

A Preclinical Investigation of Estrogenic Bone Protection in a Hypertensive Rat Model Under Gender-Affirming Hormone Therapy
 , , , and
, , , and
Simple Summary
Abstract
1. Introduction
2. Material and Methods
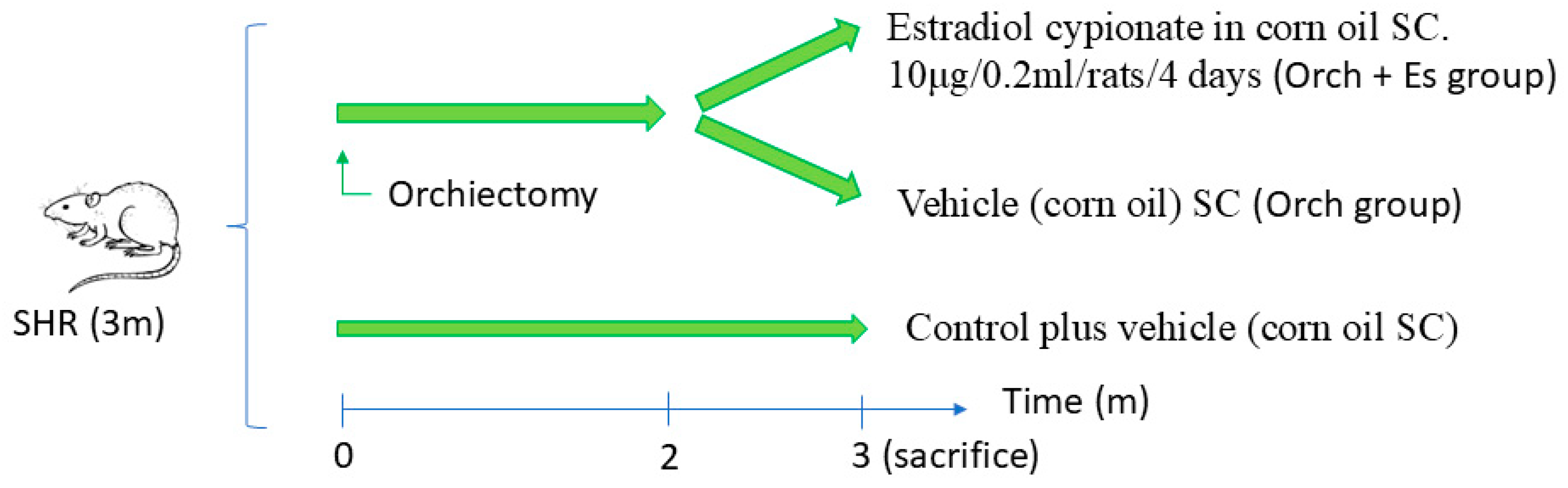
2.1. Animal Treatments
2.2. Cell Cultures
2.2.1. Bone Marrow Progenitor Cell Isolation and Incubation
2.2.2. Osteogenic Differentiation of BMPCs
2.2.3. PCR Evaluation of Osteogenic Markers
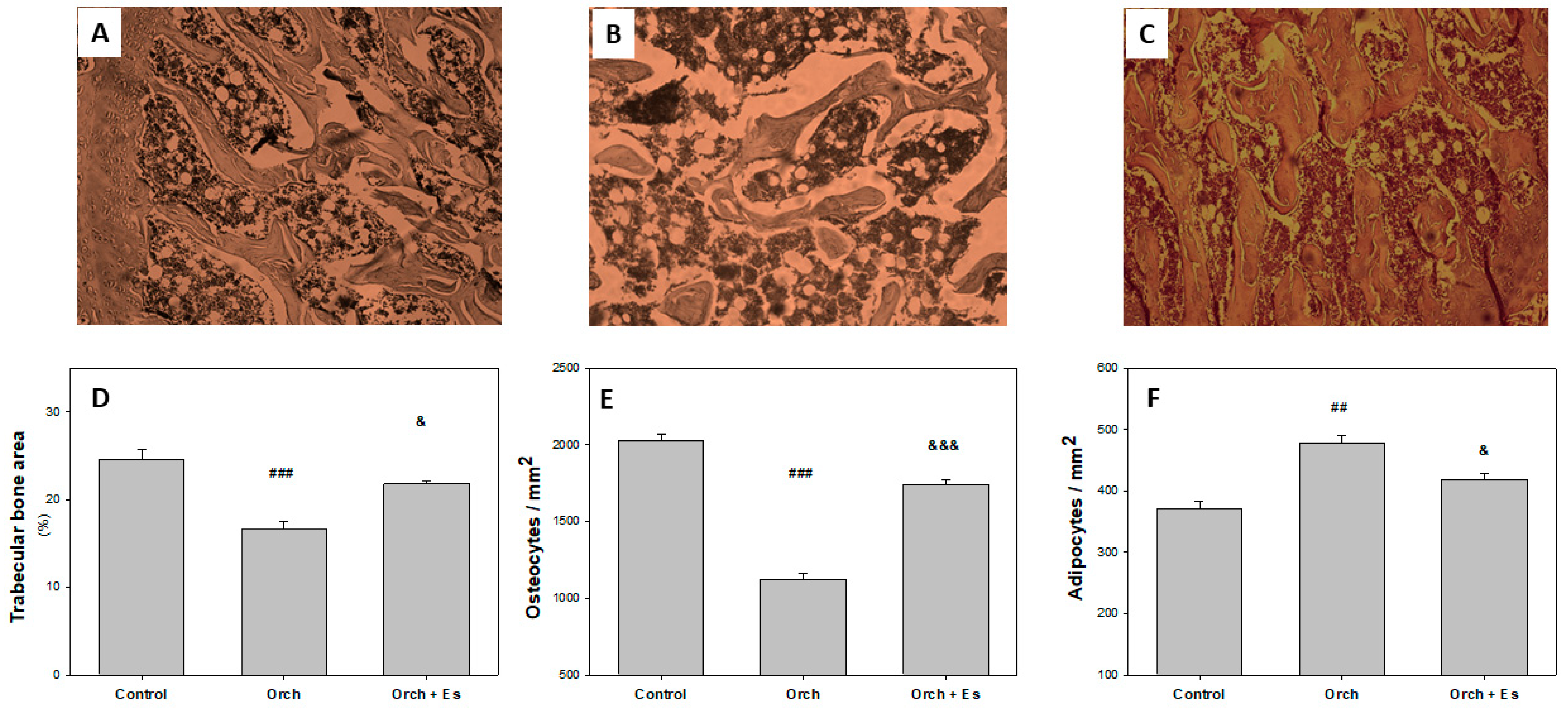
2.3. Bone Histomorphometric Examination
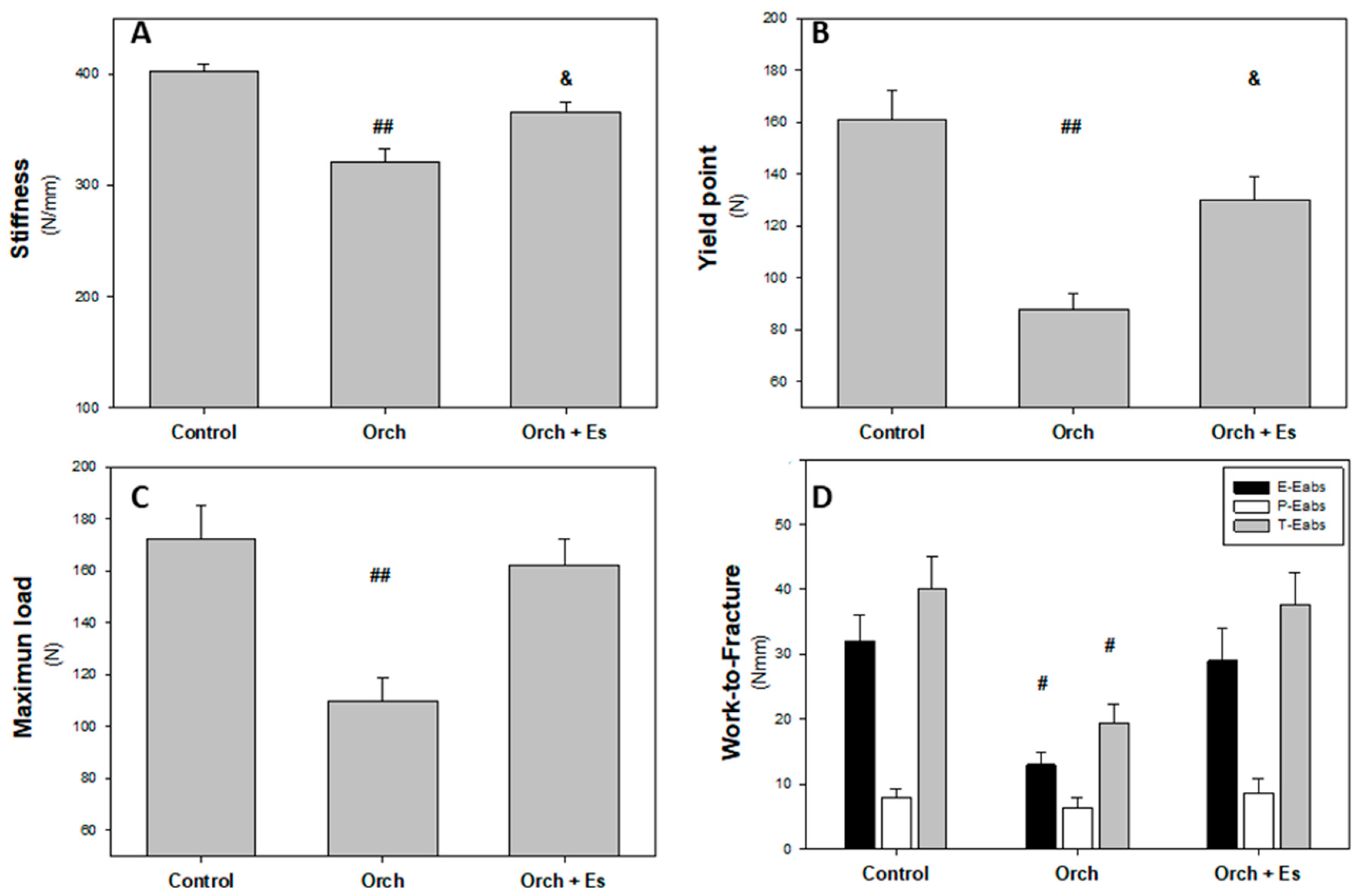
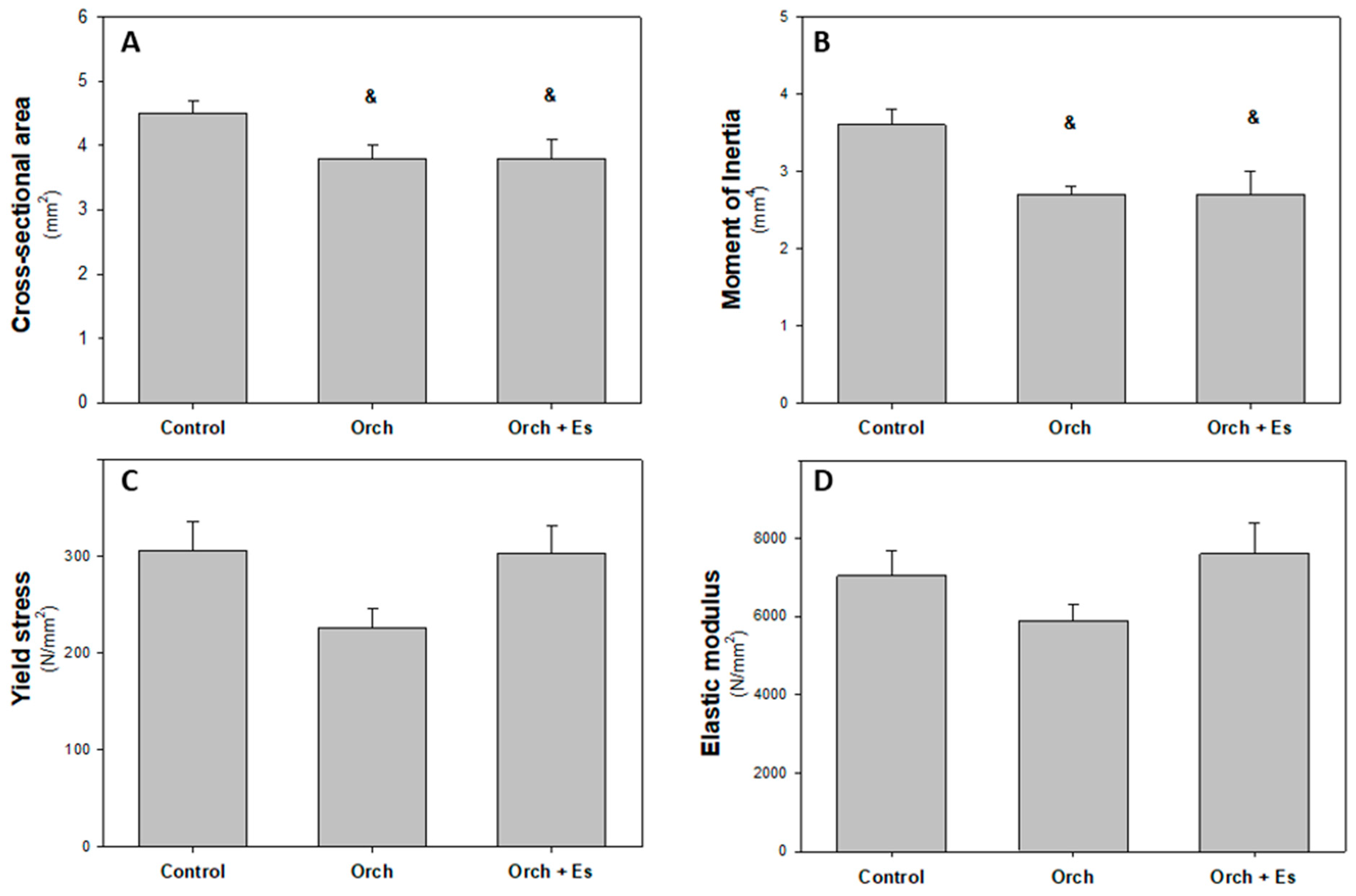
2.4. Mechanical Three-Point Bending Analysis
2.5. Statistical Analysis
3. Results
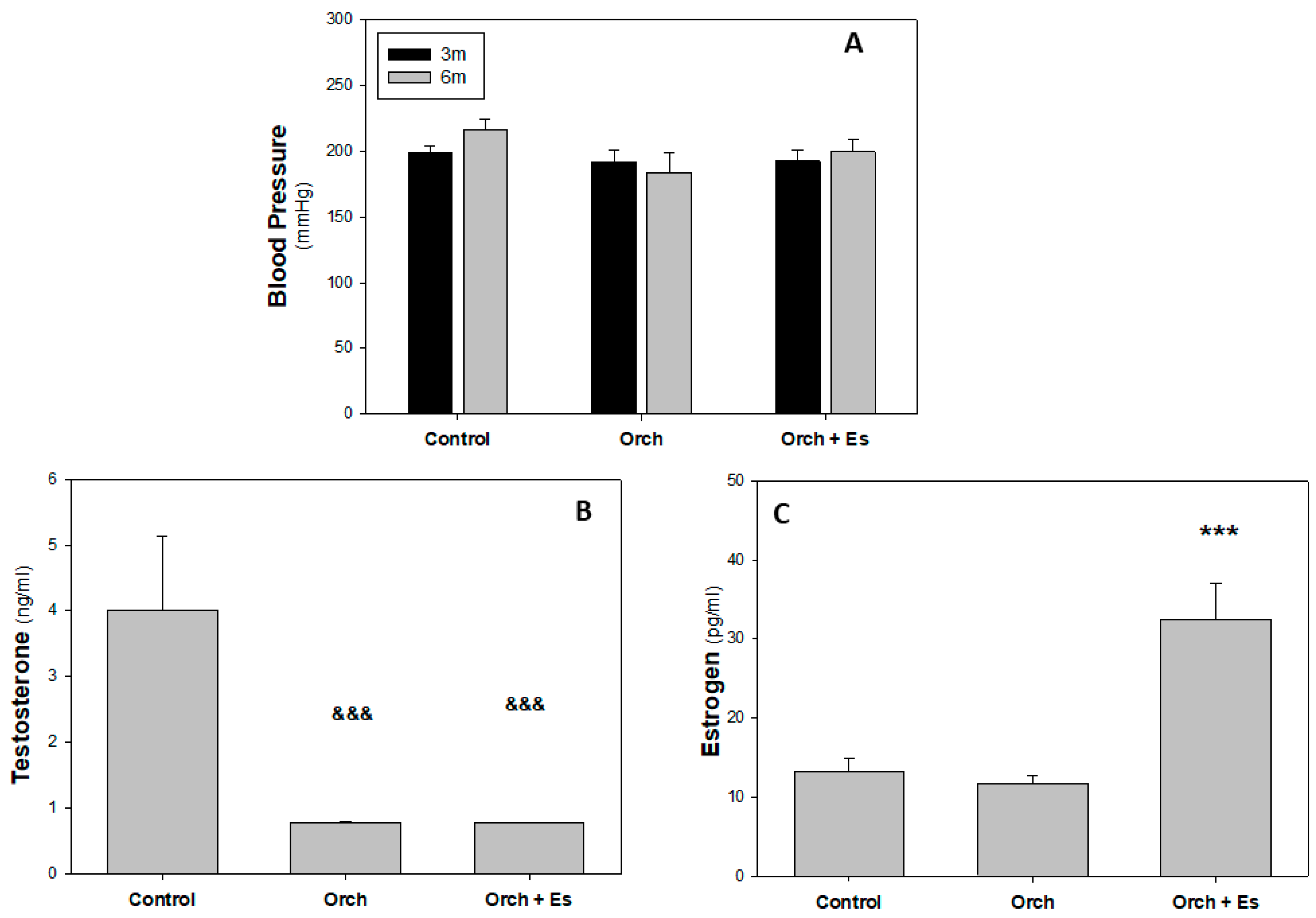
3.1. Blood Pressure and Hormonal Profile Analysis
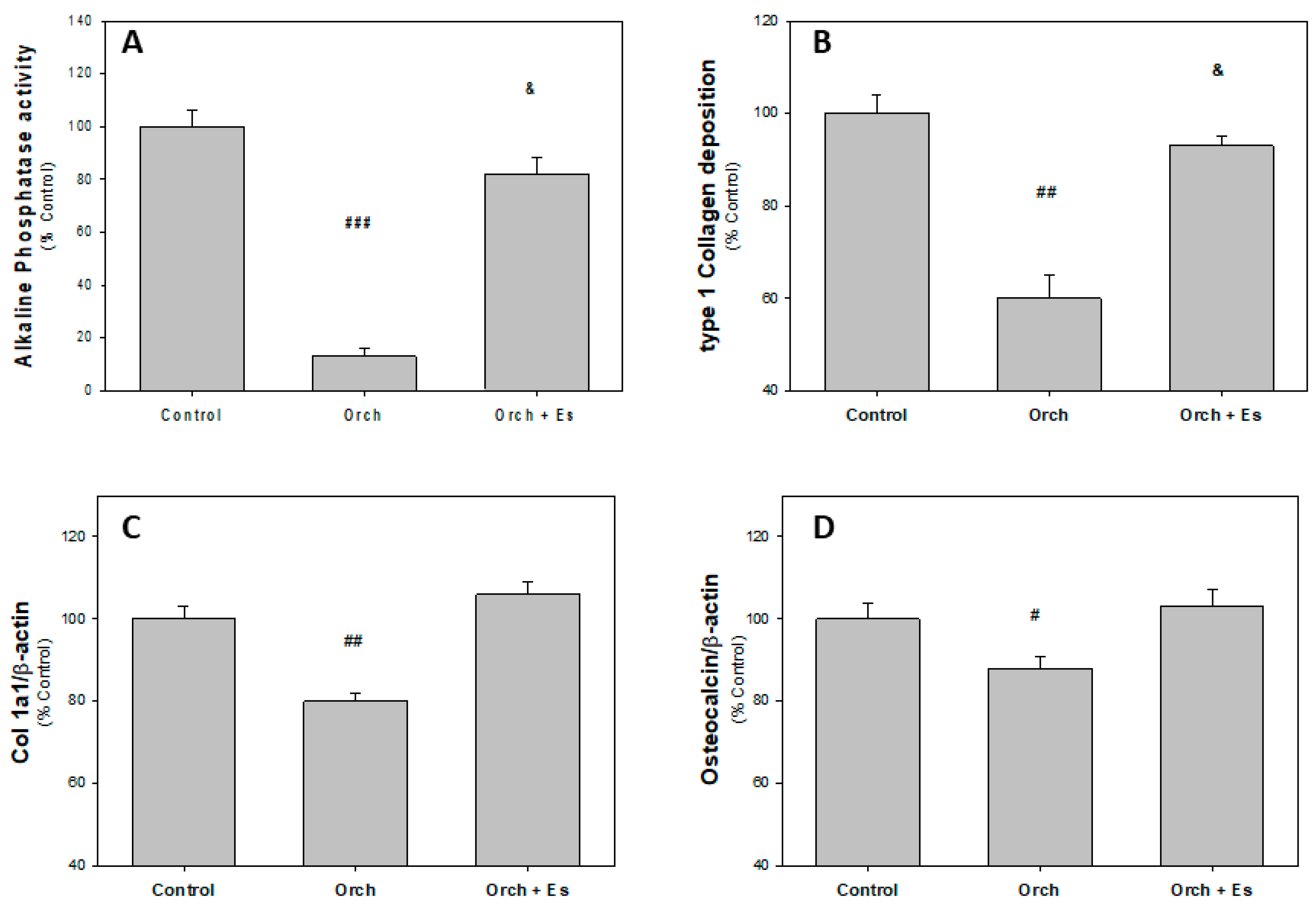
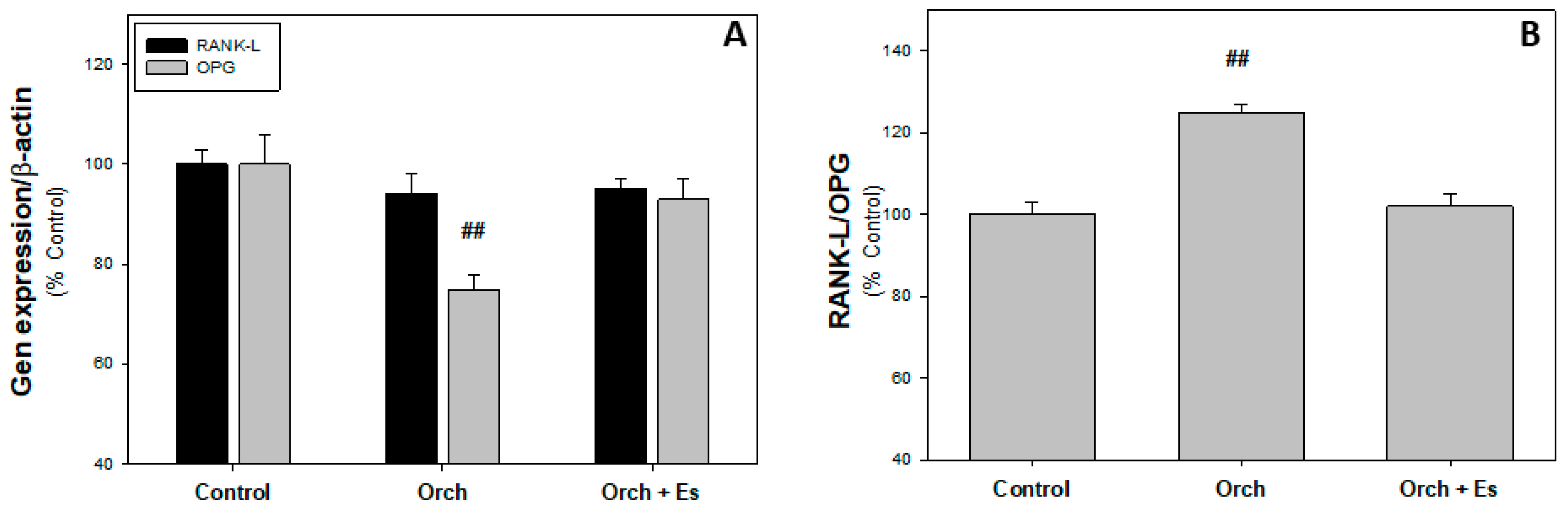
3.2. Ex Vivo Studies of BMPCs from Orchiectomized Rats Treated with Estradiol
3.3. Protective Effect of Estradiol on Bone Microarchitecture
3.4. Effect of Estradiol on the Biomechanical Behavior of the Femur
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | alkaline phosphatase activity |
| BMD | bone mineral density |
| BMPCs | bone marrow progenitor cells |
| CFU-GM | Colony-forming unit–granulocyte macrophage |
| Col 1a1 | collagen type 1 alpha 1 chain |
| Dy | yield point displacement |
| Eabs | energy absorbed |
| ERβ | estrogen receptor β |
| Es | estrogen |
| Fmax | maximun load |
| Fy | load in yield point |
| GAHT | gender-affirming hormone therapy |
| GnRH | gonadotropin-releasing hormone |
| Ix | second moment of inertia of cortical bone |
| Ob | osteoblast |
