
Effects of Dietary Tussah Immunoreactive Pupa Powder on Growth, Gonad Quality, Antioxidant Capacity, and Gut Microbiota of the Sea Urchin Strongylocentrotus intermedius

 ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sea Urchins
2.2. Experimental Design
2.3. Experimental Feed Preparation
2.4. Sample Collection
2.5. Growth Indicators
2.6. Antioxidant Capacity
2.7. Gonad Color Determination
2.8. Gonad Texture Properties
2.9. Routine Nutritional Composition
2.10. Sequencing of Gut Microbiota
2.11. Data Analysis
3. Results
3.1. Sea Urchins and Gonads Growth
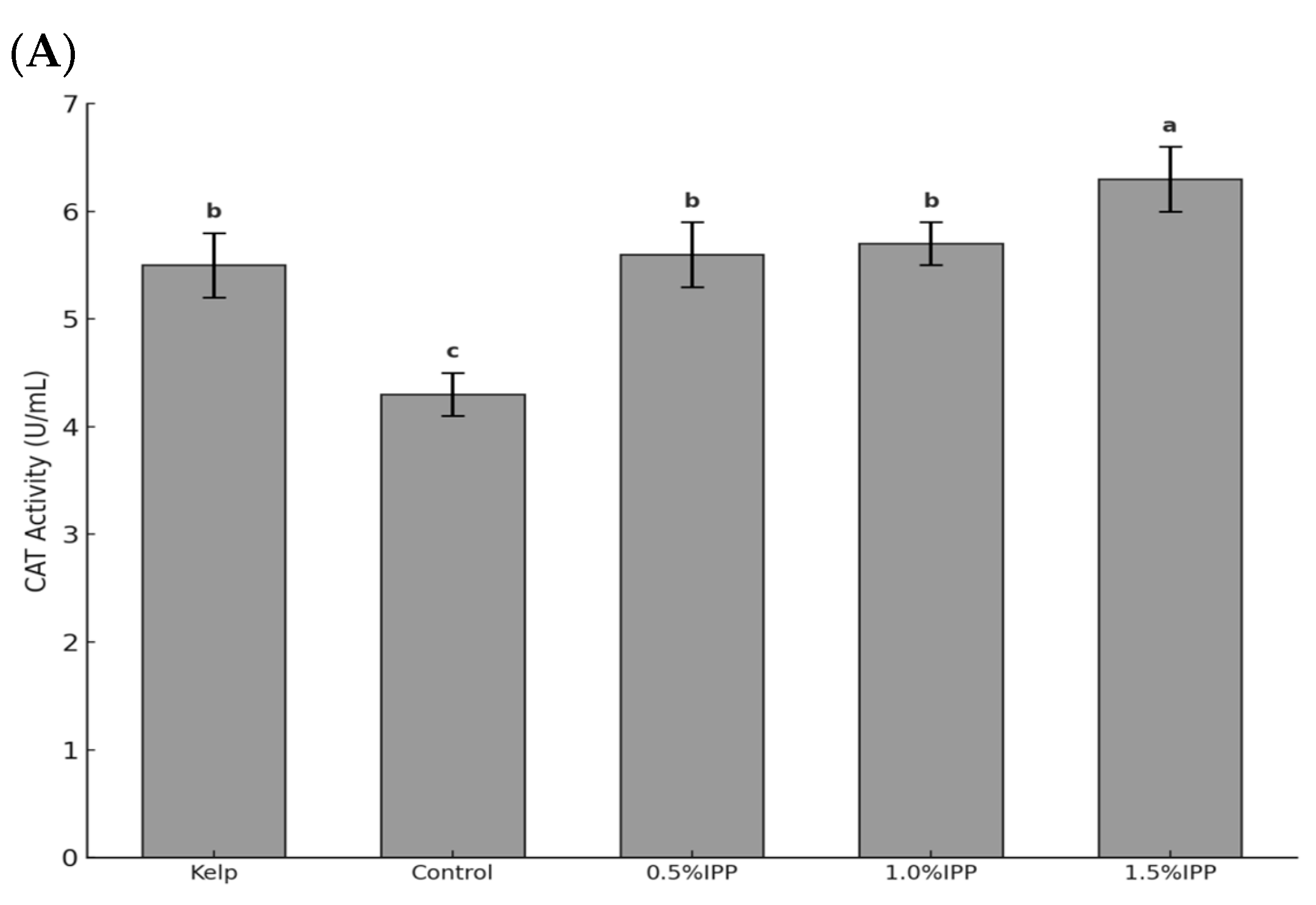
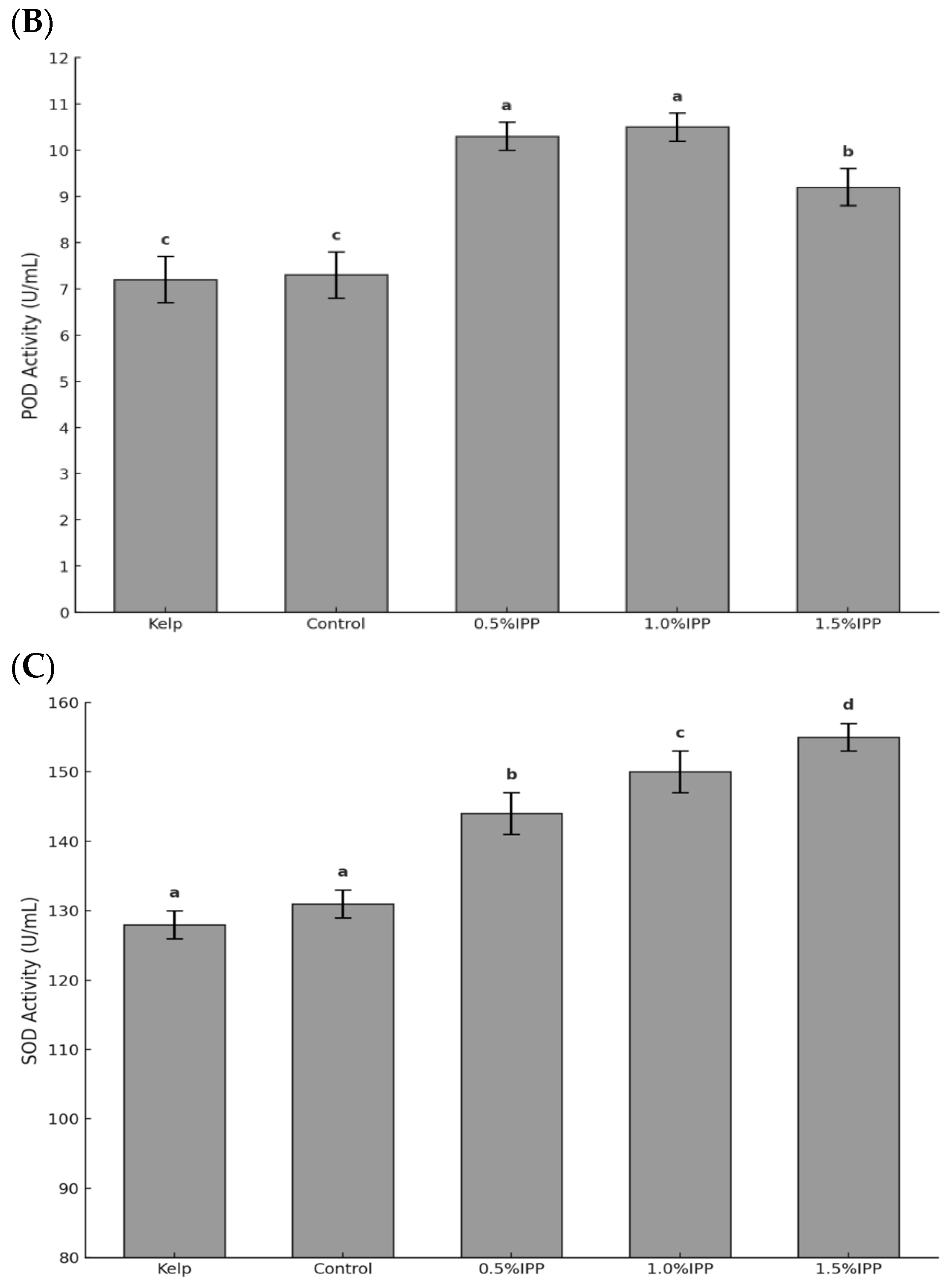
3.2. Antioxidant Enzyme Activity
3.3. Gonad Color
3.4. Gonad Texture
3.5. Routine Nutritional Composition of Gonads
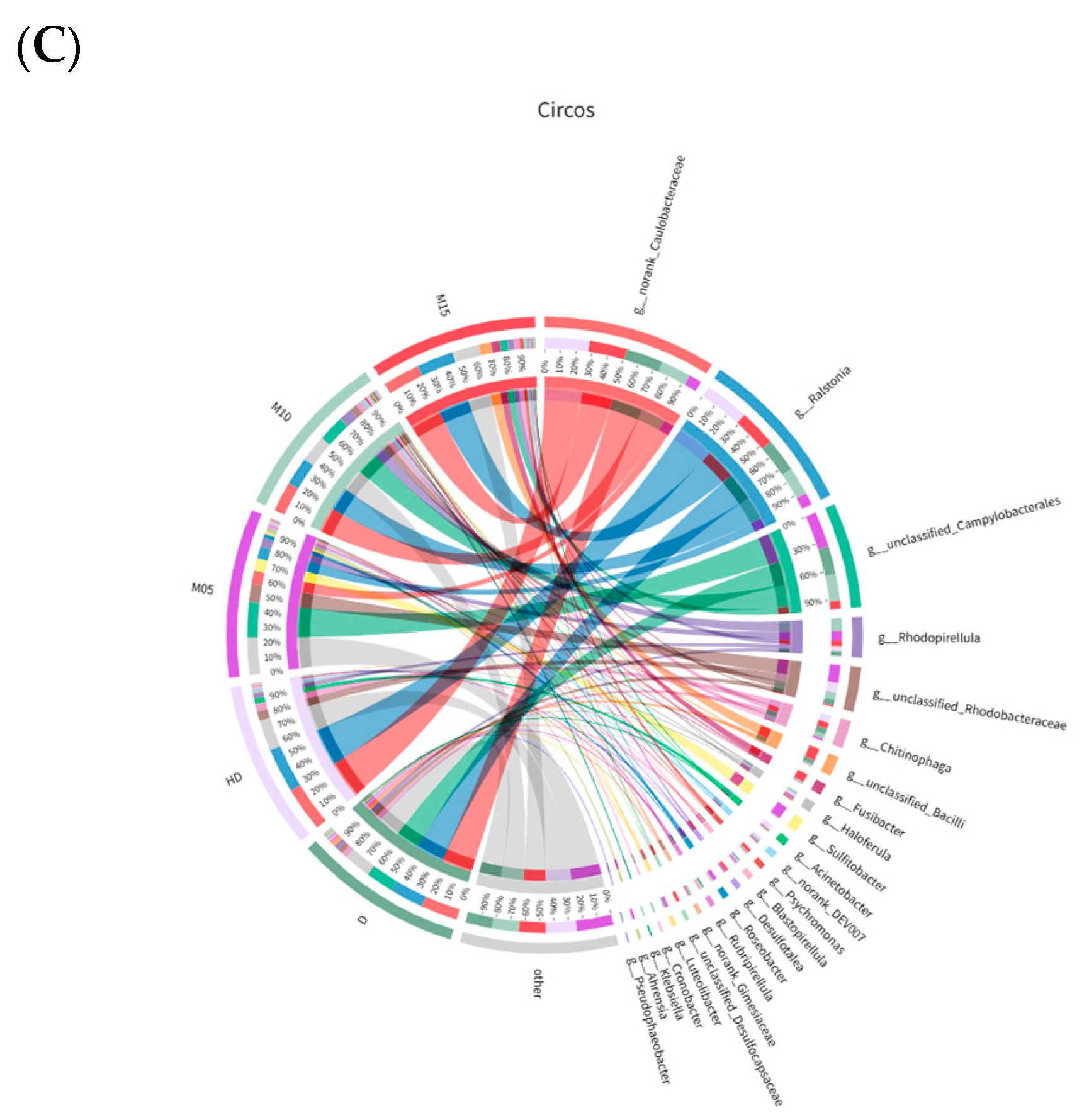
3.6. Gut Microbiota
4. Discussion
4.1. Effects of Adding IPP to the Feed on Growth of Sea Urchins
4.2. Effects of Adding IPP to the Feed on Gonad Traits of Sea Urchins
4.2.1. Effects of Adding IPP to the Feed on Gonad Growth of Sea Urchins
4.2.2. Effects of Adding IPP to the Feed on Gonad Color of Sea Urchins
4.2.3. Effects of Adding IPP to the Feed on Gonad Texture of Sea Urchins
4.3. Effects of Adding IPP to the Feed on Antioxidant Capacity of Sea Urchins
4.4. Effects of Adding IPP to the Feed on Gut Microbiota of Sea Urchins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tavazzi, L.; Maggioni, A.P.; Marchioli, R.; Barlera, S.; Franzosi, M.G.; Latini, R.; Lucci, D.; Nicolosi, G.L.; Porcu, M.; Tognoni, G. Effect of n-3 polyunsaturated fatty acids in patients with chronic heart failure (the GISSI-HF trial): A randomised, double-blind, placebo-controlled trial. Lancet 2008, 372, 1223–1230. [Google Scholar] [PubMed]
- Rubilar, T.; Epherra, L.; Deias-Spreng, J.; De Vivar, M.E.D.; Avaro, M.; Lawrence, A.L.; Lawrence, J.M. Ingestion, absorption and assimilation efficiencies, and production in the sea urchin Arbacia dufresnii fed a formulated feed. J. Shellfish Res. 2016, 35, 1083–1093. [Google Scholar] [CrossRef]
- Ding, J.; Chang, Y. Research progress in conservation and utilization of economic echinoderm: A review. J. Dalian Ocean Univ. 2020, 35, 645–656. (In Chinese) [Google Scholar]
- Knip, D.M.; Scheibling, R.E. Invertebrate fauna associated with kelp enhances reproductive output of the green sea urchin Strongylocentrotus droebachiensis. J. Exp. Mar. Biol. Ecol. 2007, 351, 150–159. [Google Scholar] [CrossRef]
- Li, M.; Gou, D.; Gong, P.; Di, W.; Wang, L.; Ding, J.; Chang, Y.; Zuo, R. An Investigation on the effects of dietary vitamin e on juvenile sea urchin (Strongylocentrotus intermedius): Growth, intestinal microbiota, immune response, and related gene expression. Biology 2023, 12, 1523. [Google Scholar] [CrossRef]
- Lv, D.; Zhang, F.; Ding, J.; Chang, Y.; Zuo, R. Effects of dietary n-3 LC-PUFA on the growth performance, gonad development, fatty acid profile, transcription of related genes and intestinal microflora in adult sea urchin (Strongylocentrotus intermedius). Aquac. Res. 2021, 52, 1431–1441. [Google Scholar] [CrossRef]
