Odor Communication and Mate Choice in Rodents

{kind=link}
Abstract
:1. Introduction and Overview
1.1. Scent Marks as Indicators of Nutritional Status of Condition of Potential Mates
1.2. Scent Marks as Indicators of Age of Potential Mates
1.3. Genotype, Scent Marks and Mate Choice
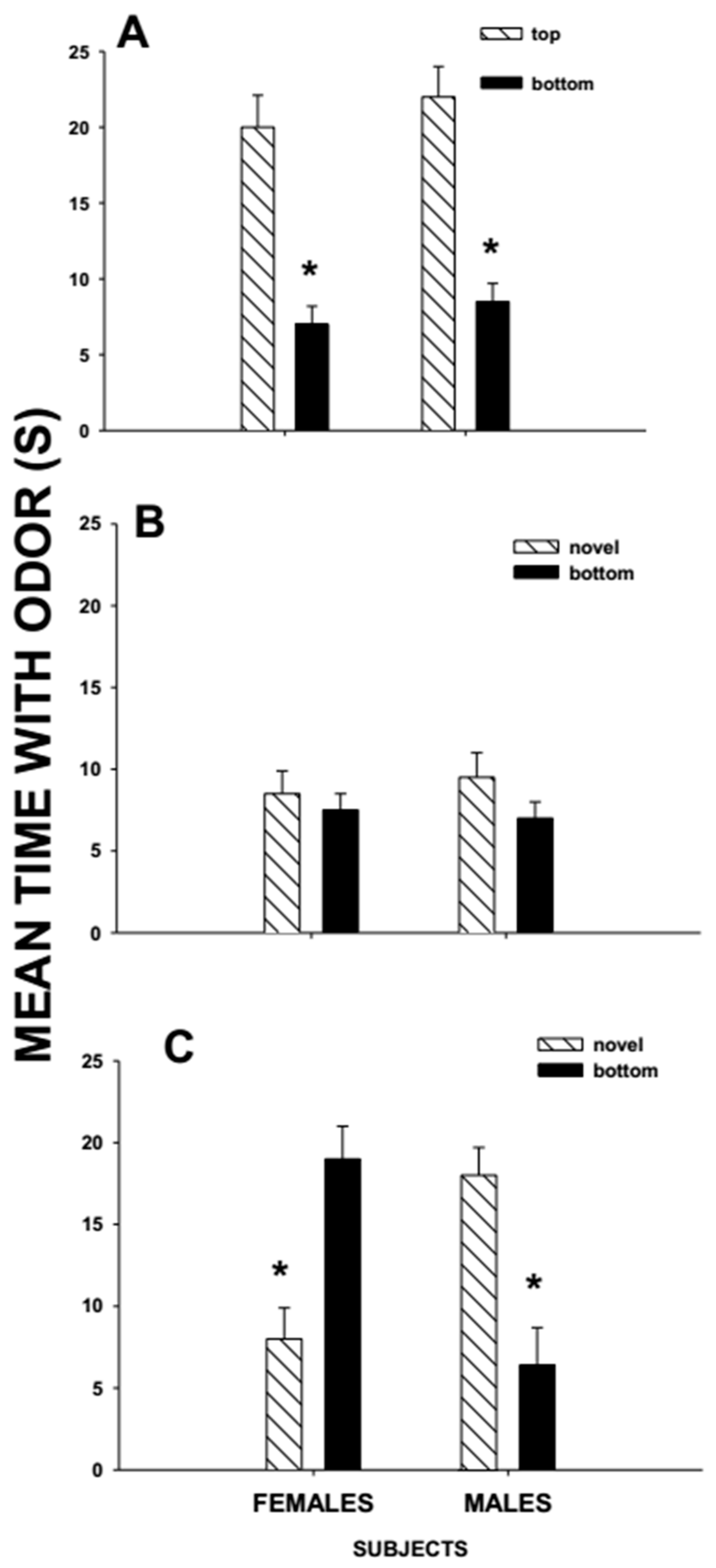
1.4. Over-Marking and Mate Choice
The Number of Potential Mates Affects the Responses to Over-Marks
1.5. Self-Grooming and Mate Choice
1.6. Sequential Mating and Mate Choice
1.7. Mate Choice and Public Information
2. Conclusions
Acknowledgments
Conflicts of Interest
References
- Johnston, R.E.; delBarco-Trillo, J. Communication by chemical signals: Behavior, Social recognition, hormones and the role of the vomeronasal and olfactory systems. In Hormones, Brain and Behavior; Pffaf, D., Arnold, A.P., Fahrbach, S.E., Etgen, A.M., Rubin, R.T., Eds.; Elsevier Science: New York, NY, USA, 2009; Volume 2, pp. 797–870. [Google Scholar]
- Ferkin, M.H.; delBarco-Trillo, J.; Petrulis, A. Communication by chemical signals: Physiological mechanisms, ontogeny and learning, function, evolution and cognition. In Hormones, Brain and Behavior, 3rd ed.; Pfaff, D.W., Joëls, M., Eds.; Elsevier Press: Oxfordshire, UK, 2017; pp. 285–327. [Google Scholar]
- Brown, R.E.; Macdonald, D.W. Social Odours in Mammals; Clarendon Press: Oxford, UK, 1985. [Google Scholar]
- Ferkin, M.H. The response of rodents to scent marks: Four broad hypotheses. Horm. Behav. 2015, 68, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Johnston, R.E. Chemical signals and reproductive behavior. In Pheromones and Reproduction in Mammals; Vandenbergh, J.G., Ed.; Academic Press: New York, NY, USA, 1983; pp. 3–37. [Google Scholar]
- Kavaliers, M.; Wiebe, J.P.; Galea, L.A. Male preference for the odors of estrous female mice is enhanced by the neurosteroid 3 alpha-hydroxy-4-pregnen-20-one (3-alpha HP). Brain Res. 1994, 646, 140–144. [Google Scholar] [CrossRef]
- delBarco-Trillo, J.; LaVenture, A.B.; Johnston, R.E. Male hamsters discriminate estrous state from vaginal secretions and individuals from flank marks. Behav. Proc. 2009, 82, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Kimura, T. Sex-attractant emitted by female mice. Physiol. Behav. 1974, 13, 563–567. [Google Scholar] [CrossRef]
- Huck, U.W.; Banks, E.M. Social olfaction in male brown lemmings (Lemmus sibiricus trimucronatus) and collared lemmings (Dicrostonyx groenlandicus): I. Discrimination of species, sex and estrous condition. J. Comp. Psychol. 1984, 98, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Ferkin, M.H.; Johnston, R.E. Effects of pregnancy, lactation and postpartum oestrus on odour signals and the attraction to odours in female meadow voles, Microtus pennsylvanicus. Anim. Behav. 1995, 49, 1211–1217. [Google Scholar] [CrossRef]
- Kumari, S.; Prakash, I. Perception of oestrous odours by male Meriones hurrianae. Anim. Behav. 1984, 32, 927–929. [Google Scholar] [CrossRef]
- Block, M.L.; Volpe, L.C.; Hayes, M.J. Saliva as a chemical cue in the development of social behavior. Science 1981, 211, 1062–1064. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A.S.; Chee, P.M.; Vaccarino, F. Sexual behaviour and its olfactory control in the desert woodrat (Neotoma lepida lepida). Anim. Behav. 1981, 29, 727–745. [Google Scholar] [CrossRef]
- Harris, M.A.; Murie, J.O. Discrimination of oestrous status by scent in Columbian ground squirrels. Anim. Behav. 1984, 32, 939–940. [Google Scholar] [CrossRef]
- Dunbar, I.F. Olfactory preferences in dogs: The response of male and female Beagles to conspecific odors. Behav. Biol. 1977, 20, 471–481. [Google Scholar] [CrossRef]
- Lindsay, D.R. The importance of olfactory stimuli in the mating behaviour of the ram. Anim. Behav. 1965, 13, 75–78. [Google Scholar] [CrossRef]
- Converse, L.J.; Carlson, A.A.; Ziegler, T.E.; Snowdon, C.T. Communication of ovulatory state to mates by female pygmy marmosets, Cebuella pygmaea. Anim. Behav. 1995, 49, 615–621. [Google Scholar] [CrossRef]
- Huck, U.W.; Lisk, R.D.; Gore, A.C. Scent marking and mate choice in the golden hamster. Physiol. Behav. 1985, 35, 389–393. [Google Scholar] [CrossRef]
