
Artificial Light at Night Alters Photosynthetic Electron Transport in Two Deciduous Species


Simple Summary
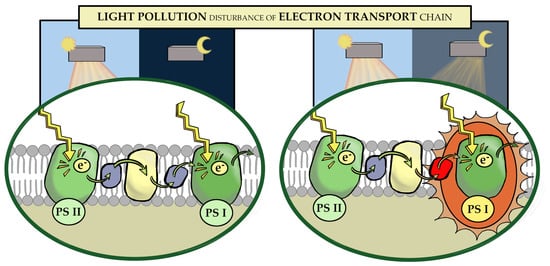
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Light Pollution Treatment
2.3. Measurements of Chlorophyll a Fluorescence
2.4. Data Processing and Statistical Analysis
3. Results and Discussion
3.1. The OJIP Chlorophyll a Fluorescence Transient Curve and Specific Events in the OJIP Curves
3.2. JIP Test Parameters
3.3. Diurnal Light Conditions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galvão, V.C.; Fankhauser, C. Sensing the light environment in plants: Photoreceptors and early signaling steps. Curr. Opin. Neurobiol. 2015, 34, 46–53. [Google Scholar] [CrossRef]
- Paik, I.; Huq, E. Plant photoreceptors: Multi-functional sensory proteins and their signaling networks. Semin. Cell Dev. Biol. 2019, 92, 114–121. [Google Scholar] [CrossRef]
- Neff, M.M.; Fankhauser, C.; Chory, J. Light: An indicator of time and place. Genes Dev. 2000, 14, 257–271. [Google Scholar] [CrossRef]
- De Wit, M.; Galvão, V.C.; Fankhauser, C. Light-mediated hormonal regulation of plant growth and development. Annu. Rev. Plant Biol. 2016, 67, 513–537. [Google Scholar] [CrossRef]
- Wang, X.; Zhou, P.; Huang, R.; Zhang, J.; Ouyang, X. A daylength recognition model of photoperiodic flowering. Front. Plant Sci. 2021, 12, 778515. [Google Scholar] [CrossRef]
- Lau, Z.X.; Grubb, L.E.; Mehta, D.; Uhrig, R.G. The discovery of twilight length sensing in plants and its implications for models of plant photoperiodism. New Phytol. 2025, 249, 123–129. [Google Scholar] [CrossRef]
- Bao, Y.; Liu, X.; Feng, C.H.; Niu, M.X.; Liu, C.; Wang, H.L.; Yin, W.; Xia, X. Light and light signals regulate growth and development in woody plants. Forests 2024, 15, 523. [Google Scholar] [CrossRef]
- Roeber, V.M.; Bajaj, I.; Rohde, M.; Schmülling, T.; Cortleven, A. Light acts as a stressor and influences abiotic and biotic stress responses in plants. Plant Cell Environ. 2021, 44, 645–664. [Google Scholar] [CrossRef]
- Kyba, C.C.; Altıntaş, Y.Ö.; Walker, C.E.; Newhouse, M. Citizen scientists report global rapid reductions in the visibility of stars from 2011 to 2022. Science 2023, 379, 265–268. [Google Scholar] [CrossRef]
- Bará, S.; Falchi, F. Artificial light at night: A global disruptor of the night-time environment. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2023, 378, 20220352. [Google Scholar] [CrossRef]
- Linares Arroyo, H.; Abascal, A.; Degen, T.; Aubé, M.; Espey, B.R.; Gyuk, G.; Hölker, F.; Jechow, A.; Kuffer, M.; Sánchez de Miguel, A.; et al. Monitoring, trends and impacts of light pollution. Nat. Rev. Earth Environ. 2024, 5, 417–430. [Google Scholar] [CrossRef]
- Science Special Issue. Available online: https://www.science.org/toc/science/380/6650 (accessed on 30 December 2025).
- Sustainable Lighting and Light Pollution, Sustainability Special Issue. Available online: https://www.mdpi.com/journal/sustainability/special_issues/sus_light_pollution (accessed on 30 December 2025).