| OC | osteocalcin |
| OPG | osteoprotegerin |
| Orch | orchidectomy |
| Ot | osteocyte |
| RANK | receptor activator of nuclear factor-κβ |
| RANK-L | receptor activator of nuclear factor-κβ ligand |
| WHO | World Health Organization |
| μCT | micro-computed tomography |
References
- Wiik, A.; Andersson, D.P.; Brismar, T.B.; Chanpen, S.; Dhejne, C.; Ekström, T.J.; Flanagan, J.N.; Holmberg, M.; Kere, J.; Lilja, M.; et al. Metabolic and functional changes in transgender individuals following cross-sex hormone treatment: Design and methods of the Gender Dysphoria Treatment in Sweden (GETS) study. Contem. Clin. Trials Commun. 2018, 10, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Tassinari, R.; Maranghi, F. Rodent Model of Gender-Affirming Hormone Therapies as Specific Tool for Identifying Susceptibility and Vulnerability of Transgender People and Future Applications for Risk Assessment. Int. J. Environ. Res. Public Health 2021, 18, 12640. [Google Scholar] [CrossRef] [PubMed]
- Coleman, E.; Radix, A.E.; Bouman, W.P.; Brown, G.R.; de Vries, A.L.C.; Deutsch, M.B.; Ettner, R.; Fraser, L.; Goodman, M.; Green, J.; et al. Standards of care for the health of transgender and gender diverse people, Version 8. Int. J. Transgend. Health 2022, 23, S1–S259. [Google Scholar] [CrossRef] [PubMed]
- Streed, C.G.; Harfouch, O.; Marvel, F.; Blumenthal, R.S.; Martin, S.S.; Mukherjee, M. Cardiovascular Disease Among Transgender Adults Receiving Hormone Therapy: A Narrative Review. Ann. Intern. Med. 2017, 167, 256. [Google Scholar] [CrossRef] [PubMed]