- Gilbert, A.N. Postpartum and lactational estrus: A comparative analysis in Rodentia. J. Comp. Psychol. 1984, 98, 232–245. [Google Scholar] [CrossRef] [PubMed]
- Ferkin, M.H.; delBarco-Trillo, J. The behavior of female meadow voles, Microtus pennsylvanicus, during postpartum estrus and the responses of males to them. Mamm. Biol. 2014, 79, 81–89. [Google Scholar] [CrossRef]
- Witt, D.M.; Carter, C.S.; Chayer, R.; Adams, K. Patterns of behavior during postpartum oestrus in prairie voles, Microtus ochrogaster. Anim. Behav. 1990, 39, 528–534. [Google Scholar] [CrossRef]
- Zeigler, T.E.; Epple, G.; Snowdon, C.T.; Porter, T.A.; Belcher, A.M.; Kuderling, I. Detection of the chemical signals of ovulation in the cotton-top tamarin, Saginus oedipus. Anim. Behav. 1993, 45, 313–322. [Google Scholar] [CrossRef]
- Ferkin, M.H.; Johnston, R.E. Meadow voles, Microtus pennsylvanicus, use multiple sources of scent for sex recognition. Anim. Behav. 1995, 49, 37–44. [Google Scholar] [CrossRef]
- Petrulis, A. Chemosignals, hormones and mammalian reproduction. Horm. Behav. 2013, 63, 723–741. [Google Scholar] [CrossRef] [PubMed]
- Petrulis, A. Chemosignals and hormones in the neural control of mammalian sexual behavior. Front. Neuroendocrinol. 2013, 34, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Ferkin, M.H.; Zucker, I. Seasonal control of odour preferences of meadow voles (Microtus pennsylvanicus) by photoperiod and ovarian hormones. J. Reprod. Fertil. 1991, 92, 433–441. [Google Scholar] [CrossRef]
- Ferkin, M.H.; Johnston, R.E. Roles of gonadal hormones in control of five sexually attractive odors of meadow voles (Microtus pennsylvanicus). Horm. Behav. 1993, 27, 523–538. [Google Scholar] [CrossRef] [PubMed]
- Ferkin, M.H.; Gorman, M.R.; Zucker, I. Gonadal hormones influence odor cues emitted by male meadow voles, Microtus pennsylvanicus. J. Reprod. Fertil. 1992, 95, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.-C.; Vasilieva, N.Y.; Johnston, R.E. Odors providing sexual information in Djungarian hamsters: Evidence for an across-odor code. Horm. Behav. 1996, 30, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Leonard, S.T.; Alizadeh-Naderi, R.; Stokes, K.; Ferkin, M.H. The role of prolaction and testosterone in mediating seasonal differences in the self-grooming behavior of male meadow voles, Microtus pennsylvanicus. Physiol. Behav. 2005, 85, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Wade, G.N.; Schneider, J.E. Metabolic fuels and reproduction in female mammals. Neurosci. Biobehav. Rev. 1992, 16, 235–272. [Google Scholar] [CrossRef]
- Wade, G.N.; Schneider, J.E.; Li, H.-Y. Control of fertility by metabolic cues. Am. J. Physiol. 1996, 270, E1–E19. [Google Scholar] [CrossRef] [PubMed]
- Gill, C.J.; Rissman, E.F. Female sexual behavior is inhibited by short and long-term food restriction. Physiol. Behav. 1997, 61, 387–394. [Google Scholar] [CrossRef]
- Temple, J.L.; Schneider, J.E.; Scott, D.K.; Korutz, A.; Rissman, E.F. Mating behavior is controlled by acute changes in metabolic fuels. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 282, R782–R790. [Google Scholar] [CrossRef] [PubMed]
- Klingerman, C.M.; Patel, A.M.; Hedges, V.L.; Meisel, R.I.; Schneider, J.E. Food restriction dissociates sexual motivation, sexual performance and the rewarding consequences of copulation in female hamsters. Behav. Brain Res. 2011, 223, 356–370. [Google Scholar] [CrossRef] [PubMed]
- Sabau, R.M.; Ferkin, M.H. Food deprivation and restriction during late gestation affect the sexual behavior of postpartum female meadow voles, Microtus pennsylvanicus. Ethology 2013, 119, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Sabau, RM.; Ferkin, M.H. Food restriction affects the maternal behavior provided by female meadow voles (Microtus pennsylvanicus). J. Mammal. 2013, 94, 1068–1076. [Google Scholar] [CrossRef]
- Schneider, J.E.; Wise, J.D.; Benton, N.A.; Brozek, J.M.; Keen-Rhinehart, E. When do we eat? Ingestive behavior, survival and reproductive success. Horm. Behav. 2013, 64, 702–728. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, R.; McColl, C.J.; Karels, T.J. Reproduction at all costs: How breeding compromises the stress response and survival in male Arctic ground squirrels. Ecology 2001, 82, 1930–1946. [Google Scholar] [CrossRef]
