
In Vivo Effects of Lipopolysaccharide on Peroxisome Proliferator-Activated Receptor Expression in Juvenile Gilthead Seabream (Sparus Aurata)

 ,
,  , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Procedure
2.2. Blood Metabolite Analyses
2.3. RNA Isolation and cDNA Synthesis
2.4. Quantitative Real-Time PCR (qPCR)
2.5. SDS-PAGE and Immunoblot Analysis
2.6. Statistical Analyses
3. Results
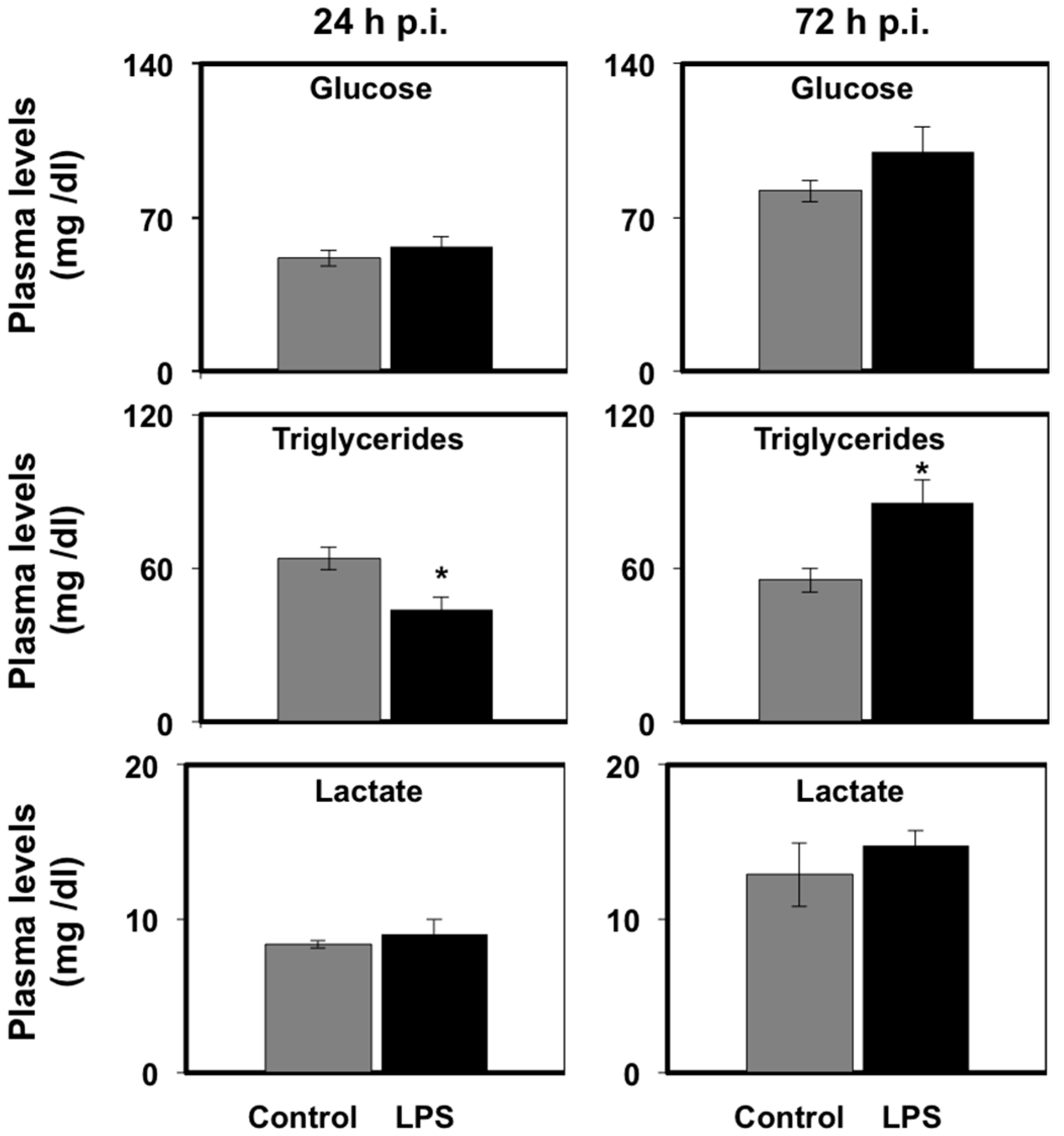
3.1. Effects of LPS Administration in Vivo on Plasma Metabolite Levels in Seabream
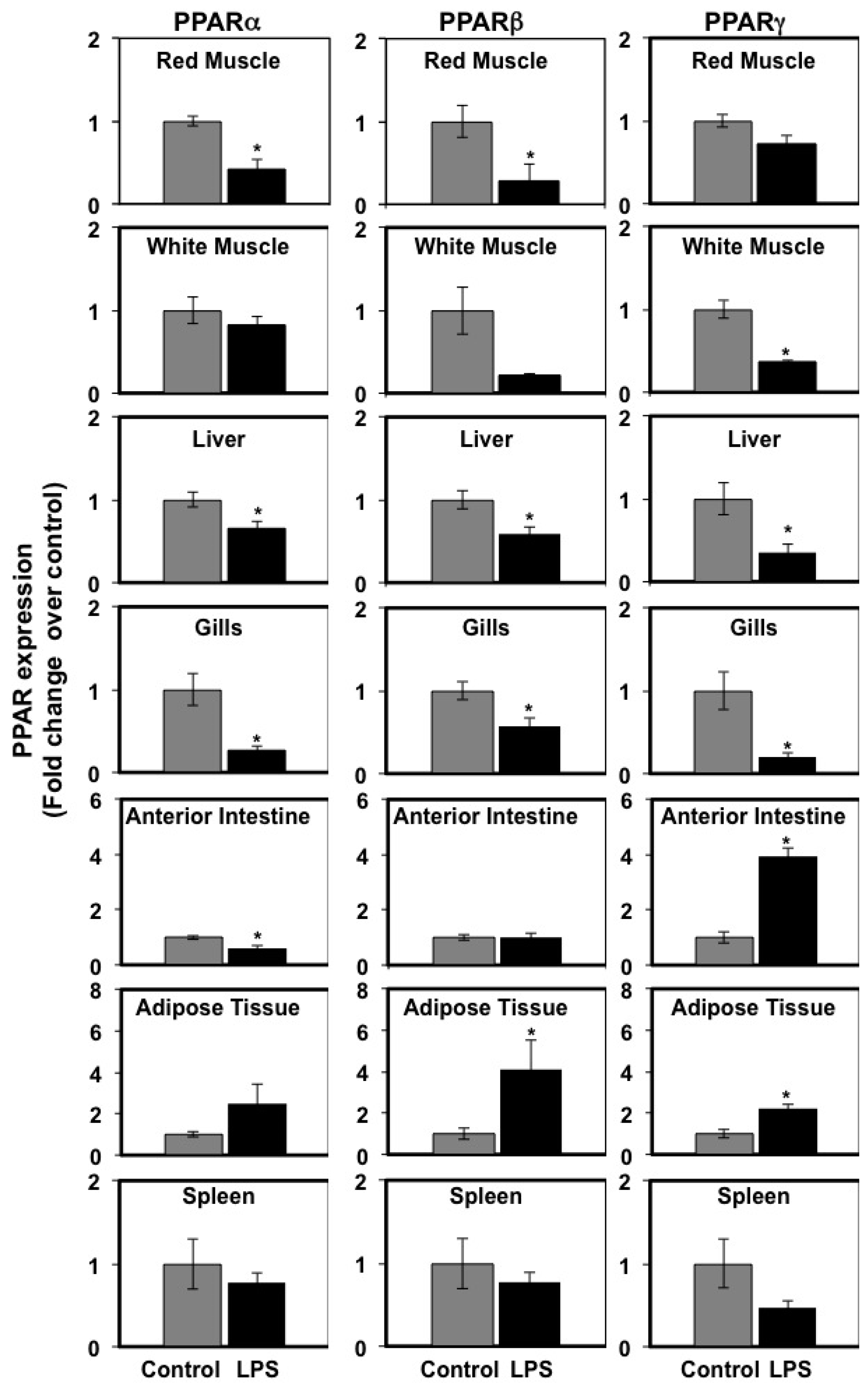
3.2. Effects of LPS Administration In Vivo on PPARα, PPARβ, and PPARγ mRNA Levels in Seabream Tissues
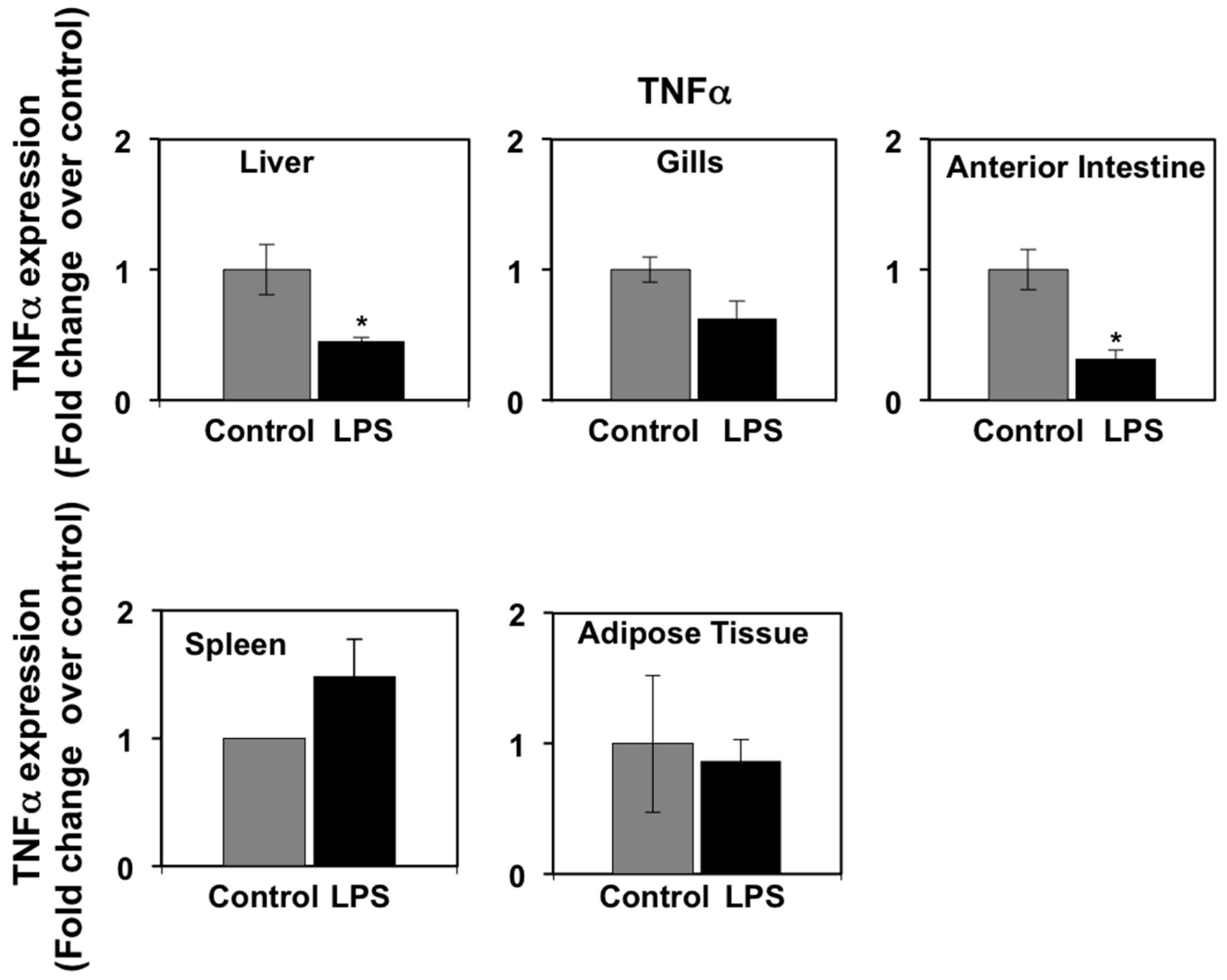
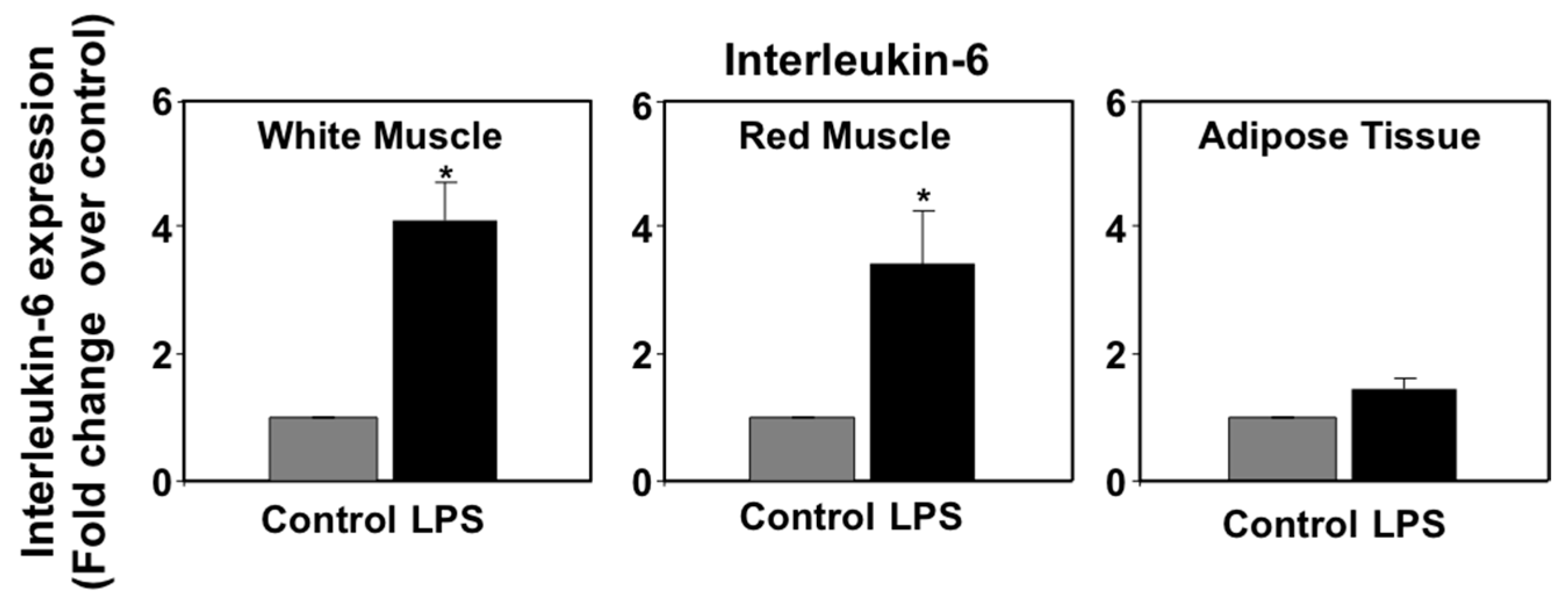
3.3. Effects of LPS Administration In Vivo on Tumor Necrosis Factor α (TNFα) and Interleukin-6 (IL-6) mRNA Levels in Seabream Tissues
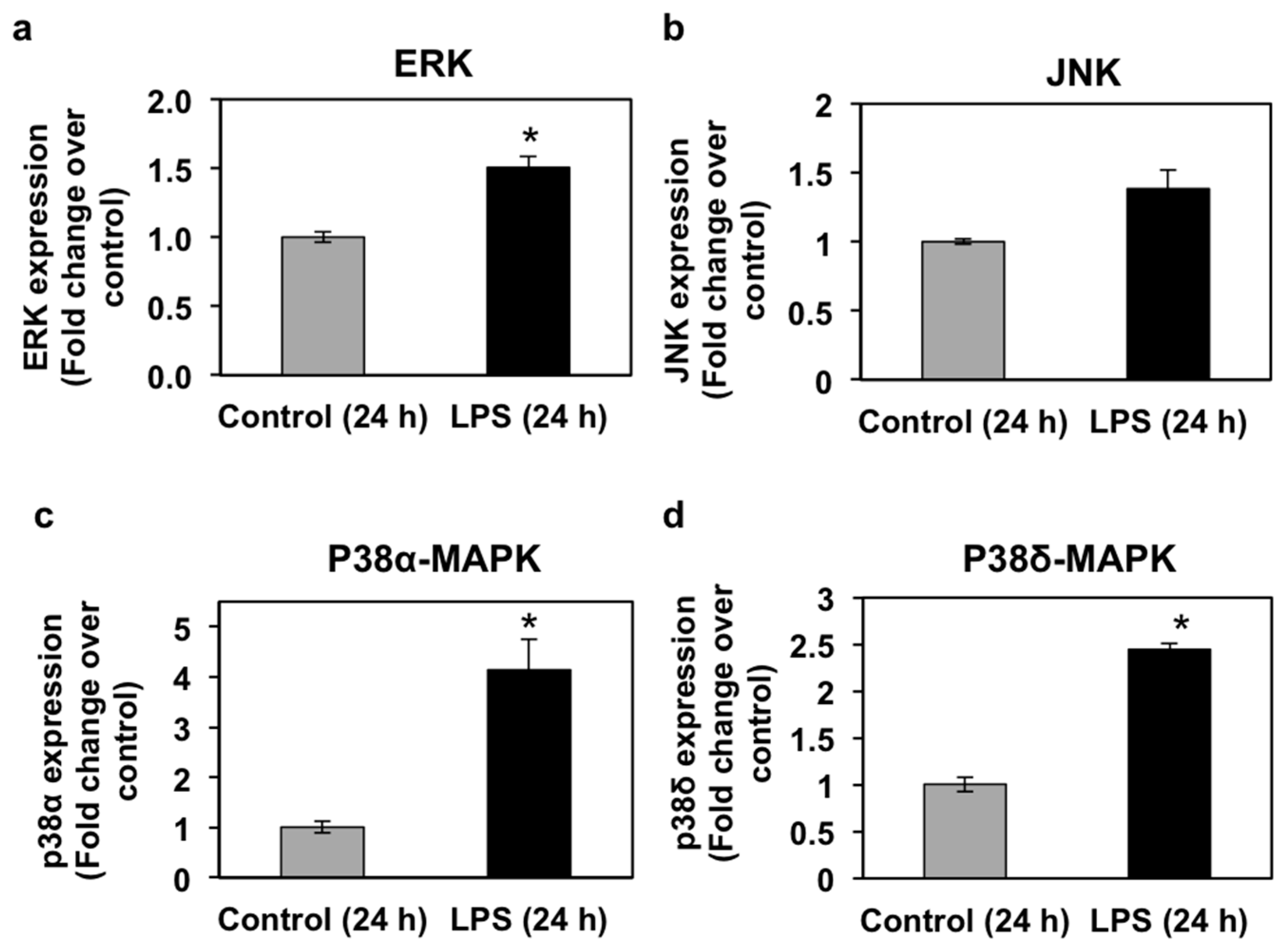
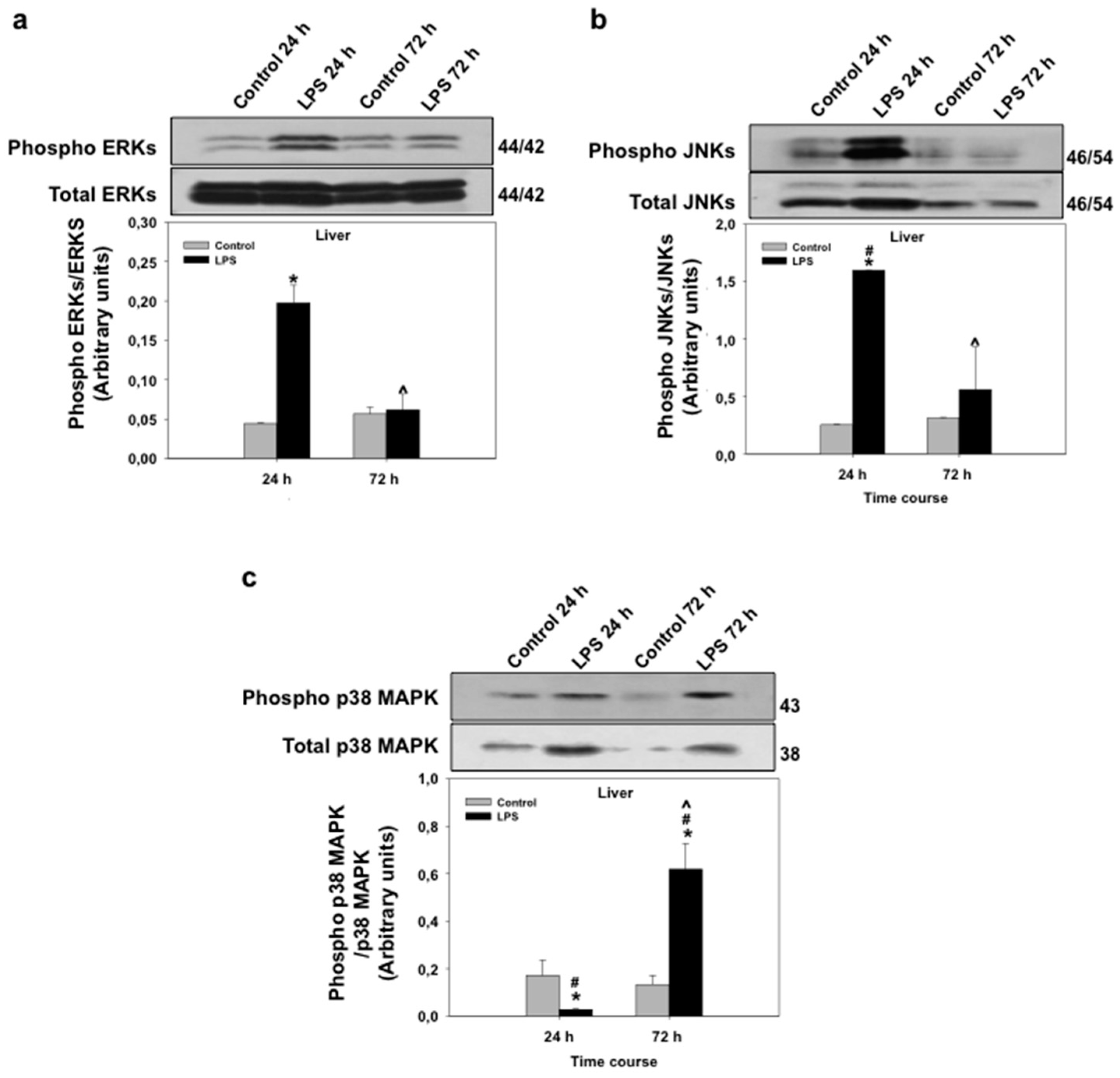
3.4. Effects of LPS Administration In Vivo on mRNA Expression and Phosphorylation Levels of Mitogen-Activated Protein Kinases (MAPKs) in the Seabream Liver
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Swain, P.; Nayak, S.; Nanda, P.; Dash, S. Biological effects of bacterial lipopolysaccharide (endotoxin) in fish: A review. Fish Shellfish Immunol. 2008, 25, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Berczi, I.; Bertók, L.; Bereznai, T. Comparative studies on the toxicity of Escherichia coli lipopolysaccharide endotoxin in various animal species. Can. J. Microbiol. 1966, 12, 1070–1071. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, S.; Balasch, J.C.; Novoa, B.; Ribas, L.; Roher, N.; Krasnov, A.; Figueras, A. Comparative analysis of the acute response of the trout, O. mykiss, head kidney to in vivo challenge with virulent and attenuated infectious hematopoietic necrosis virus and LPS-induced inflammation. BMC Genom. 2008, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Newaj-Fyzul, A.; Austin, B. Probiotics, immunostimulants, plant products and oral vaccines, and their role as feed supplements in the control of bacterial fish diseases. J. Fish Dis. 2015, 38, 937–955. [Google Scholar] [CrossRef] [PubMed]
- Iliev, D.B.; Roach, J.C.; MacKenzie, S.; Planas, J.V.; Goetz, F.W. Endotoxin recognition: In fish or not in fish? FEBS Lett. 2005, 579, 6519–6528. [Google Scholar] [CrossRef] [PubMed]