- Nolan, B.J.; Cheung, A.S. Gender-affirming hormone therapy for transgender and gender-diverse adults in Australia. Intern. Med. J. 2024, 54, 1450–1457. [Google Scholar] [CrossRef]
- Connelly, P.J.; Marie Freel, E.; Perry, C.; Ewan, J.; Touyz, R.M.; Currie, G.; Delles, C. Gender-Affirming hormone terapy, vascular health and cardiovascular disease in transgender adults. Hypertension 2019, 74, 1266–1274. [Google Scholar] [CrossRef]
- Singh-Ospina, N.; Maraka, S.; Rodriguez-Gutierrez, R.; Davidge-Pitts, C.; Nippoldt, T.B.; Prokop, J.L.; Murad, M.H. Effect of Sex Steroids on the Bone Health of Transgender Individuals: A Systematic Review and Meta-Analysis. J. Clin. Endocrinol. Metab. 2017, 102, 3904–3913. [Google Scholar] [CrossRef]
- Elichiry, M. Guia de Tratamientos de Modificación Corporal Hormonal Para Personas Trans. 2020. Available online: https://www.ms.gba.gov.ar/sitios/generoydiversidad/wp-content/uploads/sites/216/2023/12/Guia-TMCH-1-Version-2020.pdf (accessed on 2 June 2020).
- Fighera, T.M.; Ziegelmann, P.K.; Rasia da Silva, T.; Spritzer, P.M. Bone mass effects of cross-sex hormone therapy in transgender people: Updated systematic review and meta-analysis. J. Endocr. Soc. 2019, 3, 943–964. [Google Scholar] [CrossRef]
- Vlot, M.C.; Klink, D.T.; den Heijer, M.; Blankenstein, M.A.; Rotteveel, J.; Heijboer, A.C. Effect of pubertal suppression and cross-sex hormone therapy on bone turnover markers and bone mineral apparent density (BMAD) in transgender adolescents. Bone 2017, 95, 11–19. [Google Scholar] [CrossRef]
- Streicher, C.; Heyny, A.; Andrukhova, O.; Haigl, B.; Slavic, S.; Schüler, C.; Kollmann, K.; Kantner, I.; Sexl, V.; Kleiter, M.; et al. Estrogen Regulates Bone Turnover by Targeting RANKL Expression in Bone Lining Cells. Sci. Rep. 2017, 7, 6460. [Google Scholar] [CrossRef]