- Dickman, C.R.; Braithwaite, R.W. Postmating mortality of males in the dasyurid marsupials, Dasyurus and Parantechinus. J. Mammal. 1992, 73, 143–147. [Google Scholar] [CrossRef]
- Trivers, R.L. Parental investment and sexual selection. In Sexual Selection and the Descent of Man; Campbell, B., Ed.; Heinemann Press: London, UK, 1972; pp. 136–179. [Google Scholar]
- Ferkin, M.H. The effects of food availability on the sexual behavior of meadow voles. Integr. Comp. Biol. 2017, 57, 1240–1244. [Google Scholar] [CrossRef] [PubMed]
- Zala, S.M.; Potts, W.K.; Penn, D.J. Scent-marking displays provide honest signals of health and infection. Behav. Ecol. 2004, 15, 338–344. [Google Scholar] [CrossRef]
- Roberts, S.C. Scent marking. In Rodent Societies: An Ecological and Evolutionary Perspective; Wolff, J.O., Sherman, P.W., Eds.; The University of Chicago Press: Chicago, IL, USA, 2007; pp. 255–266. [Google Scholar]
- Hobbs, N.J.; Ferkin, M.H. Effect of protein content of the diet on scent marking and over-marking behavior in meadow voles, Microtus pennsylvanicus. Behaviour 2011, 148, 1027–1044. [Google Scholar] [CrossRef]
- Hobbs, N.J.; Ferkin, M.H. Dietary protein content affects the responses of meadow voles, Microtus pennsylvanicus, to over-marks. Acta Ethol. 2011, 14, 57–64. [Google Scholar] [CrossRef]
- Pierson, L.M.; Ferkin, M.H. The impact of phytoestrogens on sexual behavior and cognition in rodents. Mammal. Biol. 2015, 80, 148–154. [Google Scholar] [CrossRef]
- Pierce, A.A.; Ferkin, M.H.; Williams, T.K. Food-deprivation-induced changes in sexual behavior of meadow voles, Microtus pennsylvanicus. Anim. Behav. 2005, 70, 339–348. [Google Scholar] [CrossRef]
- Pierce, A.A.; Iwueke, I.; Ferkin, M.H. Food deprivation and the role of estradiol in mediating sexual behaviors in meadow voles. Physiol. Behav. 2007, 90, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Appleby, M.C. Social rank and food access in red deer stags. Behaviour 1980, 74, 294–309. [Google Scholar] [CrossRef]
- Stears, K.; Kerley, G.I.H.; Shrader, A.M. Group-living herbivores weigh up food availability and dominance status when making patch-joining decisions. PLoS ONE 2014, 9, E109. [Google Scholar] [CrossRef] [PubMed]
- Meikle, D.B.; Kruper, J.H.; Browning, C.R. Adult male house mice born to undernourished mothers are unattractive to oestrous females. Anim. Behav. 1995, 50, 753–758. [Google Scholar] [CrossRef]
- Liang, H.; Zhang, J.; Zhang, Z. Food restriction in pregnant rat-like hamsters (Cricetulus triton) affects endocrine, immune function and odor attractiveness of male offspring. Physiol. Behav. 2004, 82, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Sabau, R.M.; Ferkin, M.H. Maternal food restriction during lactation affects body weight and sexual behavior of male offspring in meadow voles (Microtus pennsylvanicus). Ethology 2014, 120, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Sabau, R.M.; Pierson, L.M.; Ferkin, M.H. Body mass and attractivity of female offspring are negatively affected by food restriction of meadow vole (Microtus pennsylvanicus) dams during lactation. Behaviour 2014, 151, 1847–1868. [Google Scholar] [CrossRef]
- Harriman, A.E.; Thiessen, D.D. Harderian letdown in male Mongolian gerbils (Meriones unguiculatus) contributes to proceptive behavior. Horm. Behav. 1986, 19, 213–219. [Google Scholar] [CrossRef]
- Ferkin, M.H.; Leonard, S.T. Self-grooming as a form of olfactory communication in meadow voles and prairie voles (Microtus spp.). In Neurobiology of Grooming Behavior; Kalueff, A.V., LaPorte, J.L., Bergner, C., Eds.; Cambridge University Press: Oxford, UK, 2010; pp. 19–47. [Google Scholar]
- Hobbs, N.J.; Ferkin, M.H. Effects of food availability on proceptivity: A test of the reproduction at all costs and metabolic fuels hypotheses. Behav. Proc. 2012, 91, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Osada, K.; Kashiwayanagi, M.; Izumi, H. Profiles of volatiles in male rat urine: The effect of puberty on the female attraction. Chem. Senses 2009, 34, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Ferkin, M.H. Attractiveness of opposite-sex odor and responses to it vary with age and sex in meadow voles (Microtus pennsylvanicus). J. Chem. Ecol. 1999, 25, 757–769. [Google Scholar] [CrossRef]
