Evaluating Mathematical Concordance Between Taxonomic and Functional Diversity Metrics in Benthic Macroinvertebrate Communities

 ,
,  ,
,  , ,
, ,  and
and 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling Design
2.2.1. Benthic Macroinvertebrate Sampling
2.2.2. The Fluvial Habitat Index (FHI)
2.3. Evaluated Taxonomic Metrics
2.4. Using Biological Traits of Macroinvertebrates into Functional Diversity Calculations
2.5. Functional Diversity Metrics
2.6. Rationale for Metric Selection
2.7. Data Analysis
2.7.1. Clustering Sampling Points upon Their Fluvial Habitat Index Values
2.7.2. Evaluation of Mathematical Concordance Between Taxonomic and Functional Metrics
3. Results
3.1. Outcomes of Clustering Sampling Points by Fluvial Habitat Index Values
3.2. Clustering of Taxonomic Metrics as a Function of Fluvial Habitat
3.3. Clustering of Functional Diversity Metrics as a Function of Fluvial Habitat
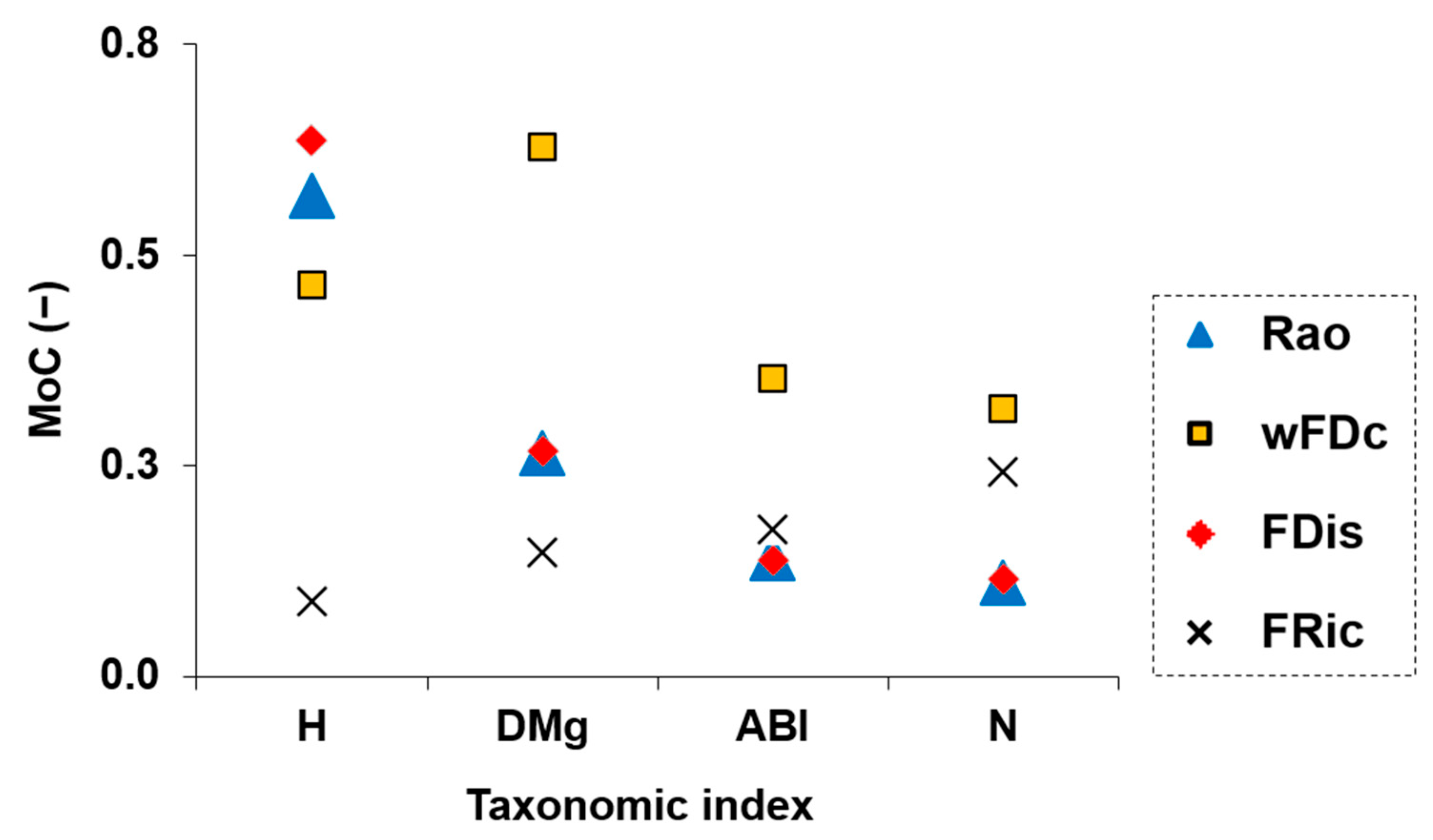
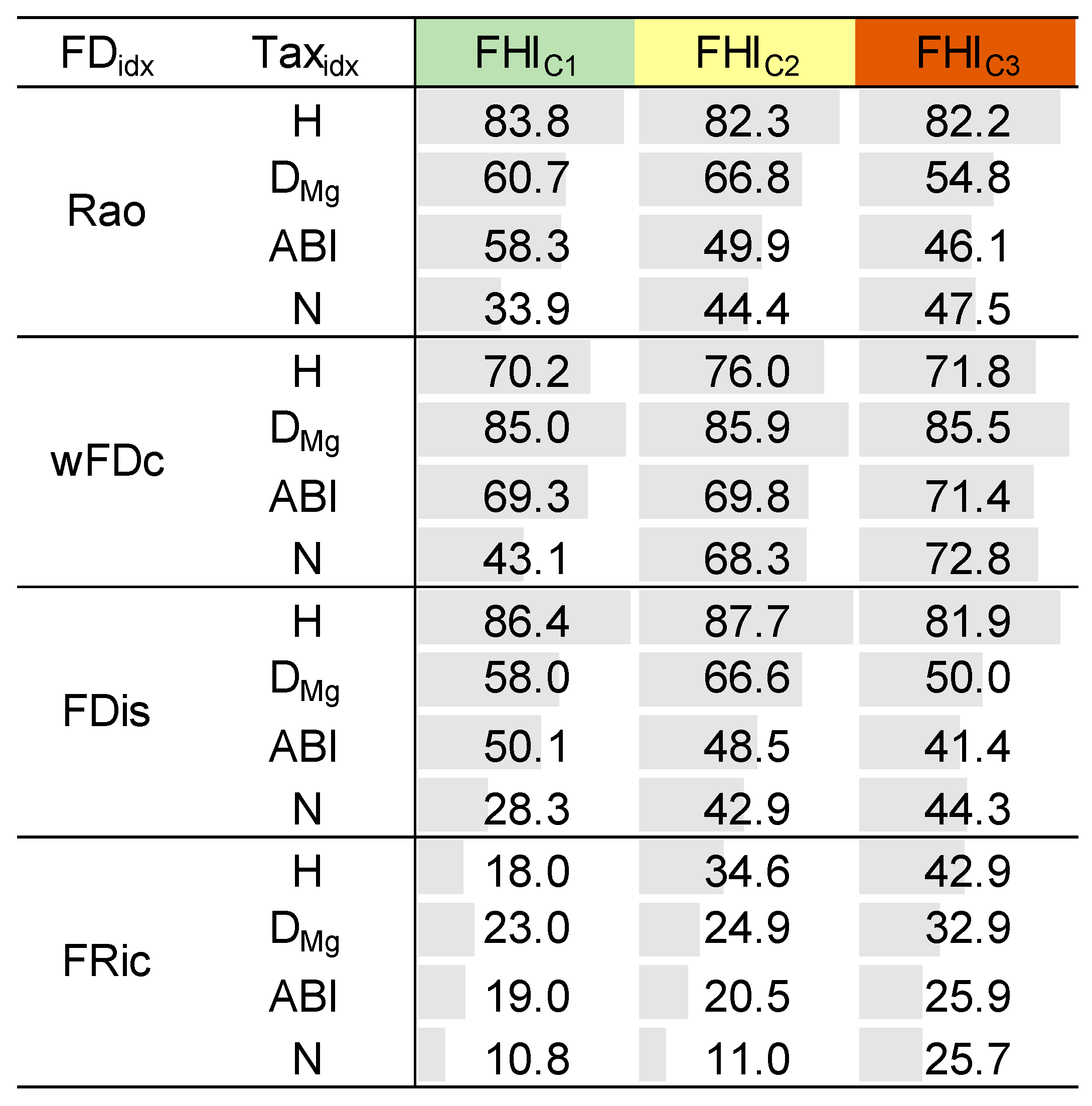
3.4. Mathematical Relationship Between Clusters from Taxonomic and Functional Metrics
3.5. Relationship Between Taxonomic and Functional Diversity Clusters as a Function of Fluvial Habitat
4. Discussion
4.1. Statistical Approach
4.2. Implications for Biomonitoring
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herman, M.R.; Nejadhashemi, A.P. A review of macroinvertebrate- and fish-based stream health indices. Ecohydrol. Hydrobiol. 2015, 15, 53–67. [Google Scholar] [CrossRef]
- Armitage, P.D.; Moss, D.; Wright, J.F.; Furse, M.T. The performance of a new biological water quality score system based on macroinvertebrates over a wide range of unpolluted running-water sites. Water Res. 1983, 17, 333–347. [Google Scholar] [CrossRef]
- Zakharova, L.; Meyer, K.M.; Seifan, M. Trait-based modelling in ecology: A review of two decades of research. Ecol. Modell. 2019, 407, 108703. [Google Scholar] [CrossRef]
- Schmera, D.; Heino, J.; Podani, J.; Erős, T.; Dolédec, S. Functional diversity: A review of methodology and current knowledge in freshwater macroinvertebrate research. Hydrobiologia 2017, 787, 27–44. [Google Scholar] [CrossRef]
- Pla, L.; Casanoves, F.; Di Rienzo, J. Quantifying Functional Biodiversity; Springer Briefs in Environmental Science; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Tomanova, S.; Moya, N.; Oberdorff, T. Using macroinvertebrate biological traits for assessing biotic integrity of neotropical streams. River Res. Appl. 2008, 24, 1230–1239. [Google Scholar] [CrossRef]
- Reynaga, M.C.; Nieto, C.; Rodríguez, J.S.; Izquierdo, A.E. Biological traits of macroinvertebrates from Puna peatbogs: Patterns along spatial environmental gradients. Freshw. Sci. 2020, 39, 137–146. [Google Scholar] [CrossRef]