- Cook, E.; Kelly, M. Enhanced production of the sea urchin Paracentrotus lividus in integrated open-water cultivation with Atlantic salmon Salmo salar. Aquaculture 2007, 273, 573–585. [Google Scholar] [CrossRef]
- Raposo, A.I.; Ferreira, S.M.; Ramos, R.; Santos, P.M.; Anjos, C.; Baptista, T.; Tecelão, C.; Costa, J.L.; Pombo, A. Effect of three diets on the gametogenic development and fatty acid profile of Paracentrotus lividus (Lamarck, 1816) gonads. Aquac. Res. 2019, 50, 2023–2038. [Google Scholar] [CrossRef]
- Zuo, R.; Li, M.; Ding, J.; Chang, Y. Higher dietary arachidonic acid levels improved the growth performance, gonad development, nutritional value, and antioxidant enzyme activities of adult sea urchin (Strongylocentrotus intermedius). J. Ocean Univ. China 2018, 17, 932–940. (In Chinese) [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Distribution patterns and biological aspects of Strongylocentrotus droebachiensis (Echinoidea: Echinoida) in Russian waters of the Barents Sea: Implications for commercial exploration. Rev. Fish Biol. Fish. 2024, 34, 1215–1229. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Aquaculture of green sea urchin in the Barents Sea: A brief review of Russian studies. Rev. Aquac. 2020, 12, 2080–2090. [Google Scholar] [CrossRef]
- Asia, F.B.; Villamor, J.G.C.; Faylogna, J.C. The effect of prepared diet on the somatic and gonad growth performance of the sea urchin Tripneustes gratilla (LINNAEUS, 1758). E-Int. Sci. Res. J. 2012, 4, 214–228. [Google Scholar]
- Li, M.; Tang, L.; Heqiu, Y.; Lv, D.; Ding, J.; Chang, Y.; Zuo, R. Effects of Oxidized Fish Oil on the Growth, Immune and Antioxidant Capacity, Inflammation-Related Gene Expression, and Intestinal Microbiota Composition of Juvenile Sea Urchin (Strongylocentrotus intermedius). Aquac. Nutr. 2022, 2022, 2340308. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Y.; Guo, Z.; Zhang, R.; Guo, J.; Wang, F.; Han, Y.; Zhao, X.; Ren, T. Effects of a dietary probiotic on growth, gonadal development and quality, antioxidant capacity, intestinal health, and non-specific immunity of a sea urchin (Strongylocentrotus intermedius). Anim. Feed Sci. Technol. 2025, 319, 116184. [Google Scholar] [CrossRef]
- Ma, S.; Sun, Y.; Wang, F.; Mi, R.; Wen, Z.; Li, X.; Meng, N.; Li, Y.; Du, X.; Li, S. Effects of tussah immunoreactive substances on growth, immunity, disease resistance against Vibrio splendidus and gut microbiota profile of Apostichopus japonicus. Fish Shellfish Immunol. 2017, 63, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Tian, B.; Sun, Y.; Li, X.; Mi, R.; Wen, Z.; Meng, N.; Li, J.; Li, S.; Du, X. Effect of Tussah Immunoreactive Substances and Tussah Antibacterial Peptide on Growth, and Immunity of Yesso Scallop Patinopecten yessoensis. Fish. Sci. 2023, 42, 878–883. (In Chinese) [Google Scholar]
