
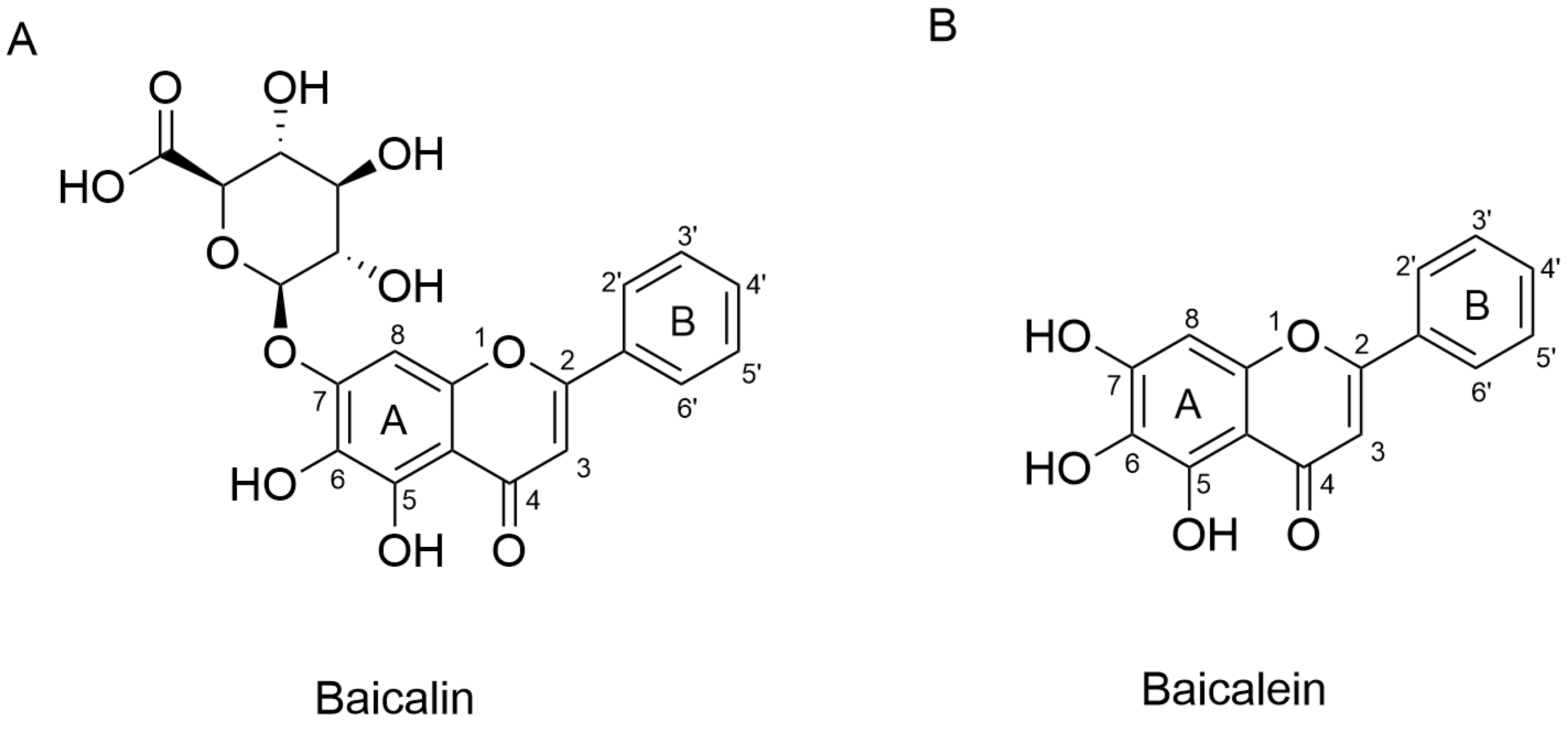
Recent Advances in the Therapeutic Effects and Molecular Mechanisms of Baicalin
Simple Summary
Abstract
1. Introduction
2. Antitumor Effects
3. Cardiovascular Protection Effects
3.1. Protection of the Heart
3.2. Regulation of Blood Vessels
4. Neurological Disorders
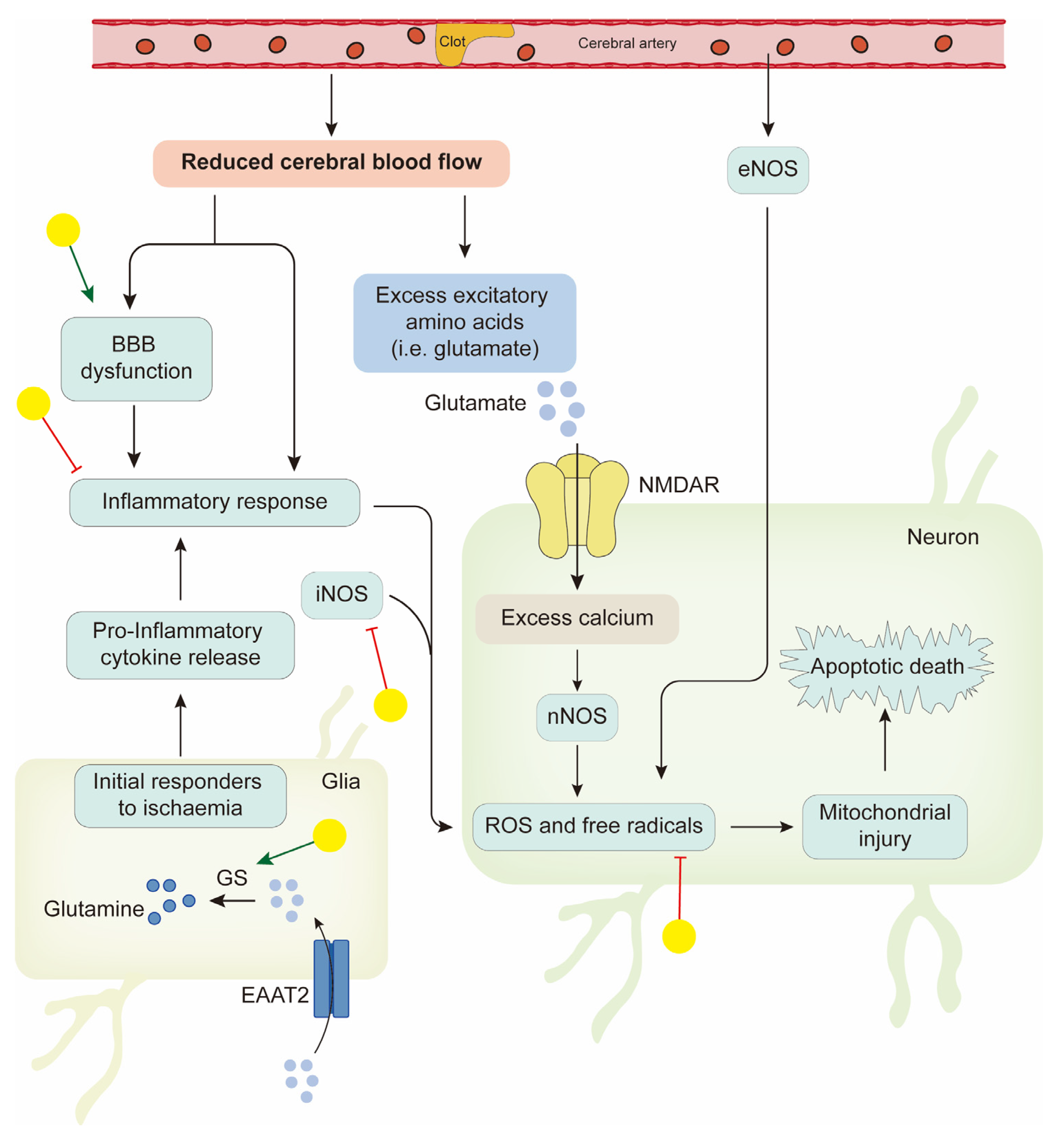
4.1. Neuroprotection in Ischemic Stroke
