
Sunflower HaGLK Enhances Photosynthesis, Grain Yields, and Stress Tolerance of Rice


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Bioinformatic Analysis
2.3. PCR Cloning of HaGLK
2.4. Vector Construction and Rice Genetic Transformation
2.5. Gene Expression Analysis
2.6. Content Measurement of Photosynthetic Pigments
2.7. Photosynthetic Assay
2.8. Ultrastructural Analysis of Chloroplasts
2.9. Statistical Measurement of Rice Agronomic Traits
2.10. Stress Treatments
3. Results
3.1. Bioinformatic Identification of HaGLK Gene
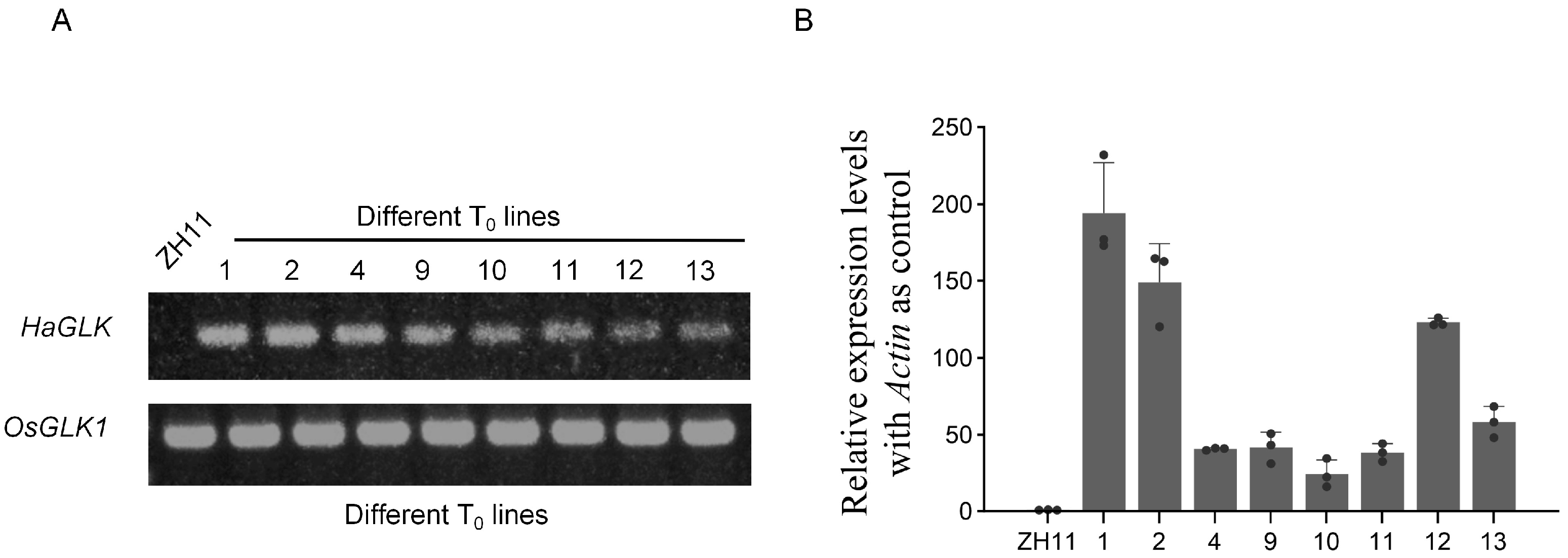
3.2. Generation and Identification of HaGLK Transgenic Rice
3.3. Photosynthetic Capacity Analysis in HaGLK Transgenic Rice
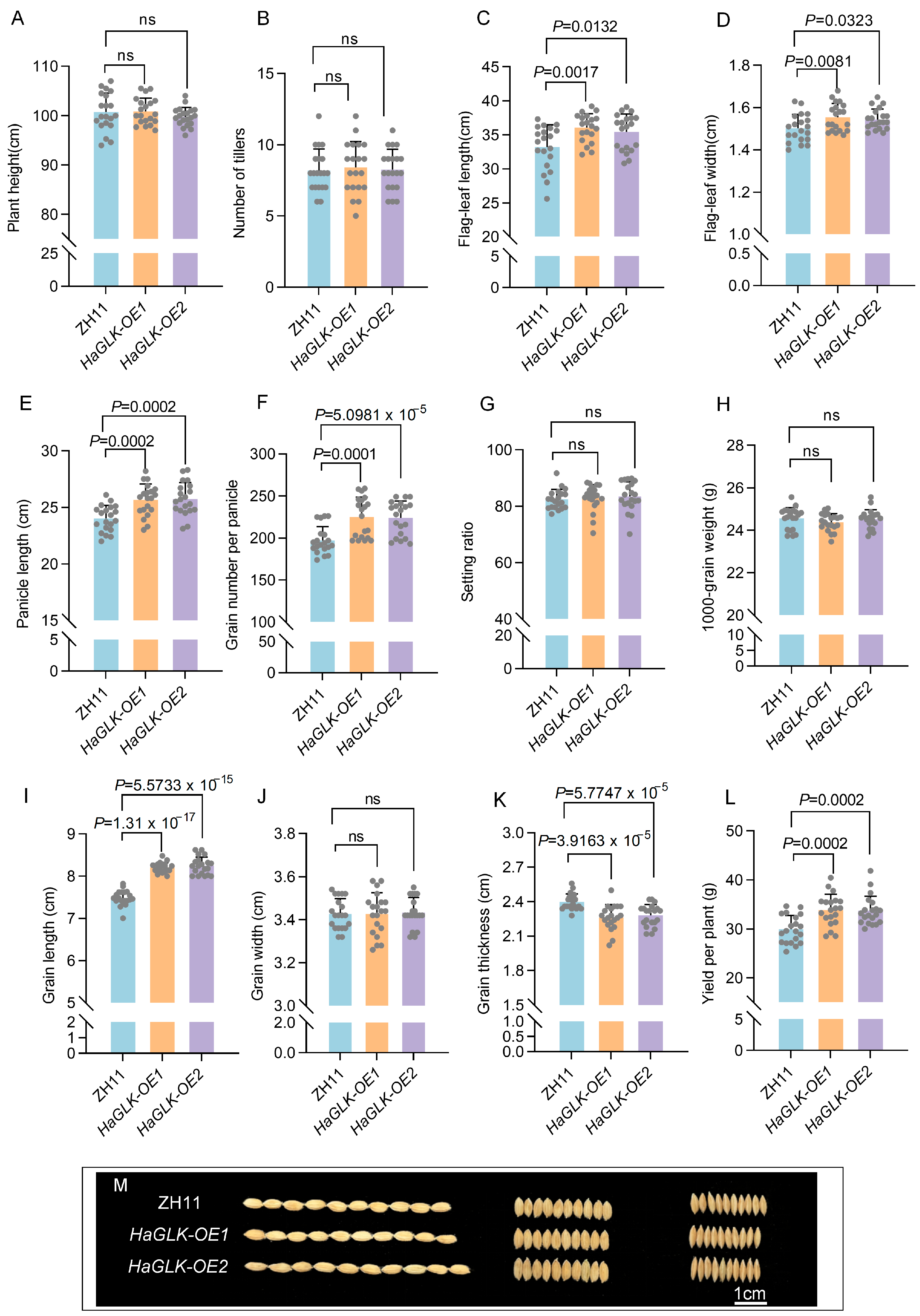
3.4. Agronomic Trait Survey of HaGLK Transgenic Rice
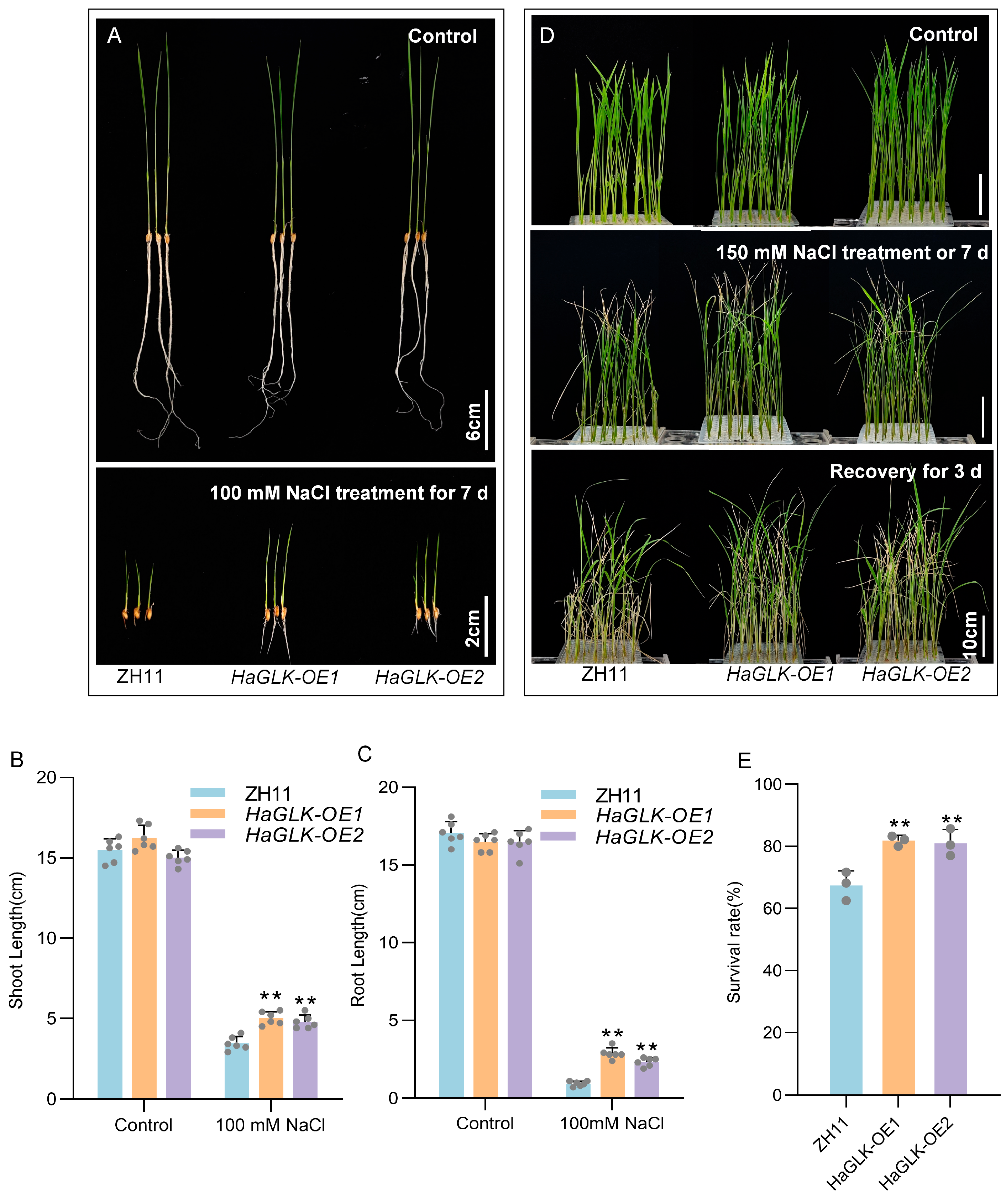
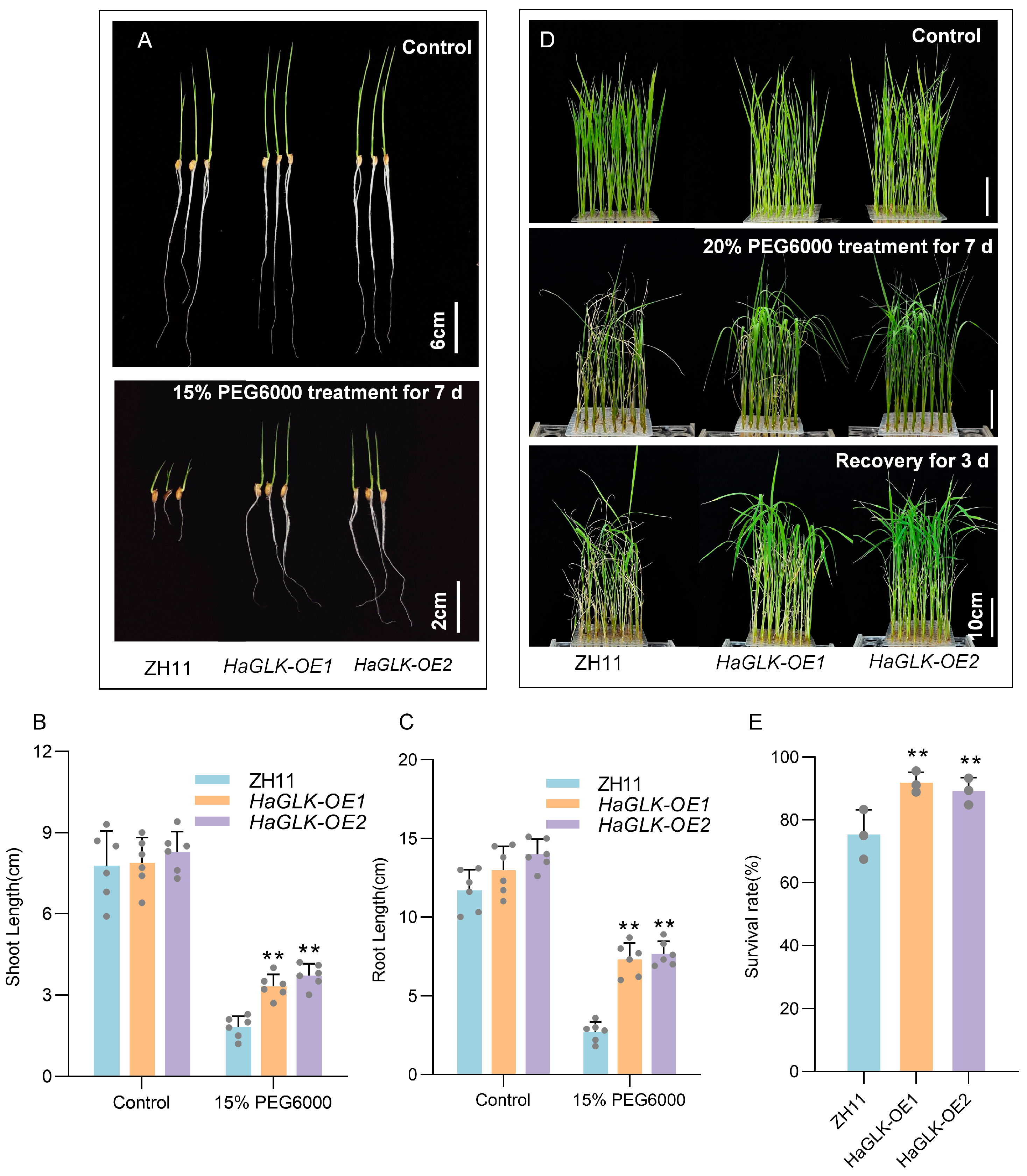
3.5. Stress Tolerance Analysis of HaGLK Transgenic Rice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gotoh, E.; Suetsugu, N.; Yamori, W.; Ishishita, K.; Kiyabu, R.; Fukuda, M.; Higa, T.; Shirouchi, B.; Wada, M. Chloroplast accumulation response enhances leaf photosynthesis and plant biomass production. Plant Physiol. 2018, 178, 1358–1369. [Google Scholar] [CrossRef]
- Shen, S.R.; Yaun, J.J.; Xu, Y.L. Biological function and molecular mechanism of the transcription factor GLKs in plants: A review. Chin. J. Biotechnol. 2022, 38, 2700–2712. [Google Scholar] [CrossRef]
- Wang, H.T.; Xu, F.F. Identification and expression analysis of the GLK gene family in tea plant (Camellia sinensis) and a functional study of CsGLK54 under low-temperature stress. Sci. Rep. 2024, 14, 12465. [Google Scholar] [CrossRef]
