeon Pyrococcus abyssi. J. Biol. Chem. 1999, 274, 6122–6129. [Google Scholar] [CrossRef] [PubMed]
- Purcarea, C.; Simon, V.; Prieur, D.; Herve, G. Purification and characterization of carbamoyl-phosphate synthetase from the deep-sea hyperthermophilic archaebacterium Pyrococcus abyssi. Eur. J. Biochem. 1996, 236, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Ramon-Maiques, S.; Marina, A.; Uriarte, M.; Fita, I.; Rubio, V. The 1.5 Å resolution crystal structure of the carbamate kinase-like carbamoyl phosphate synthetase from the hyperthermophilic archaeon Pyrococcus furiosus, bound to ADP, confirms that this thermostable enzyme is a carbamate kinase, and provides insight into substrate binding and stability in carbamate kinases. J. Mol. Biol. 2000, 299, 463–476. [Google Scholar] [PubMed]
- Uriarte, M.; Marina, A.; Ramon-Maiques, S.; Fita, I.; Rubio, V. The carbamoyl-phosphate synthetase of Pyrococcus furiosus is enzymologically and structurally a carbamate kinase. J. Biol. Chem. 1999, 274, 16295–16303. [Google Scholar] [CrossRef] [PubMed]
- Uriarte, M.; Marina, A.; Ramon-Maiques, S.; Rubio, V.; Durbecq, V.; Legrain, C.; Glansdorff, N. Carbamoyl phosphate synthesis: Carbamate kinase from Pyrococcus furiosus. Methods Enzymol. 2001, 331, 236–247. [Google Scholar] [PubMed]
- Tricot, C.; Villeret, V.; Sainz, G.; Dideberg, O.; Stalon, V. Allosteric regulation in Pseudomonas aeruginosa catabolic ornithine carbamoyltransferase revisited: Association of concerted homotropic cooperative interactions and local heterotropic effects. J. Mol. Biol. 1998, 283, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Sainz, G.; Tricot, C.; Foray, M.F.; Marion, D.; Dideberg, O.; Stalon, V. Kinetic studies of allosteric catabolic ornithine carbamoyltransferase from Pseudomonas aeruginosa. Eur. J. Biochem. 1998, 251, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.T.; Tu, B.P.; Tang, Y. Eight kinetically stable but thermodynamically activated molecules that power cell metabolism. Chem. Rev. 2018, 118, 1460–1494. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Allewell, N.M.; Tuchman, M. From genome to structure and back again: A family portrait of the transcarbamylases. Int. J. Mol. Sci. 2015, 16, 18836–18864. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.L.; Dalisay, D.S.; Andersen, R.J.; Ryan, K.S. N-carbamoylation of 2,4-diaminobutyrate reroutes the outcome in padanamide biosynthesis. Chem. Biol. 2013, 20, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.G.; Chan, Y.A.; Ozanick, S.G. Deciphering tuberactinomycin biosynthesis: Isolation, sequencing, and annotation of the viomycin biosynthetic gene cluster. Antimicrob. Agents Chemother. 2003, 47, 2823–2830. [Google Scholar] [CrossRef] [PubMed]
- Felnagle, E.A.; Rondon, M.R.; Berti, A.D.; Crosby, H.A.; Thomas, M.G. Identification of the biosynthetic gene cluster and an additional gene for resistance to the antituberculosis drug capreomycin. Appl. Environ. Microbiol. 2007, 73, 4162–4170. [Google Scholar] [CrossRef] [PubMed]
- Kevany, B.M.; Rasko, D.A.; Thomas, M.G. Characterization of the complete zwittermicin a biosynthesis gene cluster from Bacillus cereus. Appl. Environ. Microbiol. 2009, 75, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, P.; Kang, Q.; Ma, J.; Bai, L.; Deng, Z. Dual carbamoylations on the polyketide and glycosyl moiety by asm21 result in extended ansamitocin biosynthesis. Chem. Biol. 2011, 18, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Parthier, C.; Gorlich, S.; Jaenecke, F.; Breithaupt, C.; Brauer, U.; Fandrich, U.; Clausnitzer, D.; Wehmeier, U.F.; Bottcher, C.; Scheel, D.; et al. The O-carbamoyltransferase TobZ catalyzes an ancient enzymatic reaction. Angew. Chem. Int. Ed. Engl. 2012, 51, 4046–4052. [Google Scholar] [CrossRef] [PubMed]
- Petras, D.; Kerwat, D.; Pesic, A.; Hempel, B.F.; von Eckardstein, L.; Semsary, S.; Araste, J.; Marguerettaz, M.; Royer, M.; Cociancich, S.; et al. The O-carbamoyl-transferase alb15 is responsible for the modification of albicidin. ACS Chem. Biol. 2016, 11, 1198–1204. [Google Scholar] [CrossRef] [PubMed]
- Shomura, Y.; Higuchi, Y. Structural basis for the reaction mechanism of S-carbamoylation of HypE by HypF in the maturation of [NiFe]-hydrogenases. J. Biol. Chem. 2012, 287, 28409–28419. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Salo, W.L.; Lusty, C.J.; Anderson, P.M. Carbamyl phosphate synthetase III, an evolutionary intermediate in the transition between glutamine-dependent and ammonia-dependent carbamyl phosphate synthetases. J. Mol. Biol. 1994, 243, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Windmueller, H.G.; Spaeth, A.E. Source and fate of circulating citrulline. Am. J. Physiol. 1981, 241, E473–E480. [Google Scholar] [CrossRef] [PubMed]