- Li, M.; Cao, X.; Wei, C. Biological Features of Insect Antimicrobial Peptides and its Advances in the Development for Drug. Biotechnol. Commun. 2007, 18, 706–710. (In Chinese) [Google Scholar]
- Boman, H.G.; Nilsson, I.; Rasmuson, B. Inducible antibacterial defence system in Drosophila. Nature 1972, 237, 232–235. [Google Scholar] [CrossRef]
- Steiner, H.; Hultmark, D.; Engström, Å.; Bennich, H.; Boman, H.G. Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature 1981, 292, 246–248. [Google Scholar] [CrossRef]
- Keymanesh, K.; Soltani, S.; Sardari, S. Application of antimicrobial peptides in agriculture and food industry. World J. Microbiol. Biotechnol. 2009, 25, 933–944. [Google Scholar] [CrossRef]
- Tessera, V.; Guida, F.; Juretić, D.; Tossi, A. Identification of antimicrobial peptides from teleosts and anurans in expressed sequence tag databases using conserved signal sequences. FEBS J. 2012, 279, 724–736. [Google Scholar] [CrossRef]
- Viljakainen, L.; Pamilo, P. Selection on an antimicrobial peptide defensin in ants. J. Mol. Evol. 2008, 67, 643–652. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Marr, A.K.; Gooderham, W.J.; Hancock, R.E. Antibacterial peptides for therapeutic use: Obstacles and realistic outlook. Curr. Opin. Pharmacol. 2006, 6, 468–472. [Google Scholar] [CrossRef]
- Dong, X.Q.; Zhang, D.M.; Chen, Y.K.; Wang, Q.J.; Yang, Y.Y. Effects of antimicrobial peptides (AMPs) on blood biochemical parameters, antioxidase activity, and immune function in the common carp (Cyprinus carpio). Fish Shellfish Immunol. 2015, 47, 429–434. [Google Scholar] [CrossRef]
- Qin, Z.; Liang, J.; Long, S.; Niu, Z.; He, C.; Huang, J. Effects of antibacterial peptides on growth performance and body composition of Penaeus vannamei. J. South. Agric. 2016, 47, 674–678. (In Chinese) [Google Scholar]
- Chaturvedi, P.; Bhat, R.A.H.; Pande, A. Antimicrobial peptides of fish: Innocuous alternatives to antibiotics. Rev. Aquac. 2020, 12, 85–106. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- McBride, S.C.; Price, R.J.; Tom, P.D.; Lawrence, J.M.; Lawrence, A.L. Comparison of gonad quality factors: Color, hardness, and resilience, of Strongylocentrotus franciscanus between sea urchins fed prepared feed or algal diets and sea urchins harvested from the Northern California fishery. Aquaculture 2004, 233, 405–422. [Google Scholar] [CrossRef]
- Martinez, O.; Salmerón, J.; Guillén, M.; Casas, C. Texture profile analysis of meat products treated with commercial liquid smoke flavourings. Food Control 2004, 15, 457–461. [Google Scholar] [CrossRef]
- International, A. Official Methods of Analysis of AOAC International, 16th ed.; AOAC International: Rockville, MA, USA, 1995; Volume 1. [Google Scholar]
- Fontana, P. Nitrogen determination of Kjeldahl’s method. Mem. Do Inst. Oswaldo Cruz 1953, 51, 277–288. [Google Scholar] [CrossRef]
