
Delimitation and Phylogeny in Fritillaria Species (Liliaceae) Endemic to Alps

 ,
,  , , , and
, , , and
Simple Summary
Abstract
1. Introduction
- (a)
- The phylogenetic relationships between F. tubaeformis and F. burnatii.
- (b)
- The phylogenetic placement of F. tubaeformis and F. burnatii with respect to the central European F. meleagris and to the SW Alpine endemic F. involucrata.
- (c)
- The taxonomic status of the two putative subspecies within F. tubaeformis.
2. Materials and Methods
2.1. DNA Extraction and PCR Amplification
2.2. Species Distribution Ranges in the F. tubaeformis Species Complex
2.3. Sequence Alignment, Dataset Assembly, and Phylogenetic Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | References | matK | ndhF | rpl16 | rpoC1 | petA-psbJ |
|---|---|---|---|---|---|---|
| F. burnatii (TEN1-20p2017) | this study | MN917987 | MN917996 | MN918014 | - | MN918005 |
| F. burnatii (TEN2-04p2013) | this study | - | MN917995 | MN918015 | - | MN918004 |
| F. burnatii (SER-05p2013) | this study | MN917986 | - | MN918013 | MN918023 | MN918003 |
| F. burnatii (TN01) | this study | - | OP021726 | - | OP021738 | OP021733 |
| F. burnatii (TN02) | this study | - | OP021727 | - | OP021739 | OP021732 |
| F. burnatii (TN03) | this study | - | OP021728 | - | OP021740 | OP021734 |
| F. burnatii (TN04) | this study | - | OP021729 | - | OP021741 | OP021735 |
| F. involucrata (MEN-14p2016) | this study | MN917988 | MN917997 | MN918016 | - | - |
| F. involucrata (HYE-CB976) | this study | - | OP021730 | OP021736 | OP021742 | - |
| F. meleagris (GER-11p2015) | this study | MN917989 | MN917998 | MN918020 | MN918025 | MN918008 |
| F. meleagris (Chase 2566 (K), Kew 1990-3088) | [11] | AY624445 | AY624393 | |||
| F. moggridgei (MAR-07p2014) | this study | MN917992 | MN917999 | PV832077 | - | MN918012 |
| F. moggridgei (CRA-12p2014) | this study | MN917990 | PV832078 | MN918017 | - | MN918009 |
| F. moggridgei (FRO-00p2013) | this study | MN917991 | - | MN918021 | MN918027 | MN918010 |