- Sepulcre, M.P.; Alcaraz-Pérez, F.; López-Muñoz, A.; Roca, F.J.; Meseguer, J.; Cayuela, M.L.; Mulero, V. Evolution of lipopolysaccharide (LPS) recognition and signaling: Fish TLR4 does not recognize LPS and negatively regulates NF-kappaB activation. J. Immunol. 2009, 182, 1836–1845. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, C.; Charette, J.; Catchen, J.; Lage, C.R.; Giasson, G.; Postlethwait, J.H.; Millard, P.J.; Kim, C.H. The gene history of zebrafish TLR4a and TLR4b is predictive of their divergent functions. J. Immunol. 2009, 183, 5896–5908. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, S.; Iliev, D.; Liarte, C.; Koskinen, H.; Planas, J.V.; Goetz, F.W.; Mölsä, H.; Krasnov, A.; Tort, L. Transcriptional analysis of LPS-stimulated activation of trout (Oncorhynchus mykiss) monocyte/macrophage cells in primary culture treated with cortisol. Mol. Immunol. 2006, 43, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Kaitetzidou, E.; Crespo, D.; Vraskou, Y.; Antonopoulou, E.; Planas, J.V. Transcriptomic Response of Skeletal Muscle to Lipopolysaccharide in the Gilthead Seabream (Sparus aurata). Mar. Biotechnol. 2012, 14, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, S.A.; Roher, N.; Boltana, S.; Goetz, F.W. Peptidoglycan, not endotoxin, is the key mediator of cytokine gene expression induced in rainbow trout macrophages by crude LPS. Mol. Immunol. 2010, 47, 1450–1457. [Google Scholar] [CrossRef] [PubMed]
- Roher, N.; Callol, A.; Planas, J.V.; Goetz, F.W.; Mackenzie, S.A. Endotoxin recognition in fish results in inflammatory cytokine secretion not gene expression. Innate Immun. 2011, 17, 16–28. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, S.; Montserrat, N.; Mas, M.; Acerete, L.; Tort, L.; Krasnov, A.; Goetz, F.W.; Planas, J.V. Bacterial lipopolysaccharide induces apoptosis in the trout ovary. Reprod. Biol. Endocrinol. 2006, 4, 46. [Google Scholar] [CrossRef] [PubMed]
- Crespo, D.; Planas, J.V.; Bobe, J. Lipopolysaccharide administration in preovulatory rainbow trout (Oncorhynchus mykiss) reduces egg quality. Aquaculture 2010, 300, 240–242. [Google Scholar] [CrossRef]
