1. Introduction
Precise spatial and quantitative regulation of transcription factors is essential for robust plant morphogenesis. Among these regulators, class III homeodomain–leucine zipper (
HD-ZIP III) genes constitute a family that has undergone evolutionary duplication and increased functional redundancy. These genes act as key determinants of developmental patterning (reviewed in [
1]).
Genetic studies have revealed a complex regulatory architecture underlying HD-ZIP III function. In
Arabidopsis and other species, single loss-of-function mutants often exhibit mild or no visible phenotypes due to functional redundancy among family members, whereas higher-order mutants display severe developmental defects. These findings demonstrate that
HD-ZIP III genes are required for the establishment of the shoot apical meristem (SAM), maintenance of meristematic activity and organogenesis [
2,
3,
4].
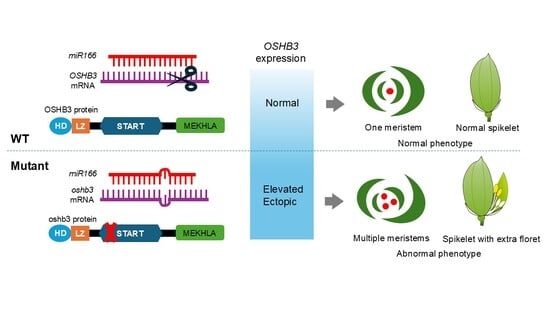
HD-ZIP III transcripts are post-transcriptionally regulated by microRNA (miRNA), specifically miR165/166, which restrict their expression domains and help establish developmental boundaries. Mutations in the miRNA target site generate gain-of-function mutants, which produce dramatic and distinct types of abnormalities compared to loss-of-function mutants. Classical mutants such as
phabulosa and
phavoluta in
Arabidopsis and
rolled in maize carry nucleotide substitutions in the miRNA target sites that impair miRNA-mediated repression and lead to ectopic
HD-ZIP III expression, resulting in severe leaf polarity defects in those mutants [
5,
6]. These findings illustrate that although the
HD-ZIP III gene exhibits a high degree of redundancy in response to decreased expression, even a modest increase in the expression of a single gene can profoundly affect development.
In rice,
HD-ZIP III genes function has been analyzed using single mutants, RNAi knock-down lines and strong overexpression systems. A single recessive mutant of
OSHB3, a member of the
HD-ZIP III family, produced ectopic meristem just after germination [
7], whereas an RNAi knock-down of
OsHox33/
OSHB3 accelerated leaf senescence [
8]. Overexpression of miRNA166-resistant versions of the
OSHB3 gene generated ectopic leaf margins, shoots, and radialized leaves, while
OSHB1 overexpression resulted in a milder phenotype with partially adaxialized leaves [
9]. A single dominant mutant,
lateral floret 1 (
lf1) in which
OSHB1 is ectopically expressed, promoted lateral floret meristem development [
10]. These results indicate that phenotypic changes resulting from endogenous gene upregulation under normal regulatory conditions differ from those resulting from artificial overexpression. Despite this, the developmental consequences of moderate and spatially altered expression of endogenous
OSHB3 genes under native regulatory contexts remain poorly understood in rice.
Here, we report the isolation of a semi-dominant rice mutant with altered meristem initiation carrying a single-nucleotide substitution in the miRNA target site of OSHB3. We observed ectopic expression of OSHB3 in the mutant, leading to dosage-dependent defects in meristem initiation, with homozygous plants exhibiting more severe phenotypes than heterozygotes. Our findings suggest that fine-tuned dosage control of HD-ZIP III genes is critical for maintaining proper meristem patterning in rice.
2. Materials and Methods
2.1. Plant Materials
The mutant line was isolated by screening M2 populations of Taichung 65 (T65) backgrounds treated with N-methyl-N-nitrosourea. Since homozygous mutants are sterile, we used heterozygous plants for maintaining dominant mutations.
For rough mapping, heterozygous mutants were crossed with Kasalath (Ka) cultivars. A total of 44 F2 progenies displaying the wild-type phenotype were used for mapping the mutation.
Plants were sown and grown on Murashige–Skoog medium at 28 °C under continuous light conditions after surface-sterilizing and washing with water. Seeds were soaked overnight in a 200-fold diluted solution of fungicide (Kumiai Chemical Industry Co., Ltd., Tokyo, Japan) at room temperature and washed with water. After 2 days in water at 15 °C, the seeds were transferred to a 28 °C incubator overnight and placed in soil after germination.
2.2. Genotyping
Genotyping was performed to distinguish homozygous and heterozygous individuals of each mutant. DNA was extracted from individual plants by grinding a small amount of plant tissue in liquid nitrogen with a multi-bead shocker (Yasui Instruments Co., Ltd., Osaka, Japan), adding 300 µL of TPS buffer (0.1 M Tris-HCl pH 8.0, 1 M KCl, 0.01 M EDTA) and incubating at 70 °C for 30 min. For precipitation, samples were centrifuged at 2900 rpm for 15 min at 4 °C. A total of 200 µL of isopropanol was added to the supernatant and the mixture was centrifuged at 4 °C, 2900 rpm for 15 min after mixing. Samples were then washed with 300 µL of 70% ethanol solution followed by a centrifugation at 4 °C, 2900 rpm for 15 min. The pellet was dissolved in 50 µL of sterile water.