- Ali, N.; Chen, H.; Zhang, C.; Khan, S.A.; Gandeka, M.; Xie, D.; Zhuang, W. Ectopic expression of AhGLK1b (GOLDEN2-like transcription factor) in Arabidopsis confers dual resistance to fungal and bacterial pathogens. Genes 2020, 11, 343. [Google Scholar] [CrossRef]
- Botero, K.; Restrepo, S.; Andres, A.P. A genome-scale metabolic model of potato late blight suggests a photosynthesis suppression mechanism. BMC Genom. 2018, 19, 31–44. [Google Scholar] [CrossRef]
- Mai, K.K.K.; Lara, M.V.; Chuong, S.D.X.; Jansonius, N.M.; Weber, A.P.M.; Edwards, G.E.; Franceschi, V.R. Electron tomography analysis of thylakoid assembly and fission in chloroplasts of a single-cell C4 plant, Bienertia sinuspersici. Sci. Rep. 2019, 9, 19640. [Google Scholar] [CrossRef]
- Yeh, S.Y.; Lin, H.H.; Chang, Y.M.; Chang, Y.L.; Chang, C.K.; Huang, Y.C.; Ho, Y.W.; Lin, C.Y.; Zheng, J.Z.; Jane, W.N.; et al. Maize Golden2-like transcription factors boost rice chloroplast development, photosynthesis, and grain yield. Plant Physiol. 2022, 188, 442–459. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Wang, J.; Chen, S.; Liu, H.; Zhou, J.; Yang, W. Maize GOLDEN2-LIKE proteins enhance drought tolerance in rice by promoting stomatal closure. Plant Physiol. 2024, 194, 774–786. [Google Scholar] [CrossRef]
- Liu, J.; Mehari, T.G.; Xu, Y.; Peng, R.; Zhou, L.; Zhang, X.; Chen, Y.; Wang, K. GhGLK1 a Key candidate fene from GARP family enhances cold and drought stress tolerance in cotton. Front. Plant Sci. 2021, 12, 759312. [Google Scholar] [CrossRef]