- Crespo, D.; Bonnet, E.; Roher, N.; Mackenzie, S.A.; Krasnov, A.; Goetz, F.W.; Bobe, J.; Planas, J.V. Cellular and molecular evidence for a role of tumor necrosis factor alpha in the ovulatory mechanism of trout. Reprod. Biol. Endocrinol. 2010, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Albalat, A.; Liarte, C.; MacKenzie, S.; Tort, L.; Planas, J.V.; Navarro, I. Control of adipose tissue lipid metabolism by tumor necrosis factor-alpha in rainbow trout (Oncorhynchus mykiss). J. Endocrinol. 2005, 184, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Johansen, K.A.; Sealey, W.M.; Overturf, K. The effects of chronic immune stimulation on muscle growth in rainbow trout. Comp. Biochem. Physiol. 2006, 144, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Frost, R.A.; Lang, C.H. Regulation of muscle growth by pathogen-associated molecules. J. Anim. Sci. 2007, 86, E84–E93. [Google Scholar] [CrossRef] [PubMed]
- Escher, P.; Wahli, W. Peroxisome proliferator-activated receptors: Insight into multiple cellular functions. Mutat. Res. 2000, 448, 121–138. [Google Scholar] [CrossRef]
- Vamecq, J.; Latruffe, N. Medical significance of peroxisome proliferator-activated receptors. Lancet 1999, 354, 141–148. [Google Scholar] [CrossRef]
- Ruyter, B.; Andersen, O.; Dehli, A.; Ostlund Farrants, A.K.; Gjøen, T.; Thomassen, M.S. Peroxisome proliferator activated receptors in Atlantic salmon (Salmo salar): Effects on PPAR transcription and acyl-CoA oxidase activity in hepatocytes by peroxisome proliferators and fatty acids. Biochim. Biophys. Acta 1997, 1348, 331–338. [Google Scholar] [CrossRef]
- Oku, H.; Umino, T. Molecular characterization of peroxisome proliferator-activated receptors (PPARs) and their gene expression in the differentiating adipocytes of red sea bream Pagrus major. Comp. Biochem. Physiol. 2008, 151, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Leaver, M.J.; Ezaz, M.T.; Fontagne, S.; Tocher, D.R.; Boukouvala, E.; Krey, G. Multiple peroxisome proliferator-activated receptor subtypes from Atlantic salmon (Salmo salar). J. Mol. Endocrinol. 2007, 38, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Boukouvala, E.; Antonopoulou, E.; Favre-Krey, L.; Diez, A.; Bautista, J.M.; Leaver, M.J.; Tocher, D.R.; Krey, G. Molecular characterization of three peroxisome proliferator-activated receptors from the sea bass (Dicentrarchus labrax). Lipids 2004, 39, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Leaver, M.J.; Boukouvala, E.; Antonopoulou, E.; Diez, A.; Favre-Krey, L.; Ezaz, M.T.; Bautista, J.M.; Tocher, D.R.; Krey, G. Three Peroxisome Proliferator-Activated Receptor Isotypes from Each of Two Species of Marine Fish. Endocrinology 2005, 146, 3150–3162. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.-L.; Chen, H.-Y.; Tseng, M.-C.; Chang, R.-C. Cloning of peroxisome proliferators activated receptors in the cobia (Rachycentron canadum) and their expression at different life-cycle stages under cage aquaculture. Gene 2008, 425, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Huang, Y.; Xie, F.; Huang, X.; Liu, Y.; Wang, W.; Qin, Q. Molecular cloning, characterization and expression analysis of PPAR gamma in the orange-spotted grouper (Epinephelus coioides) after the Vibrio alginolyticus challenge. Fish Shellfish Immunol. 2015, 43, 310–324. [Google Scholar] [CrossRef] [PubMed]
- He, A.-Y.; Liu, C.-Z.; Chen, L.-Q.; Ning, L.-J.; Qin, J.-G.; Li, J.-M.; Zhang, M.-L.; Du, Z.-Y. Molecular characterization, transcriptional activity and nutritional regulation of peroxisome proliferator activated receptor gamma in Nile tilapia (Oreochromis niloticus). Gen. Comp. Endocrinol. 2015, 223, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Gao, J.; Li, D.; Cao, X. Cloning and expressions of peroxisome proliferator activated receptor alpha1 and alpha2 (PPARα1 and PPARα2) in loach (Misgurnus anguillicaudatus) and in response to different dietary fatty acids. Biochem. Biophys. Res. Commun. 2016, 481, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhao, Y.; Li, Y.; Gao, J. Identification and structural characterization of two peroxisome proliferator activated receptors and their transcriptional changes at different developmental stages and after feeding with different fatty acids. Comp. Biochem. Physiol. 2016, 193, 9–16. [Google Scholar] [CrossRef] [PubMed]
- You, C.; Jiang, D.; Zhang, Q.; Xie, D.; Wang, S.; Dong, Y.; Li, Y. Cloning and expression characterization of peroxisome proliferator-activated receptors (PPARs) with their agonists, dietary lipids, and ambient salinity in rabbitfish Siganus canaliculatus. Comp. Biochem. Physiol. 2017, 206, 54–64. [Google Scholar] [CrossRef] [PubMed]