- Menezes, S.; Baird, D.J.; Soares, A.M.V.M. Beyond taxonomy: A review of macroinvertebrate trait-based community descriptors as tools for freshwater biomonitoring. J. Appl. Ecol. 2010, 47, 711–719. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Carscadden, K.; Mirotchnick, N. Beyond species: Functional diversity and the maintenance of ecological processes and services. J. Appl. Ecol. 2011, 48, 1079–1087. [Google Scholar] [CrossRef]
- Sroczyńska, K.; Chainho, P.; Vieira, S.; Adão, H. What makes a better indicator? Taxonomic vs functional response of nematodes to estuarine gradient. Ecol. Indic. 2021, 121, 107113. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Z.; Castro, D.M.; Tan, X.; Jiang, X.; Meng, X.; Ge, Y.; Xie, Z. Effects of different types of land-use on taxonomic and functional diversity of benthic macroinvertebrates in a subtropical river network. Environ. Sci. Pollut. Res. 2021, 28, 44339–44353. [Google Scholar] [CrossRef]
- Laini, A.; Viaroli, P.; Bolpagni, R.; Cancellario, T.; Racchetti, E.; Guareschi, S. Taxonomic and functional responses of benthic macroinvertebrate communities to hydrological and water quality variations in a heavily regulated river. Water 2019, 11, 1478. [Google Scholar] [CrossRef]
- Lee, D.Y.; Lee, D.S.; Park, Y.S. Taxonomic and Functional Diversity of Benthic Macroinvertebrate Assemblages in Reservoirs of South Korea. Int. J. Environ. Res. Public Health 2023, 20, 673. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, R.A.; Tejerina-Garro, F.L. Relationships between taxonomic and functional components of diversity: Implications for conservation of tropical freshwater fishes. Freshw Biol. 2015, 60, 1854–1862. [Google Scholar] [CrossRef]
- Sagouis, A.; Jabot, F.; Argillier, C. Taxonomic versus functional diversity metrics: How do fish communities respond to anthropogenic stressors in reservoirs? Ecol. Freshw. Fish 2017, 26, 621–635. [Google Scholar] [CrossRef]
- da Silva Camilo, G.; de Freitas Terra, B.; Araújo, F.G. Using the relationship between taxonomic and functional diversity to assess functional redundancy in streams of an altered tropical watershed. Environ. Biol. Fish. 2018, 101, 1395–1405. [Google Scholar] [CrossRef]
- Warton, D.I.; Blanchet, F.G.; O’Hara, R.B.; Ovaskainen, O.; Taskinen, S.; Walker, S.C.; Hui, F.K. So Many Variables: Joint Modeling in Community Ecology. Trends Ecol. Evol. 2015, 30, 766–779. [Google Scholar] [CrossRef]
- Morris, A. Forestry and land-use conflicts in Cuenca, Ecuador. Mt. Res. Dev. 1985, 5, 183–196. [Google Scholar] [CrossRef]