- Dhanakoti, S.N.; Brosnan, J.T.; Herzberg, G.R.; Brosnan, M.E. Renal arginine synthesis: Studies in vitro and in vivo. Am. J. Physiol. 1990, 259, E437–E442. [Google Scholar] [CrossRef] [PubMed]
- Trotta, P.P.; Burt, M.E.; Haschemeyer, R.H.; Meister, A. Reversible dissociation of carbamyl phosphate synthetase into a regulated synthesis subunit and a subunit required for glutamine utilization. Proc. Natl. Acad. Sci. USA 1971, 68, 2599–2603. [Google Scholar] [CrossRef] [PubMed]
- Post, L.E.; Post, D.J.; Raushel, F.M. Dissection of the functional domains of Escherichia coli carbamoyl phosphate synthetase by site-directed mutagenesis. J. Biol. Chem. 1990, 265, 7742–7747. [Google Scholar] [PubMed]
- Guy, H.I.; Evans, D.R. Function of the major synthetase subdomains of carbamyl-phosphate synthetase. J. Biol. Chem. 1996, 271, 13762–13769. [Google Scholar] [CrossRef] [PubMed]
- Alonso, E.; Rubio, V. Affinity cleavage of carbamoyl-phosphate synthetase I localizes regions of the enzyme interacting with the molecule of ATP that phosphorylates carbamate. Eur. J. Biochem. 1995, 229, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Guillou, F.; Rubino, S.D.; Markovitz, R.S.; Kinney, D.M.; Lusty, C.J. Escherichia coli carbamoyl-phosphate synthetase: Domains of glutaminase and synthetase subunit interaction. Proc. Natl. Acad. Sci. USA 1989, 86, 8304–8308. [Google Scholar] [CrossRef] [PubMed]
- Galperin, M.Y.; Koonin, E.V. Divergence and convergence in enzyme evolution. J. Biol. Chem. 2012, 287, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Jin, Z.; Wang, Y.; Allewell, N.M.; Tuchman, M.; Shi, D. Structure and function of Escherichia coli RimK, an ATP-grasp fold, l-glutamyl ligase enzyme. Proteins 2013, 81, 1847–1854. [Google Scholar] [CrossRef] [PubMed]
- Galperin, M.Y.; Koonin, E.V. A diverse superfamily of enzymes with ATP-dependent carboxylate-amine/thiol ligase activity. Protein Sci. 1997, 6, 2639–2643. [Google Scholar] [CrossRef] [PubMed]
- Mouilleron, S.; Golinelli-Pimpaneau, B. Conformational changes in ammonia-channeling glutamine amidotransferases. Curr. Opin. Struct. Biol. 2007, 17, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Diez-Fernandez, C.; Haberle, J. Targeting CPS1 in the treatment of Carbamoyl phosphate synthetase 1 (CPS1) deficiency, a urea cycle disorder. Expert Opin. Ther. Targets 2017, 21, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.I.; Perez-Arellano, I.; Pekkala, S.; Barcelona, B.; Cervera, J. Genetic, structural and biochemical basis of carbamoyl phosphate synthetase 1 deficiency. Mol. Genet. Metab. 2010, 101, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Diez-Fernandez, C.; Martinez, A.I.; Pekkala, S.; Barcelona, B.; Perez-Arellano, I.; Guadalajara, A.M.; Summar, M.; Cervera, J.; Rubio, V. Molecular characterization of carbamoyl-phosphate synthetase (CPS1) deficiency using human recombinant cps1 as a key tool. Hum. Mutat. 2013, 34, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Diez-Fernandez, C.; Gallego, J.; Haberle, J.; Cervera, J.; Rubio, V. The study of carbamoyl phosphate synthetase 1 deficiency sheds light on the mechanism for switching on/off the urea cycle. J. Genet. Genom. 2015, 42, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, A.; Purcarea, C.; Guy, H.I.; Evans, D.R. A novel carbamoyl-phosphate synthetase from Aquifex aeolicus. J. Biol. Chem. 2001, 276, 45694–45703. [Google Scholar] [CrossRef] [PubMed]
- Popa, E.; Perera, N.; Kibedi-Szabo, C.Z.; Guy-Evans, H.; Evans, D.R.; Purcarea, C. The smallest active carbamoyl phosphate synthetase was identified in the human gut archaeon Methanobrevibacter smithii. J. Mol. Microbiol. Biotechnol. 2012, 22, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Nyunoya, H.; Lusty, C.J. The carB gene of Escherichia coli: A duplicated gene coding for the large subunit of carbamoyl-phosphate synthetase. Proc. Natl. Acad. Sci. USA 1983, 80, 4629–4633. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J.N.; Chen, K.C.; Jamison, R.S.; Musmanno, L.A.; Kern, C.B. The evolutionary history of the first three enzymes in pyrimidine biosynthesis. Bioessays 1993, 15, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Nyunoya, H.; Broglie, K.E.; Lusty, C.J. The gene coding for carbamoyl-phosphate synthetase I was formed by fusion of an ancestral glutaminase gene and a synthetase gene. Proc. Natl. Acad. Sci. USA 1985, 82, 2244–2246. [Google Scholar] [CrossRef] [PubMed]
- Rubio, V.; Cervera, J.; Lusty, C.J.; Bendala, E.; Britton, H.G. Domain structure of the large subunit of Escherichia coli carbamoyl phosphate synthetase. Location of the binding site for the allosteric inhibitor ump in the cooh-terminal domain. Biochemistry 1991, 30, 1068–1075. [Google Scholar] [CrossRef] [PubMed]