- Ffrench-Constant, R.H.; Somers-Yeates, R.; Bennie, J.; Economou, T.; Hodgson, D.; Spalding, A.; McGregor, P.K. Light pollution is associated with earlier tree budburst across the United Kingdom. Proc. Biol. Sci. 2016, 283, 20160813. [Google Scholar] [CrossRef] [PubMed]
- Heinen, R. A spotlight on the phytobiome: Plant-mediated interactions in an illuminated world. Basic Appl. Ecol. 2021, 57, 146–158. [Google Scholar] [CrossRef]
- Czaja, M.; Kołton, A. How light pollution can affect spring development of urban trees and shrubs. Urban For. Urban Green. 2022, 77, 127753. [Google Scholar] [CrossRef]
- Matzke, E.B. The effect of street lights in delaying leaf-fall in certain trees. Am. J. Bot. 1936, 23, 446–452. [Google Scholar] [CrossRef]
- Škvareninová, J.; Tuhárska, M.; Škvarenina, J.; Babálová, D.; Slobodníková, L.; Slobodník, B.; Středová, H.; Minďaš, J. Effects of light pollution on tree phenology in the urban environment. Morav. Geogr. Rep. 2017, 25, 282–290. [Google Scholar] [CrossRef]
- Meng, L.; Zhou, Y.; Román, M.O.; Stokes, E.C.; Wang, Z.; Asrar, G.R.; Mao, J.; Richardson, A.D.; Gu, L.; Wang, Y. Artificial light at night: An underappreciated effect on phenology of deciduous woody plants. PNAS Nexus 2022, 1, pgac046. [Google Scholar] [CrossRef]
- Friulla, L.; Varone, L. Artificial Light at Night (ALAN) as an Emerging Urban Stressor for Tree Phenology and Physiology: A Review. Urban Sci. 2025, 9, 14. [Google Scholar] [CrossRef]
- Singh, R.K.; Svystun, T.; AlDahmash, B.; Jönsson, A.M.; Bhalerao, R.P. Photoperiod-and temperature-mediated control of phenology in trees—A molecular perspective. New Phytol. 2017, 213, 511–524. [Google Scholar] [CrossRef]
- Wang, L.; Meng, L.; Richardson, A.D.; Hölker, F.; Li, H.; Mao, J.; Longcore, T.; Xia, J.; She, D. Artificial light at night outweighs temperature in lengthening urban growing seasons. Nat. Cities 2025, 2, 506–517. [Google Scholar] [CrossRef]
- Piccolo, E.L.; Torre, S.; Lauria, G.; De Quattro, C.; Sebastiani, F.; Guidi, L.; Remorini, D.; Massai, R.; Landi, M. LED streetlamps alter tree architecture, downregulate the photosynthetic process and alter the sugar metabolism of Populus alba L. Environ. Exp. Bot. 2024, 226, 105861. [Google Scholar] [CrossRef]
- Kolman, F.; Dani, M.; Molnár, P.; Skribanek, A. The effect of three types of street lighting on the morphology and physiology of the common hackberry (Celtis occidentalis L.) leaves. Bot. Lett. 2023, 170, 600–611. [Google Scholar] [CrossRef]
- Kwak, M.J.; Lee, S.H.; Khaine, I.; Je, S.M.; Lee, T.Y.; You, H.N.; Lee, H.K.; Jang, J.H.; KIm, I.; Woo, S.Y. Stomatal movements depend on interactions between external night light cue and internal signals activated by rhythmic starch turnover and abscisic acid (ABA) levels at dawn and dusk. Acta Physiol. Plant 2017, 39, 162. [Google Scholar] [CrossRef]
- Ashraf, M.H.P.J.C.; Harris, P.J. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Muhammad, I.; Shalmani, A.; Ali, M.; Yang, Q.H.; Ahmad, H.; Li, F.B. Mechanisms regulating the dynamics of photosynthesis under abiotic stresses. Front. Plant Sci. 2021, 11, 615942. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Singh Sidhu, G.P.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic response of plants under different abiotic stresses: A review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Chauhan, J.; Prathibha, M.D.; Singh, P.; Choyal, P.; Mishra, U.N.; Saha, D.; Kumar, R.; Anuragi, H.; Pandey, S.; Bose, B.; et al. Plant photosynthesis under abiotic stresses: Damages, adaptive, and signaling mechanisms. Plant Stress 2023, 10, 100296. [Google Scholar] [CrossRef]