- Sotomayor, G. A Functional and Numerical Ecology Approach to Assess the Water Quality of the Rivers in the Paute Basin (Ecuador). Ph.D. Thesis, Ghent University, Gent, Belgium, 2023. [Google Scholar]
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish, 2nd ed.; EPA 841-B-99-002; US Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999. [Google Scholar]
- Von Ellenrieder, N. Composition and structure of aquatic insect assemblages of Yungas mountain cloud forest streams in NW Argentina. Rev. Soc. Entomol. Argent 2007, 66, 57–76. [Google Scholar]
- Roldán, G. Bioindicación de la Calidad del Agua en Colombia: Propuesta Para el Uso del Método BMWP Col; Universidad de Antioquia: Medellín, Colombia, 2003. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Developments in Environmental Modelling, 20; Elsevier Science B.V.: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Hammer, Ø. PAST: Paleontological Statistics Version 4.03—Reference Manual; Natural History Museum—University of Oslo: Oslo, Norway, 2020; pp. 1–283. [Google Scholar]
- Ríos-Touma, B.; Acosta, R.; Prat, N. The Andean biotic index (ABI): Revised tolerance to pollution values for macroinvertebrate families and index performance evaluation. Rev. Biol. Trop. 2014, 62, 249–273. [Google Scholar] [CrossRef]
- Tomanova, S.; Usseglio-Polatera, P. Patterns of benthic community traits in neotropical streams: Relationship to mesoscale spatial variability. Fundam. Appl. Limnol. 2007, 170, 243–255. [Google Scholar] [CrossRef]
- Chevenet, F.; Dolédec, S.; Chessel, D. A fuzzy coding approach for the analysis of long-term ecological data. Freshw. Biol. 1994, 31, 295–309. [Google Scholar] [CrossRef]
- Sotomayor, G.; Hampel, H.; Vázquez, R.F.; Forio, M.A.E.; Goethals, P.L.M. Implications of macroinvertebrate taxonomic resolution for freshwater assessments using functional traits: The Paute River Basin (Ecuador) case. Divers. Distrib. 2021, 28, 1735–1747. [Google Scholar] [CrossRef]
- Pla, L.; Casanoves, F.; Di Rienzo, J.; Fernandez, F.; Finegan, B. Confidence intervals for functional diversity indices considering species abundance. In Proceedings of the XXIV International Biometric Conference, Dublin, Ireland, 13–18 July 2008. [Google Scholar]
- Laliberte, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.R. Diversity and dissimilarity coefficients: A unified approach. Theor. Popul. Biol. 1982, 21, 24–43. [Google Scholar] [CrossRef]
- Dalu, T.; Wasserman, R.J.; Tonkin, J.D.; Alexander, M.E.; Dalu, M.T.; Motitsoe, S.N.; Manungo, K.I.; Bepe, O.; Dube, T. Assessing drivers of benthic macroinvertebrate community structure in African highland streams: An exploration using multivariate analysis. Sci. Total Environ. 2017, 601–602, 1340–1348. [Google Scholar] [CrossRef]