- Alcantara, C.; Cervera, J.; Rubio, V. Carbamate kinase can replace in vivo carbamoyl phosphate synthetase. Implications for the evolution of carbamoyl phosphate biosynthesis. FEBS Lett. 2000, 484, 261–264. [Google Scholar] [CrossRef]
- Durbecq, V.; Legrain, C.; Roovers, M.; Pierard, A.; Glansdorff, N. The carbamate kinase-like carbamoyl phosphate synthetase of the hyperthermophilic archaeon Pyrococcus furiosus, a missing link in the evolution of carbamoyl phosphate biosynthesis. Proc. Natl. Acad. Sci. USA 1997, 94, 12803–12808. [Google Scholar] [CrossRef] [PubMed]
- Marina, A.; Alzari, P.M.; Bravo, J.; Uriarte, M.; Barcelona, B.; Fita, I.; Rubio, V. Carbamate kinase: New structural machinery for making carbamoyl phosphate, the common precursor of pyrimidines and arginine. Protein Sci. 1999, 8, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Purcarea, C.; Herve, G.; Cunin, R.; Evans, D.R. Cloning, expression, and structure analysis of carbamate kinase-like carbamoyl phosphate synthetase from pyrococcus abyssi. Extremophiles 2001, 5, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Massant, J.; Glansdorff, N. New experimental approaches for investigating interactions between Pyrococcus furiosus carbamate kinase and carbamoyltransferases, enzymes involved in the channeling of thermolabile carbamoyl phosphate. Archaea 2005, 1, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Abdelal, A.T. Arginine catabolism by microorganisms. Ann. Rev. Microbiol. 1979, 33, 139–168. [Google Scholar] [CrossRef] [PubMed]
- Gallego, P.; Planell, R.; Benach, J.; Querol, E.; Perez-Pons, J.A.; Reverter, D. Structural characterization of the enzymes composing the arginine deiminase pathway in mycoplasma penetrans. PLoS ONE 2012, 7, e47886. [Google Scholar] [CrossRef] [PubMed]
- Ramon-Maiques, S.; Marina, A.; Guinot, A.; Gil-Ortiz, F.; Uriarte, M.; Fita, I.; Rubio, V. Substrate binding and catalysis in carbamate kinase ascertained by crystallographic and site-directed mutagenesis studies: Movements and significance of a unique globular subdomain of this key enzyme for fermentative ATP production in bacteria. J. Mol. Biol. 2010, 397, 1261–1275. [Google Scholar] [CrossRef] [PubMed]
- Ramon-Maiques, S.; Marina, A.; Gil-Ortiz, F.; Fita, I.; Rubio, V. Structure of acetylglutamate kinase, a key enzyme for arginine biosynthesis and a prototype for the amino acid kinase enzyme family, during catalysis. Structure 2002, 10, 329–342. [Google Scholar] [CrossRef]
- Marco-Marin, C.; Gil-Ortiz, F.; Rubio, V. The crystal structure of Pyrococcus furiosus ump kinase provides insight into catalysis and regulation in microbial pyrimidine nucleotide biosynthesis. J. Mol. Biol. 2005, 352, 438–454. [Google Scholar] [CrossRef] [PubMed]
- Marco-Marin, C.; Gil-Ortiz, F.; Perez-Arellano, I.; Cervera, J.; Fita, I.; Rubio, V. A novel two-domain architecture within the amino acid kinase enzyme family revealed by the crystal structure of Escherichia coli glutamate 5-kinase. J. Mol. Biol. 2007, 367, 1431–1446. [Google Scholar] [CrossRef] [PubMed]
- Pakhomova, S.; Bartlett, S.G.; Augustus, A.; Kuzuyama, T.; Newcomer, M.E. Crystal structure of fosfomycin resistance kinase foma from streptomyces wedmorensis. J. Biol. Chem. 2008, 283, 28518–28526. [Google Scholar] [CrossRef] [PubMed]
- Kotaka, M.; Ren, J.; Lockyer, M.; Hawkins, A.R.; Stammers, D.K. Structures of R- and T-state Escherichia coli aspartokinase III. Mechanisms of the allosteric transition and inhibition by lysine. J. Biol. Chem. 2006, 281, 31544–31552. [Google Scholar] [CrossRef] [PubMed]
- Baur, H.; Luethi, E.; Stalon, V.; Mercenier, A.; Haas, D. Sequence analysis and expression of the arginine-deiminase and carbamate-kinase genes of pseudomonas aeruginosa. Eur. J. Biochem. 1989, 179, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Rubio, V.; Cervera, J. The carbamoyl-phosphate synthase family and carbamate kinase: Structure-function studies. Biochem. Soc. Trans. 1995, 23, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Galkin, A.; Kulakova, L.; Lim, K.; Chen, C.Z.; Zheng, W.; Turko, I.V.; Herzberg, O. Structural basis for inactivation of giardia lamblia carbamate kinase by disulfiram. J. Biol. Chem. 2014, 289, 10502–10509. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jin, Z.; Yu, X.; Allewell, N.M.; Tuchman, M.; Shi, D. The ygew encoded protein from Escherichia coli is a knotted ancestral catabolic transcarbamylase. Proteins 2011, 79, 2327–2334. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Yu, X.; Zhao, G.; Ho, J.; Lu, S.; Allewell, N.M.; Tuchman, M. Crystal structure and biochemical properties of putrescine carbamoyltransferase from Enterococcus faecalis: Assembly, active site, and allosteric regulation. Proteins 2012, 80, 1436–1447. [Google Scholar] [CrossRef] [PubMed]