- Garratt, M.; Stockley, P.; Armstrong, S.D.; Beynon, R.J.; Hurst, J.L. The scent of senescence: Sexual signaling and female preference in house mice. J. Evol. Biol. 2011, 24, 2398–2409. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L.; Kamath, V. The influences of age on olfaction: A review. Front. Psychol. 2014, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Mobley, A.S.; Rodriguez-Gil, D.J.; Imamura, F.; Greer, C.A. Aging in the olfactory system. Trends Neurosci. 2014, 37, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Ferkin, M.H.; Vlautin, C.T.; Pierson, L.M. Age-related effects on individual discrimination among meadow voles, Microtus pennsylvanicus. In Chemical Signals in Vertebrates; Schulte, B.A., Goodwin, T.E., Ferkin, M.H., Eds.; Springer International Publishing Co. AG: Cham, Switzerland, 2016; Volume 13, pp. 121–136. [Google Scholar]
- Hurst, J.L.; Payne, C.E.; Nevison, C.M.; Marie, A.D.; Humphries, R.E.; Robertson, D.H.L.; Cavaggioni, A.; Beynon, R.J. Individual recognition in mice mediated by major urinary proteins. Nature 2001, 414, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Baena, G.; Armstrong, S.D.; Phelan, M.M.; Hurst, J.L.; Beynon, R.J. The major urinary protein system in the rat. Biochem. Soc. Trans. 2014, 42, 886–892. [Google Scholar] [CrossRef] [PubMed]
- Cheetham, S.A.; Thom, M.D.; Jury, F.; Ollier, W.E.R.; Beynon, R.J.; Hurst, J.L. The genetic basis of individual-recognition signals in the mouse. Curr. Biol. 2007, 17, 1771–1777. [Google Scholar] [CrossRef] [PubMed]
- Green, J.P.; Holmes, A.M.; Davidson, A.J.; Paterson, S.; Stockley, P.; Beynon, R.J.; Hurst, J.L. The genetic basis of kin recognition in a cooperatively breeding mammal. Curr. Biol. 2015, 25, 2631–2641. [Google Scholar] [CrossRef] [PubMed]
- Kaur Angeldeep, W.; Ackels, T.; Kuo, T.-H.; Cichy, A.; Dey, S.; Hays, C.; Kateri, M.; Logan, D.W.; Marton, T.F.; Spehr, M.; et al. Murine pheromone proteins constitute a context-dependent combinatorial code governing multiple social behaviors. Cell 2014, 157, 676–688. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.A.; Simpson, D.M.; Armstrong, S.D.; Davidson, A.J.; Robertson, D.H.; McLean, L.; Beynon, R.J.; Hurst, J.L. Darcin: A male pheromone that stimulates female memory and sexual attraction to an individual male’s odour. BMC Biol. 2010, 8, 75. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.A.; Davidson, S.J.; Beynon, R.J.; Hurst, J.L. Female attraction to mate scent and associative learning: The house mouse as a mammalian model. Anim. Behav. 2014, 97, 313–321. [Google Scholar] [CrossRef]
- Thoß, M.; Luzynski, K.; Ante, M.; Miller, I.; Penn, D.J. Major urinary protein (MUP) profiles show dynamic changes rather than individual “barcode” signatures. Front. Ecol. Evol. 2015, 3, 71. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.; Macrides, F.; Clancy, A.N.; Agosta, W.C. Purification and analysis of a proteinaceous aphrodisiac pheromone from hamster vaginal discharge. J. Biol. Chem. 1986, 261, 13323–13326. [Google Scholar] [PubMed]
- Macrides, F.; Clancy, A.N.; Singer, A.G.; Agosta, W.C. Male hamster investigatory and copulatory responses to vaginal discharge: An attempt to impart sexual significance to an arbitrary chemosensory stimulus. Physiol. Behav. 1986, 33, 627–632. [Google Scholar] [CrossRef]
- Stopková, R.; Zdráhal, Z.; Ryba, S.; Šedo, O.; Šandera, M.; Stopka, P. Novel OBP genes similar to hamster Aphrodisin in the bank vole, Myodes glareolus. BMC Genom. 2010, 11, 45. [Google Scholar] [CrossRef] [PubMed]
- Penn, D.J.; Potts, W.K. The evolution of mating preferences and major histocompatibility complex genes. Am. Nat. 1999, 153, 145–164. [Google Scholar] [CrossRef]
- Penn, D.; Potts, W.K. Chemical signals and parasite-mediated sexual selection. Trends Ecol. Evol. 1998, 13, 391–396. [Google Scholar] [CrossRef]
- Potts, W.K.; Wakeland, E.K. Evolution of MHC genetic diversity: A tale of incest, pestilence and sexual preference. Trends Genet. 1993, 9, 408–412. [Google Scholar] [CrossRef]