- Piccolo, E.L.; Lauria, G.; Guidi, L.; Remorini, D.; Massai, R.; Landi, M. Shedding light on the effects of LED streetlamps on trees in urban areas: Friends or foes? Sci. Total Environ. 2023, 865, 161200. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Li, Z.; Zhang, J.; Hu, D. Influence of night-time light pollution on the photosynthesis and physiological characteristics of the urban plants Euonymus japonicus and Rosa hybrida. Ecol. Processes 2023, 12, 38. [Google Scholar] [CrossRef]
- Stirbet, A.; Lazár, D.; Kromdijk, J.; Govindjee. Chlorophyll a fluorescence induction: Can just a one-second measurement be used to quantify abiotic stress responses? Photosynthetica 2018, 56, 86–104. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Rastogi, A.; Živčák, M.; Brestic, M.; Daszkowska-Golec, A.; Sitko, K.; Alsharafa, K.Y.; Lotfi, R.; Stypiński, P.; Samborska, I.A.; et al. Prompt chlorophyll fluorescence as a tool for crop phenotyping: An example of barley landraces exposed to various abiotic stress factors. Photosynthetica 2018, 56, 953–961. [Google Scholar] [CrossRef]
- Tsimilli-Michael, M. Revisiting JIP-test: An educative review on concepts, assumptions, approximations, definitions and terminology. Photosynthetica 2020, 58, 275–292. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Dangre, D.; Rai, M. Biophysical phenomics reveals functional building blocks of plants systems biology: A case study for the evaluation of the impact of mycorrhization with Piriformospora indica. In Advanced Techniques in Soil Microbiology; Varma, A., Oelmüller, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 319–341. [Google Scholar] [CrossRef]
- Jedmowski, C.; Ashoub, A.; Brüggemann, W. Reactions of Egyptian landraces of Hordeum vulgare and Sorghum bicolor to drought stress, evaluated by the OJIP fluorescence transient analysis. Acta Physiol. Plant 2013, 35, 345–354. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanisms, Regulation and Adaptation, 1st ed.; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor & Francis: London, UK, 2000; Volume 25, pp. 445–483. [Google Scholar]
- Kalaji, H.M.; Loboda, T. Photosystem II of barley seedlings under cadmium and lead stress. Plant Soil Environ. 2007, 53, 511–516. [Google Scholar] [CrossRef]
- Fattahi, M.; Nasrollahpourmoghadam, S. Evaluation of Photosystem II Activity in Two Almond Genotypes under Drought Stress by Exploring the OJIP Test. Evaluation 2024, 37, 901–914, (In Persian with English abstract). [Google Scholar] [CrossRef]
- Chen, X.; Liu, X.; Cong, Y.; Jiang, Y.; Zhang, J.; Yang, Q.; Liu, H. Melatonin alleviates photosynthetic injury in tomato seedlings subjected to salt stress via OJIP chlorophyll fluorescence kinetics. Plants 2025, 14, 824. [Google Scholar] [CrossRef]
- Sabouri, H.; Kazerani, B.; Taliei, F.; Ghasemi, S. Genome-Wide Composite Interval Mapping Reveal Closely Linked Quantitative Genes Related to OJIP Test Parameters under Chilling Stress Condition in Barley. Agriculture 2024, 14, 1272. [Google Scholar] [CrossRef]
- Aucique-Perez, C.; Shahid, I.; Shahid, M. Photosynthetic Damage in Citrus Leaves Under Freezing Stress Revealed by OJIP Chlorophyll Fluorescence Kinetics. Poster Published in The 2nd International Electronic Conference on Horticulturae Session Abiotic and Biotic Stress. 2025; Available online: https://sciforum.net/manuscripts/23048/slides.pdf (accessed on 30 December 2025).
