


Effects of Chronic Low-Salinity Stress on Growth, Survival, Antioxidant Capacity, and Gene Expression in Mizuhopecten yessoensis

 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Animals and Stress Design
2.3. Sample Collection
2.4. Growth and Survival Assessment
2.5. Measurement of Oxygen Consumption and Ammonia Excretion Rates
2.6. Enzyme Activity Assays
2.7. Sample Collection and RNA Extraction
2.8. Library Construction and High-Throughput Sequencing
2.9. Quality Control and Sequence Assembly
2.10. Differential Gene Expression Analysis
2.11. Data Analysis
3. Results
3.1. Effects of Different Low-Salinity Levels on Shell Length Growth Rate, Wet Weight Gain, and Mortality of M. yessoensis
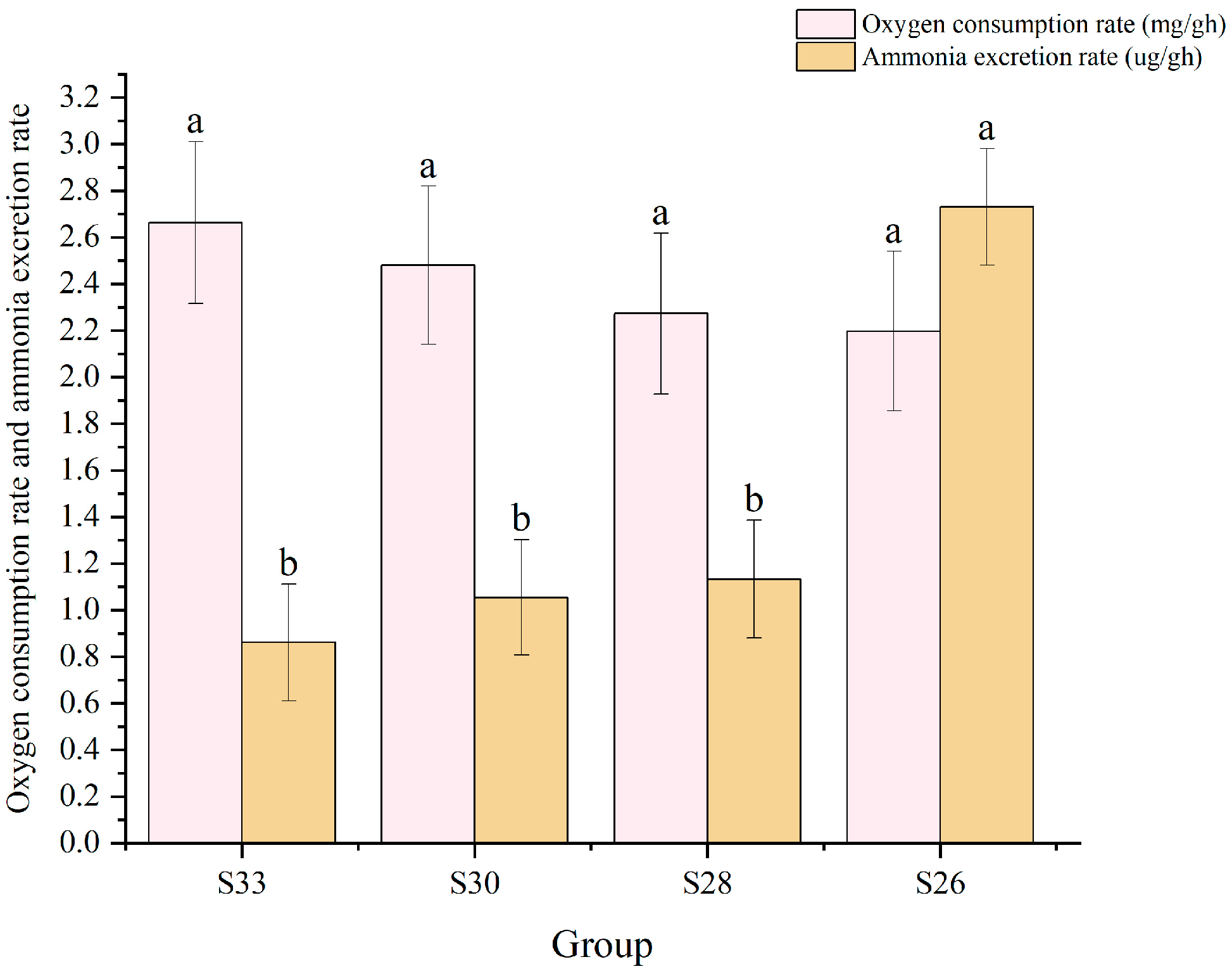
3.2. Effects of Different Low-Salinity Levels on Oxygen Consumption and Ammonia Excretion