4.2. Neuroprotection in Neurodegenerative Diseases
4.3. Antidepression
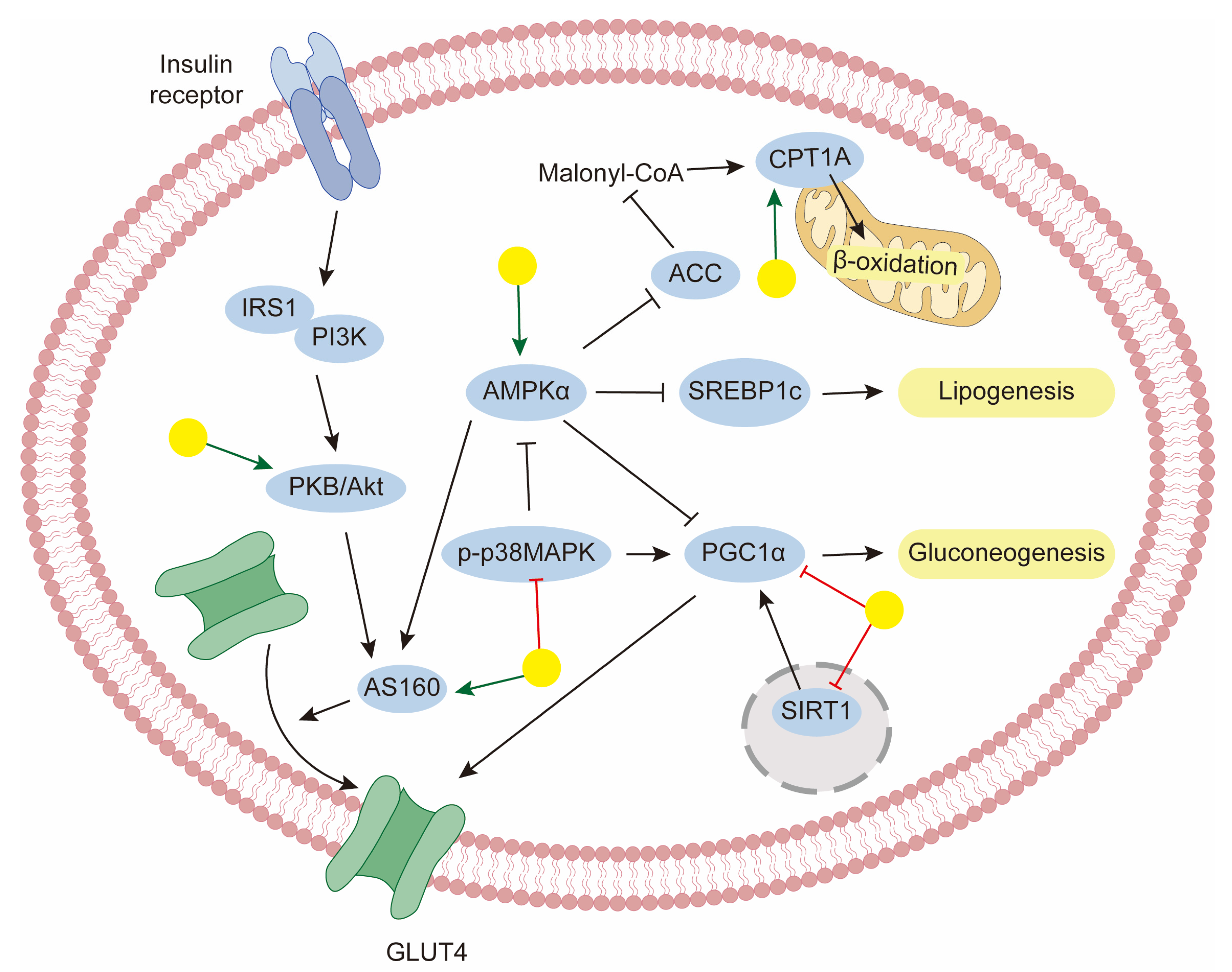
5. Regulation of Metabolic Disorders
5.1. Obesity and NAFLD
5.2. Diabetes
6. Discussion
- Identifying specific molecular targets in different diseases;
- Exploring the dose–response relationship between baicalin and the tissue-specific response mechanism under multi-organ and multi-pathological conditions to delineate its tissue selectivity;
- Standardizing baicalin preparations, clarifying its pharmacokinetic characteristics, and exploring its application in combination therapies to bridge the gap between bench and bedside;
- Optimizing chemical structure or delivery systems to enhance its oral bioavailability, stability, and overall pharmacological efficacy.