- Zhu, L.; Wen, W.; Thorpe, M.R.; Hocart, C.H.; Song, X. Combining Heat Stress with Pre-Existing Drought Exacerbated the Effects on Chlorophyll Fluorescence Rise Kinetics in Four Contrasting Plant Species. Int. J. Mol. Sci. 2021, 22, 10682. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Kumar, D.; Soni, V. Performance of chlorophyll a fluorescence parameters in Lemna minor under heavy metal stress induced by various concentration of copper. Sci. Rep. 2022, 12, 10620. [Google Scholar] [CrossRef] [PubMed]
- Kalisz, A.; Kornaś, A.; Skoczowski, A.; Oliwa, J.; Jurkow, R.; Gil, J.; Sękara, A.; Caruso, G. Leaf chlorophyll fluorescence and reflectance of oakleaf lettuce exposed to metal and metal (oid) oxide nanoparticles. BMC Plant Biol. 2023, 23, 329. [Google Scholar] [CrossRef]
- Ren, J.; Guo, P.; Zhao, X.; Ma, X.; Ai, X.; Wang, J.; Zou, H.; Yu, H. Differential photosynthetic responses to drought stress in peanut varieties: Insights from transcriptome profiling and JIP-Test analysis. BMC Plant Biol. 2025, 25, 957. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Račková, L.; Paganová, V.; Swoczyna, T.; Rusinowski, S.; Sitko, K. Can chlorophyll-a fluorescence parameters be used as bio-indicators to distinguish between drought and salinity stress in Tilia cordata Mill? Environ. Exp. Bot. 2018, 152, 149–157. [Google Scholar] [CrossRef]
- Kwak, M.J.; Je, S.M.; Cheng, H.C.; Seo, S.M.; Park, J.H.; Baek, S.G.; Khaine, I.; Lee, T.; Jang, J.; Li, Y.; et al. Night Light-Adaptation Strategies for Photosynthetic Apparatus in Yellow-Poplar (Liriodendron tulipifera L.) Exposed to Artificial Night Lighting. Forests 2018, 9, 74. [Google Scholar] [CrossRef]
- Liu, M.S.; Huang, X.H.; Wang, R.J.; Xu, H.Y.; Zhu, F. Inhibition of photosynthesis in Melia azedarach and Ligustrum lucidum induced by manganese toxicity using OJIP chlorophyll a fluorescence transient. Photosynthetica 2021, 59, 148–159. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, Y.; Lu, Y.; Shi, J.; Chen, S.; Strasser, R.J.; Qiang, S.; Hu, Z. Special issue in honour of Prof. Reto J. Strasser Effect of AtLFNR1 deficiency on chlorophyll a fluorescence rise kinetics OJIP of Arabidopsis. Photosynthetica 2020, 58, 391–398. [Google Scholar] [CrossRef]
- Antunović Dunić, J.; Mlinarić, S.; Pavlović, I.; Lepeduš, H.; Salopek-Sondi, B. Comparative Analysis of Primary Photosynthetic Reactions Assessed by OJIP Kinetics in Three Brassica Crops after Drought and Recovery. Appl. Sci. 2023, 13, 3078. [Google Scholar] [CrossRef]
- Pollastrini, M.; Salvatori, E.; Fusaro, L.; Manes, F.; Marzuoli, R.; Gerosa, G.; Brüggemann, W.; Strasser, R.J.; Bussotti, F. Selection of tree species for forests under climate change: Is PSI functioning a better predictor for net photosynthesis and growth than PSII? Tree Physiol. 2020, 40, 1561–1571. [Google Scholar] [CrossRef]
- Dos Santos, V.A.H.F.; Nelson, B.W.; Rodrigues, J.V.F.C.; Garcia, M.N.; Ceron, J.V.B.; Ferreira, M.J. Fluorescence parameters among leaf photosynthesis-related traits are the best proxies for CO2 assimilation in Central Amazon trees. Braz. J. Bot. 2019, 42, 239–247. [Google Scholar] [CrossRef]