- Brown, J.L.; Eklund, A. Kin recognition and the major histocompatibility complex: An integrative review. Am. Nat. 1994, 143, 435–461. [Google Scholar] [CrossRef]
- Yamazaki, K.; Yamaguchi, M.; Boyse, E.A.; Thomas, L. The major histocompatibility complex as a source of odors imparting individuality among mice. In Chemical Signals: Vertebrates and Aquatic Invertebrates; Müller-Schwarze, D., Silverstein, R.M., Eds.; Plenum Press: New York, NY, USA, 1980; pp. 267–273. [Google Scholar]
- Brown, R.E.; Roser, B.; Singh, P.B. The MHC and individual odors in rats. In Chemical Signals in Vertebrates; Macdonald, D.W., Müller-Schwarze, D., Natynczuk, S.E., Eds.; Oxford University Press: New York, NY, USA, 1990; Volume 5, pp. 228–243. [Google Scholar]
- Egid, K.; Brown, J.L. The major histocompatibility complex and female mating preferences in mice. Anim. Behav. 1989, 38, 548–549. [Google Scholar] [CrossRef]
- Wedekind, C.; Seebeck, T.; Bettens, F.; Paepke, A.J. MHC-Dependent mate preferences in humans. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1995, 260, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Sherborne, A.L.; Thom, M.D.; Paterson, S.; Jury, F.; Ollier, W.E.R.; Stockley, P.; Beynon, R.J.; Hurst, J.L. The genetic basis of inbreeding avoidance in house mice. Curr. Biol. 2007, 17, 2061–2066. [Google Scholar] [CrossRef] [PubMed]
- Johnston, R.E.; Chiang, G.; Tung, C. The information in scent over-marks of golden hamsters. Anim. Behav. 1994, 48, 323–330. [Google Scholar] [CrossRef]
- Kappeler, P.M. To whom it may concern: The transmission and function of chemical signals in Lemur catta. Behav. Ecol. Sociobiol. 1998, 42, 411–421. [Google Scholar] [CrossRef]
- Ferkin, M.H.; Pierce, A.A. Perspectives on over-marking: Is it good to be on top? J. Ethol. 2007, 25, 107–116. [Google Scholar] [CrossRef]
- Hurst, J.L.; Beynon, R.J. Scent wars: The chemobiology of competitive signalling in mice. Bioessays 2004, 26, 1288–1298. [Google Scholar] [CrossRef] [PubMed]
- Ferkin, M.H.; Hobbs, N.J.; Ferkin, B.D.; Ferkin, A.C.; Ferkin, D.A. Male and female meadow voles, Microtus pennsylvanicus, respond differently to scent marks from the top- middle- and bottom-scent donors of an over-mark. Curr. Zool. 2011, 57, 441–447. [Google Scholar] [CrossRef]
- Johnston, R.E.; Munver, R.; Tung, C. Scent counter marks: Selective memory for the top scent by golden hamsters. Anim. Behav. 1995, 49, 1435–1442. [Google Scholar] [CrossRef]
- Woodward, R.L.; Schmick, M.K.; Ferkin, M.H. Response of prairie voles, Microtus ochrogaster (Rodentia, Arvicolidae), to scent over-marks of two same-sex conspecifics: A test of the scent masking hypothesis. Ethology 1999, 105, 1009–1017. [Google Scholar] [CrossRef]
- Johnston, R.E. Chemical communication in rodents: From pheromones to individual recognition. J. Mammal. 2003, 84, 1141–1162. [Google Scholar] [CrossRef]
- Ferkin, M.H. Odor-related behavior and cognition in meadow voles, Microtus pennsylvanicus (Arvicolidae, Rodentia). Folia Zool. 2011, 60, 262–276. [Google Scholar] [CrossRef]
- Leonard, S.T.; Ferkin, M.H.; Johnson, M.M. The response of meadow voles to an over-mark in which the two donors differ in gonadal hormone status. Anim. Behav. 2001, 62, 1171–1177. [Google Scholar] [CrossRef]
- Hobbs, N.J.; Ferkin, M.H. The response of male meadow voles, Microtus pennsylvanicus, to same- and mixed- sex over-marks depends on the reproductive state of the top- and bottom-female scent donors. Behaviour 2012, 149, 705–722. [Google Scholar] [CrossRef]
- Ferkin, M.H.; Ferkin, D.A.; Ferkin, B.D.; Vlautin, C.T. Olfactory experience affects the response of meadow voles to the opposite-sex scent donor of mixed-sex over-marks. Ethology 2010, 116, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Hurst, J.L. Urine marking in populations of wild house mice Mus domesticus Rutty. III. Communication between the sexes. Anim. Behav. 1990, 40, 233–243. [Google Scholar] [CrossRef]
- Woodward, R.L.; Bartos, K.; Ferkin, M.H. Meadow voles (Microtus pennsylvanicus) and prairie voles (M. ochrogaster) differ in their responses to over-marks from opposite- and same-sex conspecifics. Ethology 2000, 106, 979–992. [Google Scholar] [CrossRef]