| F. moggridgei (PLU-08p2014) | this study | PV832076 | MN918000 | MN918022 | MN918026 | MN918011 |
| F. moggridgei (HYE-CB6181) | this study | - | OP021731 | OP021737 | - | - |
| F. tubaeformis (GLE-10p2013) | this study | MN917993 | MN918001 | MN918018 | MN918028 | MN918006 |
| F. tubaeformis (GLE-09p2015) | this study | MN917994 | MN918002 | MN918019 | MN918024 | MN918007 |
| L. superbum | [52] | KP462883 | KP462883 | KP462883 | KP462883 | KP462883 |
| L. bakerianum | [53] | KY748301 | KY748301 | KY748301 | KY748301 | KY748301 |
| Taxa | References | ITS |
|---|---|---|
| F. burnatii (TEN1-20p2017) | this study | MT522874 |
| F. burnatii (TEN2-04p2013) | this study | MT522877 |
| F. burnatii (SER-05p2013) | this study | PV816804 |
| F. involucrata (MEN-14p2016) | this study | MT522876 |
| F. meleagris (Chase 2566 (K)) | [11] | AY616730 |
| F. meleagris (K:DNA:MWC12064) | [54] | HE656029 |
| F. moggridgei (MAR-07p2014) | this study | PV816806 |
| F. moggridgei (CRA-12p2014) | this study | MT522875 |
| F. moggridgei (PLU-08p2014) | this study | PV816805 |
| F. tubaeformis (GLE-10p2013) | this study | MT522878 |
| F. tubaeformis (GLE-09p2015) | this study | MT522879 |
| L. superbum | [55] | JF829386 |
| L. bakerianum (G2013001) | [56] | KF851093 |
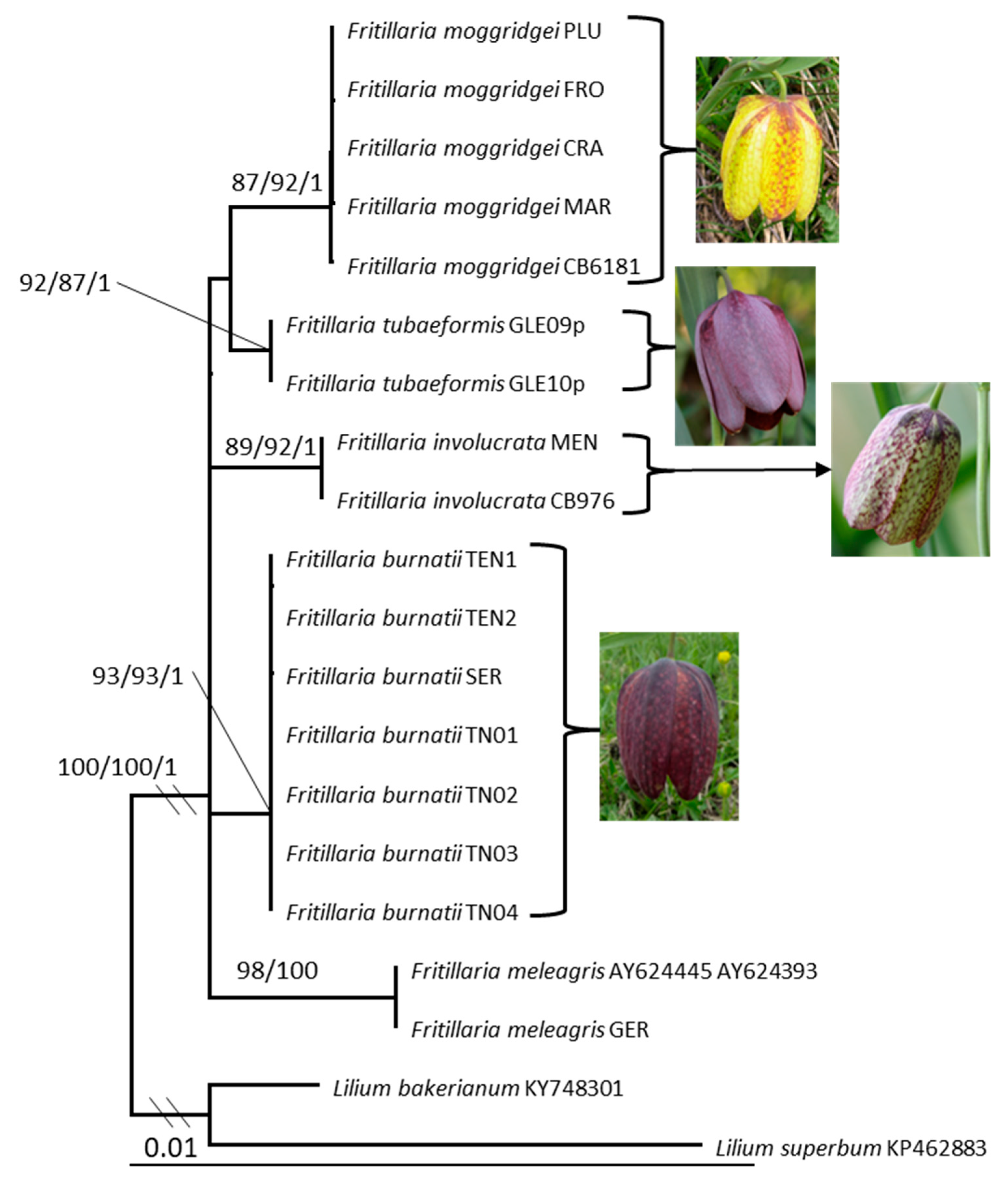
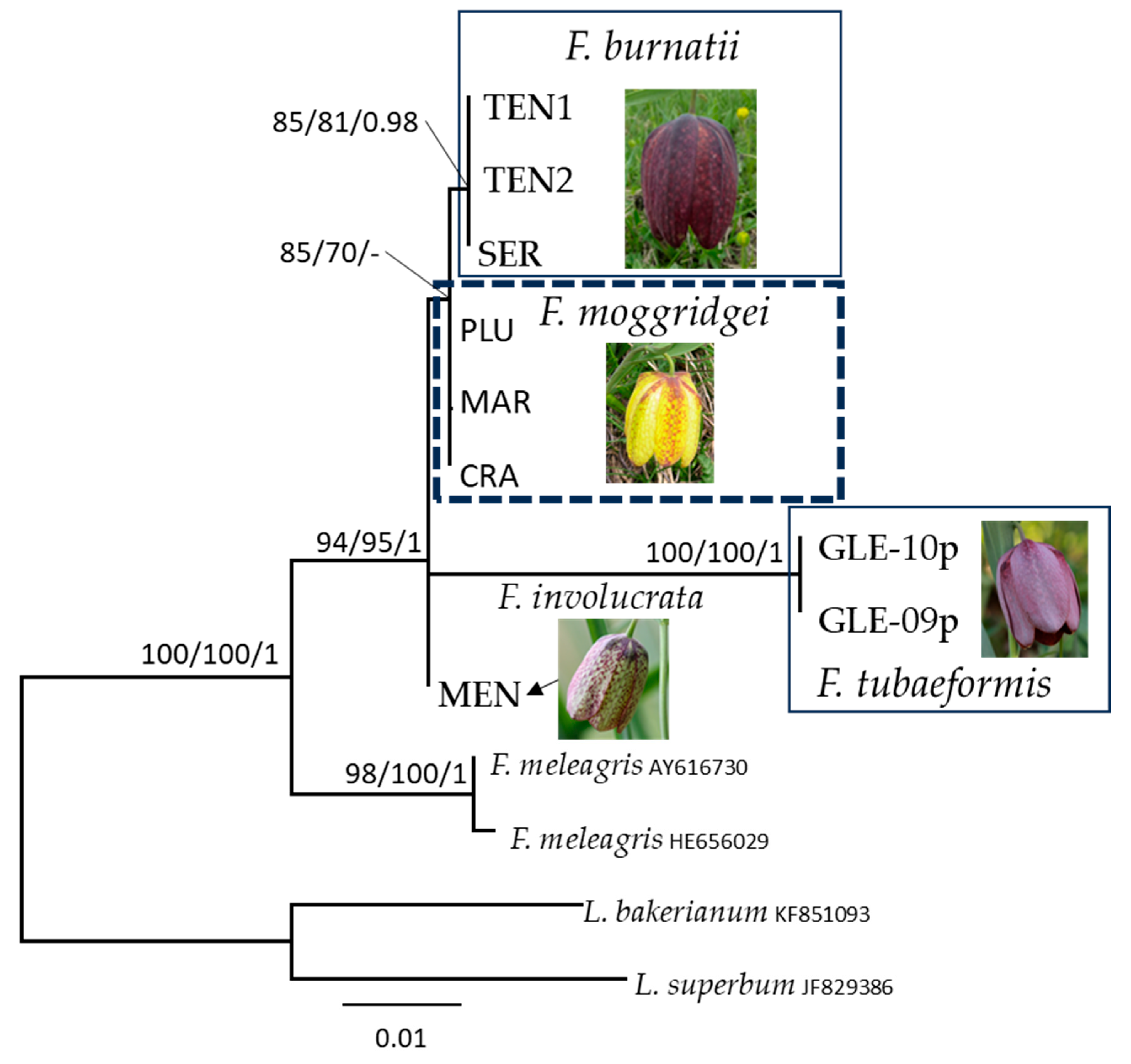
3. Results
Molecular and Phylogenetic Analysis
4. Discussion
4.1. Fritillaria meleagris
4.2. Fritillaria burnatii
4.3. Fritillaria tubaeformis and F. moggridgei
4.4. Fritillaria involucrata
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tekşen, M.; Aytaç, Z. The revision of the genus Fritillaria L. (Liliaceae) in the Mediterranean region (Turkey). Turk J. Bot. 2011, 35, 447–478. [Google Scholar] [CrossRef]
- Givnish, T.J.; Zuluaga, A.; Marques, I.; Lam, V.K.; Gomez, M.S.; Iles, W.J.; Ames, M.; Spalink, D.; Moeller, J.R.; Briggs, B.G. Phylogenomics and historical biogeography of the monocot order Liliales: Out of Australia and through Antarctica. Cladistics 2016, 32, 581–605. [Google Scholar] [CrossRef]