- Thiex, N.J.; Anderson, S.; Gildemeister, B. Crude fat, hexanes extraction, in feed, cereal grain, and forage (Randall/Soxtec/submersion method): Collaborative study. J. AOAC Int. 2003, 86, 899–908. [Google Scholar] [CrossRef]
- Zeng, F.; Wang, L.; Zhen, H.; Guo, C.; Liu, A.; Xia, X.; Pei, H.; Dong, C.; Ding, J. Nanoplastics affect the growth of sea urchins (Strongylocentrotus intermedius) and damage gut health. Sci. Total Environ. 2023, 869, 161576. [Google Scholar] [CrossRef]
- Huang, Y. Illumina-based analysis of endophytic bacterial diversity of four Allium species. Sci. Rep. 2019, 9, 15271. [Google Scholar] [CrossRef]
- Zhou, Y. Development and application of protein feed for insects. Feed Res. 2005, 02, 21–23. (In Chinese) [Google Scholar]
- Hammer, B.; Hammer, H.S.; Watts, S.A.; Desmond, R.; Lawrence, J.M.; Lawrence, A. The effects of dietary protein concentration on feeding and growth of small Lytechinus variegatus (Echinodermata: Echinoidea). Mar. Biol. 2004, 145, 1143–1157. [Google Scholar] [CrossRef]
- Sun, Y.; Ma, S.; Wang, Q.; Ren, T.; Du, X.; Li, S. Effects of dietary immunity induced pupae powder of Antheraea pernyi on growth, non-specific immunity and disease resistance of tiger puffer Takifugu rubripes. Fish. Sci. 2022, 41, 236–242. (In Chinese) [Google Scholar]
- McLaughlin, G.; Kelly, M.S. Effect of artificial diets containing carotenoid-rich microalgae on gonad growth and color in the sea urchin Psammechinus miliaris (GMELIN). J. Shellfish Res. 2001, 20, 377–382. [Google Scholar]
- Hammer, H.; Hammer, B.; Watts, S.; Lawrence, A.; Lawrence, J. The effect of dietary protein and carbohydrate concentration on the biochemical composition and gametogenic condition of the sea urchin Lytechinus variegatus. J. Exp. Mar. Biol. Ecol. 2006, 334, 109–121. [Google Scholar] [CrossRef]
- Ning, Y.; Zhang, F.; Tang, L.; Song, J.; Ding, J.; Chang, Y.; Zuo, R. Effects of dietary lipid sources on the growth, gonad development, nutritional and organoleptic quality, transcription of fatty acid synthesis related genes and antioxidant capacity during cold storage in adult sea urchin (Strongylocentrotus intermedius). Aquaculture 2022, 548, 737688. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, T.; Chiou, T.; Hwang, D. Seasonal variation on general composition, free amino acids and fatty acids in the gonad of Taiwan’s sea urchin Tripneustes gratilla. J. Mar. Sci. Technol. 2013, 21, 14. [Google Scholar]
- Cyrus, M.D.; Bolton, J.J.; Macey, B.M. The role of the green seaweed Ulva as a dietary supplement for full life-cycle grow-out of Tripneustes gratilla. Aquaculture 2015, 446, 187–197. [Google Scholar] [CrossRef]
- Symonds, R.C.; Kelly, M.S.; Caris-Veyrat, C.; Young, A.J. Carotenoids in the sea urchin Paracentrotus lividus: Occurrence of 9′-cis-echinenone as the dominant carotenoid in gonad colour determination. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2007, 148, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Symonds, R.C.; Kelly, M.S.; Suckling, C.C.; Young, A.J. Carotenoids in the gonad and gut of the edible sea urchin Psammechinus miliaris. Aquaculture 2009, 288, 120–125. [Google Scholar] [CrossRef]