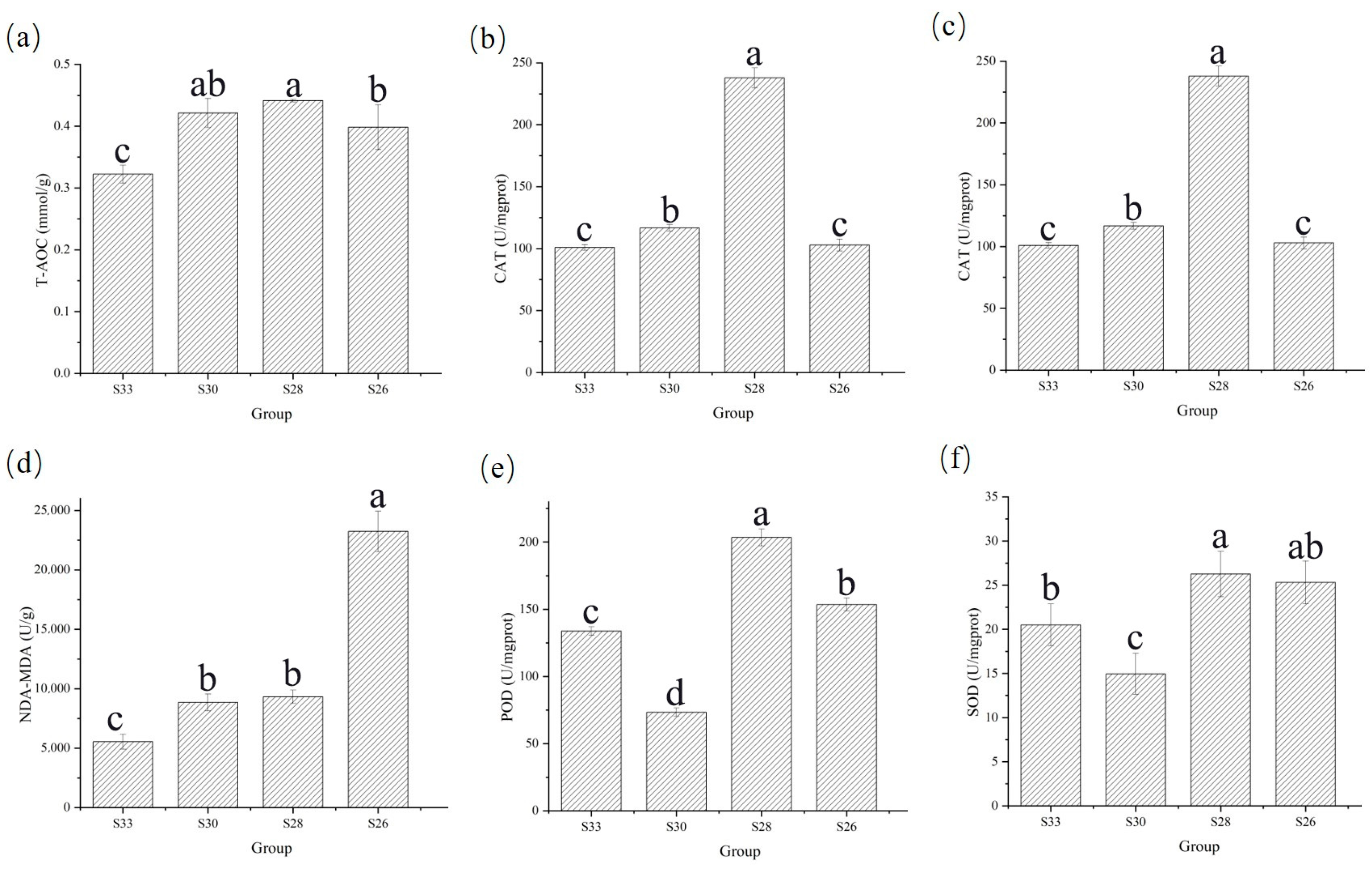
3.3. Effects of Different Low-Salinity Levels on Enzyme Activities in M. yessoensis
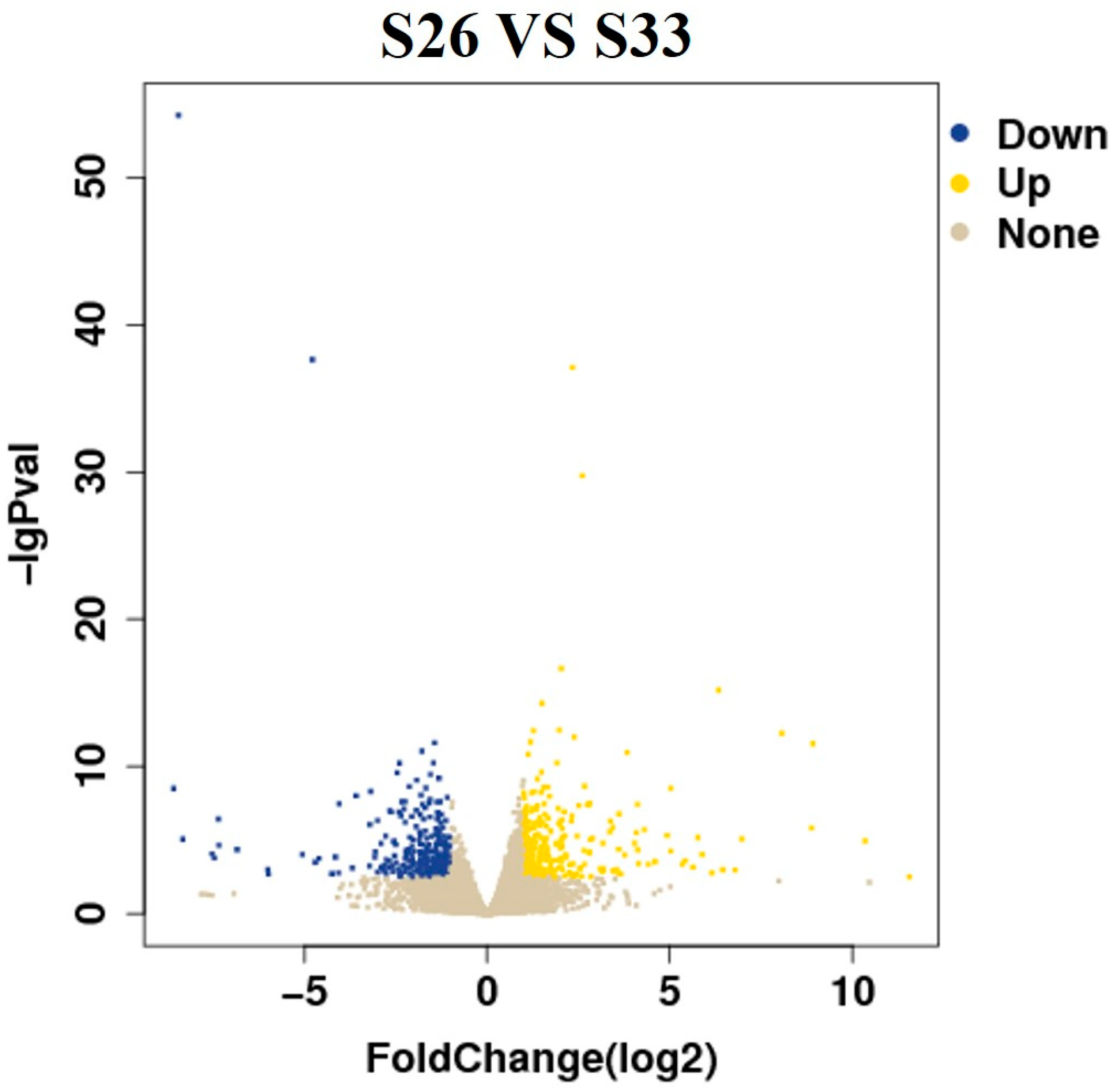
3.4. Differential Gene Expression
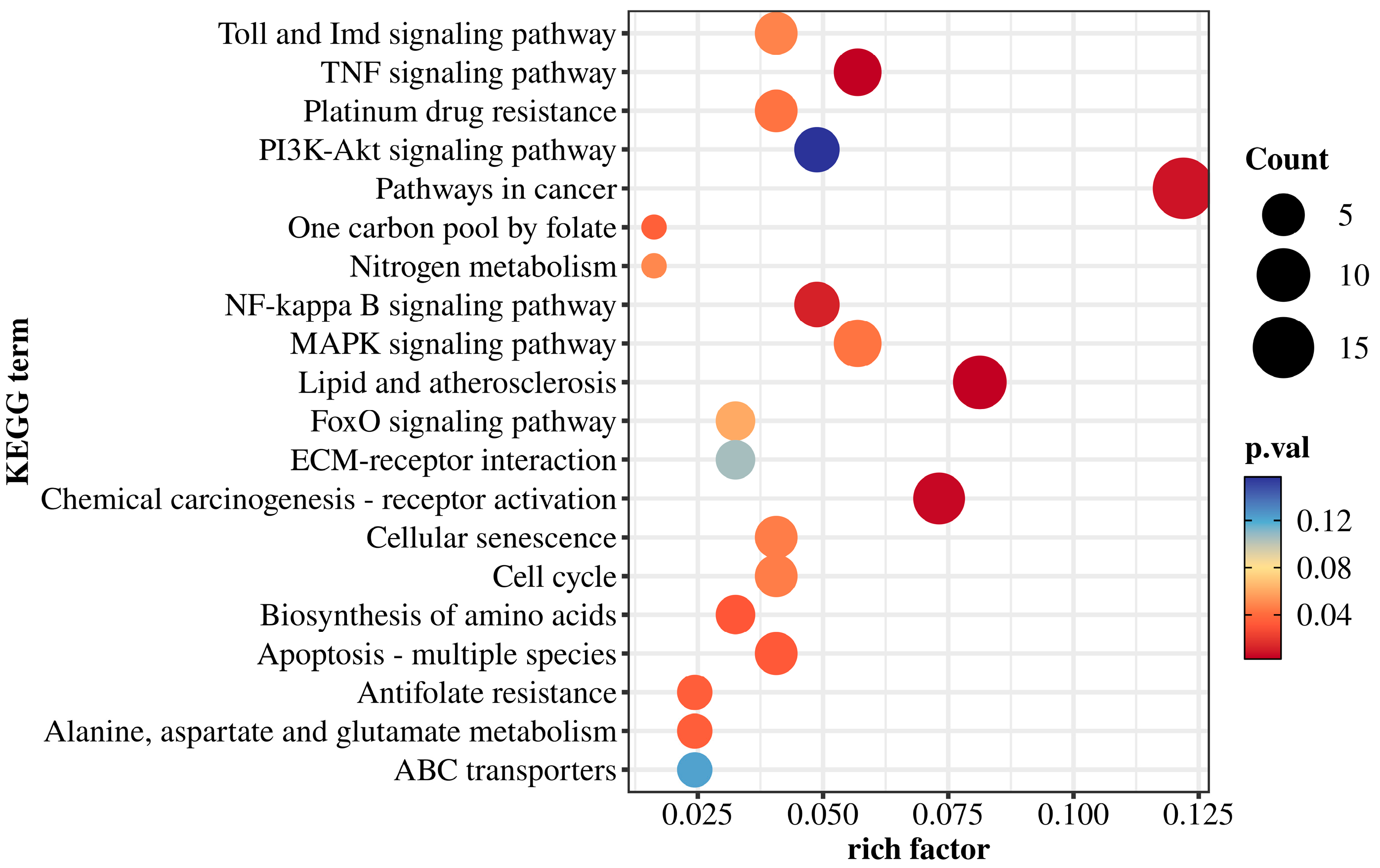
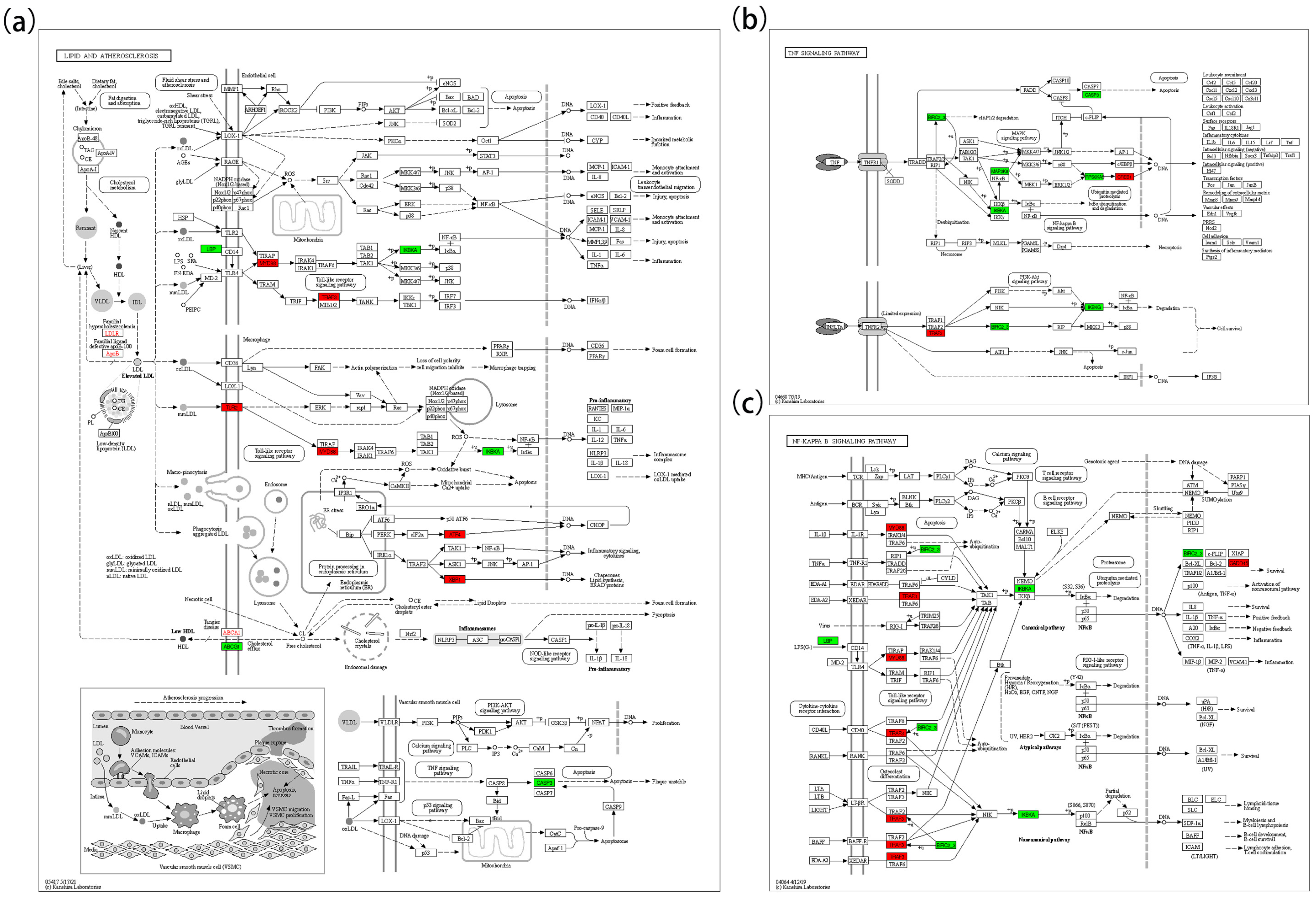
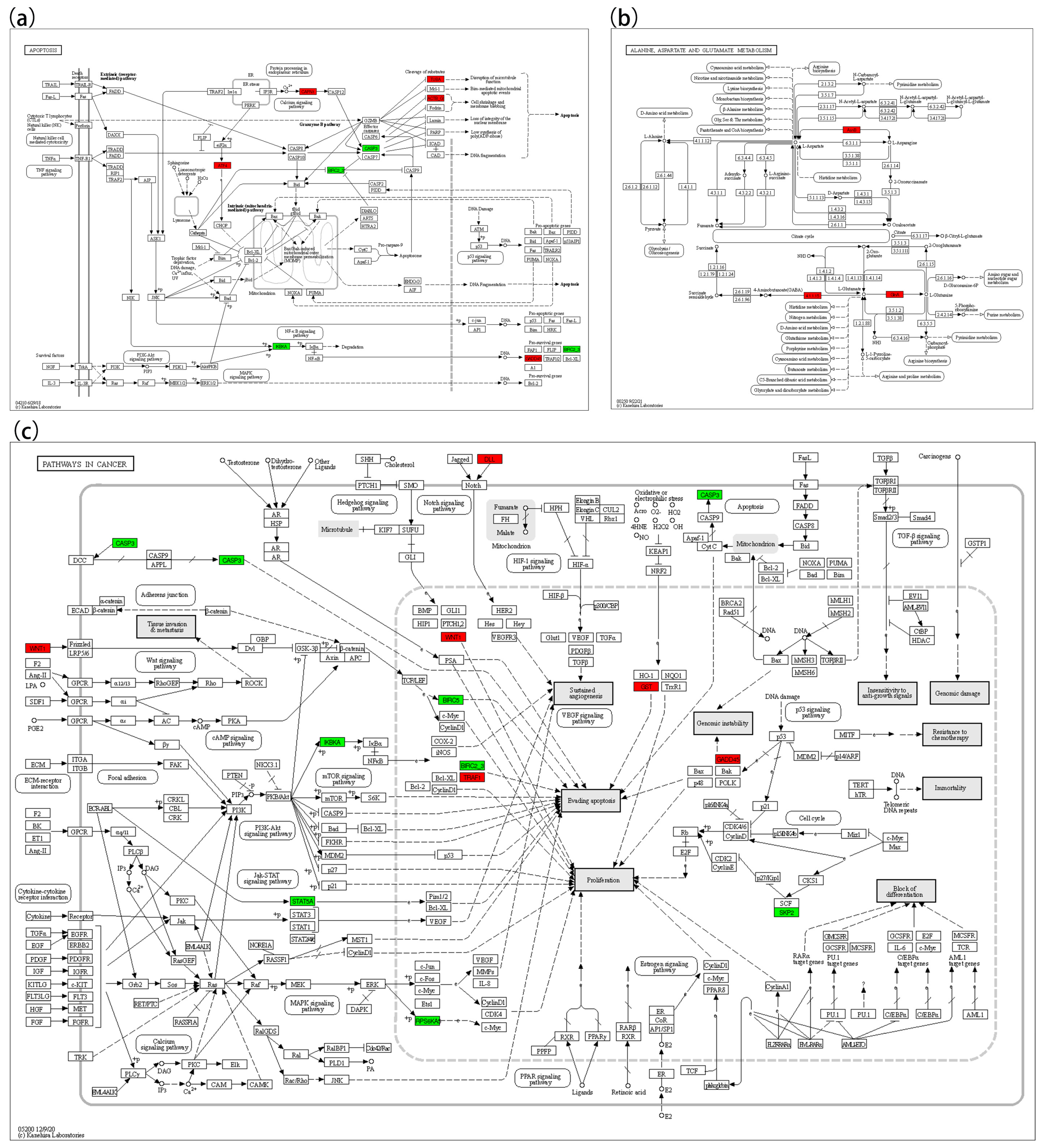
3.5. KEGG Pathway Enrichment
3.6. qRT-PCR and RNA-Seq
4. Discussion
4.1. Antioxidant Response
4.2. Regulation of Apoptosis Signaling
4.3. Adaptation of Energy Metabolism
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nan, X.; Wei, H.; Zhang, H.; Nie, H. Factors influencing the interannual variation in biomass of bottom-cultured Yesso scallop (Patinopecten yessoensis) in the Changhai Sea area, China. Front. Mar. Sci. 2022, 8, 798359. [Google Scholar] [CrossRef]