- Nagatoshi, Y.; Mitsuda, N.; Hayashi, M.; Kubo, A.; Yamaguchi, M.; Ito, T.; Matsuda, M.; Nakamura, T.; Ohme-Takagi, M. GOLDEN 2-LIKE transcription factors for chloroplast development affect ozone tolerance through the regulation of stomatal movement. Proc. Natl. Acad. Sci. USA 2016, 113, 4218–4223. [Google Scholar] [CrossRef]
- Lv, R.; Li, Z.; Li, M.; Zhang, Y.; Yang, X.; Wang, Y.; Zhang, X.; Li, F. Uncoupled expression of nuclear and plastid photosynthesis-associated genes contributes to cell death in a lesion mimic mutant. Plant Cell 2019, 31, 210–230. [Google Scholar] [CrossRef]
- Li, M.; Lee, K.P.; Liu, T.; Ma, J.; Zhang, Y.; Lin, J.; Sun, L.; Deng, X.W. Antagonistic modules regulate photosynthesis-associated nuclear genes via GOLDEN2-LIKE transcription factors. Plant Physiol. 2022, 188, 2308–2324. [Google Scholar] [CrossRef]
- Potter, J.R. Maintenance of high photosynthetic rates during the accumulation of high leaf starch levels in sunflower and soybean. Plant Physiol. 1980, 66, 528–531. [Google Scholar] [CrossRef]
- Li, W.; Zeng, Y.; Yin, F.; Zhang, X.; Liu, J.; Chen, H.; Wang, Y.; Yang, X.; Zhou, M.; Liu, Y. Genome-wide identification and comprehensive analysis of the NAC transcription factor family in sunflower during salt and drought stress. Sci. Rep. 2021, 11, 19865. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, X.; Wu, Y.; Li, Y.; Wang, J.; Liu, H.; Chen, S.; Yang, W.; Zhou, J. Genome-wide identification and evolution of the SAP gene family in sunflower (Helianthus annuus L.) and expression analysis under salt and drought stress. PeerJ 2024, 12, e17808. [Google Scholar] [CrossRef]
- Niu, S.; Tang, M.; Du, C.; Zhang, Y.; Wang, X.; Li, J.; Chen, L. Molecular bases of rice quality regulation and efects of abiotic stress on rice quality. China Rice 2022, 28, 10–19. [Google Scholar] [CrossRef]