PCR was performed using CAPS primers and the PCR products were subjected to restriction enzyme analysis. Primer pairs for each mutant allele and enzymes used for genotyping are listed in
Table S1. PCR reactions were performed with KOD neo (TOYOBO Co., Ltd., Osaka, Japan) under the following conditions: initial denaturation: 94 °C for 3 min; (denaturation at 94 °C for 30 s, annealing at 67 °C for 30 s, extension at 68 °C for 90 s) × 6 cycles; (annealing temperature decreased by 0.5 °C per cycle); (denaturation at 94 °C for 30 s, annealing at 62 °C for 30 s, extension at 68 °C for 90 s) × 29 cycles. After the PCR reaction, Mbo I (New England Biolabs, Boston, MA, USA) was used to digest the products and digested samples were electrophoresed to detect polymorphisms.
2.3. Mapping
In the first step, we conducted an experiment using pooled DNA and 48 sets of markers to determine the chromosome on which the gene responsible for the mutant was located. Markers located at 12.3 cM, 47.2 cM, 95 cM, and 104 cM on chromosome 12 (which was identified as carrying the causative gene) were used. The regions near 104 cM emerged as candidate regions; therefore, markers were developed at 96 cM and 102 cM to perform a slightly finer mapping. The number of recombinants at each marker position was as follows: 96cM:14; 102cM:1; and 104cM:0. This suggested that the candidate region was likely to be located toward the telomeric end of the long arm of chromosome 12, relative to the marker at 102 cM. The OSHB3 gene was located within this region.
2.4. Sectioning
Samples from the mutant and wild-type plants were fixed with 4% (w/v) paraformaldehyde (PFA) (Sigma-aldrich, St Louis, MO, USA) and 1% Triton-X (FUJIFILM Co., Tokyo, Japan) in 0.1 M sodium phosphate buffer for 48 h at 4 °C. Samples were then dehydrated in a gradual ethanol series, substituted with t-butanol (FUJIFILM Co., Tokyo, Japan), and embedded in Paraplast Plus (McCormick Scientific, MO, USA). Sectioning was performed at 10 µm thickness using a rotary microtome (Microm, Walldorf, Germany). The sections were stained with hematoxylin (SAKURA Finetek, Tokyo, Japan) and observed under a light microscope (BX-51, Olympus, Tokyo, Japan). For resin sectioning, we followed the same procedure as for preparing paraffin sections up to the 100% ethanol replacement step, then immersed the sample in a mixture of replacement solution (Technovit 7100, Heraeus Kulzer GmbH, Wehrheim, Germany) and ethanol. The concentration of Technovit 7100 was gradually increased every half day until it reached 100%. Subsequently, the resin was poured into a mold and the sample was embedded, covered with parafilm (amcor, Zurich, Switzerland) and placed in the refrigerator overnight to harden. Sections were cut at 5 μm thickness using a microtome and arranged on microscope slides, stained with toluidine blue (Waldeck, GmbH, Munster, Germany), and then observed under a light microscope.
2.5. SEM Observation
Samples were fixed in 4% of PFA at 4 °C overnight. Then, they were dehydrated through an ethanol series and isoamyl acetate (KANTO CHEMICAL Co., Ltd., Tokyo, Japan). Subsequently, samples were dried using a JCPD-5 (JEOL Ltd., Tokyo, Japan) critical point drying system. After ion coating, the samples were observed using a TM3030Plus Miniscope (Hitach high-tech, Tokyo, Japan).
2.6. RNA Extraction and Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
Total RNA was extracted using TRIzol (Invitrogen, Carlsbad, CA, USA), following the manufacturer’s protocol.
A total of 500 µg of RNA was used for first-strand cDNA synthesis using the ReverTra Ace qPCR RT Master Mix with gDNA Remover (TOYOBO Co., Ltd., Osaka, Japan). The cDNA was diluted 20 times and used for real-time PCR. For quantification of the genes, KOD SYBR qPCR Mix (TOYOBO Co., Ltd., Osaka, Japan) was used. The expression level of each sample was normalized to that of an internal control,
UBIQUITIN5 (
UBQ5). The primers for the detection of
OSHB3/ABL2 and
UBQ5 are listed in
Table S1. Amplification conditions were initial denaturation at 98 °C for 2 min, denaturation at 98 °C for 10 s, annealing at 60 °C for 10 s, and extension at 68 °C for 30 s: this cycle was repeated 40 times. Finally, a melting curve analysis was performed. Settings were adjusted to standard settings for each instrument.
2.7. In Situ Hybridization
For studying
ABL2/OSHB3 and
OSH1 expression patterns by in situ hybridization, we followed the protocol noted in [
9]. Samples were fixed using the same procedure described above.
4. Discussion
The present study suggests that a defining characteristic of a gain-of-function mutant with a single-nucleotide substitution in OSHB3 is the repeated formation of ectopic meristems adjacent to the primary meristems throughout the plant’s lifecycle.