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TCM | traditional Chinese medicine |
| HCC | hepatocellular carcinoma |
| TME | tumor microenvironment |
| TAM | tumor-associated macrophage |
| ROS | reactive oxygen species |
| CVDs | cardiovascular diseases |
| H/R | hypoxia/reoxygenation |
| IR | ischemia–reperfusion |
| Ang II | angiotensin II |
| AAC | abdominal aortic constriction |
| VSMCs | vascular smooth muscle cells |
| PDGF | platelet-derived growth factor |
| VEGF | vascular endothelial growth factor |
| VDCCs | voltage-dependent Ca2+ channels |
| BK | large-conductance Ca2+-activated channels |
| BBB | Blood–brain barrier |
| NMDA | N-methyl-d-aspartate |
| NOS | nitric oxide synthase |
| GS | glutamine synthetase |
| pMCAO | permanent middle cerebral artery occlusion |
| TNFα | tumor necrosis factor α |
| AD | Alzheimer’s disease |
| PD | Parkinson’s disease |
| CNS | central nervous system. |
| BSCB | blood–spinal cord barrier |
| PPAR | peroxisome proliferator-activated receptor |
| T2DM | type 2 diabetes mellitus |
| NAFLD | non-alcoholic fatty liver disease |
| SREBP-1c | sterol-CoA response element binding protein-1c |
| AMPK | AMP-activated protein kinase |
| HFD | high-fat diet |
| CPT1 | carnitine palmitoyl-transferase1 |
| GLUT4 | glucose transporter isoform 4 |
| DARTS | drug affinity responsive target stability |
References
- Corson, T.W.; Crews, C.M. Molecular Understanding and Modern Application of Traditional Medicines: Triumphs and Trials. Cell 2007, 130, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Kase, Y.; Saitoh, K.; Makino, B.; Hashimoto, K.; Ishige, A.; Komatsu, Y. Relationship between the antidiarrhoeal effects of Hange-Shashin-To and its active components. Phytother. Res. 1999, 13, 468–473. [Google Scholar] [CrossRef]
- Yang, H.J.; Ma, J.Y.; Weon, J.B.; Lee, B.; Ma, C.J. Qualitative and quantitative simultaneous determination of six marker compounds in Soshiho-tang by HPLC-DAD-ESI-MS. Arch. Pharm. Res. 2012, 35, 1785–1791. [Google Scholar] [CrossRef]
- Zhang, D.; Fu, M.; Song, C.; Wang, C.; Lin, X.; Liu, Y. Expressions of apoptosis-related proteins in rats with focal cerebral ischemia after Angong Niuhuang sticker point application. Neural Regen. Res. 2012, 7, 2347–2353. [Google Scholar]
- Zhu, H.; Qian, Z.; He, F.; Liu, M.; Pan, L.; Zhang, Q.; Tang, Y. Novel pharmacokinetic studies of the Chinese formula Huang-Lian-Jie-Du-Tang in MCAO rats. Phytomedicine 2013, 20, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, J.; Liu, J.; Qian, G.; Fu, C. Quality evaluation of Huaijiao pill by chromatographic fingerprint and simultaneous determination of its major bioactive components. J. Pharm. Anal. 2016, 6, 249–255. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, D.; Yang, X.; Fu, C.; Zou, L.; Zhang, J. Traditional Chinese medicine Gegen Qinlian decoction ameliorates irinotecan chemotherapy-induced gut toxicity in mice. Biomed. Pharmacother. 2019, 109, 2252–2261. [Google Scholar] [CrossRef]
- Su, H.X.; Yao, S.; Zhao, W.F.; Li, M.J.; Liu, J.; Shang, W.J.; Xie, H.; Ke, C.Q.; Hu, H.C.; Gao, M.N.; et al. Anti-SARS-CoV-2 activities in vitro of Shuanghuanglian preparations and bioactive ingredients. Acta Pharmacol. Sin. 2020, 41, 1167–1177. [Google Scholar] [CrossRef]