- La Cour Poulsen, L.; Siersbæk, M.; Mandrup, S. PPARs: Fatty acid sensors controlling metabolism. Sem. Cell Dev. Biol. 2012, 23, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Delerive, P.; Fruchart, J.C.; Staels, B. Peroxisome proliferator-activated receptors in inflammation control. J. Endocrinol. 2001, 169, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Venteclef, N.; Jakobsson, T.; Steffensen, K.R.; Treuter, E. Metabolic nuclear receptor signaling and the inflammatory acute phase response. Trends Endocrinol. Metab. 2011, 22, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Wahli, W.; Michalik, L. PPARs at the crossroads of lipid signaling and inflammation. Trends Endocrinol. Metab. 2012, 23, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Feidantsis, K.; Kaitetzidou, E.; Mavrogiannis, N.; Michaelidis, B.; Kotzamanis, Y.; Antonopoulou, E. Effect of taurine-enriched diets on the Hsp expression, MAPK activation and the antioxidant defence of the European sea bass (Dicentrarchus labrax). Aquacul. Nutr. 2014, 20, 431–442. [Google Scholar] [CrossRef]
- Antonopoulou, E.; Kentepozidou, E.; Feidantsis, K.; Roufidou, C.; Despoti, S.; Chatzifotis, S. Starvation and re-feeding affect Hsp expression, MAPK activation and antioxidant enzymes activity of European sea bass (Dicentrarchus labrax). Comp. Biochem. Physiol. 2013, 165, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Villacorta, L.; Garcia-Barrio, M.T.; Chen, Y.E. Transcriptional Regulation of Peroxisome Proliferator–Activated Receptors and Liver X Receptors. Curr. Atheroscler. Rep. 2007, 9, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Ye, J. Regulation of PPARgamma function by TNF-alpha. Biochem. Biophys. Res. Commun. 2008, 374, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Burns, K.; Vandenheuvel, J. Modulation of PPAR activity via phosphorylation. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 2007, 1771, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Gelman, L.; Michalik, L.; Desvergne, B.; Wahli, W. Kinase signaling cascades that modulate peroxisome proliferator-activated receptors. Curr. Opin. Cell Biol. 2005, 17, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Hardardóttir, I.; Grunfeld, C.; Feingold, K.R. Effects of endotoxin on lipid metabolism. Biochem. Soc. Trans. 1995, 23, 1013–1017. [Google Scholar] [CrossRef] [PubMed]