- Arakawa, H.; Blanchard, D.C.; Blanchard, R.J. Central oxytocin regualtes social familiarity and scent marking behavior that involves amicable odor signals between male mice. Physiol. Behav. 2015, 146, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Dröscher, I.; Kappeler, P.M. Maintenance of familiarity and social bonding via communal latrine use in a solitary primate (Lepilemur leucopus). Behav. Ecol. Sociobiol. 2014, 68, 2043–2058. [Google Scholar] [CrossRef] [PubMed]
- Ferkin, M.H.; Briley, D.; Ferkin, B.D.; Hardaway, A.; Applebury, T. Responses of meadow voles, Microtus pennsylvanicus, to areas containing over-marks and single scent marks of two opposite-sex conspecifics. Behaviour 2011, 148, 117–130. [Google Scholar] [CrossRef]
- Ferkin, M.H.; Pierce, A.A.; Sealand, R.O.; delBarco-Trillo, J. Meadow voles, Microtus pennsylvanicus, can distinguish more over-marks from fewer over-marks. Anim. Cognit. 2005, 8, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, A.A.; Ferkin, M.H. The presence and number of male competitor’s scent marks and female reproductive state affects the response of male meadow voles to female conspecifics’ odours. Behaviour 2011, 48, 927–943. [Google Scholar] [CrossRef]
- Ferkin, M.H.; Hobbs, N.J. Female meadow voles, Microtus pennsylvanicus, respond differently to the scent marks of multiple male conspecifics. Anim. Cognit. 2014, 17, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Ferkin, M.H.; Li, H.Z.; Leonard, S.T. Meadow voles and prairie voles differ in the percentage of conspecific marks that they over-mark. Acta Ethol. 2004, 7, 1–7. [Google Scholar] [CrossRef]
- Thiessen, D.D. Thermoenergetics and the evolution of pheromone communication. In Progress in Psychobiology and Physiological Psychology; Sprague, J.M., Epstein, A.N., Eds.; Academic Press: New York, NY, USA, 1977; Volume 7, pp. 91–191. [Google Scholar]
- Wiepkema, P.R. The social significance of self grooming in rats. Neth. J. Zool. 1979, 29, 622–623. [Google Scholar]
- Achiraman, S.; Ganesh, D.S.; Kannan, S.; Kamalakkannan, S.; Nimala, N.; Archunan, G. Response of male mice to odours of female mice in different stages of oestrous cycle: Self-grooming behaviour and the effect of castration. Indian J. Exp. Biol. 2014, 52, 30–35. [Google Scholar] [PubMed]
- Yu, H.; Zhao, X.; Yue, P.; Sun, P. Chemical communication in mammal populations: Urinary olfactory chemosignals in lactating female root voles (Microtus oeconomus pallas). Pol. J. Ecol. 2010, 58, 153–165. [Google Scholar]
- Hobbs, N.L.; Aven, A.M.; Ferkin, M.H. Self-grooming responses of meadow voles to the odors of opposite-sex conspecifics in relation to the dietary protein content of both sexes. Ethology 2008, 14, 1210–1217. [Google Scholar] [CrossRef]
- Paz-y-Miño, G.C.; Leonard, C.T.; Ferkin, M.H.; Trimble, J.F. Self-grooming and sibling recognition in meadow voles, Microtus pennsylvanicus and prairie voles, M. ochrogaster. Anim. Behav. 2002, 63, 331–338. [Google Scholar] [CrossRef]
- Huck, U.W.; Banks, E.M.; Coopersmith, C.B. Social olfaction in male brown lemmings (Lemmus sibiricus = trimucronatus) and collared lemmings (Dicrostonyx groenlandicus): II. Discrimination of mated and unmated females. J. Comp. Physiol. Psychol. 1984, 98, 60–65. [Google Scholar] [CrossRef]
- Dewsbury, D.A.; Ferguson, B.; Hodges, A.W.; Taylor, S.A. Tests of preferences of deer mice (Peromyscus maniculatus bairdi) for individuals and their odors as a function of sex and estrous condition. J. Comp. Psychol. 1986, 100, 117–127. [Google Scholar] [CrossRef]
- Jennions, M.D.; Petrie, M. Variation in mate choice and mating preferences: A review of causes and consequences. Biol. Rev. 1997, 72, 283–327. [Google Scholar] [CrossRef] [PubMed]
- Bonduriansky, R. The evolution of male mate choice in insects: A synthesis of ideas and evidence. Biol. Rev. Camb. Philos. Soc. 2001, 76, 305–339. [Google Scholar] [CrossRef] [PubMed]
- Barry, K.L.; Kokko, H. Male mate choice: Why sequential choice can make its evolution difficult. Anim. Behav. 2010, 80, 163–169. [Google Scholar] [CrossRef]