- Wen, Y.; Han, C.; Liu, T.; Wang, R.; Cai, W.; Yang, J.; Liang, G.; Yao, L.; Shi, N.; Fu, X.; et al. Chaiqin chengqi decoction alleviates severity of acute pancreatitis via inhibition of TLR4 and NLRP3 inflammasome: Identification of bioactive ingredients via pharmacological sub-network analysis and experimental validation. Phytomedicine 2020, 79, 153328. [Google Scholar] [CrossRef]
- Wu, X.; Shen, A.; Bao, L.; Wu, M.; Lin, X.; Wang, H.; Chen, Y.; Cai, Q.; Lin, S.; Zhou, X.; et al. Qingda granules attenuate hypertensive cardiac remodeling and inflammation in spontaneously hypertensive rats. Biomed. Pharmacother. 2020, 129, 110367. [Google Scholar] [CrossRef]
- Ma, Q.; Wang, Z.; Chen, R.; Lei, B.; Liu, B.; Jiang, H.; Chen, Z.; Cai, X.; Guo, X.; Zhou, M.; et al. Effect of Jinzhen granule on two coronaviruses: The novel SARS-CoV-2 and the HCoV-229E and the evidences for their mechanisms of action. Phytomedicine 2022, 95, 153874. [Google Scholar] [CrossRef]
- Chen, X.; Yang, J.B.; Cao, H.H.; Fang, X.C.; Liu, S.H.; Zou, L.F.; Yu, J.H.; Zuo, J.P.; Zhao, W.; Lu, Z.B.; et al. Liang-Ge-San inhibits dengue virus serotype 2 infection by reducing caveolin1-induced cytoplasmic heat shock protein 70 translocation into the plasma membrane. Phytomedicine 2023, 119, 154977. [Google Scholar] [CrossRef]
- Liu, S.; Ding, P.; Wu, M.; Zhu, Z.; Tao, J.; Wang, J.; Xue, Z.; Wang, R. Screening quality markers (Q-markers) of Xiaoer Chaige Tuire Oral Liquid by in vitro sequential metabolism and in vivo biopharmaceutical analysis. Phytomedicine 2023, 116, 154844. [Google Scholar] [CrossRef]
- Ma, H.; Fu, W.; Yu, H.; Xu, Y.; Xiao, L.; Zhang, Y.; Wu, Y.; Liu, X.; Chen, Y.; Xu, T. Exploration of the anti-inflammatory mechanism of Lanqin oral solution based on the network pharmacology analysis optimized by Q-markers selection. Comput. Biol. Med. 2023, 154, 106607. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Chen, C.; Zheng, Y.; Zhang, S.; Yan, J.; Cheng, F.; Wang, X.; Wang, Q.; Li, C. Refined Qingkailing protects the in vitro neurovascular unit against oxygen-glucose deprivation and re-oxygenation-induced injury. Biomed. Pharmacother. 2023, 167, 115580. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, J.; Jin, X.; Ma, Q.; Li, H.; Zhou, Q.; Chen, W. Bu-Shen-Ning-Xin decoction ameliorates premature ovarian insufficiency by suppressing oxidative stress through rno_circRNA_012284/rno_miR-760-3p/HBEGF pathway. Phytomedicine 2024, 133, 155920. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Zhang, G.; Wu, Y.; Hua, Y.; Ding, W.; Han, X.; Liu, B.; Zhou, C.; Li, A. Protective effects of Wenqingyin on sepsis-induced acute lung injury through regulation of the receptor for advanced glycation end products pathway. Phytomedicine 2024, 129, 155654. [Google Scholar] [CrossRef]
- Ma, S.; Wei, T.; Zhang, B.; Zhang, Y.; Lai, J.; Qu, J.; Liu, J.; Yin, P.; Shang, D. Integrated pharmacokinetic properties and tissue distribution of multiple active constituents in Qing-Yi Recipe: A comparison between granules and decoction. Phytomedicine 2024, 129, 155645. [Google Scholar] [CrossRef]
- Lai, M.; Sun, S.; Zuo, T.; Li, L.; Zhao, Q.; Li, W.; Zheng, J.; Hong, M. Sanfeng Tongqiao Dripping Pills alleviate House Dust Mite-induced allergic rhinitis in mice by inhibiting Th2 differentiation and repairing the nasal epithelial barrier. Phytomedicine 2024, 132, 155899. [Google Scholar] [CrossRef]