- Smyčka, J.; Roquet, C.; Renaud, J.; Thuiller, W.; Zimmermann, N.E.; Lavergne, S. Disentangling drivers of plant endemism and diversification in the European Alps—A phylogenetic and spatially explicit approach. Perspect. Plant Ecol. Evol. Syst. 2017, 28, 9–27. [Google Scholar] [CrossRef]
- Ebersbach, J.; Schnitzler, J.; Favre, A.; Muellner-Riehl, A.N. Evolutionary radiations in the species-rich mountain genus Saxifraga L. BMC Evol. Biol. 2017, 17, 119. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biog. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Casazza, G.; Barberis, G.; Guerrina, M.; Zappa, E.; Mariotti, M.; Minuto, L. The plant endemism in the Maritime and Ligurian Alps. Biogeographia 2016, 31, 73–88. [Google Scholar] [CrossRef]
- Schönswetter, P.; Stehlik, I.; Holderegger, R.; Tribsch, A. Molecular evidence for glacial refugia of mountain plants in the European Alps. Mol. Ecol. 2005, 14, 3547–3555. [Google Scholar] [CrossRef]
- Holderegger, R.; Thiel-Egenter, C. A discussion of different types of glacial refugia used in mountain biogeography and phylogeography. J. Biogeogr. 2009, 36, 476–480. [Google Scholar] [CrossRef]
- Rix, E.M. Fritillaria. A Revised Classification Together with an Updated List of Species; The Fritillaria Group of the Alpine Garden Society: Edinburgh, UK, 2001; pp. 1–14. [Google Scholar]
- Peruzzi, L. A new infrafamilial taxonomic setting for Liliaceae, with a key to genera and tribes. Plant Biosyst. 2016, 150, 1341–1347. [Google Scholar] [CrossRef]
- Rønsted, N.; Law, S.; Thornton, H.; Fay, M.F.; Chase, M.W. Molecular phylogenetic evidence for the monophyly of Fritillaria and Lilium (Liliaceae; Liliales) and the infrageneric classification of Fritillaria. Molec. Phyl. Evol. 2005, 35, 509–527. [Google Scholar] [CrossRef] [PubMed]
- Day, P.D.; Berger, M.; Hill, L.; Fay, M.F.; Leitch, A.R.; Leitch, I.J.; Kelly, L.J. Evolutionary relationships in the medicinally important genus Fritillaria L. (Liliaceae). Molec. Phyl. Evol. 2014, 80, 11–19. [Google Scholar] [CrossRef]