- Alam, T.; Najam, L.; Al Harrasi, A. Extraction of natural pigments from marine algae. J. Agric. Mar. Sci. (JAMS) 2018, 23, 81–91. [Google Scholar] [CrossRef]
- Pearce, C.M.; Daggett, T.L.; Robinson, S.M. Effect of urchin size and diet on gonad yield and quality in the green sea urchin (Strongylocentrotus droebachiensis). Aquaculture 2004, 233, 337–367. [Google Scholar] [CrossRef]
- Zuo, R.; Hou, S.; Wu, F.; Song, J.; Zhang, W.; Zhao, C.; Chang, Y. Higher dietary protein increases growth performance, anti-oxidative enzymes activity and transcription of heat shock protein 70 in the juvenile sea urchin (Strongylocentrotus intermedius) under a heat stress. Aquac. Fish. 2017, 2, 18–23. [Google Scholar] [CrossRef]
- Zhang, X. Research advances on superoxide dismutase (SOD). Inn. Mong. Petrochem. 2010, 116, 14–15. [Google Scholar]
- Abd El-Gawad, E.A.; Abd El-latif, A.M.; Amin, A.A.; Abd-El-Azem, M. Effect of dietary fructooligosaccharide on bacterial Infection, oxidative stress, and histopathological alterations in Nile tilapia (Oreochromis niloticus). Glob. Vet. 2015, 15, 339–350. [Google Scholar]
- Zuo, R.; Mai, K.; Xu, W.; Turchini, G.M.; Ai, Q. Dietary ALA, but not LNA, increase growth, reduce inflammatory processes, and increase anti-oxidant capacity in the marine finfish Larimichthys crocea: Dietary ALA, but not LNA, increase growth, reduce inflammatory processes, and increase anti-oxidant capacity in the large yellow croaker. Lipids 2015, 50, 149–163. [Google Scholar]
- Li, Z.; Xie, S.; Wang, J.; Sales, J.; Li, P.; Chen, D. Effect of intermittent starvation on growth and some antioxidant indexes of Macrobrachium nipponense (De Haan). Aquac. Res. 2009, 40, 526–532. [Google Scholar] [CrossRef]
- Muniswamy, D.; Munaswamy, V.; Halappa, R.; Marigoudar, S. Impact of sodium cyanide on catalase activity in the freshwater exotic carp, Cyprinus carpio (Linnaeus). Pestic. Biochem. Physiol. 2008, 92, 15–18. [Google Scholar] [CrossRef]
- Li, H.; Feng, L.; Jiang, W.; Liu, Y.; Jiang, J.; Li, S.; Zhou, X.-Q. Oxidative stress parameters and anti-apoptotic response to hydroxyl radicals in fish erythrocytes: Protective effects of glutamine, alanine, citrulline, and proline. Aquat. Toxicol. 2013, 126, 169–179. [Google Scholar] [CrossRef]
- Chandra, R.K. Nutrition and the immune system: An introduction. Am. J. Clin. Nutr. 1997, 66, 460S–463S. [Google Scholar] [CrossRef]
- Ramírez, C.; Romero, J. Fine flounder (Paralichthys adspersus) microbiome showed important differences between wild and reared specimens. Front. Microbiol. 2017, 8, 271. [Google Scholar] [CrossRef]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef]