- Ma, W.; Liu, T.; Ogaji, O.D.; Li, J.; Du, K.; Chang, Y. Recent advances in Scutellariae Radix: A comprehensive review on ethnobotanical uses, processing, phytochemistry, pharmacological effects, quality control and influence factors of biosynthesis. Heliyon 2024, 10, e36146. [Google Scholar] [CrossRef]
- Wang, F.; Wang, B.; Wang, L.; Xiong, Z.-Y.; Gao, W.; Li, P.; Li, H.-J. Discovery of discriminatory quality control markers for Chinese herbal medicines and related processed products by combination of chromatographic analysis and chemometrics methods: Radix Scutellariae as a case study. J. Pharm. Biomed. Anal. 2017, 138, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Bochoráková, H.; Paulová, H.; Slanina, J.; Musil, P.; Táborská, E. Main flavonoids in the root of Scutellaria baicalensis cultivated in Europe and their comparative antiradical properties. Phytother. Res. 2003, 17, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Chen, X.; Zhong, D. Absorption and enterohepatic circulation of baicalin in rats. Life Sci. 2005, 78, 140–146. [Google Scholar] [CrossRef]
- Wu, H.; Long, X.; Yuan, F.; Chen, L.; Pan, S.; Liu, Y.; Stowell, Y.; Li, X. Combined use of phospholipid complexes and self-emulsifying microemulsions for improving the oral absorption of a BCS class IV compound, baicalin. Acta Pharm. Sin. B 2014, 4, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Guo, J.; Zheng, X.; Wu, J.; Zhou, Y.; Yu, Y.; Ye, Y.; Zhang, L.; Zhao, L. Preparation, pharmacokinetics and biodistribution of baicalin-loaded liposomes. Int. J. Nanomed. 2014, 9, 3623–3630. [Google Scholar]
- Kim, Y.S.; Kim, J.J.; Cho, K.H.; Jung, W.S.; Moon, S.K.; Park, E.K.; Kim, D.H. Biotransformation of ginsenoside Rb1, crocin, amygdalin, geniposide, puerarin, ginsenoside Re, hesperidin, poncirin, glycyrrhizin, and baicalin by human fecal microflora and its relation to cytotoxicity against tumor cells. J. Microbiol. Biotechnol. 2008, 18, 1109–1114. [Google Scholar]
- Kim, D.H.; Jung, E.A.; Sohng, I.S.; Han, J.A.; Kim, T.H.; Han, M.J. Intestinal bacterial metabolism of flavonoids and its relation to some biological activities. Arch. Pharm. Res. 1998, 21, 17–23. [Google Scholar] [CrossRef]
- Mano, E.C.C.; Scott, A.L.; Honorio, K.M. UDP-glucuronosyltransferases: Structure, Function and Drug Design Studies. Curr. Med. Chem. 2018, 25, 3247–3255. [Google Scholar] [CrossRef]
- Kang, M.J.; Ko, G.S.; Oh, D.G.; Kim, J.S.; Noh, K.; Kang, W.; Yoon, W.K.; Kim, H.C.; Jeong, H.G.; Jeong, T.C. Role of metabolism by intestinal microbiota in pharmacokinetics of oral baicalin. Arch. Pharm. Res. 2014, 37, 371–378. [Google Scholar] [CrossRef]
- Chen, H.; Gao, Y.; Wu, J.; Chen, Y.; Chen, B.; Hu, J.; Zhou, J. Exploring therapeutic potentials of baicalin and its aglycone baicalein for hematological malignancies. Cancer Lett. 2014, 354, 5–11. [Google Scholar] [CrossRef]
- Lai, M.Y.; Hsiu, S.L.; Tsai, S.Y.; Hou, Y.C.; Chao, P.D. Comparison of metabolic pharmacokinetics of baicalin and baicalein in rats. J. Pharm. Pharmacol. 2003, 55, 205–209. [Google Scholar] [CrossRef]