- Huang, J.; Yang, L.Q.; Yu, Y.; Liu, Y.M.; Xie, D.F.; Li, J.; He, X.J.; Zhou, S.D. Molecular phylogenetics and historical biogeography of the tribe Lilieae (Liliaceae): Bi-directional dispersal between biodiversity hotspots in Eurasia. Ann. Bot. 2018, 122, 1245–1262. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D. Plant Evolution in the Mediterranean; Oxford University Press: Oxford, UK, 2005; ISBN 9780198515340. [Google Scholar]
- Huang, J.; Yu, Y.; Liu, Y.M.; Xie, D.F.; He, X.J.; Zhou, S.D. Comparative chloroplast genomics of Fritillaria (Liliaceae), inferences for phylogenetic relationships between Fritillaria and Lilium and plastome evolution. Plants 2020, 9, 133. [Google Scholar] [CrossRef] [PubMed]
- Kadereit, J.W. The role of in situ species diversification for the evolution of high vascular plant species diversity in the European Alps—A review and interpretation of phylogenetic studies of the endemic flora of the Alps. Perspect. Plant Ecol. Evol. Syst. 2017, 26, 28–38. [Google Scholar] [CrossRef]
- Rix, E.M. Fritillaria involucrata: Liliaceae. Curtis’s Bot. Mag. 2000, 17, 179–181. [Google Scholar] [CrossRef]
- Noble, V.; Diadema, K. La Flore des Alpes-Maritimes et de la Principauté de Monaco. Originalité et Diversité; Naturalia Publications: Turriers, France, 2011; p. 504. [Google Scholar]
- Tison, J.M.; Jauzein, P.J.; Michaud, H. Flore de la France Méditerranèenne Continentale; Naturalia Publications: Turriers, France, 2014; pp. 1–2080. ISBN 9782909717906. [Google Scholar]
- Selvaggi, A.; Ebone, A.; Bombonati, D.; Gallino, B. Fritillaria involucrata All. (Liliaceae). In: Selvaggi, A.; Soldano, A.; Pascale, M.; Pascal, R. (eds.). Note Floristiche Piemontesi n. 176-245. Riv. Piem. St. Nat. 2009, 30, 313–340. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Alessandrini, A.; Ardenghi, N.M.G.; Bacchetta, G.; Banfi, E. A second update to the checklist of the vascular flora native to Italy. Plant Biosyst. 2024, 158, 219–296. [Google Scholar] [CrossRef]
- Available online: https://www.gbif.org/occurrence/search?/ (accessed on 8 April 2022).