- Naumoff, D.G.; Xu, Y.; Stalon, V.; Glansdorff, N.; Labedan, B. The difficulty of annotating genes: The case of putrescine carbamoyltransferase. Microbiology 2004, 150, 3908–3911. [Google Scholar] [CrossRef] [PubMed]
- Naumoff, D.G.; Xu, Y.; Glansdorff, N.; Labedan, B. Retrieving sequences of enzymes experimentally characterized but erroneously annotated: The case of the putrescine carbamoyltransferase. BMC Genom. 2004, 5, 52. [Google Scholar] [CrossRef] [PubMed]
- Griswold, A.R.; Chen, Y.Y.; Burne, R.A. Analysis of an agmatine deiminase gene cluster in streptococcus mutans UA159. J. Bacteriol. 2004, 186, 1902–1904. [Google Scholar] [CrossRef] [PubMed]
- Barba, M.; Dutoit, R.; Legrain, C.; Labedan, B. Identifying reaction modules in metabolic pathways: Bioinformatic deduction and experimental validation of a new putative route in purine catabolism. BMC Syst. Biol. 2013, 7, 99. [Google Scholar] [CrossRef] [PubMed]
- Barcelona-Andres, B.; Marina, A.; Rubio, V. Gene structure, organization, expression, and potential regulatory mechanisms of arginine catabolism in Enterococcus faecalis. J. Bacteriol. 2002, 184, 6289–6300. [Google Scholar] [CrossRef] [PubMed]
- Llacer, J.L.; Polo, L.M.; Tavarez, S.; Alarcon, B.; Hilario, R.; Rubio, V. The gene cluster for agmatine catabolism of Enterococcus faecalis: Study of recombinant putrescine transcarbamylase and agmatine deiminase and a snapshot of agmatine deiminase catalyzing its reaction. J. Bacteriol. 2007, 189, 1254–1265. [Google Scholar] [CrossRef] [PubMed]
- Stalon, V.; Ramos, F.; Pierard, A.; Wiame, J.M. Regulation of the catabolic ornithine carbamoyltransferase of pseudomonas fluorescens. A comparison with the anabolic transferase and with a mutationally modified catabolic transferase. Eur. J. Biochem. 1972, 29, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Villeret, V.; Tricot, C.; Stalon, V.; Dideberg, O. Crystal structure of pseudomonas aeruginosa catabolic ornithine transcarbamoylase at 3.0-A resolution: A different oligomeric organization in the transcarbamoylase family. Proc. Natl. Acad. Sci. USA 1995, 92, 10762–10766. [Google Scholar] [CrossRef] [PubMed]
- De Las Rivas, B.; Fox, G.C.; Angulo, I.; Ripoll, M.M.; Rodriguez, H.; Munoz, R.; Mancheno, J.M. Crystal structure of the hexameric catabolic ornithine transcarbamylase from Lactobacillus hilgardii: Structural insights into the oligomeric assembly and metal binding. J. Mol. Biol. 2009, 393, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Galkin, A.; Kulakova, L.; Wu, R.; Gong, M.; Dunaway-Mariano, D.; Herzberg, O. X-ray structure and kinetic properties of ornithine transcarbamoylase from the human parasite giardia lamblia. Proteins 2009, 76, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Wargnies, B.; Lauwers, N.; Stalon, V. Structure and properties of the putrescine carbamoyltransferase of Streptococcus faecalis. Eur. J. Biochem. 1979, 101, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.P.; Stalon, V. Enzymes of agmatine degradation and the control of their synthesis in Streptococcus faecalis. J. Bacteriol. 1982, 152, 676–681. [Google Scholar] [PubMed]
- Polo, L.M.; Gil-Ortiz, F.; Cantin, A.; Rubio, V. New insight into the transcarbamylase family: The structure of putrescine transcarbamylase, a key catalyst for fermentative utilization of agmatine. PLoS ONE 2012, 7, e31528. [Google Scholar] [CrossRef] [PubMed]
- Tigier, H.; Grisolia, S. Induction of carbamyl-p specific oxamate transcarbamylase by parabanic acid in a streptococcus. Biochem. Biophys. Res. Commun. 1965, 19, 209–214. [Google Scholar] [CrossRef]
- Vander Wauven, C.; Simon, J.P.; Slos, P.; Stalon, V. Control of enzyme synthesis in the oxalurate catabolic pathway of Streptococcus faecalis ATCC 11700: Evidence for the existence of a third carbamate kinase. Arch. Microbiol. 1986, 145, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Xi, H.; Schneider, B.L.; Reitzer, L. Purine catabolism in Escherichia coli and function of xanthine dehydrogenase in purine salvage. J. Bacteriol. 2000, 182, 5332–5341. [Google Scholar] [CrossRef] [PubMed]
- Cusa, E.; Obradors, N.; Baldoma, L.; Badia, J.; Aguilar, J. Genetic analysis of a chromosomal region containing genes required for assimilation of allantoin nitrogen and linked glyoxylate metabolism in Escherichia coli. J. Bacteriol. 1999, 181, 7479–7484. [Google Scholar] [PubMed]