- Miao, S.; Han, B.; Hu, J.; Zhu, J.; Zhang, X.; Dong, X.; Chen, G. Effects of different dietary concentrations of tetracycline on growth performance, intestinal microbiota composition, and morphology of Channa argus. Chin. J. Anim. Nutr. 2019, 31, 5813–5822. (In Chinese) [Google Scholar]
- Wu, S.; Gao, T.; Zheng, Y.; Wang, W.; Cheng, Y.; Wang, G. Microbial diversity of intestinal contents and mucus in yellow catfish (Pelteobagrus fulvidraco). Aquaculture 2010, 303, 1–7. [Google Scholar] [CrossRef]
- Mackie, R.I.; White, B.A. Recent advances in rumen microbial ecology and metabolism: Potential impact on nutrient output. J. Dairy Sci. 1990, 73, 2971–2995. [Google Scholar] [CrossRef]
- Guo, W.; Zhou, X.; Li, X.; Zhu, Q.; Peng, J.; Zhu, B.; Zheng, X.; Lu, Y.; Yang, D.; Wang, B. Depletion of gut microbiota impairs gut barrier function and antiviral immune defense in the liver. Front. Immunol. 2021, 12, 636803. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lv, Z.; Guo, M. Research advancement of Apostichopus japonicus from 2000 to 2021. Front. Mar. Sci. 2022, 9, 931903. [Google Scholar] [CrossRef]
- Yang, M.; Song, H.; Sun, L.; Yu, Z.; Hu, Z.; Wang, X.; Zhu, J.; Zhang, T. Effect of temperature on the microflora community composition in the digestive tract of the veined rapa whelk (Rapana venosa) revealed by 16S rRNA gene sequencing. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 29, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Hoyles, L.; McCartney, A.L. What do we mean when we refer to Bacteroidetes populations in the human gastrointestinal microbiota? FEMS Microbiol. Lett. 2009, 299, 175–183. [Google Scholar] [CrossRef]
- Li, Y.-F.; Yang, N.; Liang, X.; Yoshida, A.; Osatomi, K.; Power, D.; Batista, F.M.; Yang, J.-L. Elevated seawater temperatures decrease microbial diversity in the gut of Mytilus coruscus. Front. Physiol. 2018, 9, 839. [Google Scholar] [CrossRef]
- Liu, G.; Luo, X.; Zhao, X.; Zhang, A.; Jiang, N.; Yang, L.; Huang, M.; Xu, L.; Ding, L.; Li, M. Gut microbiota correlates with fiber and apparent nutrients digestion in goose. Poult. Sci. 2018, 97, 3899–3909. [Google Scholar] [CrossRef]
- Kyselková, M.; Kopecký, J.; Frapolli, M.; Défago, G.; Ságová-Marečková, M.; Grundmann, G.L.; Moënne-Loccoz, Y. Comparison of rhizobacterial community composition in soil suppressive or conducive to tobacco black root rot disease. ISME J. 2009, 3, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Meirelles, P.M.; Mino, S.; Suda, W.; Oshima, K.; Hattori, M.; Thompson, F.L.; Sakai, Y.; Sawabe, T. Individual Apostichopus japonicus fecal microbiome reveals a link with polyhydroxybutyrate producers in host growth gaps. Sci. Rep. 2016, 6, 21631. [Google Scholar] [CrossRef]
- Pei, P.; Liu, X.; Chen, Y.; Wu, J.; Zhong, M.; Lin, Q.; Du, H. Effects of biological water purification grid on microbial community of culture environment and intestine of the shrimp Litopenaeus vannamei. Aquac. Res. 2019, 50, 1300–1312. [Google Scholar] [CrossRef]