- Hasselgren, P.O. Muscle protein metabolism during sepsis. Biochem. Soc. Trans. 1995, 23, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Saera-Vila, A.; Calduch-Giner, J.A.; Navarro, I.; Pérez-Sánchez, J. Tumour necrosis factor (TNF)α as a regulator of fat tissue mass in the Mediterranean gilthead sea bream (Sparus aurata L.). Comp. Biochem. Physiol. 2007, 146, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S.; Stienstra, R. The role and regulation of the peroxisome proliferator activated receptor alpha in human liver. Biochimie 2017, 136, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Yoon, S.; Tindberg, N.; Järveläinen, H.A.; Lindros, K.O.; Ingelman-Sundberg, M. Hepatic expression of multiple acute phase proteins and down-regulation of nuclear receptors after acute endotoxin exposure. Biochem. Pharmacol. 2004, 67, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Garcia, L.; Sánchez-Gurmaches, J.; Monroy, M.; Gutiérrez, J.; Navarro, I. Regulation of lipid metabolism and peroxisome proliferator-activated receptors in rainbow trout adipose tissue by lipolytic and antilipolytic endocrine factors. Domest. Anim. Endocrinol. 2015, 51, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Garcia, L.; Saera-Vila, A.; Navarro, I.; Calduch-Giner, J.; Perez-Sanchez, J. Targets for TNF-induced lipolysis in gilthead sea bream (Sparus aurata L.) adipocytes isolated from lean and fat juvenile fish. J. Exp. Biol. 2009, 212, 2254–2260. [Google Scholar] [CrossRef] [PubMed]
- Acerete, L.; Balasch, J.C.; Castellana, B.; Redruello, B.; Roher, N.; Canario, A.V.; Planas, J.V.; MacKenzie, S.; Tort, L. Cloning of the glucocorticoid receptor (GR) in gilthead seabream (Sparus aurata). Differential expression of GR and immune genes in gilthead seabream after an immune challenge. Comp. Biochem. Physiol. 2007, 148, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Doñate, C.; Roher, N.; Balasch, J.C.; Ribas, L.; Goetz, F.W.; Planas, J.V.; Tort, L.; MacKenzie, S. CD83 expression in sea bream macrophages is a marker for the LPS-induced inflammatory response. Fish Shellfish Immunol. 2007, 23, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Castellana, B.; Iliev, D.B.; Sepulcre, M.P.; MacKenzie, S. Molecular characterization of interleukin-6 in the gilthead seabream (Sparus aurata). Mol. Immunol. 2008, 45, 3363–3370. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Fischer, C.P. Beneficial health effects of exercise—The role of IL-6 as a myokine. Trends Pharmacol. Sci. 2007, 28, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Päth, G.; Bornstein, S.R.; Gurniak, M.; Chrousos, G.P.; Scherbaum, W.A.; Hauner, H. Human Breast Adipocytes Express Interleukin-6 (IL-6) and Its Receptor System: Increased IL-6 Production by β-Adrenergic Activationj and Effects of IL-6 on Adipocyte Function. J. Clin. Endocr. Metab. 2001, 86, 2281–2288. [Google Scholar] [PubMed]