- Newell, R.I. Ecosystem influences of natural and cultivated populations of suspension-feeding bivalve molluscs: A review. J. Shellfish Res. 2004, 23, 51–62. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Biological aspects, fisheries, and aquaculture of yesso scallops in Russian waters of the Sea of Japan. Diversity 2022, 14, 399. [Google Scholar] [CrossRef]
- Namikawa, Y.; Suzuki, M. Atmospheric CO2 Sequestration in Seawater Enhanced by Molluscan Shell Powders. Environ. Sci. Technol. 2024, 58, 2404–2412. [Google Scholar] [CrossRef]
- Li, X.; Li, D.; Tian, Y.; Chang, Y.; Hao, Z. Behavioral characteristics and related physiological and ecological indexes of cultured scallops (Mizuhopecten yessoensis) in response to predation by the crab Charybdis japonica. Fishes 2024, 9, 389. [Google Scholar] [CrossRef]
- Gibson, R.; Barnes, M.; Atkinson, R. Impact of changes in flow of freshwater on estuarine and open coastal habitats and the associated organisms. Oceanogr. Mar. Biol. Annu. Rev. 2002, 40, 233. [Google Scholar] [CrossRef]
- Mackay, A. Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. J. Environ. Qual. 2022, 37, 3056. [Google Scholar] [CrossRef]
- Hutchings, J.A.; Cote, I.M.; Dodson, J.J.; Fleming, I.A.; Jennings, S.; Mantua, N.J.; Peterman, R.M.; Riddell, B.E.; Weaver, A.J. Climate change, fisheries, and aquaculture: Trends and consequences for Canadian marine biodiversity. Environ. Rev. 2012, 20, 220–311. [Google Scholar] [CrossRef]
- Hao, Z.; Tang, X.; Zhan, Y.; Tian, Y.; Yang, L.; Ding, J.; Chang, Y. A Comparative Study of Survival, Metabolism, Immune Indicators, and Proteomics, in Five Batches of Japanese Scallop Mizuhopecten yessoensis under Short-Term High Temperature Stress. Isr. J. Aquac. 2016, 68, 20828. [Google Scholar] [CrossRef]
- Peng, M.; Liu, X.; Niu, D.; Ye, B.; Lan, T.; Dong, Z.; Li, J. Survival, growth and physiology of marine bivalve (Sinonovacula constricta) in long-term low-salt culture. Sci. Rep. 2019, 9, 2819. [Google Scholar] [CrossRef]
- Liu, J.; Changg, Y.; Yang, Y.; Liu, Y.; Zhang, J.; Zhang, W.; Wang, Y. Effects of Gradual Salinity Changes on Immune Parameters of Scallop (Patinopecten yessoensis). J. Agric. Sci. Technol. 2011, 13, 7. [Google Scholar]
- LüFu; Pan, L.Q.; Wang, A.M.; Hu, Y. Effects of salinity on oxygen consumption rate and ammonia excretion rate of allogynogemetic crucian carp. Acta Hydrobiol. Sin. 2010, 34, 1000–3207. Available online: http://ssswxb.ihb.ac.cn/en/article/id/23af8a68-c7ef-4ade-9274-a28dd2aceb5f (accessed on 30 January 2025).