- Pollastrini, M.; Nogales, A.G.; Benavides, R.; Bonal, D.; Finer, L.; Fotelli, M.; Gessler, A.; Grossiord, C.; Radoglou, K.; Strasser, R.J.; et al. Tree diversity affects chlorophyll a fluorescence and other leaf traits of tree species in a boreal forest. Tree Physiol. 2017, 37, 199–208. [Google Scholar] [CrossRef]
- Šrajer Gajdošik, M.; Vicić, A.; Gvozdić, V.; Galić, V.; Begović, L.; Mlinarić, S. Effect of prolonged photoperiod on light-dependent photosynthetic reactions in cannabis. Int. J. Mol. Sci. 2022, 23, 9702. [Google Scholar] [CrossRef]
- Desotgiu, R.; Cascio, C.; Pollastrini, M.; Gerosa, G.; Marzuoli, R.; Bussotti, F. Short and long term photosynthetic adjustments in sun and shade leaves of Fagus sylvatica L., investigated by fluorescence transient (FT) analysis. Plant Biosyst. 2012, 146, 206–216. [Google Scholar] [CrossRef]
- Moy, A.; Le, S.; Verhoeven, A. Different strategies for photoprotection during autumn senescence in maple and oak. Physiol. Plant 2015, 155, 205–216. [Google Scholar] [CrossRef]
- Lazár, D. Modelling of light-induced chlorophyll a fluorescence rise (OJIP transient) and changes in 820 nm-transmittance signal of photosynthesis. Photosynthetica 2009, 47, 483–498. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Kunderlikova, K.; Olsovska, K.; Allakhverdiev, S.I. Effect of photosystem I inactivation on chlorophyll a fluorescence induction in wheat leaves: Does activity of photosystem I play any role in OJIP rise? J. Photochem. Photobiol. B 2015, 152, 318–324. [Google Scholar] [CrossRef]
- Mehta, P.; Jajoo, A.; Mathur, S.; Bharti, S. Chlorophyll a fluorescence study revealing effects of high salt stress on Photosystem II in wheat leaves. Plant Physiol. Biochem. 2010, 48, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, U.; Singh, H.; Kumar, D.; Strasser, R.J.; Soni, V. Severe leaf-vein infestation upregulates antioxidant and photosynthetic activities in the lamina of Ficus religiosa. Acta Physiol. Plant 2022, 44, 15. [Google Scholar] [CrossRef]
- Swoczyna, T.; Łata, B.; Stasiak, A.; Stefaniak, J.; Latocha, P. JIP-test in assessing sensitivity to nitrogen deficiency in two cultivars of Actinidia arguta (Siebold et Zucc.) Planch. ex Miq. Photosynthetica 2019, 57, 646–658. [Google Scholar] [CrossRef]
- Le Verge-Campion, T.; Jauffrais, T.; Lefeuvre, L.; Houlbrèque, F. Commercial Arthrospira platensis extract modifies the photophysiology of Cladocopium goreaui, coral endosymbiont microalgae. Phycology 2025, 5, 50. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Carpentier, R.; Allakhverdiev, S.I.; Bosa, K. Fluorescence parameters as early indicators of light stress in barley. J. Photochem. Photobiol. B 2012, 112, 1–6. [Google Scholar] [CrossRef]
- Yan, K.; Chen, P.; Shao, H.; Shao, C.; Zhao, S.; Brestic, M. Dissection of photosynthetic electron transport process in sweet sorghum under heat stress. PLoS ONE 2013, 8, e62100. [Google Scholar] [CrossRef]
- Li, Y.; Fang, A.; Zhang, T.; Zhang, S.; Zhu, W.; Zhou, Y. Exogenous Calcium Improves Photosynthetic Capacity of Pinus sylvestris var. mongolica under Drought. Forests 2022, 13, 2155. [Google Scholar] [CrossRef]