- Mucciarelli, M.; Rosso, P.; Noble, V.; Bartolucci, F.; Peruzzi, L. A morphometric study and taxonomic revision of Fritillaria tubaeformis species complex (Liliaceae). Plant Syst. Evol. 2016, 302, 1329–1343. [Google Scholar] [CrossRef]
- Fiori, A. Nuova Flora Analitica d’Italia; Tipografia di M. Ricci: Firenze, Italy, 1923. [Google Scholar]
- Pignatti, S. Flora d’Italia 1; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Aeschimann, D.; Lauber, K.; Moser, D.M.; Theurillat, J.-P. Flora Alpina 1; Zanichelli: Bologna, Italy, 2004. [Google Scholar]
- Zangheri, P. Flora Italica; Cedam: Padova, Italy, 1976. [Google Scholar]
- Conti, F.; Abbate, G.; Alessandrini, A.; Blasi, C. An Annotated Checklist of the Italian Vascular Flora; Palombi Editori: Roma, Italy, 2005; pp. 1–428. [Google Scholar]
- Rix, E.M. Short notes (Liliaceae). Bot. J. Linn. Soc. 1978, 76, 356. [Google Scholar]
- Rix, E.M. Fritillaria L. In Flora Europaea; Tutin, T.G., Burges, N.A., Chater, A.O., Edmondson, J.R., Heywood, V.H., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1980; pp. 31–34. [Google Scholar]
- Tison, J.M.; de Foucault, B. Flora Gallica: Flore de France; Biotope Èditions: Mèze, France, 2014; pp. 1–1196. [Google Scholar]
- Mucciarelli, M.; Fay, M.F. Plastid DNA fingerprinting of the rare Fritillaria moggridgei (Liliaceae) reveals population differentiation and genetic isolation within the Fritillaria tubiformis complex. Phytotaxa 2013, 91, 1–23. [Google Scholar] [CrossRef]
- Mucciarelli, M.; Ferrazzini, D.; Belletti, P. Genetic variability and population divergence in the rare Fritillaria tubiformis subsp. moggridgei Rix (Liliaceae) as revealed by RAPD analysis. PLoS ONE 2014, 9, e101967. [Google Scholar] [CrossRef]
- Bartolucci, F.; Peruzzi, L. Typification of Fritillaria tubiformis Gren. & Godr., Fritillaria delphinensis f. moggridgei Planch. and Fritillaria delphinensis var. burnatii Planch (Liliaceae) from SW Europe. Candollea 2012, 67, 23–29. [Google Scholar] [CrossRef]
- Wang, H.; Qi, M.; Cutler, A.J. A simple method of preparing plant samples for PCR. Nuc. Acids Res. 1993, 21, 4153–4154. [Google Scholar] [CrossRef]
- Ito, M.A.; Kawamoto, Y.; Yukawa, K.T.; Kurita, S. Phylogenetic relationships of Amaryllidaceae based on matK sequence data. J. Plant Res. 1999, 112, 207–216. [Google Scholar] [CrossRef]
- Olmstead, R.G.; Sweere, J.A. Combining data in phylogenetic systematics: An empirical approach using three molecular data sets in the Solanaceae. Syst. Biol. 1994, 43, 67–481. [Google Scholar] [CrossRef]
- Marques, I.; Nieto Feliner, G.; Draper, D.; Martins-Loução, M.A.; Fuertes Aguilar, J. Unravelling cryptic reticulate relationships and the origin of orphan hybrid disjunct populations in Narcissus. Evolution 2010, 64, 2353–2368. [Google Scholar] [CrossRef]
- Cheng, T.; Xu, C.; Lei, L.; Li, C.; Zhang, Y.; Zhou, S. Barcoding the kingdom Plantae: New PCR primers for ITS regions of plants with improved universality and specificity. Mol. Ecol. Resour. 2016, 16, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Dauby, G.; Stévart, T.; Droissart, V.; Cosiaux, A.; Deblauwe, V.; Simo-Droissart, M.; Sosef, M.S.M.; Lowry, P.P.; Schatz, G.E.; Gereau, R.E.; et al. ConR: An R package to assist large-scale multispecies preliminary conservation assessments using distribution data. Ecol. Evol. 2017, 7, 11292–11303. [Google Scholar] [CrossRef]