- Banoriya, G.K.; Singh, V.K.; Maurya, R.; Kharwar, R.K. Neuro-Immuno-Endocrine Regulation of Bone Homeostasis. Discov. Med. 2025, 37, 464–485. [Google Scholar] [CrossRef] [PubMed]
- Khalid, A.B.; Krum, S.A. Estrogen receptors alpha and beta in bone. Bone 2016, 87, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Emmanuelle, N.E.; Marie-Cécile, V.; Florence, T.; Jean-François, A.; Françoise, L.; Coralie, F.; Alexia, V. Critical Role of Estrogens on Bone Homeostasis in Both Male and Female: From Physiology to Medical Implications. Int. J. Mol. Sci. 2021, 22, 1568. [Google Scholar] [PubMed]
- Colvard, D.S.; Eriksen, E.F.; Keeting, P.E.; Wilson, E.M.; Lubahn, D.B.; French, F.S.; Riggs, B.L.; Spelsberg, T.C. Identification of androgen receptors in normal human osteoblast-like cells. Proc. Natl. Acad. Sci. USA 1989, 86, 854–857. [Google Scholar] [CrossRef]
- Giacomelli, G.; Meriggiola, M.C. Bone health in transgender people: A narrative review. Ther. Adv. Endocrinol. 2022, 13, 204201882210993. [Google Scholar] [CrossRef]
- Goodrich, E.; Walcott, Q.; Dallman, J.; Crow, H.; Templeton, K. Bone health in the transgender population. JBJS Rev. 2023, 11, e23.00123. [Google Scholar] [CrossRef]
- Bishop, A.; Overcash, F.; McGuire, J.; Reicks, M. Diet and Physical Activity Behaviors Among Adolescent Transgender Students: School Survey Results. J. Adolesc. Health 2020, 66, 484–490. [Google Scholar] [CrossRef]