- Vraskou, Y.; Roher, N.; Diaz, M.; Antonescu, C.N.; MacKenzie, S.A.; Planas, J.V. Direct involvement of tumor necrosis factor-alpha in the regulation of glucose uptake in rainbow trout muscle cells. Amer. J. Physiol. Reg. Integr. Comp. Physiol. 2011, 300, R716–R723. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Mai, K.; Zhang, Y.; Xu, W.; Ai, Q. Tumour necrosis factor-α inhibits hepatic lipid deposition through GSK-3b/b-catenin signaling in juvenile turbot (Scophthalmus maximus L.). Gen. Comp. Endocrinol. 2016, 228, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Mai, K.; Zhang, Y.; Xu, W.; Ai, Q. GSK-3β participates in the regulation of hepatic lipid deposition in large yellow croaker (Larmichthys crocea). Fish Physiol. Biochem. 2016, 42, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Urbatzka, R.; Galante-Oliveira, S.; Rocha, E.; Lobo-da-Cunha, A.; Castro, L.F.C.; Cunha, I. Effects of the PPARα agonist WY-14,643 on plasma lipids, enzymatic activities and mRNA expression of lipid metabolism genes in a marine flatfish, Scophthalmus maximus. Aquat. Toxicol. 2015, 164, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Ning, L.-J.; He, A.-Y.; Li, J.-M.; Lu, D.-L.; Jiao, J.-G.; Li, L.-Y.; Li, D.-L.; Zhang, M.-L.; Chen, L.-Q.; Du, Z.-Y. Mechanisms and metabolic regulation of PPARα activation in Nile tilapia (Oreochromis niloticus). Biochim. Biophys. Acta 2016, 1861, 1036–1048. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.-C.; Wu, K.; Gao, Y.; Zhang, L.-H.; Li, D.-D.; Luo, Z. Magnesium Reduces Hepatic Lipid Accumulation in Yellow Catfish (Pelteobagrus fulvidraco) and Modulates Lipogenesis and Lipolysis via PPARA, JAK-STAT, and AMPK Pathways in Hepatocytes. J. Nutr. 2017, 147, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Mi, H.-F.; Liu, W.-B.; Wu, Y.-Y.; Wang, K.-Z.; Jiang, G.-Z. Molecular characterisation of tumour necrosis factor alpha and its potential connection with lipoprotein lipase and peroxisome proliferator-activated receptors in blunt snout bream (Megalobrama amblycephala). J. Appl. Genet. 2017, 58, 381–391. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′–3′) | Amplicon Size |