- Labedan, B.; Boyen, A.; Baetens, M.; Charlier, D.; Chen, P.; Cunin, R.; Durbeco, V.; Glansdorff, N.; Herve, G.; Legrain, C.; et al. The evolutionary history of carbamoyltransferases: A complex set of paralogous genes was already present in the last universal common ancestor. J. Mol. Evol. 1999, 49, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Labedan, B.; Xu, Y.; Naumoff, D.G.; Glansdorff, N. Using quaternary structures to assess the evolutionary history of proteins: The case of the aspartate carbamoyltransferase. Mol. Biol. Evol. 2004, 21, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Lipscomb, W.N. Aspartate transcarbamylase from Escherichia coli: Activity and regulation. Adv. Enzymol. Relat. Areas Mol. Biol. 1994, 68, 67–151. [Google Scholar] [PubMed]
- Lowenstein, J.M.; Cohen, P.P. Studies on the biosynthesis of carbamylaspartic acid. J. Biol. Chem. 1956, 220, 57–70. [Google Scholar] [PubMed]
- Lipscomb, W.N.; Kantrowitz, E.R. Structure and mechanisms of Escherichia coli aspartate transcarbamoylase. Acc. Chem. Res. 2012, 45, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Schurr, M.J.; Vickrey, J.F.; Kumar, A.P.; Campbell, A.L.; Cunin, R.; Benjamin, R.C.; Shanley, M.S.; O’Donovan, G.A. Aspartate transcarbamoylase genes of pseudomonas putida: Requirement for an inactive dihydroorotase for assembly into the dodecameric holoenzyme. J. Bacteriol. 1995, 177, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Hughes, L.E.; Hooshdaran, M.Z.; O’Donovan, G.A. Streptomyces aspartate transcarbamoylase is a dodecamer with dihydroorotase activity. Curr. Microbiol. 1999, 39, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Van Vliet, F.; Van De Casteele, M.; Legrain, C.; Cunin, R.; Glansdorff, N. Aspartate transcarbamylase from the hyperthermophilic eubacterium Thermotoga maritima: Fused catalytic and regulatory polypeptides form an allosteric enzyme. J. Bacteriol. 1998, 180, 6389–6391. [Google Scholar] [PubMed]
- Brabson, J.S.; Switzer, R.L. Purification and properties of bacillus subtilis aspartate transcarbamylase. J. Biol. Chem. 1975, 250, 8664–8669. [Google Scholar] [PubMed]
- Coleman, P.F.; Suttle, D.P.; Stark, G.R. Purification from hamster cells of the multifunctional protein that initiates de novo synthesis of pyrimidine nucleotides. J. Biol. Chem. 1977, 252, 6379–6385. [Google Scholar] [PubMed]
- Souciet, J.L.; Nagy, M.; Le Gouar, M.; Lacroute, F.; Potier, S. Organization of the yeast URA2 gene: Identification of a defective dihydroorotase-like domain in the multifunctional carbamoylphosphate synthetase-aspartate transcarbamylase complex. Gene 1989, 79, 59–70. [Google Scholar] [CrossRef]
- Khan, A.I.; Chowdhry, B.Z.; Yon, R.J. Wheat-germ aspartate transcarbamoylase: Revised purification, stability and re-evaluation of regulatory kinetics in terms of the monod-wyman-changeux model. Eur. J. Biochem. 1999, 259, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Williamson, C.L.; Slocum, R.D. Molecular cloning and characterization of the pyrB1 and pyrB2 genes encoding aspartate transcarbamoylase in pea (Pisum sativum L.). Plant Physiol. 1994, 105, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Martin, P.D.; Purcarea, C.; Vaishnav, A.; Brunzelle, J.S.; Fernando, R.; Guy-Evans, H.I.; Evans, D.R.; Edwards, B.F. Dihydroorotase from the hyperthermophile Aquifex aeolicus is activated by stoichiometric association with aspartate transcarbamoylase and forms a one-pot reactor for pyrimidine biosynthesis. Biochemistry 2009, 48, 766–778. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Morcillo, M.; Grande-Garcia, A.; Ruiz-Ramos, A.; Del Cano-Ochoa, F.; Boskovic, J.; Ramon-Maiques, S. Structural insight into the core of CAD, the multifunctional protein leading de novo pyrimidine biosynthesis. Structure 2017, 25, 912–923. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, P.; Deng, Z.; Zhao, C. Ornithine transcarbamylase ArgK plays a dual role for the self-defense of phaseolotoxin producing Pseudomonas syringae pv. phaseolicola. Sci. Rep. 2015, 5, 12892. [Google Scholar] [CrossRef] [PubMed]
- Morizono, H.; Cabrera-Luque, J.; Shi, D.; Gallegos, R.; Yamaguchi, S.; Yu, X.; Allewell, N.M.; Malamy, M.H.; Tuchman, M. Acetylornithine transcarbamylase: A novel enzyme in arginine biosynthesis. J. Bacteriol. 2006, 188, 2974–2982. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Morizono, H.; Cabrera-Luque, J.; Yu, X.; Roth, L.; Malamy, M.H.; Allewell, N.M.; Tuchman, M. Structure and catalytic mechanism of a novel N-succinyl-l-ornithine transcarbamylase in arginine biosynthesis of Bacteroides fragilis. J. Biol. Chem. 2006, 281, 20623–20631. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Morizono, H.; Yu, X.; Roth, L.; Caldovic, L.; Allewell, N.M.; Malamy, M.H.; Tuchman, M. Crystal structure of N-acetylornithine transcarbamylase from Xanthomonas campestris: A novel enzyme in a new arginine biosynthetic pathway found in several eubacteria. J. Biol. Chem. 2005, 280, 14366–14369. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kwon, Y.M. Identification of an isoform of ornithine carbamoyltransferase that can effectively utilize canaline as a substrate from the leaves of Canavalia lineata. Plant Sci. 2000, 151, 145–151. [Google Scholar] [CrossRef]