- Birkhead, T.R. Promiscuity: An Evolutionary History of Sperm Competition; Harvard University Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Bateman, P.W.; Fleming, P.S. Males are selective too: Mating but not courtship with sequential females influences choosiness in male field crickets (Gryllus bimaculatas). Behav. Ecol. Sociobiol. 2006, 59, 577–581. [Google Scholar] [CrossRef]
- Head, M.L.; Jacomb, F.; Vega-Tejo, R.; Jennions, M.D. Male mate choice and insemination success under simultaneous versus sequential choice conditions. Anim. Behav. 2015, 103, 99–105. [Google Scholar] [CrossRef]
- Jennions, M.D.; Kokko, H. Sexual selection: Mate choice. In The Princeton Guide to Evolution; Losos, J., Baum, D., Futuyama, D.J., Hoekestra, H., Lenski, R., Moore, A., Eds.; Princeton University Press: Princeton, NJ, USA, 2014; pp. 647–654. [Google Scholar]
- Ferkin, M.H.; Ferkin, A.C. The time interval between sequential mating affects the response of male meadow voles to previous mates. Behaviour 2016, 153, 1931–1949. [Google Scholar]
- Wilson, J.R.; Kuehn, R.E.; Beach, F.A. Modification in the sexual behavior of male rats produced by changing the stimulus female. J. Comp. Physiol. Psychol. 1963, 56, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Stockley, P.; Preston, B.T. Sperm competition and diversity in rodent copulatory behaviour. J. Evol. Biol. 2004, 17, 1048–1057. [Google Scholar] [CrossRef] [PubMed]
- DelBarco-Trillo, J.; Ferkin, M.H. Risk of sperm competition does not influence copulatory behavior in the promiscuous meadow vole (Microtus pennsylvanicus). J. Ethol. 2007, 25, 139–145. [Google Scholar] [CrossRef]
- Berteaux, D.; Bety, J.; Rengifo, E.; Bergeron, J. Multiple paternity in meadow voles (Microtus pennsylvanicus): Investigating the role of the female. Behav. Ecol. Sociobiol. 1999, 45, 283–291. [Google Scholar] [CrossRef]
- DelBarco-Trillo, J.; Ferkin, M.H. Male mammals respond to a risk of sperm competition conveyed by odours of conspecific males. Nature 2004, 431, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, R.; Xia, X.; Pavone, L. Mating system of the meadow vole, Microtus pennsylvanicus. Behav. Ecol. 1993, 4, 83–89. [Google Scholar] [CrossRef]
- Spritzer, M.D.; Meikle, D.B.; Solomon, N.G. The relationship between dominance rank and spatial ability among meadow voles (Microtus pennsylvanicus). J. Comp. Psychol. 2004, 118, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Franklin, S.; Ferkin, M.H. Using broad cognitive models to apply computational intelligence to animal cognition. In Applications of Computational Intelligence in Biology: Current Trends and Open Problems; Smolinski, T.G., Milanova, M.M., Hassanien, A.-E., Eds.; Springer: Berlin, Germany, 2008; pp. 363–394. [Google Scholar]
- Nadeau, J.H. Ontogeny. In Biology of New World Microtus; Tamarin, R.H., Ed.; American Society of Mammalogists Special Publications: Lawrence, KS, USA, 1985; Volume 8, pp. 254–285. [Google Scholar]
- Gorman, M.R.; Ferkin, M.H.; Dark, J. Melatonin influences sex-specific prenatal mortality in meadow voles. Biol. Reprod. 1994, 51, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Insel, T.R.; Preston, S.; Winslow, J.T. Mating in the monogamous male: Behavioral consequences. Physiol. Behav. 1995, 57, 615–627. [Google Scholar] [CrossRef]
- Curtis, J.T. Does fertility trump monogamy? Anim. Behav. 2010, 80, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Storey, A. Behavioral interactions increase pregnancy blocking by unfamiliar male meadow voles. Physiol. Behav. 1996, 60, 1093–1098. [Google Scholar] [CrossRef]
- DeCatanzaro, D. Blastocyst implantation is vulnerable to stress-induced rises in endogenous estrogens and also to excretions of estrogens by proximate males. J. Reprod. Immunol. 2011, 90, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Ferkin, M.H.; Ferkin, A.C. The number of male conspecifics affects the odor preferences and the copulatory behavior of male meadow voles, Microtus pennsylvanicus. Behaviour 2017, 154, 413–433. [Google Scholar] [CrossRef]