- Wang, P.; Fouracres, J.; Kelly, S.; Karki, S.; Lago, C.; Hudson, D.; Powell, B.; Monson, R.K.; Quick, W.P. Evolution of GOLDEN2-LIKE gene function in C3 and C4 plants. Planta 2013, 237, 481–495. [Google Scholar] [CrossRef]
- Chun, Y.; Fang, J.; Savelieva, E.M.; Hu, X.; Zhou, H.; Wang, Y.; Li, Y.; Wang, J.; Zhang, Y.; Wang, Z.; et al. The cytokinin receptor OHK4/OsHK4 regulates inflorescence architecture in rice via an IDEAL PLANT ARCHITECTURE1/WEALTHY FARMER’S PANICLE-mediated positive feedback circuit. Plant Cell 2023, 36, 40–64. [Google Scholar] [CrossRef]
- Toki, S.; Hara, N.; Ono, K.; Saito, H.; Matsuoka, M.; Tanaka, H.; Ohtomo, S.; Mori, M.; Nakamura, C. Early infection of scutellum tissue with Agrobacterium allows high- speed transformation of rice. Plant J. 2006, 47, 969–976. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Porra, R.J. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynth. Res. 2002, 73, 149–156. [Google Scholar] [CrossRef]
- Ned, F. Plant Microtechnique: Some principles and new methods. Am. J. Bot. 1968, 55, 123–142. [Google Scholar] [CrossRef]
- Dubos, C.; Strakraba, T.; Schmidt-Eddy, B. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Park, B.; Burke, J.M. Phylogeography and the evolutionary history of sunflower (Helianthus annuus L.): Wild diversity and the dynamics of domestication. Genes 2020, 11, 266. [Google Scholar] [CrossRef]
- Martín, G.; Leivar, P.; Ludevid, D.; Tepperman, J.M.; Quail, P.H.; Monte, E. Phytochrome and retrograde signalling pathways converge to antagonistically regulate a light-induced transcriptional network. Nat. Commun. 2016, 7, 11431. [Google Scholar] [CrossRef]