- Lee, J.L.; Finlayson, C.; Olson-Kennedy, J.; Garofalo, R.; Chan, Y.M.; Glidden, D.V.; Rosenthal, S.M. Low bone mineral density in early pubertal transgender/gender diverse youth: Findings from the trans youth care study. J. Endocr. Soc. 2020, 4, bvaa065. [Google Scholar] [CrossRef]
- Lee, J.Y. Bone health in the transgender and gender diverse youth population. Curr. Osteoporos. Rep. 2023, 21, 459–471. [Google Scholar] [CrossRef]
- Lightner, J.S.; Schneider, J.; Grimes, A.; Wigginton, M.; Curran, L.; Gleason, T.; Prochnow, T. Physical activity among transgender individuals: A systematic review of quantitative and qualitative studies. PLoS ONE 2024, 19, e0297571. [Google Scholar] [CrossRef]
- Motta, G.; Marinelli, L.; Barale, M.; Brustio, P.R.; Manieri, C.; Ghigo, E.; Procopio, M. Lanfranco F, Fracture risk assessment in an Italian group of transgender women after gender-confirming surgery. J. Bone Miner. Metab. 2020, 38, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Van Caenegem, E.; Taes, Y.; Wierckx, K.; Vandewalle, S.; Toye, K.; Kaufman, J.M.; Schreiner, T.; Haraldsen, I.; T’Sjoen, G. Low bone mass is prevalent in male-to-female transsexual persons before the start of crosssex hormonal therapy and gonadectomy. Bone 2013, 54, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Yang, K.; Hu, Z.; Li, M.; Wei, H.; Tang, Z.; Chen, B.; Su, C.; Cai, D.; Xu, J. Determining the association between hypertension and bone metabolism markers in osteoporotic patients. Medicine 2021, 100, e26276. [Google Scholar] [CrossRef]
- World Health Organization. Global Report on Hypertension: The Care Against a Silent Killer. 2023. Available online: https://www.who.int/publications/i/item/9789240081062 (accessed on 19 September 2023).