- Huang, Y.; Hu, J.; Zheng, J.; Li, J.; Wei, T.; Zheng, Z.; Chen, Y. Down-regulation of the PI3K/Akt signaling pathway and induction of apoptosis in CA46 Burkitt lymphoma cells by baicalin. J. Exp. Clin. Cancer Res. 2012, 31, 48. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.Y.; Wang, N.; Man, K.; Tsao, S.W.; Che, C.M.; Feng, Y. Autophagy-induced RelB/p52 activation mediates tumour-associated macrophage repolarisation and suppression of hepatocellular carcinoma by natural compound baicalin. Cell Death Dis. 2015, 6, e1942. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, L.; Su, M.; Zhou, Y.; Mao, K.; Li, C.; Peng, G.; Zhou, C.; Shen, B.; Dou, J. Baicalin induces cellular senescence in human colon cancer cells via upregulation of DEPP and the activation of Ras/Raf/MEK/ERK signaling. Cell Death Dis. 2018, 9, 217. [Google Scholar] [CrossRef] [PubMed]
- Zakki, S.A.; Cui, Z.G.; Sun, L.; Feng, Q.W.; Li, M.L.; Inadera, H. Baicalin Augments Hyperthermia-Induced Apoptosis in U937 Cells and Modulates the MAPK Pathway via ROS Generation. Cell Physiol. Biochem. 2018, 45, 2444–2460. [Google Scholar] [CrossRef]
- Kong, N.; Chen, X.; Feng, J.; Duan, T.; Liu, S.; Sun, X.; Chen, P.; Pan, T.; Yan, L.; Jin, T.; et al. Baicalin induces ferroptosis in bladder cancer cells by downregulating FTH1. Acta Pharm. Sin. B 2021, 11, 4045–4054. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.R.; Jiang, Z.B.; Xu, C.; Meng, W.Y.; Liu, P.; Zhang, Y.Z.; Xie, C.; Xu, J.Y.; Xie, Y.J.; Liang, T.L.; et al. Andrographolide suppresses non-small-cell lung cancer progression through induction of autophagy and antitumor immune response. Pharmacol. Res. 2022, 179, 106198. [Google Scholar] [CrossRef]
- Shehatta, N.H.; Okda, T.M.; Omran, G.A.; Abd-Alhaseeb, M.M. Baicalin; a promising chemopreventive agent, enhances the antitumor effect of 5-FU against breast cancer and inhibits tumor growth and angiogenesis in Ehrlich solid tumor. Biomed Pharmacother. 2022, 146, 112599. [Google Scholar] [CrossRef]
- Liu, H.; Liu, H.; Zhou, Z.; Chung, J.; Zhang, G.; Chang, J.; Parise, R.A.; Chu, E.; Schmitz, J.C. Scutellaria baicalensis enhances 5-fluorouracil-based chemotherapy via inhibition of proliferative signaling pathways. Cell Commun. Signal 2023, 21, 147. [Google Scholar] [CrossRef]
- Sun, J.; Yang, X.; Sun, H.; Huang, S.; An, H.; Xu, W.; Chen, W.; Zhao, W.; He, C.; Zhong, X.; et al. Baicalin inhibits hepatocellular carcinoma cell growth and metastasis by suppressing ROCK1 signaling. Phytother. Res. 2023, 37, 4117–4132. [Google Scholar] [CrossRef]
- Wen, R.J.; Dong, X.; Zhuang, H.W.; Pang, F.X.; Ding, S.C.; Li, N.; Mai, Y.X.; Zhou, S.T.; Wang, J.Y.; Zhang, J.F. Baicalin induces ferroptosis in osteosarcomas through a novel Nrf2/xCT/GPX4 regulatory axis. Phytomedicine 2023, 116, 154881. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Khan, S.U.; Yan, J.; Lu, J.; Yang, C.; Tong, Q. Baicalin enhances the efficacy of 5-Fluorouracil in gastric cancer by promoting ROS-mediated ferroptosis. Biomed. Pharmacother. 2023, 164, 114986. [Google Scholar] [CrossRef] [PubMed]
- He, X.W.; Yu, D.; Li, W.L.; Zheng, Z.; Lv, C.L.; Li, C.; Liu, P.; Xu, C.Q.; Hu, X.F.; Jin, X.P. Anti-atherosclerotic potential of baicalin mediated by promoting cholesterol efflux from macrophages via the PPARγ-LXRα-ABCA1/ABCG1 pathway. Biomed. Pharmacother. 2016, 83, 257–264. [Google Scholar] [CrossRef]