- Ferraro, S.P.; Cole, F.A. Taxonomic level sufficient for assessing pollution impacts on the southern California Bight macrobenthos-revisited. Environ. Toxicol. Chem. Int. J. 1995, 14, 1031–1040. [Google Scholar]
- Corbi, J.J.; Trivinho-Strixino, S. Influence of taxonomic resolution of stream macroinvertebrate communities on the evaluation of different land uses. Acta Limnol. Bras. 2006, 18, 469–475. [Google Scholar]
- Mueller, M.; Pander, J.; Geist, J. Taxonomic sufficiency in freshwater ecosystems: Effects of taxonomic resolution, functional traits, and data transformation. Freshw. Sci. 2013, 32, 762–778. [Google Scholar] [CrossRef]
- Bo, T.; Doretto, A.; Levrino, M.; Fenoglio, S. Contribution of beta diversity in shaping stream macroinvertebrate communities among hydro-ecoregions. Aquat. Ecol. 2020, 54, 957–971. [Google Scholar] [CrossRef]
- Eriksen, T.E.; Brittain, J.E.; Søli, G.; Jacobsen, D.; Goethals, P.; Friberg, N. A global perspective on the application of riverine macroinvertebrates as biological indicators in Africa, South-Central America, Mexico and Southern Asia. Ecol. Indic. 2021, 126, 107609. [Google Scholar] [CrossRef]
- Sotomayor, G.; Hampel, H.; Vázquez, R.F.; Forio, M.A.E.; Goethals, P.L.M. Selection of an adequate functional diversity index for stream assessment based on biological traits of macroinvertebrates. Ecol. Indic. 2023, 151, 110335. [Google Scholar] [CrossRef]
- Frank, I.E.; Todeschini, R. The Data Analysis Handbook; B.V., Elsevier Science: Amsterdam, The Netherlands, 1994; Volume 14. [Google Scholar] [CrossRef]
- Dou, Q.; Du, X.; Cong, Y.; Wang, L.; Zhao, C.; Song, D.; Liu, H.; Huo, T. Influence of environmental variables on macroinvertebrate community structure in Lianhuan Lake. Ecol. Evol. 2022, 12, e8553. [Google Scholar] [CrossRef]
- Wu, J. Advances in K-Means Clustering: A Data Mining Thinking; Springer Science & Business Media: New York, NJ, USA, 2012. [Google Scholar] [CrossRef]
- Tibshirani, R.; Walther, G.; Hastie, T. Estimating the number of clusters in a data set via the gap statistic. J. R. Stat. Soc. Ser. B Stat. Methodol. 2001, 63, 411–423. [Google Scholar] [CrossRef]
- Dudoit, S.; Fridlyand, J. A prediction-based resampling method for estimating the number of clusters in a dataset. Genome Biol. 2002, 3, 1–21. [Google Scholar] [CrossRef]
- Todeschini, R.; Ballabio, D.; Termopoli, V.; Consonni, V. Extended multivariate comparison of 68 cluster validity indices. A review. Chemom. Intell. Lab. Syst. 2024, 251, 105117. [Google Scholar] [CrossRef]