During early development, both homozygous and heterozygous mutants exhibited phenotypes such as aberrant phyllotaxy and the formation of ectopic meristems, but these phenotypic changes were more frequent in the homozygous mutants (
Figure 4A–D;
Figure S2;
Table 1). Furthermore, homozygous mutants remained dwarf in stature and perished early, whereas heterozygous mutants survived until the reproductive growth stage. The formation patterns and arrangements of axillary meristems in heterozygous mutants were unique. Ectopic meristems differentiated at multiple stages of axillary bud development; in particular, the earliest ectopic meristems formed adjacent to the prophyll at the onset of prophyll formation, and additional ectopic meristems differentiated within the prophylls themselves. As a result, several prophylls developed in parallel or nested configurations (
Figure 4H). As they entered the reproductive development, the mutants developed additional phenotypic changes, such as the formation of extra flowers, changes in the number of lemma and palea, and a reduction in the number of stamens (
Figure 5;
Table S2). In contrast, in the gain-of-function mutant of
OSHB1,
lf1 [
10,
11], the only abnormal phenotype observed was the formation of extra flowers, and the lemma, palea, lodicules, and stamens showed normal development. In other words, while both
OSHB1 and
OSHB3 shared a similar function in activating extra meristems in the spikelets during reproductive development, there were differences in the function of these genes during flower development.
The phenotypic changes in the homozygous mutant were more pronounced than those in the heterozygous mutant, and this extent of change correlated with the level of
OSHB3 expression and the size of the expression domain. Among the heterozygous mutants exhibiting individual variation in phenotypic severity, those with visible abnormalities exhibited higher
OSHB3 expression in their flowers than those without (
Table S2;
Figure 2G). These results suggest
OSHB3 as an important regulator of meristem initiation and underscore the significance of precise dosage control of
HD-ZIP III genes in rice development. Previous studies using
ACTIN promoter-driven overexpression of miRNA-resistant
OSHB genes showed ectopic shoots and aberrant leaf morphology [
7], establishing essential roles for
HD-ZIP III genes in polarity and meristem maintenance during vegetative growth. However, the contribution of
OSHB3 to subsequent developmental processes remained unclear and is addressed in this study.
The contrasting difference in the effects of loss-of-function and gain-of-function mutations in
HD-ZIP III genes is intriguing. Loss-of-function mutations in individual
HD-ZIP III genes often produce weak or undetectable phenotypes due to functional redundancy among family members [
2,
3,
4]. In contrast, gain-of-function mutations frequently result in severe developmental abnormalities [
5,
6]. Both previous studies [
9] and our present study show that
OSHB3 mutants follow this pattern. These observations are consistent with the idea that HD-ZIP III activity must be maintained within a relatively narrow range during shoot development. The observation that both the recessive and gain-of-function mutants produce bipolar leaves supports the hypothesis.
Several hypotheses can be proposed to explain the mutation responsible for the phenotype observed in this gain-of-function mutant. First, because the mapping resolution is limited, we cannot exclude the possibility that causal mutations reside outside the
OSHB3 locus and indirectly affect
OSHB3 expression or localization, thereby leading to the observed phenotype. However, the strong correlations between genotypes, phenotypes and extent of gene expression make this unlikely. Second, even if the mutation identified in exon 5 of
OSHB3 is causal, its effect may not necessarily involve miRNA-mediated regulation. The amino acid substitution within the START domain could alter protein function, for example, by affecting dimerization or downstream gene regulation [
12], which in turn may influence
OSHB3 expression patterns and contribute to the phenotype. One plausible explanation for the increased and ectopic expression of
OSHB3 involves disruption of miR166-mediated regulation. The mutation we identified is located within the predicted miR166 target site in
OSHB3. Notably, it corresponds to the sixth nucleotide from the 5′ end of miR166, within the seed region that is critical for target recognition. Mismatches in this region are known to strongly affect miRNA–target interactions [
13,
14]. Therefore, this mutation may impair miR166 binding, which in turn could cause changes in
OSHB3 expression and contribute to the mutant phenotype. Furthermore, the morphology of the homozygous mutant was very similar to that of
ACTIN promoter-driven overexpression plants of miRNA-resistant
OSHB3, in that both were dwarf plants with narrowly rolled leaves [
7]. However, ectopic ligules and filamentous leaves that were frequently observed in the overexpression lines were not seen in the homozygous mutants; instead, another phenotype characterized by the absence of leaf auricles was observed (
Figure S3). These minor differences could be attributed to the fact that the overexpression lines were generated using the
ACTIN promoter, but the
OSHB3 expression in the mutants analyzed in this study could be regulated by a miRNA-mediated mechanism.
Taken together, our findings indicate that the dose of OSHB3 expression can alter meristem initiation patterns in rice. Since the initiation patterns of meristems are related to the overall architecture of plants, the insights we have gained regarding these genes may be useful for understanding how the architecture of rice is regulated throughout its life cycle. From an agronomic perspective, increasing the meristems in the axillary buds can increase the number of panicles, and increasing the meristems in the spikelets can increase the number of grains; therefore, OSHB3 may represent a potential target for improving rice yields.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}