- Hembree, W.C.; Cohen-Kettenis, P.T.; Gooren, L.; Hannema, S.E.; Meyer, W.J.; Murad, M.H.; Rosenthal, S.M.; Safer, J.D.; Tangpricha, V.; T’Sjoen, G.G. Endocrine treatment of gender-dysphoric/gender-incongruent persons: An endocrine society clinical practice. J. Clin. Endocrinol. Meta 2017, 102, 1–35. [Google Scholar] [CrossRef]
- Ciancia, S.; Dubois, V.; Cools, M. Impact of gender-affirming treatment on bone health in transgender and gender diverse youth. Endocr. Connect. 2022, 11, e220280. [Google Scholar] [CrossRef] [PubMed]
- Boogers, L.; Reijtenbagh, S.; Wiepjes, C.; van Trotsenburg, A.; den Heijer, M.; Hannema, S. Time Course of Body Composition Changes in Transgender Adolescents During Puberty Suppression and Sex Hormone Treatment. J. Clin. Endocrinol. Metab. 2024, 109, e1593–e1601. [Google Scholar] [CrossRef]
- Nagelberg, A.; Abdala, R.; Larroude, M.; Farias, V.; Garabito, M.; Balogna, M.; Giacoia, E.; Karlsbrum, S.; Pavlove, M.; Frigeri, A.; et al. Valoración de la Densidad Mineral Ósea en Personas Transgénero. Medicina 2024, 84, 125–137. [Google Scholar]
- Glintborg, D.; T’Sjoen, G.; Ravn, P.; Andersen, M.S. Management of Endocrine Disease: Optimal feminizing hormone treatment in transgender people. Eur. J. Endocrinol. 2021, 185, R49–R63. [Google Scholar] [CrossRef]
- Haraldsen, I.R.; Haug, E.; Falch, J.; Egeland, T.; Opjordsmoen, S. Cross-sex pattern of bone mineral density in early onset gender identity disorder. Horm. Behav. 2007, 52, 334–343. [Google Scholar] [CrossRef]
- Wiepjes, C.M.; Den Heijer, M.; T’Sjoen, G.G. Bone health in adult trans persons: An update of the literature. Curr. Opin. Endocrinol. Diabetes Obes. 2019, 26, 296–300. [Google Scholar] [CrossRef]
- Gomes, N.A.; Guarenhi, G.G.; Valenga, H.M.; von Stein Cubas Warnavin, S.; Paganella Chaves, J.D.; Cardoso, A.C.; Steffens, J.P. Mandibular-related bone metabolism in orchiectomized rats treated with sex hormones. Arch. Oral. Biol. 2021, 122, 105000. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, P.; Rau, A.; Seidlová-Wuttke, D.; Jarry, H.; Schminke, B.; Matthes, S.; Wiese, K.G. Effect of dihydrotestosterone, 17-β-estrogen, genistein and equol on remodeling and morphology of bone in osteoporotic male rats during bone healing. Bone Rep. 2020, 13, 100300. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.Z.; Qi, H.; Crawfort, D.T.; Chidsey-Frink, K.L.; Simmons, H.A.; Thompson, D.D. Lasofoxifene (CP-336,156), a selective estrogen receptor modulator, prevents bone loss induced by aging and orchidectomy in the adult rat. Endocrinology 2000, 141, 1338–1344. [Google Scholar] [CrossRef]
- Komrakova, M.; Büchler, G.; Böker, K.O.; Lehmann, W.; Schilling, A.F.; Roch, P.J.; Taudien, S.; Hoffmann, D.B.; Sehmish, S. A combined treatment with selective androgen and estrogen receptor modulators prevents bone loss in orchiectomized rats. J. Endocrinol. Invest. 2022, 45, 2299–2311. [Google Scholar] [CrossRef] [PubMed]
- Reis, L.O.; Zani, E.L.; Garcia-Perdomo, H.A. Estrogen therapy in patients with prostate cancer: A contemporary systematic review. Int. Urol. Nephrol. 2018, 50, 993–1003. [Google Scholar] [CrossRef]
- Tassinari, R.; Tammaro, A.; Lori, G.; Tait, S.; Martinelli, A.; Cancemi, L.; Frassanito, P.; Maranghi, F. Risk Assessment of Transgender People: Development of Rodent Models Mimicing Gender-Affirming Hormone Therapies and Identification of Sex-Dimorphic Liver Genes as Novel Biomarkers of Sex Transition. Cells 2023, 12, 474. [Google Scholar] [CrossRef]
- Szawka, R.E.; Rodovalho, G.V.; Monteiro, P.M.; Carrer, H.F.; Anselmo-Franci, J.A. Ovarian-steroid modulation of locus coeruleus activity in female rats: Involvement in luteinising hormone regulation. J. Neuroendocrinol. 2009, 21, 629–639. [Google Scholar] [CrossRef]
- Escudero, D.S.; Brea, M.S.; Caldiz, C.I.; Amarillo, M.E.; Aranda, J.O.; Portiansky, E.L.; Pérez, N.G.; Díaz, R.G. PDE5 inhibition improves cardiac morphology and function in SHR by reducing NHE1 activity: Repurposing Sildenafil for the treatment of hypertensive cardiac hypertrophy. Eur. J. Pharmacol. 2021, 891, 173724. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.L.; Wanionok, N.E.; McCarthy, A.D.; Morel, G.R.; Fernández, J.M. Systemic oxidative stress in old rats is associated with both osteoporosis and cognitive impairment. Exp. Gerontol. 2021, 156, 111596. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Álvarez-Lloret, P.; Fernández, J.M.; Molinuevo, M.S.; Lino, A.B.; Ferretti, J.L.; Capozza, R.F.; Cortizo, A.M.; McCarthy, A.D. Multi-Scale Approach for the Evaluation of Bone Mineralization in Strontium Ranelate-Treated Diabetic Rats. Biol. Trace Elem. Res. 2018, 186, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Jepsen, K.J.; Silva, M.J.; Vashishth, D.; Guo, X.E.; Van Der Meulen, M.C. Establishing Biomechanical Mechanisms in Mouse Models: Practical Guidelines for Systematically Evaluating Phenotypic Changes in the Diaphyses of Long Bones. J. Bone Miner. Res. 2015, 30, 951–966. [Google Scholar] [CrossRef]