- Nguyen, C.V.; Vrebalov, J.T.; Gapper, N.E.; Zheng, Y.; Zhong, S.; Fei, Z. Tomato GOLDEN2-LIKE transcription factors reveal molecular gradients that function during fruit development and ripening. Plant Cell 2014, 26, 585–601. [Google Scholar] [CrossRef]
- Kobayashi, K.; Baba, S.; Obayashi, T.; Sato, M.; Toyooka, K.; Keränen, M. Regulation of root greening by light and auxin/cytokinin signaling in Arabidopsis. Plant Cell 2012, 24, 1081–1095. [Google Scholar] [CrossRef]
- Hubbart, S.; Smillie, I.R.A.; Heatley, M.; Swarup, R.; Foo, C.C.; Zhao, L.; Murchie, E.H. Enhanced thylakoid photoprotection can increase yield and canopy radiation use efficiency in rice. Commun. Biol. 2018, 1, 22. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Kim, E.Y.; Han, S.H.; Piao, W.; An, G.; Todaka, D.; Yamaguchi-Shinozaki, K.; Peak, N.C. Rice phytochrome-interacting factor-like1 (OsPIL1) is involved in the promotion of chlorophyll biosynthesis through feed-forward regulatory loops. J. Exp. Bot. 2017, 68, 4103–4114. [Google Scholar] [CrossRef]
- Yang, Z.; Bai, T.; Zhiguo, E.; Niu, B.; Chen, C. OsNF-YB7 inactivates OsGLK1 to inhibit chlorophyll biosynthesis in rice embryo. eLife 2024, 13, RP96553. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, L.; Kuang, C.; Ma, X.; Zhang, Y.; Wang, J.; Zhou, X.; Liu, S.; Wang, P.; Huang, J. Important photosynthetic contribution of silique wall to seed yield-related traits in Arabidopsis thaliana. Photosynth. Res. 2018, 137, 493–501. [Google Scholar] [CrossRef]