- Slocum, R.D. Genes, enzymes and regulation of arginine biosynthesis in plants. Plant Physiol. Biochem. 2005, 43, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Yu, X.; Cabrera-Luque, J.; Chen, T.Y.; Roth, L.; Morizono, H.; Allewell, N.M.; Tuchman, M. A single mutation in the active site swaps the substrate specificity of n-acetyl-l-ornithine transcarbamylase and N-succinyl-l-ornithine transcarbamylase. Protein Sci. 2007, 16, 1689–1699. [Google Scholar] [CrossRef] [PubMed]
- Barkei, J.J.; Kevany, B.M.; Felnagle, E.A.; Thomas, M.G. Investigations into viomycin biosynthesis by using heterologous production in Streptomyces lividans. Chembiochem 2009, 10, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Brewer, S.J.; Taylor, P.M.; Turner, M.K. An adenosine triphosphate-dependent carbamoylphosphate--3-hydroxymethylcephem O-carbamoyltransferase from Streptomyces clavuligerus. Biochem. J. 1980, 185, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Freel Meyers, C.L.; Oberthur, M.; Xu, H.; Heide, L.; Kahne, D.; Walsh, C.T. Characterization of NovP and NovN: Completion of novobiocin biosynthesis by sequential tailoring of the noviosyl ring. Angew. Chem. Int. Ed. Engl. 2004, 43, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Haydock, S.F.; Appleyard, A.N.; Mironenko, T.; Lester, J.; Scott, N.; Leadlay, P.F. Organization of the biosynthetic gene cluster for the macrolide concanamycin A in Streptomyces neyagawaensis ATCC 27449. Microbiology 2005, 151, 3161–3169. [Google Scholar] [CrossRef] [PubMed]
- Higashide, E.; Asai, M.; Ootsu, K.; Tanida, S.; Kozai, Y.; Hasegawa, T.; Kishi, T.; Sugino, Y.; Yoneda, M. Ansamitocin, a group of novel maytansinoid antibiotics with antitumour properties from nocardia. Nature 1977, 270, 721–722. [Google Scholar] [CrossRef] [PubMed]
- Wehmeier, U.F.; Piepersberg, W. Enzymology of aminoglycoside biosynthesis-deduction from gene clusters. Methods Enzymol. 2009, 459, 459–491. [Google Scholar] [PubMed]
- Chen, W.; Huang, T.; He, X.; Meng, Q.; You, D.; Bai, L.; Li, J.; Wu, M.; Li, R.; Xie, Z.; et al. Characterization of the polyoxin biosynthetic gene cluster from Streptomyces cacaoi and engineered production of polyoxin H. J. Biol. Chem. 2009, 284, 10627–10638. [Google Scholar] [CrossRef] [PubMed]
- Kharel, M.K.; Basnet, D.B.; Lee, H.C.; Liou, K.; Woo, J.S.; Kim, B.G.; Sohng, J.K. Isolation and characterization of the tobramycin biosynthetic gene cluster from Streptomyces tenebrarius. FEMS Microbiol. Lett. 2004, 230, 185–190. [Google Scholar] [CrossRef]
- Jabbouri, S.; Fellay, R.; Talmont, F.; Kamalaprija, P.; Burger, U.; Relic, B.; Prome, J.C.; Broughton, W.J. Involvement of nodS in N-methylation and nodU in 6-O-carbamoylation of Rhizobium sp. NGR234 Nod Factors. J. Biol. Chem. 1995, 270, 22968–22973. [Google Scholar] [CrossRef] [PubMed]
- Kellmann, R.; Mihali, T.K.; Neilan, B.A. Identification of a saxitoxin biosynthesis gene with a history of frequent horizontal gene transfers. J. Mol. Evol. 2008, 67, 526–538. [Google Scholar] [CrossRef] [PubMed]
- Coque, J.J.; Perez-Llarena, F.J.; Enguita, F.J.; Fuente, J.L.; Martin, J.F.; Liras, P. Characterization of the cmcH genes of Nocardia lactamdurans and streptomyces clavuligerus encoding a functional 3′-hydroxymethylcephem O-carbamoyltransferase for cephamycin biosynthesis. Gene 1995, 162, 21–27. [Google Scholar] [CrossRef]
- Forzi, L.; Sawers, R.G. Maturation of [NiFe]-hydrogenases in Escherichia coli. Biometals 2007, 20, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Leach, M.R.; Zamble, D.B. Metallocenter assembly of the hydrogenase enzymes. Curr. Opin. Chem. Biol. 2007, 11, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Shima, S.; Thauer, R.K. A third type of hydrogenase catalyzing H2 activation. Chem. Rec. 2007, 7, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Cunin, R.; Glansdorff, N.; Pierard, A.; Stalon, V. Biosynthesis and metabolism of arginine in bacteria. Microbiol. Rev. 1986, 50, 314–352. [Google Scholar] [PubMed]