- Sotomayor, G.; Hampel, H.; Vázquez, R.F. Water quality assessment with emphasis in parameter optimisation using pattern recognition methods and genetic algorithm. Water Res. 2018, 130, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Pfitzner, D.; Leibbrandt, R.; Powers, D. Characterization and evaluation of similarity measures for pairs of clusterings. Knowl. Inf. Syst. 2009, 19, 361–394. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar] [CrossRef]
- Casanoves, F.; Pla, L.; Di Rienzo, J.A.; Díaz, S. FDiversity: A software package for the integrated analysis of functional diversity. Methods Ecol. Evol. 2011, 2, 233–237. [Google Scholar] [CrossRef]
- Litchman, E.; Klausmeier, C.A. Trait-based community ecology of phytoplankton. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 615–639. [Google Scholar] [CrossRef]
- Saito, V.S.; Siqueira, T.; Fonseca-Gessner, A.A. Should phylogenetic and functional diversity metrics compose macroinvertebrate multimetric indices for stream biomonitoring? Hydrobiologia 2015, 745, 167–179. [Google Scholar] [CrossRef]
- Quenta-Herrera, E.; Crespo-Pérez, V.; Mark, B.G.; Gonzales, A.L.; Kulonen, A. Mountain freshwater ecosystems and protected areas in the tropical Andes: Insights and gaps for climate change adaptation. Environ. Conserv. 2022, 49, 17–26. [Google Scholar] [CrossRef]
- Culp, J.M.; Armanini, D.G.; Dunbar, M.J.; Orlofske, J.M.; Poff, N.L.; Pollard, A.I.; Yates, A.G.; Hose, G.C. Incorporating traits in aquatic biomonitoring to enhance causal diagnosis and prediction. Integr. Environ. Assess. Manag. 2011, 7, 187–197. [Google Scholar] [CrossRef]
- Ofogh, A.R.E.; Dorche, E.E.; Birk, S.; Fathi, P.; Shahraki, M.Z.; Bruder, A. Improving the performance of macroinvertebrate based multi-metric indices by incorporating functional traits and an index performance-driven approach. Sci. Total Environ. 2024, 931, 172850. [Google Scholar] [CrossRef]
- Morelli, F.; Benedetti, Y.; Perna, P.; Santolini, R. Associations among taxonomic diversity, functional diversity and evolutionary distinctiveness vary among environments. Ecol. Indic. 2018, 88, 8–16. [Google Scholar] [CrossRef]
- Piano, E.; Doretto, A.; Mammola, S.; Falasco, E.; Fenoglio, S.; Bona, F. Taxonomic and functional homogenisation of macroinvertebrate communities in recently intermittent Alpine watercourses. Freshw. Biol. 2020, 65, 2096–2107. [Google Scholar] [CrossRef]
- Mouillot, D.; Villéger, S.; Scherer-Lorenzen, M.; Mason, N.W.H. Functional structure of biological communities predicts ecosystem multifunctionality. PLoS ONE 2011, 6, e17476. [Google Scholar] [CrossRef]