- Chen, L.S.; Cheng, L. The acceptor side of photosystem II is damaged more severely than the donor side of photosystem II in ‘Honeycrisp’apple leaves with zonal chlorosis. Acta Physiol. Plant. 2010, 32, 253–261. [Google Scholar] [CrossRef]
- Jiang, H.X.; Chen, L.S.; Zheng, J.G.; Han, S.; Tang, N.; Smith, B.R. Aluminum-induced effects on Photosystem II photochemistry in Citrus leaves assessed by the chlorophyll a fluorescence transient. Tree Physiol. 2008, 28, 1863–1871. [Google Scholar] [CrossRef]
- Jiang, Y.; Feng, X.; Wang, H. Heat-induced down-regulation of photosystem II protects photosystem I in honeysuckle (Lonicera japonica). J. Plant Res. 2021, 134, 1311–1321. [Google Scholar] [CrossRef]
- Yan, K.; Han, G.; Ren, C.; Zhao, S.; Wu, X.; Bian, T. Fusarium solani infection depressed photosystem performance by inducing foliage wilting in apple seedlings. Front. Plant Sci. 2018, 9, 479. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R. The oxygen-evolving complex: A super catalyst for life on earth, in response to abiotic stresses. Plant Signal Behav. 2020, 15, 1824721. [Google Scholar] [CrossRef]
- Nohales, M.A.; Kay, S.A. Molecular mechanisms at the core of the plant circadian oscillator. Nat. Struct. Mol. Biol. 2016, 23, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Seo, P.J.; Mas, P. STRESSing the role of the plant circadian clock. Trends Plant Sci. 2015, 20, 230–237. [Google Scholar] [CrossRef]
- Srivastava, D.; Shamim, M.; Kumar, M.; Mishra, A.; Maurya, R.; Sharma, D.; Pandey, P.; Singh, K.N. Role of circadian rhythm in plant system: An update from development to stress response. Environ. Exp. Bot. 2019, 162, 256–271. [Google Scholar] [CrossRef]
- Blair, E.J.; Bonnot, T.; Hummel, M.; Hay, E.; Marzolino, J.M.; Quijada, I.A.; Nagel, D.H. Contribution of time of day and the circadian clock to the heat stress responsive transcriptome in Arabidopsis. Sci. Rep. 2019, 9, 4814. [Google Scholar] [CrossRef]
- Dubois, M.; Claeys, H.; Van den Broeck, L.; Inzé, D. Time of day determines Arabidopsis transcriptome and growth dynamics under mild drought. Plant Cell Environ. 2017, 40, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, O.; Waldron, L.; Nahal, H.; Provart, N.J.; Campbell, M.M. Genotype and time of day shape the Populus drought response. Plant J. 2009, 60, 703–715. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Czaja, M.A.; Kołton, A. Artificial Light at Night Alters Photosynthetic Electron Transport in Two Deciduous Species. Biology 2026, 15, 272. https://doi.org/10.3390/biology15030272
Czaja MA, Kołton A. Artificial Light at Night Alters Photosynthetic Electron Transport in Two Deciduous Species. Biology. 2026; 15(3):272. https://doi.org/10.3390/biology15030272
Chicago/Turabian StyleCzaja, Monika A., and Anna Kołton. 2026. "Artificial Light at Night Alters Photosynthetic Electron Transport in Two Deciduous Species" Biology 15, no. 3: 272. https://doi.org/10.3390/biology15030272
APA StyleCzaja, M. A., & Kołton, A. (2026). Artificial Light at Night Alters Photosynthetic Electron Transport in Two Deciduous Species. Biology, 15(3), 272. https://doi.org/10.3390/biology15030272