- Ryan, M.P.; Adley, C.C. Ralstonia spp.: Emerging global opportunistic pathogens. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 291–304. [Google Scholar] [CrossRef]
- Fowler, E.C.; Poudel, P.; White, B.; St-Pierre, B.; Brown, M. Effects of a bioprocessed soybean meal ingredient on the intestinal microbiota of hybrid striped bass, Morone chrysops x M. saxatilis. Microorganisms 2021, 9, 1032. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Tajima, K.; Ezura, Y. Resuscitation of Tenacibaculum sp., the causative bacterium of spotting disease of sea urchin Strongylocentrotus intermedius, from the viable but non-culturable state. Fish. Sci. 2004, 70, 277–284. [Google Scholar] [CrossRef]
- Tajima, K.; Hirano, T.; Shimizu, M.; Ezura, Y. Isolation and pathogenicity of the causative bacterium of spotting disease of sea urchin Strongylocentrotus intermedius. Fish. Sci. 1997, 63, 249–252. [Google Scholar] [CrossRef]
- Hao, P.; Han, L.; Quan, Z.; Jin, X.; Li, Y.; Wu, Y.; Zhang, X.; Wang, W.; Gao, C.; Wang, L. Integrative mRNA-miRNA interaction analysis associated with the immune response of Strongylocentrotus intermedius to Vibrio harveyi infection. Fish Shellfish Immunol. 2023, 134, 108577. [Google Scholar] [CrossRef] [PubMed]
- Habibi, M.; Golmakani, M.T.; Eskandari, M.H.; Hosseini, S.M.H. Potential prebiotic and antibacterial activities of fucoidan from Laminaria japonica. Int. J. Biol. Macromol. 2024, 268, 131776. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Kelp Group | Feed Group | |||
|---|---|---|---|---|---|
| Control Group | 0.5% IPP Group | 1.0% IPP Group | 1.5% IPP Group | ||
| Survival rate/% | 100 ± 0 | 100 ± 0 | 100 ± 0 | 100 ± 0 | 100 ± 0 |
| Initial body weight/g | 17.26 ± 0.55 | 17.53 ± 0.53 | 17.18 ± 0.33 | 17.25 ± 0.58 | 17.14 ± 0.59 |
| Final body weight/g | 30.86 ± 0.34 c | 28.98 ± 0.41 b | 29.75 ± 0.22 a | 30.59 ± 0.19 a | 30.24 ± 0.30 a |
| Weight gain rate/% | 78.80 ± 2.11 c | 65.32 ± 2.14 b | 73.17 ± 1.82 a | 77.33 ± 1.73 a | 76.42 ± 2.16 a |
| Wet gonad weight/g | 6.20 ± 0.03 ab | 6.09 ± 0.06 b | 6.26 ± 0.02 ab | 6.58 ± 0.09 a | 6.41 ± 0.02 a |
| Gonad index/% | 20.09 ± 0.21 a | 21.01 ± 0.20 ab | 21.04 ± 0.15 ab | 21.51 ± 0.26 b | 21.20 ± 0.29 bc |
| Items | Kelp Group | Feed Group | |||
|---|---|---|---|---|---|
| Control Group | 0.5% IPP Group | 1.0% IPP Group | 1.5% IPP Group | ||
| L* 1 | 70.69 ± 1.41 c | 75.99 ± 1.21 a | 73.12 ± 1.68 b | 71.72 ± 1.20 bc | 75.90 ± 0.99 a |
| a* 2 | 28.12 ± 1.76 a | 23.09 ± 1.24 b | 24.78 ± 1.11 c | 26.51 ± 0.81 bc | 26.94 ± 0.66 bcd |
| b* 3 | 45.42 ± 1.97 a | 40.57 ± 1.27 c | 41.91 ± 2.15 b | 44.43 ± 2.06 ab | 45.13 ± 0.96 ab |
| ΔE1 4 | 15.10 ± 0.98 a | 21.79 ± 1.01 d | 19.37 ± 0.62 c | 16.36 ± 0.49 ab | 16.89 ± 0.44 bc |
| ΔE2 5 | 21.05 ± 1.23 a | 26.18 ± 1.35 d | 24.55 ± 1.26 c | 21.97 ± 0.98 b | 21.08 ± 1.33 ab |
| Items | Kelp Group | Feed Group | |||
|---|---|---|---|---|---|