- Falfushynska, H.; Wu, F.; Sokolov, E.P.; Sokolova, I.M. Salinity variation modulates cellular stress response to ZnO nanoparticles in a sentinel marine bivalve, the blue mussel Mytilus sp. Mar. Environ. Res. 2023, 183, 105834. [Google Scholar] [CrossRef]
- Knowles, G.; Handlinger, J.; Jones, B.; Moltschaniwskyj, N. Hemolymph chemistry and histopathological changes in Pacific oysters (Crassostrea gigas) in response to low salinity stress. J. Invertebr. Pathol. 2014, 121, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Peteiro, L.G.; Woodin, S.A.; Wethey, D.S.; Costas-Costas, D.; Martínez-Casal, A.; Olabarria, C.; Vázquez, E. Responses to salinity stress in bivalves: Evidence of ontogenetic changes in energetic physiology on Cerastoderma edule. Sci. Rep. 2018, 8, 8329. [Google Scholar] [CrossRef] [PubMed]
- Berezina, N. Biomarkers of stress in common coastal amphipods and bivalves under salinity gradient and pollution influence (the White Sea). Žurnal Obŝej Biol. 2024, 85, 445–459. [Google Scholar] [CrossRef]
- Gostyukhina, O.L.; Kladchenko, E.S.; Chelebieva, E.S.; Tkachuk, A.A.; Lavrichenko, D.S.; Andreyeva, A.Y. Short-time salinity fluctuations are strong activators of oxidative stress in Mediterranean mussel (Mytilus galloprovincialis). Ecol. Montenegrina 2023, 63, 46–58. [Google Scholar] [CrossRef]
- Braga, A.C.; Pereira, V.; Marçal, R.; Marques, A.; Guilherme, S.; Costa, P.R.; Pacheco, M. DNA damage and oxidative stress responses of mussels Mytilus galloprovincialis to paralytic shellfish toxins under warming and acidification conditions—Elucidation on the organ-specificity. Aquat. Toxicol. 2020, 228, 105619. [Google Scholar] [CrossRef]
- Yang, H.; Wang, P.; Zhang, T.; Wang, J.; He, Y.; Zhang, F. Effects of reduced salinity on oxygen consumption and ammonia-N excretion of Chlamys farreri. Chin. J. Oceanol. Limnol. 1999, 17, 207–211. [Google Scholar] [CrossRef]
- Wei, S.; Xie, Z.; Liu, C.; Sokolova, I.; Sun, B.; Mao, Y.; Xiong, K.; Peng, J.; Fang, J.K.-H.; Hu, M. Antioxidant response of the oyster Crassostrea hongkongensis exposed to diel-cycling hypoxia under different salinities. Mar. Environ. Res. 2022, 179, 105705. [Google Scholar] [CrossRef]
- Yan, L.; Su, J.; Wang, Z.; Yan, X.; Yu, R.; Ma, P.; Li, Y.; Du, J. Transcriptomic analysis of Crassostrea sikamea × Crassostrea angulata hybrids in response to low salinity stress. PLoS ONE 2017, 12, e0171483. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.-m.; Yao, X.-m.; Cheng, Y.; Xing, Y.-j.; Sun, Y.; Hua, Q.; Wan, S.-j.; Meng, X.-j. Metformin enhances METTL14-Mediated m6A methylation to alleviate NIT-1 cells apoptosis induced by hydrogen peroxide. Heliyon 2024, 10, e24432. [Google Scholar] [CrossRef] [PubMed]
- Zucca, P.; Fernandez-Lafuente, R.; Sanjust, E. Agarose and its derivatives as supports for enzyme immobilization. Molecules 2016, 21, 1577. [Google Scholar] [CrossRef] [PubMed]
- Kenneth, J.L.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar]
- Yao, H.; Hu, Y.; Tong, H.; Shi, S. Dimethylglycine Alleviates Metabolic Dysfunction-Associated Fatty Liver Disease by Improving the Circulating Estrogen Level via Gut Staphylococcus. J. Agric. Food Chem. 2023, 72, 2708–2717. [Google Scholar] [CrossRef] [PubMed]
- Song, J.A.; Choi, C.Y. Temporal changes in physiological responses of Bay Scallop: Performance of antioxidant mechanism in Argopecten irradians in response to sudden changes in habitat salinity. Antioxidants 2021, 10, 1673. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, J.; Dong, H.; Wang, Y.; Liu, Q.; Li, H. Effect of desiccation and resubmersion on the oxidative stress response of the kuruma shrimp Marsupenaeus japonicus. Fish Shellfish Immunol. 2016, 49, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Yang, A.-G.; Yan, J.-K.; Liu, Z.-H.; Sun, X.-J.; Zhou, L.-Q.; Zhang, G.-M. Effects of acute low-salinity stress on the activities of catalase (CAT), superoxide dismutase (SOD) and glutathiones-transferase (GST) in Scapharca broughtonii. J. Mol. Biol. Res. 2019, 9, 172. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, H.; Cheng, D.; Tan, K.; Ye, T.; Ma, H.; Li, S.; Zheng, H. Differential responses of a pi-class glutathione S-transferase (CnGSTp) expression and antioxidant status between golden and brown noble scallops under pathogenic stress. Fish Shellfish Immunol. 2020, 105, 144–151. [Google Scholar] [CrossRef]
- Tang, D.; Wu, Y.; Huang, S.; Wu, L.; Luo, Y.; Wang, Z. Transcriptome reveals the mechanism of immunity in the low salinity stress of the Chinese shrimp (Fenneropenaeus chinensis). Thalass. Int. J. Mar. Sci. 2022, 38, 977–987. [Google Scholar] [CrossRef]