- Griswold, A.R.; Jameson-Lee, M.; Burne, R.A. Regulation and physiologic significance of the agmatine deiminase system of Streptococcus mutans UA159. J. Bacteriol. 2006, 188, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Ruepp, A.; Soppa, J. Fermentative arginine degradation in Halobacterium salinarium (formerly Halobacterium halobum): Gene products, and transcripts of the arcRACB gene cluster. Plant Physiol. 1993, 101, 429–434. [Google Scholar]
- Schofield, P.J.; Edwards, M.R.; Matthews, J.; Wilson, J.R. The pathway of arginine catabolism in giardia intestinalis. Mol. Biochem. Parasitol. 1992, 51, 29–36. [Google Scholar] [CrossRef]
- Yarlett, N.; Lindmark, D.G.; Goldberg, B.; Moharrami, M.A.; Bacchi, C.J. Subcellular localization of the enzymes of the arginine dihydrolase pathway in Trichomonas vaginalis and Tritrichomonas foetus. J. Eukaryot. Microbiol. 1994, 41, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Schofield, P.J.; Costello, M.; Edwards, M.R.; O’Sullivan, W.J. The arginine dihydrolase pathway is present in Giardia intestinalis. Int. J. Parasitol. 1990, 20, 697–699. [Google Scholar] [CrossRef]
- Wang, Q.; Xia, J.; Guallar, V.; Krilov, G.; Kantrowitz, E.R. Mechanism of thermal decomposition of carbamoyl phosphate and its stabilization by aspartate and ornithine transcarbamoylases. Proc. Natl. Acad. Sci. USA 2008, 105, 16918–16923. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.M., Jr.; Jones, M.E. Decomposition of carbamylphosphate in aqueous solutions. Biochemistry 1964, 3, 1238–1247. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.M.; Cockrell, G.M.; Puleo, D.E.; Kantrowitz, E.R. Crystallographic snapshots of the complete catalytic cycle of the unregulated aspartate transcarbamoylase from Bacillus subtilis. J. Mol. Biol. 2011, 411, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ramos, A.; Velazquez-Campoy, A.; Grande-Garcia, A.; Moreno-Morcillo, M.; Ramon-Maiques, S. Structure and functional characterization of human aspartate transcarbamoylase, the target of the anti-tumoral drug PALA. Structure 2016, 24, 1081–1094. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Morizono, H.; Yu, X.; Tong, L.; Allewell, N.M.; Tuchman, M. Human ornithine transcarbamylase: Crystallographic insights into substrate recognition and conformational changes. Biochem. J. 2001, 354, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Yu, X.; Roth, L.; Morizono, H.; Tuchman, M.; Allewell, N.M. Structures of N-acetylornithine transcarbamoylase from Xanthomonas campestris complexed with substrates and substrate analogs imply mechanisms for substrate binding and catalysis. Proteins 2006, 64, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Penverne, B.; Belkaid, M.; Herve, G. In situ behavior of the pyrimidine pathway enzymes in Saccharomyces cerevisiae 4. The channeling of carbamylphosphate to aspartate transcarbamylase and its partition in the pyrimidine and arginine pathways. Arch. Biochem. Biophys. 1994, 309, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, M.; Penverne, B.; Herve, G. In situ behavior of the pyrimidine pathway enzymes in Saccharomyces cerevisiae: 3. Catalytic and regulatory properties of carbamylphosphate synthetase: Channeling of carbamylphosphate to aspartate transcarbamylase. Arch. Biochem. Biophys. 1988, 262, 171–180. [Google Scholar] [CrossRef]
- Williams, L.G.; Bernhardt, S.; Davis, R.H. Copurification of pyrimidine-specific carbamyl phosphate synthetase and aspartate transcarbamylase of Neurospora crassa. Biochemistry 1970, 9, 4329–4335. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.G.; Bernhardt, S.A.; Davis, R.H. Evidence for two discrete carbamyl phosphate pools in neurospora. J. Biol. Chem. 1971, 246, 973–978. [Google Scholar] [PubMed]
- Christopherson, R.I.; Jones, M.E. The overall synthesis of L-5,6-dihydroorotate by multienzymatic protein pyr1-3 from hamster cells. Kinetic studies, substrate channeling, and the effects of inhibitors. J. Biol. Chem. 1980, 255, 11381–11395. [Google Scholar] [PubMed]
- Irvine, H.S.; Shaw, S.M.; Paton, A.; Carrey, E.A. A reciprocal allosteric mechanism for efficient transfer of labile intermediates between active sites in cad, the mammalian pyrimidine-biosynthetic multienzyme polypeptide. Eur. J. Biochem. 1997, 247, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.S.; Cheung, C.W.; Sijuwade, E.; Raijman, L. Kinetic properties of carbamoyl-phosphate synthase (ammonia) and ornithine carbamoyltransferase in permeabilized mitochondria. Biochem. J. 1992, 282, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Wanders, R.J.; Van Roermund, C.W.; Meijer, A.J. Analysis of the control of citrulline synthesis in isolated rat-liver mitochondria. Eur. J. Biochem. 1984, 142, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Van de Casteele, M.; Legrain, C.; Desmarez, L.; Chen, P.G.; Pierard, A.; Glansdorff, N. Molecular physiology of carbamoylation under extreme conditions: What can we learn from extreme thermophilic microorganisms? Comp. Biochem. Physiol. Part A Physiol. 1997, 118, 463–473. [Google Scholar] [CrossRef]