- Wanionok, N.E.; Molinuevo, M.S.; Fernández, J.M.; Besadas, L.; Cortizo, A.M.; Castillo, E.J.; Jiron, J.M.; Sedlinsky, C.; Shurman, L.; Aguirre, J.I.; et al. Skeletal Effects of a Prolonged Oral Metformin Treatment in Adult Wistar Rats. Exp. Clin. Endocrinol. Diabetes 2024, 132, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Boyer, P.; Compagnucci, G.E.; Olivera, M.I.; Bozzini, C.; Roig, M.C.; Campagnucci, C.V.; Alippi, R.M. Bone status in an animal model of chronic sub-optimal nutrition: A morphometric, densitometric and mechanical study. Br. J. Nutr. 2005, 93, 663–669. [Google Scholar] [CrossRef]
- Bozzini, C.; Picasso, E.O.; Champin, G.M.; Alippi, R.M.; Bozzini, C.E. Biomechanical properties of the mid-shaft femur in middle-aged hypophysectomized rats as assessed by bending test. Endocrine 2012, 42, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Tiyasatkulkovit, W.; Promruk, W.; Rojviriya, C.; Pakawanit, P.; Chaimongkolnukul, K.; Kengkoom, K.; Teerapornpuntakit, J.; Panupinthu, N.; Charoenphandhu, N. Impairment of bone microstructure and upregulation of osteoclastogenic markers in spontaneously hypertensive rats. Sci. Rep. 2019, 9, 12293. [Google Scholar] [CrossRef]
- Ye, Z.; Lu, H.; Liu, P. Association between essential hypertension and bone mineral density: A systematic review and meta-analysis. Oncotarget 2017, 8, 68916–68927. [Google Scholar] [CrossRef]
- Kapsdorferová, V.; Grešová, S.; Švorc, P. Measurement of blood pressure in rats: Invasive or noninvasive methods? Physiol. Rep. 2024, 12, e70041. [Google Scholar] [CrossRef]
- Venkatesh, V.S.; Nie, T.; Golub, S.; Stok, K.S.; Hemmatian, H.; Desai, R.; Handelsman, D.J.; Zajac, J.D.; Grossmann, M.; Davey, R.A. High circulating concentrations of estradiol are anabolic for bone mass and strength in an adult male to female transgender mouse model. Bone 2024, 186, 117143. [Google Scholar] [CrossRef]
- Abdala, R.; Nagelberg, A.; Brance, M.L. Salud ósea en personas transgénero. Actualizaciones en Osteología 2020, 16, 176–186. [Google Scholar]
- Bonanno, M.S.; Saravia, M.R.; Seijo, M.; Zeni, S.N. Rol de la Osteocalcina mas alla del hueso. Actualizaciones en Osteología 2019, 15, 78–93. [Google Scholar]
- Liu, S.; Pang, Q.; Guan, W.; Yu, F.; Wang, O.; Li, M.; Xing, X.; Yu, W.; Jiang, Y.; Xia, W. Association of serum osteocalcin with bone microarchitecture and muscle mass in Beijing community-dwelling postmenopausal women. Endocrine 2024, 84, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Van Caenegem, E.; Wierckx, K.; Taes, Y.; Schreiner, T.; Vandewalle, S.; Toye, K.; Kaufman, J.M.; T’Sjoen, G. Preservation of volumetric bone density and geometry in trans women during cross-sex hormonal therapy: A prospective observational study. Osteoporos. Int. 2015, 26, 35–47. [Google Scholar] [CrossRef]
- Delgado-Ruiz, R.; Swanson, P.; Romanos, G. Systematic Review of the Long-Term Effects of Transgender Hormone Therapy on Bone Markers and Bone Mineral Density and Their Potential Effects in Implant Therapy. J. Clin. Med. 2019, 8, 784. [Google Scholar] [CrossRef]
- Gava, G.; Cerpolini, S.; Martelli, V.; Battista, G.; Seracchioli, R.; Meriggiola, M.C. Cyproterone acetate vs leuprolide acetate in combination with transdermal oestradiol in transwomen: A comparison of safety and effectiveness. Clin. Endocrinol. 2016, 85, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Vlot, M.C.; Wiepjes, C.M.; De Jongh, R.T.; T’Sjoen, G.; Heijboer, A.C.; Den Heijer, M. Gender-Affirming Hormone Treatment Decreases Bone Turnover in Transwomen and Older Transmen. J. Bone Miner. Res. 2019, 34, 1862–1872. [Google Scholar] [CrossRef]
- Wanionok, N.E.; Morel, G.R.; Fernández, J.M. Osteoporosis and Alzheimer’s disease (or Alzheimer’s disease and Osteoporosis). Ageing Res. Rev. 2024, 99, 102408. [Google Scholar] [CrossRef]
- Chi, G.; Qiu, L.; Ma, J.; Wu, W.; Zhang, Y. The association of osteoprotegerin and RANKL with osteoporosis: A systematic review with meta-analysis. J. Orthop. Surg. Res. 2023, 18, 839. [Google Scholar] [CrossRef]
- Chen, Q.; Kaji, H.; Kanatani, M.; Sugimoto, T.; Chihara, K. Testosterone increases Osteoprotegerin mRNA expression in mouse osteoblast cells. Horm. Metab. Res. 2004, 36, 674–678. [Google Scholar] [CrossRef]