- Hong, Y.; Boiti, A.; Vallone, D.; Foulkes, N.S. Reactive Oxygen Species Signaling and Oxidative Stress: Transcriptional Regulation and Evolution. Antioxidants 2024, 13, 312. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jin, Y.; Xu, Z.; Zheng, Z.; Wan, S. Involvement of caspase-3 activity and survivin downregulation in cinobufocini-induced apoptosis in A 549 cells. Exp. Biol. Med. 2009, 234, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Tamm, I.; Wang, Y.; Sausville, E.; Scudiero, D.A.; Vigna, N.; Oltersdorf, T.; Reed, J.C. IAP-family protein survivin inhibits caspase activity and apoptosis induced by Fas (CD95), Bax, caspases, and anticancer drugs. Cancer Res. 1998, 58, 5315–5320. [Google Scholar] [PubMed]
- Xu, J.-H.; Wang, A.-x.; Huang, H.-Z.; Wang, J.-G.; Pan, C.-B.; Zhang, B. Survivin shRNA induces caspase-3-dependent apoptosis and enhances cisplatin sensitivity in squamous cell carcinoma of the tongue. Oncol. Res. 2010, 18, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, L.; Li, G.; Meng, K.; Guan, X.; Wang, C. Expressions of survivin and caspase-3 in human non-small cell lung cancer and their relationship with cell apoptosis. Zhongguo Fei Ai Za Zhi 2005, 8, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Cramer, A.; Hecla, J.; Wu, D.; Lai, X.; Boers, T.; Yang, K.; Moulton, T.; Kenyon, S.; Arzoumanian, Z.; Krull, W. Stationary computed tomography for space and other resource-constrained environments. Sci. Rep. 2018, 8, 14195. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, B.; Talikoti, A.T.; Nelamangala, K.; Krishnamurthy, D. Effect of Oral Pregabalin as Preemptive Analgesic in Patients Undergoing Lower Limb Orthopedic Surgeries under Spinal Anaesthesia. J. Clin. Diagn. Res. JCDR 2016, 10, Uc01–Uc04. [Google Scholar] [CrossRef]
- Ding, L.; Li, W.; Li, N.; Liang, L.; Zhang, X.; Jin, H.; Shi, H.; Storey, K.B.; Hong, M. Antioxidant responses to salinity stress in an invasive species, the red-eared slider (Trachemys scripta elegans) and involvement of a TOR-Nrf2 signaling pathway. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 219, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.J.; Lopez-Chavez, A.; Citrin, D.; Janik, J.E.; Morris, J.C. Impacting tumor cell-fate by targeting the inhibitor of apoptosis protein survivin. Mol. Cancer 2011, 10, 35. [Google Scholar] [CrossRef]
- Feng, K.; Lu, J.; Chen, Y.; Luo, Y.; Hu, Y.; Li, X.; Zhong, S.; Cheng, L. The coordinated alterations in antioxidative enzymes, PeCu/ZnSOD and PeAPX2 expression facilitated in vitro Populus euphratica resistance to salinity stress. Plant Cell Tissue Organ Cult. 2022, 150, 399–416. [Google Scholar] [CrossRef]
- Santhanam, M.; Pandey, K.; Shteinfer-Kuzmine, A.; Paul, A.; Abusiam, N.; Zalk, R.; Shoshan-Barmatz, V. Interaction of SMAC/Diablo with a survivin/BIRC5-derived peptide alters essential cancer hallmarks: Tumor growth, inflammation, and immunosuppression. Mol. Ther. 2024, 32, 1934–1955. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Q.; Zhang, X.; Gao, G.; Niu, M.; Wang, H.; Chen, L.; Wang, C.; Mu, C.; Wang, F. Metabolic Response in the Gill of Portunus trituberculatus Under Short-Term Low Salinity Stress Based on GC-MS Technique. Front. Mar. Sci. 2022, 9, 881016. [Google Scholar] [CrossRef]
- Arense, P.; Bernal, V.; Iborra, J.; Cánovas, M. Metabolic adaptation of Escherichia coli to long-term exposure to salt stress. Process Biochem. 2010, 45, 1459–1467. [Google Scholar] [CrossRef]
- Liu, T.; Nie, H.; Ding, J.; Huo, Z.; Yan, X.-W. Physiological and transcriptomic analysis provides new insights into osmoregulation mechanism of Ruditapes philippinarum under low and high salinity stress. Sci. Total Environ. 2024, 935, 173215. [Google Scholar] [CrossRef]
- Giffard-Mena, I.; Ponce-Rivas, E.; Sigala-Andrade, H.; Uganda-Solís, C.; Re, A.D.; Díaz, F.; Camacho-Jiménez, L. Evaluation of the osmoregulatory capacity and three stress biomarkers in white shrimp Penaeus vannamei exposed to different temperature and salinity conditions: Na+/K+ ATPase, Heat Shock Proteins (HSP), and Crustacean Hyperglycemic Hormones (CHHs). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2024, 271, 110942. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Liu, X. Expression of Na+/K+-ATPase Was Affected by Salinity Change in Pacific abalone Haliotis discus hannai. Front. Physiol. 2018, 9, 1244. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Shen, Y.; Li, X.; Wu, Z.; Jiao, L.; Li, J.; Zhou, Q.-C.; Jin, M. A New Insight Into the Underlying Adaptive Strategies of Euryhaline Marine Fish to Low Salinity Environment: Through Cholesterol Nutrition to Regulate Physiological Responses. Front. Nutr. 2022, 9, 855369. [Google Scholar] [CrossRef] [PubMed]
- La Fortezza, M.; Schenk, M.; Cosolo, A.; Kolybaba, A.; Grass, I.; Classen, A.-K. JAK/STAT signalling mediates cell survival in response to tissue stress. Development 2016, 143, 2907–2919. [Google Scholar] [CrossRef]