- Zhang, Q.; Mao, Y.; Zhao, Z.; Li, X.; Yang, J.; Wang, H.; Liu, X.; Zhang, Y.; Li, J.; Liu, F. A Golden2-like transcription factor, BnGLK1a, improves chloroplast development, photosynthesis, and seed weight in rapeseed. J. Integr. Agric. 2024, 23, 1481–1493. [Google Scholar] [CrossRef]
- Alam, I.; Jin, Y.; Zhang, Q.; Yang, Y.; Liu, J.; Liu, Y.; Luo, H.; Zheng, H.; Wang, X.; Yu, F. Identification of GOLDEN2-like transcription factor genes in soybeans and their role in regulating plant development and metal ion stresses. Front. Plant Sci. 2022, 13, 1052659. [Google Scholar] [CrossRef]
- Li, X.; Wang, P.; Li, J.; Gao, Y.; Liu, M.; Li, Y.; Li, C.; Xia, X.; Xiao, L.; Zhang, J. Maize GOLDEN2-LIKE genes enhance biomass and grain yields in rice by improving photosynthesis and reducing photoinhibition. Commun. Biol. 2020, 3, 151. [Google Scholar] [CrossRef]
- Rucker, K.S.; Kvien, C.K.; Holbrook, C.C.; Hook, J.E.; Bridges, D.C. Identification of peanut genotypes with improved drought avoidance traits 1. Peanut Sci. 1995, 22, 14–18. [Google Scholar] [CrossRef]
- Luo, X.; Sun, M.; Xu, R.; Jia, B.; Zhang, H.; Li, Y.; Yang, J.; Jia, L.; Zhang, Z. ABA signaling is negatively regulated by GbWRKY1 through JAZ1 and ABI1 to affect salt and drought tolerance. Plant Cell Rep. 2020, 39, 181–194. [Google Scholar] [CrossRef]
- Wu, R.; Guo, L.; Guo, Y.; Wang, H.; Liu, X.; Zhang, J.; Li, Y.; Chen, S. The G2-Like gene family in Populus trichocarpa: Identification, evolution and expression profiles. BMC Genom. Data 2023, 24, 37. [Google Scholar] [CrossRef]
- Liu, X.; Li, Y.; Zhong, S.; Zhang, H.; Wang, X.; Zhou, B.; Zhang, Q.; Wang, J. AhGLK1 affects chlorophyll biosynthesis and photosynthesis in peanut leaves during recovery from drought. Sci. Rep. 2018, 8, 2250. [Google Scholar] [CrossRef]