- Matuoka, M.A.; Benchimol, M.; de Almeida-Rocha, J.M.; Morante-Filho, J.C. Effects of anthropogenic disturbances on bird functional diversity: A global meta-analysis. Ecol. Indic. 2020, 116, 106471. [Google Scholar] [CrossRef]
- van der Hoef, H.; Warrens, M.J. Understanding information theoretic measures for comparing clusterings. Behaviormetrika 2019, 46, 353–370. [Google Scholar] [CrossRef]
- Manfrin, A.; Pilotto, F.; Larsen, S.; Tonkin, J.D.; Lorenz, A.W.; Haase, P.; Stoll, S. Taxonomic and functional reorganization in Central European stream macroinvertebrate communities over 25 years. Sci. Total Environ. 2023, 889, 164278. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Romero, A.J.; Rico-Sánchez, A.E.; Sedeño-Díaz, J.E.; López-López, E. Characterization of the multidimensional functional space of the aquatic macroinvertebrate assemblages in a biosphere reserve (Central México). Diversity 2021, 13, 546. [Google Scholar] [CrossRef]
- Wang, L.; Xia, L.; Li, J.; Wan, L.; Yang, H. Winter dynamics of functional diversity and redundancy of riffle and pool macroinvertebrates after defoliation in a temperate forest stream. Front. Microbiol. 2023, 14, 1105323. [Google Scholar] [CrossRef] [PubMed]
- Casanoves, F.; Di Rienzo, J.A.; Pla, L. User Manual F-Diversity: Statistical Software for the Analysis of Functional Diversity; FDiversity: Buenos Aires, Argentina, 2010. [Google Scholar]
- Gamito, S. Caution is needed when applying Margalef diversity index. Ecol. Indic. 2010, 10, 550–551. [Google Scholar] [CrossRef]
- Saito, V.S.; Siqueira, T.; Bini, L.M.; Costa-Pereira, R.; Santos, E.P.; Pavoine, S. Comparing taxon- and trait-environment relationships in stream communities. Ecol. Indic. 2020, 117, 106625. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, L.; Li, K.; Zhang, L.; Cai, Y.; Wang, X.; Heino, J. Nutrient enrichment homogenizes taxonomic and functional diversity of benthic macroinvertebrate assemblages in shallow lakes. Limnol. Oceanogr. 2019, 64, 1047–1058. [Google Scholar] [CrossRef]
- Niu, K.; Choler, P.; de Bello, F.; Mirotchnick, N.; Du, G.; Sun, S. Fertilization decreases species diversity but increases functional diversity: A three-year experiment in a Tibetan alpine meadow. Agric. Ecosyst. Environ. 2014, 182, 106–112. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Category |
|---|---|
| Feeding habits | Collector–Filterer (C-Ft) |
| Collector–Gatherer (CG) | |
| Piercers (Pc) | |
| Predators (Pr) | |
| Scrapers (Sc) | |
| Shredders (Sh) | |
| Parasite (PA) | |
| Respiration | Tegument (Teg) |
| Gill | |
| Plastron (Pla) | |