- GBIF.org GBIF Occurrence Download. Available online: https://www.gbif.org/occurrence/download/0252572-210914110416597 (accessed on 28 April 2022). [CrossRef]
- GBIF.org GBIF Occurrence Download. Available online: https://www.gbif.org/occurrence/download/0252573-210914110416597 (accessed on 28 April 2022). [CrossRef]
- Argagnon, A.; De Barros, G.; Noble, V. SIMETHIS-Flore-CBNMed-Database of Southeastern France vegetation. Veg. Classif. Surv. 2022, 3, 119–120. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Le Vinh, S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological hylogenetic analyses. Molec. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.F.K.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Mennes, C.B.; Lam, V.K.Y.; Rudall, P.J.; Lyon, S.P.; Graham, S.W.; Smets, E.F.; Merckx, V.S.F.T. Ancient Gondwana break-up explains the distribution of the mycoheterotrophic family Corsiaceae (Liliales). J. Biogeogr. 2015, 42, 1123–1136. [Google Scholar] [CrossRef]
- Du, Y.P.; Bi, Y.; Yang, F.P.; Zhang, M.F.; Chen, X.Q.; Xue, J.; Zhang, X.H. Complete chloroplast genome sequences of Lilium: Insights into evolutionary dynamics and phylogenetic analyses. Sci. Rep. 2017, 7, 5751. [Google Scholar] [CrossRef]
- Clennett, J.C.B.; Chase, M.W.; Forest, F.; Maurin, O.; Wilkin, P. Phylogenetic systematics of Erythronium (Liliaceae): Morphological and molecular analyses. Bot. J. Linn. Soc. 2012, 170, 504–528. [Google Scholar] [CrossRef]
- Douglas, N.A.; Wall, W.A.; Xiang, Q.Y.; Hoffmann, W.A.; Wentworth, T.R.; Gray, J.B.; Hohmann, M.G. Recent Vicariance and the Origin of the Rare, Edaphically Specialized Sandhills Lily, Lilium pyrophilum (Liliaceae): Evidence from Phylogenetic and Coalescent Analyses. Mol. Ecol. 2021, 20, 2901–2915. [Google Scholar] [CrossRef]
- Gao, Y.D.; Harris, A.; He, X.J. Morphological and ecological divergence of Lilium and Nomocharis within the Hengduan Mountains and Qinghai-Tibetan Plateau may result from habitat specialization and hybridization. BMC Evol. Biol. 2015, 15, 147. [Google Scholar] [CrossRef] [PubMed]
- Day, P. Studies in the genus Fritillaria L. (Liliaceae). Ph.D. Thesis, Queen Mary University, London, UK, 2017. [Google Scholar]
- Turrill, W.B. The Snake’s Head (Fritillaria meleagris). In The Lily Year Book 1951-2 (Number Fifteen); Synge, P.M., Ed.; Royal Horticultural Society: London, UK, 1951; pp. 108–116. [Google Scholar]
- Tatarenko, I.; Walker, K.; Dyson, M. Biological Flora of Britain and Ireland: Fritillaria meleagris. J. Ecol. 2022, 299, 1704–1726. [Google Scholar] [CrossRef]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia 2; Edagricole: Bologna, Italy, 2017. [Google Scholar]
- Bovio, M. Flora vascolare della Valle d’Aosta [Vascular flora of Valle d’Aosta]. Testolin Editore: Aosta, Italy, 2014; p. 335. ISBN 978-88-909-4664-6. [Google Scholar]
- Kamari, G.; Zahos, A.; Siagou, I. A new yellow-flowered Fritillaria species (Liliaceae) from Mt. Tisseon, continental Greece and its taxonomic relationships. Phytotaxa 2017, 328, 227–242. [Google Scholar] [CrossRef]
- Planchon, J.E. Sur les espèces de fritillaires de France, à propos des Icones et d’un manuscrit inédit de Pierre de Belleval. Bull. Soc. Bot. France 1873, 20, 96–124. [Google Scholar] [CrossRef]