- Ahmad, R.; Liu, Y.; Wang, T.J.; Meng, X.; Zhang, H.; Pham, B.; Kim, S.H.; Yun, D.J. GOLDEN2-LIKE transcription factors regulate WRKY40 expression in response to abscisic acid. Plant Physiol. 2019, 179, 1844–1860. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| HaGLK | CACTCACCATCACCACCACCA | GGATACCAGGGACCGCTACAT |
| Lhca1 | GAGTTCGTCGCCATCGCCTTCG | CGCCCGTTCTTGATCTCCTTGAGC |
| Lhca3 | GTTCGCATCCAAGCAGTCCCT | CCGTTGAACACCTCGCCGTAG |
| Lhcb2 | TGTTCTCCATGTTCGGCTTCTTCG | GCGTATGCCCAGGCGTTGTT |
| PsaG | AGCGGGAGAACGTGGCGAAGCA | GCAGGTGGCGAGGATGTAGTAGGC |
| PsbQ | GAATTGAAGGGCTGCATTTGT | CCGAAGATCCCGAAGAAGGT |
| Actin | TGGCATCTCTCAGCACATTCC | TGCACAATGGATGGGTCAGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, J.; Zheng, M.; He, J.; Lou, Y.; Ge, Q.; Ma, B.; Chen, X. Sunflower HaGLK Enhances Photosynthesis, Grain Yields, and Stress Tolerance of Rice. Biology 2025, 14, 946. https://doi.org/10.3390/biology14080946
Luo J, Zheng M, He J, Lou Y, Ge Q, Ma B, Chen X. Sunflower HaGLK Enhances Photosynthesis, Grain Yields, and Stress Tolerance of Rice. Biology. 2025; 14(8):946. https://doi.org/10.3390/biology14080946
Chicago/Turabian StyleLuo, Jie, Mengyi Zheng, Jiacheng He, Yangyang Lou, Qianwen Ge, Bojun Ma, and Xifeng Chen. 2025. "Sunflower HaGLK Enhances Photosynthesis, Grain Yields, and Stress Tolerance of Rice" Biology 14, no. 8: 946. https://doi.org/10.3390/biology14080946
APA StyleLuo, J., Zheng, M., He, J., Lou, Y., Ge, Q., Ma, B., & Chen, X. (2025). Sunflower HaGLK Enhances Photosynthesis, Grain Yields, and Stress Tolerance of Rice. Biology, 14(8), 946. https://doi.org/10.3390/biology14080946