|---|---|---|
| saPPARalpha F | GCAGCCTGTGAGTCTTGTGAGTGA | 121 bp |
| saPPARalpha R | CTCCATCAGGTCTCCACACAGC | |
| saPPARbeta F | CGTGTTCGGGATTCGGGACT | 186 bp |
| saPPARbeta R | CACCCTGTCGTGCTGCTCTGTA | |
| saPPARgamma F | CGGAGAGAGAAGCAAGAACAAGAA | 213 bp |
| saPPARgamma R | GAGGAGGAGGAGATGGAGGTGTA | |
| TNFalpha F | GCGACAAACTGGAGACGGAAACC | 221 bp |
| TNFalpha R | GCCTGTTCAGCCACAAGCGTTATC | |
| IL-6 F | GAACTTGTTACAGATCCG | 131 bp |
| IL-6 R | GGCGATGACACCTGTCACTCTCTA | |
| p38α-MAPK F | GGCTCACTCCTACTTCTC | 112 bp |
| p38α-MAPK R | TAATCGTTTCCACTCTTCG | |
| p38δ-MAPK F | CGAAGGTGCGAGGTCATC | 133 bp |
| p38δ-MAPK R | CGGTTTACAGCCAAGTTTCC | |
| JNK F | TCTCCAGCACCCTTATATCAAC | 157 bp |
| JNK R | TGTCCTCTCTTCCCAGTCC | |
| ERK F | GCTCTATGGCAAGGCTGAC | 238 bp |
| ERK R | TGCCTGGAAACGAGCTGTT | |
| 18S F | CAGACAAATCGCTCCACCAACTA | 99 bp |
| 18S R | CTCAACACGGGAAACCTCACC | |
| L13 F | TCTGGAGGACTGTCAGGGGCATGC | 148 bp |
| L13 R | AGACGACAATCTTGAGAGCAG |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antonopoulou, E.; Kaitetzidou, E.; Castellana, B.; Panteli, N.; Kyriakis, D.; Vraskou, Y.; Planas, J.V. In Vivo Effects of Lipopolysaccharide on Peroxisome Proliferator-Activated Receptor Expression in Juvenile Gilthead Seabream (Sparus Aurata). Biology 2017, 6, 36. https://doi.org/10.3390/biology6040036
Antonopoulou E, Kaitetzidou E, Castellana B, Panteli N, Kyriakis D, Vraskou Y, Planas JV. In Vivo Effects of Lipopolysaccharide on Peroxisome Proliferator-Activated Receptor Expression in Juvenile Gilthead Seabream (Sparus Aurata). Biology. 2017; 6(4):36. https://doi.org/10.3390/biology6040036
Chicago/Turabian StyleAntonopoulou, Efthimia, Elisavet Kaitetzidou, Barbara Castellana, Nikolas Panteli, Dimitrios Kyriakis, Yoryia Vraskou, and Josep V. Planas. 2017. "In Vivo Effects of Lipopolysaccharide on Peroxisome Proliferator-Activated Receptor Expression in Juvenile Gilthead Seabream (Sparus Aurata)" Biology 6, no. 4: 36. https://doi.org/10.3390/biology6040036
APA StyleAntonopoulou, E., Kaitetzidou, E., Castellana, B., Panteli, N., Kyriakis, D., Vraskou, Y., & Planas, J. V. (2017). In Vivo Effects of Lipopolysaccharide on Peroxisome Proliferator-Activated Receptor Expression in Juvenile Gilthead Seabream (Sparus Aurata). Biology, 6(4), 36. https://doi.org/10.3390/biology6040036