| Control Group | 0.5% IPP Group | 1.0% IPP Group | 1.5% IPP Group | ||
| Hardness/N 1 | 1.30 ± 0.04 c | 1.21 ± 0.03 d | 1.31 ± 0.05 c | 1.41 ± 0.02 a | 1.35 ± 0.06 b |
| Adhesiveness/N·mm 2 | 0.24 ± 0.01 | 0.26 ± 0.03 | 0.25 ± 0.02 | 0.25 ± 0.06 | 0.24 ± 0.03 |
| Cohesiveness 3 | 0.23 ± 0.02 b | 0.23 ± 0.03 b | 0.24 ± 0.02 b | 0.27 ± 0.01 a | 0.26 ± 0.01 ab |
| Springiness/mm 4 | 0.26 ± 0.02 | 0.24 ± 0.02 | 0.24 ± 0.01 | 0.25 ± 0.03 | 0.25 ± 0.01 |
| Gumminess/N 5 | 0.30 ± 0.07 c | 0.27 ± 0.02 d | 0.31 ± 0.06 c | 0.37 ± 0.05 a | 0.35 ± 0.07 b |
| Chewiness/mJ 6 | 0.08 ± 0.01 a | 0.06 ± 0.01 c | 0.07 ± 0.02 b | 0.09 ± 0.01 a | 0.09 ± 0.03 a |
| Items | Kelp Group | Feed Group | |||
|---|---|---|---|---|---|
| Control Group | 0.5% IPP Group | 1.0% IPP Group | 1.5% IPP Group | ||
| GM | 71.29 ± 1.21 c | 73.51 ± 0.98 a | 73.26 ± 1.48 ab | 70.94 ± 3.35 b | 71.34 ± 1.09 c |
| CP | 12.56 ± 0.44 d | 12.97 ± 1.12 cd | 13.18 ± 0.67 c | 13.50 ± 0.62 b | 13.82 ± 0.92 a |
| CF | 5.06 ± 0.51 a | 4.80 ± 0.66 b | 4.84 ± 0.78 b | 5.01 ± 0.74 a | 4.96 ± 1.01 ab |
| Items | KELP Group | Feed Group | |||
|---|---|---|---|---|---|
| Control Group | 0.5% IPP Group | 1.0% IPP Group | 1.5% IPP Group | ||
| ACE | 203.09 ± 31.73 e | 672.87 ± 87.17 b | 872.2 ± 104.34 a | 479.84 ± 65.7 c | 283.12 ± 42.15 d |
| Chao1 | 201.2± 62.29 e | 678.06 ± 75.19 b | 850.80 ± 96.1 a | 481.77 ± 67.9 c | 289.11 ± 60.07 d |
| Shannon | 2.16 ± 0.36 c | 2.11 ± 0.32 c | 2.69 ± 0.38 ab | 3.45 ± 0.42 a | 2.61 ± 0.38 ab |
| Simpson | 0.25 ± 0.04 a | 0.29 ± 0.05 a | 0.21 ± 0.04 b | 0.11 ± 0.04 c | 0.15 ± 0.03 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Tian, F.; Li, W.; Xiao, H.; Tian, Y.; Deng, Y.; Han, L.; Zhao, C.; Ding, J. Effects of Dietary Tussah Immunoreactive Pupa Powder on Growth, Gonad Quality, Antioxidant Capacity, and Gut Microbiota of the Sea Urchin Strongylocentrotus intermedius. Biology 2025, 14, 874. https://doi.org/10.3390/biology14070874
Li S, Tian F, Li W, Xiao H, Tian Y, Deng Y, Han L, Zhao C, Ding J. Effects of Dietary Tussah Immunoreactive Pupa Powder on Growth, Gonad Quality, Antioxidant Capacity, and Gut Microbiota of the Sea Urchin Strongylocentrotus intermedius. Biology. 2025; 14(7):874. https://doi.org/10.3390/biology14070874
Chicago/Turabian StyleLi, Shufeng, Fenglin Tian, Weiyan Li, Haoran Xiao, Ye Tian, Yajie Deng, Lingshu Han, Chong Zhao, and Jun Ding. 2025. "Effects of Dietary Tussah Immunoreactive Pupa Powder on Growth, Gonad Quality, Antioxidant Capacity, and Gut Microbiota of the Sea Urchin Strongylocentrotus intermedius" Biology 14, no. 7: 874. https://doi.org/10.3390/biology14070874
APA StyleLi, S., Tian, F., Li, W., Xiao, H., Tian, Y., Deng, Y., Han, L., Zhao, C., & Ding, J. (2025). Effects of Dietary Tussah Immunoreactive Pupa Powder on Growth, Gonad Quality, Antioxidant Capacity, and Gut Microbiota of the Sea Urchin Strongylocentrotus intermedius. Biology, 14(7), 874. https://doi.org/10.3390/biology14070874