- Bord, S.; Ireland, D.C.; Beavan, S.R.; Compston, J.E. The effects of estrogen on osteoprotegerin, RANKL, and estrogen receptor expression in human osteoblasts. Bone 2003, 32, 136–141. [Google Scholar] [CrossRef]
- Stratos, I.; Rinas, I.; Schröpfer, K.; Hink, K.; Herlyn, P.; Bäumler, M.; Histing, T.; Bruhn, S.; Müller-Hilke, B.; Menger, M.D.; et al. Effects on Bone and Muscle upon Treadmill Interval Training in Hypogonadal Male Rats. Biomedicines 2023, 11, 1370. [Google Scholar] [CrossRef] [PubMed]
- Dobrolińska, M.; Van Der Tuuk, K.; Vink, P.; Van Den Berg, M.; Schuringa, A.; Monroy-Gonzalez, A.G.; García, D.V.; Schultz, W.C.M.W.; Slart, R.H.J.A. Bone mineral density in transgender individuals after gonadectomy and long-term gender-affirming hormonal treatment. J. Sex. Med. 2019, 16, 1469–1477. [Google Scholar] [CrossRef] [PubMed]
- Snyder, P.J. Andrógenos y tracto reproductor masculino. In Goodman & Gilman: Las Bases Farmacológicas de la Terapéutica, 13th ed.; Brunton, L.L., Chabner, B.A., Knollmann, B.C., Eds.; McGraw-HillEducation: New York, NY, USA, 2019; pp. 833–843. [Google Scholar]
- Doolittle, M.L.; Saul, D.; Kaur, J.; Rowsey, J.L.; Eckhardt, B.; Vos, S.; Grain, S.; Kroupova, K.; Ruan, M.; Weivoda, M.; et al. Skeletal effects of inducible ERα deletion in osteocytes in adult mice. J. Bone Miner. Res. 2022, 37, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Nyman, J.S. Effect of Diabetes on the Fracture Resistance of Bone. Clin. Rev. Bone Miner. Metab. 2013, 11, 38–48. [Google Scholar] [CrossRef]
- Prakasam, G.; Yeh, J.K.; Chen, M.M.; Castro-Magana, M.; Liang, C.T.; Aloia, J.F. Effects of growth hormone and testosterone on cortical bone formation and bone density in aged orchiectomized rats. Bone 1999, 24, 491–497. [Google Scholar] [CrossRef]
- Rothman, M.S.; Iwamoto, S.J. Bone Health in the Transgender Population. Clin. Rev. Bone Miner. Metab. 2019, 17, 77–85. [Google Scholar] [CrossRef]
- Deckard, C.; Walker, A.; Hill, B.J.F. Using three-point bending to evaluate tibia bone strength in ovariectomized young mice. J. Biol. Phys. 2017, 43, 139–148. [Google Scholar] [CrossRef]
- Azboy, I.; Özkaya, M.; Demir, T.; Demirtaş, A.; Kağan Arslan, A.; Özkul, E.; Akcan, A.; Tolunay, T. Biomechanical properties of osteoporotic rat femurs after different hormonal treatments: Genistein, estradiol, and estradiol/progesterone. SICOT-J. 2016, 2, 24. [Google Scholar] [CrossRef]
- Rooney, A.M.; van der Meulen, M.C.H. Mouse models to evaluate the role of estrogen receptor α in skeletal maintenance and adaptation. Ann. N. Y. Acad. Sci. 2017, 1410, 85–92. [Google Scholar] [CrossRef]
- Xu, X.; Yang, H.; Bullock, W.A.; Gallant, M.A.; Ohlsson, C.; Bellido, T.M.; Main, R.P. Osteocyte Estrogen Receptor β (Ot-ERβ) Regulates Bone Turnover and Skeletal Adaptive Response to Mechanical Loading Differently in Male and Female Growing and Adult Mice. J. Bone Miner. Res. 2022, 38, 186–197. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Genebank Code | Product Size (bp) | Sequence | |
|---|---|---|---|---|
| Housekeeping gene | ||||
| β-actin | NM_031144.3 | 345 | Fw | CCTTCAACACCCCAGCCAT |
| Rv | CATAGCTCTTCTCCAGGGA | |||
| Markers evaluated for osteogenic differentiation | ||||
| Col1a1 | NM_053304.1 | 651 | Fw | GCATACACAATGGCCTAA |
| Rv | CTGTTCCAGGCAATCCAC | |||
| Osteocalcin | NM_13414.1 | 156 | Fw | CAAAGCCTTCATGTCCAA |
| Rv | TCTGATAGCTCGTCACAA | |||
| RANK-L | NM_057149.1 | 432 | Fw | TCGCTCTGTTCCTGTACTTT |
| Rv | CCCTTAGTTTTCCGTTGCTT | |||
| OPG | U94330.1 | 408 | Fw | CTCCTGGCACCTACCTAA |
| Rv | GTGTTGCATTTCCTTTCTGA | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Streckwall, L.; Colareda, G.A.; Escudero, D.; Diaz, R.G.; Fernández, J.M. A Preclinical Investigation of Estrogenic Bone Protection in a Hypertensive Rat Model Under Gender-Affirming Hormone Therapy. Biology 2025, 14, 650. https://doi.org/10.3390/biology14060650
Streckwall L, Colareda GA, Escudero D, Diaz RG, Fernández JM. A Preclinical Investigation of Estrogenic Bone Protection in a Hypertensive Rat Model Under Gender-Affirming Hormone Therapy. Biology. 2025; 14(6):650. https://doi.org/10.3390/biology14060650
Chicago/Turabian StyleStreckwall, Lucas, Germán A. Colareda, Daiana Escudero, Romina G. Diaz, and Juan M. Fernández. 2025. "A Preclinical Investigation of Estrogenic Bone Protection in a Hypertensive Rat Model Under Gender-Affirming Hormone Therapy" Biology 14, no. 6: 650. https://doi.org/10.3390/biology14060650
APA StyleStreckwall, L., Colareda, G. A., Escudero, D., Diaz, R. G., & Fernández, J. M. (2025). A Preclinical Investigation of Estrogenic Bone Protection in a Hypertensive Rat Model Under Gender-Affirming Hormone Therapy. Biology, 14(6), 650. https://doi.org/10.3390/biology14060650