- Dagnino, D.; Minuto, L.; Casazza, G. Niche divergence between putative taxa: Ecological niche models in taxonomic researches. Plant Biol. 2017, 19, 1003–1011. [Google Scholar] [CrossRef]
- Kelly, L.; Renny-Byfield, S.; Pellicer, J.; Macas, J.; Novak, P.; Neumann, P.; Lysak, M.A.; Day, P.D.; Berger, M.; Fay, M.F.; et al. Analysis of the giant genomes of Fritillaria (Liliaceae) indicates that a lack of DNA removal characterises extreme expansions in genome size. New Phytol. 2015, 208, 596–607. [Google Scholar] [CrossRef] [PubMed]


| Taxa | DNA Voucher Code (Herbarium Specimen) a | Country | Site of Sampling | Locality |
|---|---|---|---|---|
| Fritillaria burnatii | TEN1-20p2017 (TO-HP 7481) | Italy | Maritime Alps | Colle di Tenda (Cuneo) |
| Fritillaria burnatii | TEN2-04p2013 | Italy | Maritime Alps | Colle di Tenda (Cuneo) |
| Fritillaria burnatii | SER-05p2013 (TO-HP 7482) | Italy | Ligurian Alps | Vallone di Serpentera (Cuneo) |
| Fritillaria burnatii | TN01 | Italy | Trentino-Alto Adige | Bondo–Breguzzo–Daone (Trento) |
| Fritillaria burnatii | TN02 | Italy | Trentino-Alto Adige | Bondo–Breguzzo–Daone (Trento) |
| Fritillaria burnatii | TN03 | Italy | Trentino-Alto Adige | Bondo–Breguzzo–Daone (Trento) |
| Fritillaria burnatii | TN04 | Italy | Trentino-Alto Adige | Bondo–Breguzzo–Daone (Trento) |
| Fritillaria involucrata | MEN-14p2016 (TO-HG 3558) | Italy | Ligurian Alps | San Bernardo di Mendatica (Imperia) |
| Fritillaria involucrata | HYE-CB976 | France | Maritime Alps | Saint-Vallier-de-Thiey (Grasse) |
| Fritillaria meleagris | GER-11p2015 | Germany | Cultivated seedlings | Hortus Botanicus Taurinensis |
| Fritillaria moggridgei | MAR-07p2014 (TO-HP 7485) | Italy | Ligurian Alps | Vallone del Marguareis (Cuneo) |
| Fritillaria moggridgei | CRA-12p2014 | Italy | Ligurian Alps | Vallone di Cravina (Cuneo) |
| Fritillaria moggridgei | FRO-00p2013 | Italy | Ligurian Alps | Monte Frontè (Imperia) |
| Fritillaria moggridgei | PLU-08p2014 (TO-HP 7484) | Italy | Ligurian Alps | Pian del Lupo (Cuneo) |
| Fritillaria moggridgei | HYE-CB6181 | France | Maritime Alps | Tende (France) |
| Fritillaria tubaeformis | GLE-10p2013 | France | Hautes Alpes | Col de Gleize (Gap) |
| Fritillaria tubaeformis | GLE-09p2015 (TO-HG 3328) | France | Hautes Alpes | Col de Gleize (Gap) |
| Lilium superbum | Chase, M.W. 112 | n/a | - | - |
| Lilium bakerianum | BOP040929 | China | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dovana, F.; Peruzzi, L.; Noble, V.; Adamo, M.; Bonomi, C.; Mucciarelli, M. Delimitation and Phylogeny in Fritillaria Species (Liliaceae) Endemic to Alps. Biology 2025, 14, 785. https://doi.org/10.3390/biology14070785
Dovana F, Peruzzi L, Noble V, Adamo M, Bonomi C, Mucciarelli M. Delimitation and Phylogeny in Fritillaria Species (Liliaceae) Endemic to Alps. Biology. 2025; 14(7):785. https://doi.org/10.3390/biology14070785
Chicago/Turabian StyleDovana, Francesco, Lorenzo Peruzzi, Virgile Noble, Martino Adamo, Costantino Bonomi, and Marco Mucciarelli. 2025. "Delimitation and Phylogeny in Fritillaria Species (Liliaceae) Endemic to Alps" Biology 14, no. 7: 785. https://doi.org/10.3390/biology14070785
APA StyleDovana, F., Peruzzi, L., Noble, V., Adamo, M., Bonomi, C., & Mucciarelli, M. (2025). Delimitation and Phylogeny in Fritillaria Species (Liliaceae) Endemic to Alps. Biology, 14(7), 785. https://doi.org/10.3390/biology14070785