- Purcarea, C.; Ahuja, A.; Lu, T.; Kovari, L.; Guy, H.I.; Evans, D.R. Aquifex aeolicus aspartate transcarbamoylase, an enzyme specialized for the efficient utilization of unstable carbamoyl phosphate at elevated temperature. J. Biol. Chem. 2003, 278, 52924–52934. [Google Scholar] [CrossRef] [PubMed]
- Loffler, M.; Fairbanks, L.D.; Zameitat, E.; Marinaki, A.M.; Simmonds, H.A. Pyrimidine pathways in health and disease. Trends Mol. Med. 2005, 11, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Monks, A.; Chisena, C.A.; Cysyk, R.L. Influence of ammonium ions on hepatic de novo pyrimidine biosynthesis. Arch. Biochem. Biophys. 1985, 236, 1–10. [Google Scholar] [CrossRef]
- Wendler, P.A.; Blanding, J.H.; Tremblay, G.C. Interaction between the urea cycle and the orotate pathway: Studies with isolated hepatocytes. Arch. Biochem. Biophys. 1983, 224, 36–48. [Google Scholar] [CrossRef]
- Wang, Z.; Nicholls, S.J.; Rodriguez, E.R.; Kummu, O.; Horkko, S.; Barnard, J.; Reynolds, W.F.; Topol, E.J.; DiDonato, J.A.; Hazen, S.L. Protein carbamylation links inflammation, smoking, uremia and atherogenesis. Nat. Med. 2007, 13, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Kraus, L.M.; Kraus, A.P., Jr. Carbamoylation of amino acids and proteins in uremia. Kidney Int. Suppl. 2001, 78, S102–S107. [Google Scholar] [CrossRef] [PubMed]
- Sirpal, S. Myeloperoxidase-mediated lipoprotein carbamylation as a mechanistic pathway for atherosclerotic vascular disease. Clin. Sci. 2009, 116, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Manjeshwar, S.; Sheikh, A.; Pichiri-Coni, G.; Coni, P.; Rao, P.M.; Rajalakshmi, S.; Pediaditakis, P.; Michalopoulos, G.; Sarma, D.S. Orotic acid, nucleotide-pool imbalance, and liver-tumor promotion: A possible mechanism for the mitoinhibitory effects of orotic acid in isolated rat hepatocytes. Cancer Res. 1992, 52, 2078s–2081s. [Google Scholar] [PubMed]
- Wilson, J.M.; Shchelochkov, O.A.; Gallagher, R.C.; Batshaw, M.L. Hepatocellular carcinoma in a research subject with ornithine transcarbamylase deficiency. Mol. Genet. Metab. 2012, 105, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Hu, Z.; Cai, L.; Li, K.; Choi, E.; Faubert, B.; Bezwada, D.; Rodriguez-Canales, J.; Villalobos, P.; Lin, Y.F.; et al. CPS1 maintains pyrimidine pools and DNA synthesis in KRAS/LKB1-mutant lung cancer cells. Nature 2017, 546, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Celiktas, M.; Tanaka, I.; Tripathi, S.C.; Fahrmann, J.F.; Aguilar-Bonavides, C.; Villalobos, P.; Delgado, O.; Dhillon, D.; Dennison, J.B.; Ostrin, E.J.; et al. Role of CPS1 in cell growth, metabolism and prognosis in LKB1-inactivated lung adenocarcinoma. J. Natl. Cancer Inst. 2017, 109, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Haberle, J.; Shchelochkov, O.A.; Wang, J.; Katsonis, P.; Hall, L.; Reiss, S.; Eeds, A.; Willis, A.; Yadav, M.; Summar, S.; et al. Molecular defects in human carbamoy phosphate synthetase I: Mutational spectrum, diagnostic and protein structure considerations. Hum. Mutat. 2011, 32, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Ah Mew, N.; Caldovic, L. N-acetylglutamate synthase deficiency: An insight into the genetics, epidemiology, pathophysiology, and treatment. Appl. Clin. Genet. 2011, 4, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Van Karnebeek, C.D.; Sly, W.S.; Ross, C.J.; Salvarinova, R.; Yaplito-Lee, J.; Santra, S.; Shyr, C.; Horvath, G.A.; Eydoux, P.; Lehman, A.M.; et al. Mitochondrial carbonic anhydrase VA deficiency resulting from CA5A alterations presents with hyperammonemia in early childhood. Am. J. Hum. Genet. 2014, 94, 453–461. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, D.; Caldovic, L.; Tuchman, M. Sources and Fates of Carbamyl Phosphate: A Labile Energy-Rich Molecule with Multiple Facets. Biology 2018, 7, 34. https://doi.org/10.3390/biology7020034
Shi D, Caldovic L, Tuchman M. Sources and Fates of Carbamyl Phosphate: A Labile Energy-Rich Molecule with Multiple Facets. Biology. 2018; 7(2):34. https://doi.org/10.3390/biology7020034
Chicago/Turabian StyleShi, Dashuang, Ljubica Caldovic, and Mendel Tuchman. 2018. "Sources and Fates of Carbamyl Phosphate: A Labile Energy-Rich Molecule with Multiple Facets" Biology 7, no. 2: 34. https://doi.org/10.3390/biology7020034
APA StyleShi, D., Caldovic, L., & Tuchman, M. (2018). Sources and Fates of Carbamyl Phosphate: A Labile Energy-Rich Molecule with Multiple Facets. Biology, 7(2), 34. https://doi.org/10.3390/biology7020034