- Ferkin, M.H.; Pierce, A.A.; Sealand, R.O. Gonadal hormones mediate sex differences in judgments of relative numerousness in meadow voles, Microtus pennsylvanicus. Horm. Behav. 2008, 55, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Ferkin, M.H. Information by chemical signals in rodents. In Chemical Signals in Vertebrates; Buesching, C.D., Müller, C.T., Eds.; Springer International Publishing AG: Cham, Switzerland, 2019; Volume 14, submitted. [Google Scholar]
- Dewsbury, D.A. Sperm competition in muroid rodents. In Sperm Competition and the Evolution of Animal Mating Systems; Smith, R.R., Ed.; Academic Press: New York, NY, USA, 1984; pp. 547–571. [Google Scholar]
- Ramm, S.A.; Stockley, P. Sequential male mate choice under sperm competition risk. Behav. Ecol. 2014, 25, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Witte, K.; Kniel, N.; Kureck, I.M. Mate-choice copying: Status quo and where to go. Curr. Biol. 2015, 61, 1073–1081. [Google Scholar] [CrossRef]
- Galef, B.G.; Lim, T.C.W.; Gilbert, G.S. Evidence of mate choice copying in Norway rats, Rattus norvegicus. Anim. Behav. 2008, 75, 1117–1123. [Google Scholar] [CrossRef]
- Kavaliers, M.; Matta, R.; Choleris, E. Mate-choice copying, social information processing and the roles of oxytocin. Neurosci. Biobehav. Rev. 2017, 72, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Giraldeau, L.-A.; Valone, T.J.; Templeton, J.J. Potential disadvantages of using socially acquired information. Philos. Trans. R. Soc. Lond. B 2002, 357, 1559–1566. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.A.; Pizzari, T. Sperm competition and ejaculate economics. Biol. Rev. Camb. Philos. Soc. 2010, 85, 897–934. [Google Scholar] [CrossRef] [PubMed]
- DelBarco-Trillo, J.; Ferkin, M.H. Male meadow voles respond differently to risk and intensity of sperm competition. Behav. Ecol. 2006, 17, 581–585. [Google Scholar] [CrossRef]
- Vaughn, A.A.; delBarco-Trillo, J.; Ferkin, M.H. Sperm investment in male meadow voles is affected by the condition of the nearby male conspecifics. Behav. Ecol. 2008, 19, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Ramm, S.A.; Stockley, P. Male house mice do not adjust sperm allocation in response to odours from related or unrelated rivals. Anim. Behav. 2009, 78, 685–690. [Google Scholar] [CrossRef]
- Cummings, M.E.; Ramsey, M.E. Mate choice as social cognition: Predicting female behavioral and neural plasticity as a function of alternative male reproductive tactics. Curr. Opin. Behav. Sci. 2015, 6, 125–131. [Google Scholar] [CrossRef]
- Baum, M.J.; Cherry, J.A. Processing by the main olfactory system of chemosignals that facilitate mammalian reproduction. Horm. Behav. 2015, 68, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Been, L.E.; Petrulis, A. Chemosensory and hormone information are relayed directly between the medial amygdala, posterior bed nucleus of the stria terminalis, and the medial preoptic area in male Syrian hamsters (Mesocricetus auratus). Horm. Behav. 2011, 59, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Been, L.E.; Bauman, J.M.; Petrulis, A.; Chang, Y.H. X-ray kinematics analysis of vaginal scent marking in female Syrian hamsters (Mesocricetus auratus). Physiol. Behav. 2012, 105, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Petty, J.M.A.; Drea, C.M. Female rule in lemurs is ancestral and hormonally mediated. Sci. Rep. 2015, 5, e9631. [Google Scholar] [CrossRef] [PubMed]
- Kavliers, M.; Choleris, E. Social cognition and the neurobiology of rodent mate choice. Integr. Comp. Biol. 2017, 57, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Flatt, T.; Heyland, A. (Eds.) Mechanisms of Life History Evolution: The Genetics and Physiology of Life History Traits and Trade-Offs; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferkin, M.H. Odor Communication and Mate Choice in Rodents. Biology 2018, 7, 13. https://doi.org/10.3390/biology7010013
Ferkin MH. Odor Communication and Mate Choice in Rodents. Biology. 2018; 7(1):13. https://doi.org/10.3390/biology7010013
Chicago/Turabian StyleFerkin, Michael H. 2018. "Odor Communication and Mate Choice in Rodents" Biology 7, no. 1: 13. https://doi.org/10.3390/biology7010013
APA StyleFerkin, M. H. (2018). Odor Communication and Mate Choice in Rodents. Biology, 7(1), 13. https://doi.org/10.3390/biology7010013