| Spiracle (Spi) | |
| Body form | Streamlined (Str) |
| Flattened (Flat) | |
| Cylindrical (Cy) | |
| Spherical (Sph) | |
| Maximum body size (mm) | <2.5 |
| 2.5–5.0 | |
| 5–10 | |
| 10–20 | |
| 20–40 | |
| 40–80 | |
| >80 | |
| Body flexibility | None (<10°) |
| Low (10°–45°) | |
| High (>45°) | |
| Locomotion | Flier (Fli) |
| Surface swimmer (SS) | |
| Full water swimmer (FWS) | |
| Crawler (Cra) | |
| Burrower (Bur) | |
| Temporarily attached (TA) | |
| Reproduction | Asexual (As) |
| Clutches and cemented (CC) | |
| Clutches and free (CF) | |
| Clutches in vegetation (CV) | |
| Clutches and Terrestrial (CT) | |
| Isolated eggs and clutches (IEC) | |
| Isolated eggs and free (IEF) | |
| Ovoviviparity (Ovi) | |
| Hardness exoskeleton | None |
| Moderate | |
| High |
| Sampling Point | FHI | ANuR | Elevation | Slope | Strahler | |
|---|---|---|---|---|---|---|
| (−) | Cluster | (−) | (m a.s.l.) | (%) | (−) | |
| P1 | 173 ± 2.2 | FHIC1 | 19 | 2855.3 ± 500.8 | 9.3 ± 11.4 | 3 ± 1.2 |
| COL1 | 171 ± 2.5 | |||||
| SB4 | 169 ± 1.1 | |||||
| PIN5 | 154 ± 1.7 | |||||
| MAZ4 | 137 ± 1.0 | FHIC2 | 18 | 2384.1 ± 359.1 | 5.2 ± 3.7 | 4 ± 0.8 |
| PIN4 | 135 ± 1.9 | |||||
| PIN1 | 134 ± 3.2 | |||||
| MAZ1 | 130 ± 2.4 | |||||
| B8 | 107 ± 2.3 | FHIC3 | 18 | 2611.5 ± 419.0 | 11.5 ± 13.4 | 4 ± 1.7 |
| MAG2 | 105 ± 1.2 | |||||
| B2 | 96 ± 1.5 | |||||
| MAG1 | 91 ± 2.2 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sotomayor, G.; Hampel, H.; Vázquez, R.F.; Van der heyden, C.; Forio, M.A.E.; Goethals, P.L.M. Evaluating Mathematical Concordance Between Taxonomic and Functional Diversity Metrics in Benthic Macroinvertebrate Communities. Biology 2025, 14, 692. https://doi.org/10.3390/biology14060692
Sotomayor G, Hampel H, Vázquez RF, Van der heyden C, Forio MAE, Goethals PLM. Evaluating Mathematical Concordance Between Taxonomic and Functional Diversity Metrics in Benthic Macroinvertebrate Communities. Biology. 2025; 14(6):692. https://doi.org/10.3390/biology14060692
Chicago/Turabian StyleSotomayor, Gonzalo, Henrietta Hampel, Raúl F. Vázquez, Christine Van der heyden, Marie Anne Eurie Forio, and Peter L. M. Goethals. 2025. "Evaluating Mathematical Concordance Between Taxonomic and Functional Diversity Metrics in Benthic Macroinvertebrate Communities" Biology 14, no. 6: 692. https://doi.org/10.3390/biology14060692
APA StyleSotomayor, G., Hampel, H., Vázquez, R. F., Van der heyden, C., Forio, M. A. E., & Goethals, P. L. M. (2025). Evaluating Mathematical Concordance Between Taxonomic and Functional Diversity Metrics in Benthic Macroinvertebrate Communities. Biology, 14(6), 692. https://doi.org/10.3390/biology14060692