- Ding, Y.; Chen, X.; Wang, B.; Yu, B.; Ge, J. Deubiquitinase inhibitor b-AP15 activates endoplasmic reticulum (ER) stress and inhibits Wnt/Notch1 signaling pathway leading to the reduction of cell survival in hepatocellular carcinoma cells. Eur. J. Pharmacol. 2018, 825, 10. [Google Scholar] [CrossRef]
- Guan, Y.; Yao, W.; Yu, H.; Feng, Y.; Zhao, Y.; Zhan, X.; Wang, Y. Chronic stress promotes colorectal cancer progression by enhancing glycolysis through β2-AR/CREB1 signal pathway. Int. J. Biol. Sci. 2023, 19, 2006–2019. [Google Scholar] [CrossRef]
- Mambetsariev, N.; Lin, W.; Stunz, L.; Hanson, B.; Arkee, T.; Bishop, G. Nuclear TRAF3 Inhibits CREB-mediated survival and metabolic reprogramming in B lymphocytes (TUM10P.1045). J. Immunol. 2015, 194, 211-26. [Google Scholar] [CrossRef]
- Barcelos, E.S.; Rompietti, C.; Adamo, F.; Dorillo, E.; De Falco, F.; Del Papa, B.; Baldoni, S.; Nogarotto, M.; Esposito, A.; Capoccia, S.; et al. NOTCH1-mutated chronic lymphocytic leukemia displays high endoplasmic reticulum stress response with druggable potential. Front. Oncol. 2023, 13, 1218989. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Wang, C.; Yang, H.; Chen, Y.; Feng, X.; Li, B.; Fan, H. P-STAT3 Inhibition Activates Endoplasmic Reticulum Stress-Induced Splenocyte Apoptosis in Chronic Stress. Front. Physiol. 2020, 11, 680. [Google Scholar] [CrossRef] [PubMed]
- Sadoul, B.; Alfonso, S.; Cousin, X.; Prunet, P.; Bégout, M.; Leguen, I. Global assessment of the response to chronic stress in European sea bass. Aquaculture 2021, 544, 737072. [Google Scholar] [CrossRef]
- Iwamoto, S.; Kobayashi, T.; Hanamatsu, H.; Yokota, I.; Teranishi, Y.; Iwamoto, A.; Kitagawa, M.; Ashida, S.; Sakurai, A.; Matsuo, S.; et al. Tolerable glycometabolic stress boosts cancer cell resilience through altered N-glycosylation and Notch signaling activation. Cell Death Dis. 2024, 15, 53. [Google Scholar] [CrossRef]
- Habelhah, H.; Blackwell, K.; Altaeva, A.; Zhang, L.; Shi, Z. Abstract 1692: TRAF2 phosphorylation plays a critical role in cell adaptation to chronic oxidative stress. Cancer Res. 2010, 70, 1692. [Google Scholar] [CrossRef]
- Habelhah, H.; Zhang, L.; Xialikaer, A.; Blackwell, K. Abstract 3496: TRAF2 protects mammary epithelial and cancer cells from endoplasmic reticulum stress-induced apoptosis. Cancer Res. 2016, 76, 3496. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence | Gene ID |
|---|---|---|
| M.yPROTEIN3L | F: CATTGCGTACTCCCCACCTT R: GACCAATGAGCGACAGGACA | LOC110440822 |
| M.yCNDPL | F: TCTTTCCGCATTCTTTTCCTCT R: ACTTTGGATTGAGAGAGATTGAAGG | LOC110440951 |
| M.yPKD1L2 | F: TCGCTTCCTATTCTGGGGGA R: ACAAAGTCCCAAAACACGGC | LOC110441234 |
| M.yPRG4L | F: ACCCCTGAAAATGCAGCAGA R: TTCTCTTGACGGGTGAAGGC | LOC110441594 |
| M.yLIMK1L | F: CAGGACAGGGGAGTGAAACA R: GCTCCTGTTCGTCACTCTCC | LOC110441862 |
| M.yHSP70-12AL | F: GCTGGTTGGCAAATTCTCGG R: AGTGATGCATGTCCTACCGA | LOC110442164 |
| M.y2912 | F: ACCTGATACCGACAAGCATATT R: CAGGAAATGCTTTTTAACTTTGCGT | LOC110442912 |
| M.yENGASEL | F: ACCTGATACCGACAAGCATATT R: CAGGAAATGCTTTTTAACTTTGCGT | LOC110443190 |
| M.y4468 | F: TGCTTGTACATACCAGGGTCA R: ACATGGGTTGATGGAGATGACC | LOC110444468 |
| M.y4566 | F: GCAACATTAAGACCATTTTGAGGATT R: TCATTCTGACAGTAAAAGTTGGCA | LOC110444566 |
| M.yTOLLP | F: CTCCCCCGAGTTTCTCAAAG R: TCGAGACTTCCAAGGTTCCT | LOC110445648 |
| 18S | F: CTTCGAAGGCGATCAGATAC R: CTGTCAATCCTCACTGTGTC | LOC110442844 |
| Gene Name | KEGG Pathway ID | Trend |
|---|---|---|
| CASP3 | 5200, 5417, 04210, 04668 | ↓ |
| IKBKA | 5200, 5417, 0421, 04064, 04668 | ↓ |
| BIRC2-3 | 04064, 5200, 04668, 04668 | ↓ |
| LBP | 04064, 5417 | ↓ |
| MYD88 | 04064, 5417 | ↑ |
| TRAF3 | 04064, 5417, 04668 | ↑ |
| ATF4 | 04210, 5417 | ↑ |
| GADD45 | 5200, 04064, 4210 | ↑ |
| glnAGLU | 00250 | ↑ |
| gadB | 00250 | ↑ |
| ASNS | 00250 | ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, H.; Jin, X.; Wang, Z.; Ye, Q.; Li, W.; Han, L.; Ding, J. Effects of Chronic Low-Salinity Stress on Growth, Survival, Antioxidant Capacity, and Gene Expression in Mizuhopecten yessoensis. Biology 2025, 14, 759. https://doi.org/10.3390/biology14070759
Xiao H, Jin X, Wang Z, Ye Q, Li W, Han L, Ding J. Effects of Chronic Low-Salinity Stress on Growth, Survival, Antioxidant Capacity, and Gene Expression in Mizuhopecten yessoensis. Biology. 2025; 14(7):759. https://doi.org/10.3390/biology14070759
Chicago/Turabian StyleXiao, Haoran, Xin Jin, Zitong Wang, Qi Ye, Weiyan Li, Lingshu Han, and Jun Ding. 2025. "Effects of Chronic Low-Salinity Stress on Growth, Survival, Antioxidant Capacity, and Gene Expression in Mizuhopecten yessoensis" Biology 14, no. 7: 759. https://doi.org/10.3390/biology14070759
APA StyleXiao, H., Jin, X., Wang, Z., Ye, Q., Li, W., Han, L., & Ding, J. (2025). Effects of Chronic Low-Salinity Stress on Growth, Survival, Antioxidant Capacity, and Gene Expression in Mizuhopecten yessoensis. Biology, 14(7), 759. https://doi.org/10.3390/biology14070759