- Zhang, K.; Lu, J.; Mori, T.; Smith-Powell, L.; Synold, T.W.; Chen, S.; Wen, W. Baicalin increases VEGF expression and angiogenesis by activating the ERRα/PGC-1αpathway. Cardiovasc. Res. 2011, 89, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Jiang, S.; Huang, C.; Shen, A.; Zhang, X.; Yang, W.; Xiao, Y.; Gao, S.; Du, R.; Zheng, G.; et al. Baicalin inhibits pressure overload-induced cardiac hypertrophy by regulating the SIRT3-dependent signaling pathway. Phytomedicine 2023, 114, 154747. [Google Scholar] [CrossRef]
- Huang, X.; Wu, P.; Huang, F.; Xu, M.; Chen, M.; Huang, K.; Li, G.P.; Xu, M.; Yao, D.; Wang, L. Baicalin attenuates chronic hypoxia-induced pulmonary hypertension via adenosine A(2A) receptor-induced SDF-1/CXCR4/PI3K/AKT signaling. J. Biomed. Sci. 2017, 24, 52. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Cheng, Y.; Chu, J.; Wu, M.; Yan, M.; Wang, D.; Xie, Q.; Ali, F.; Fang, Y.; Wei, L.; et al. Baicalin attenuates angiotensin II-induced blood pressure elevation and modulates MLCK/p-MLC signaling pathway. Biomed. Pharmacother. 2021, 143, 112124. [Google Scholar] [CrossRef]
- Xiong, P.; Chen, X.; Guo, C.; Zhang, N.; Ma, B. Baicalin and deferoxamine alleviate iron accumulation in different brain regions of Parkinson’s disease rats. Neural Regen. Res. 2012, 7, 2092–2098. [Google Scholar]
- Bitto, A.; Giuliani, D.; Pallio, G.; Irrera, N.; Vandini, E.; Canalini, F.; Zaffe, D.; Ottani, A.; Minutoli, L.; Rinaldi, M.; et al. Effects of COX1-2/5-LOX blockade in Alzheimer transgenic 3xTg-AD mice. Inflamm. Res. 2017, 66, 389–398. [Google Scholar] [CrossRef]
- Ai, R.S.; Xing, K.; Deng, X.; Han, J.J.; Hao, D.X.; Qi, W.H.; Han, B.; Yang, Y.N.; Li, X.; Zhang, Y. Baicalin Promotes CNS Remyelination via PPARγ Signal Pathway. Neurol. Neuroimmunol. Neuroinflamm. 2022, 9, e1142. [Google Scholar] [CrossRef]
- Zhao, R.; Wu, X.; Bi, X.Y.; Yang, H.; Zhang, Q. Baicalin attenuates blood-spinal cord barrier disruption and apoptosis through PI3K/Akt signaling pathway after spinal cord injury. Neural Regen. Res. 2022, 17, 1080–1087. [Google Scholar] [PubMed]
- Lee, H.; Kang, R.; Hahn, Y.; Yang, Y.; Kim, S.S.; Cho, S.H.; Chung, S.I.; Yoon, Y. Antiobesity effect of baicalin involves the modulations of proadipogenic and antiadipogenic regulators of the adipogenesis pathway. Phytother. Res. 2009, 23, 1615–1623. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.X.; Liu, D.H.; Ma, Y.; Liu, J.F.; Wang, Y.; Du, Z.Y.; Wang, X.; Shen, J.K.; Peng, H.L. Long-term baicalin administration ameliorates metabolic disorders and hepatic steatosis in rats given a high-fat diet. Acta Pharmacol. Sin. 2009, 30, 1505–1512. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Liang, K.; Zhao, S.; Jia, W.; Liu, Y.; Wu, H.; Lv, J.; Cao, C.; Chen, T.; Zhuang, S.; et al. Chemoproteomics reveals baicalin activates hepatic CPT1 to ameliorate diet-induced obesity and hepatic steatosis. Proc. Natl. Acad. Sci. USA 2018, 115, E5896–E5905. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.; Deng, X.; Zhang, N.; Liu, B.; Xin, S.; Li, G.; Xu, K. Baicalin attenuates non-alcoholic steatohepatitis by suppressing key regulators of lipid metabolism, inflammation and fibrosis in mice. Life Sci. 2018, 192, 46–54. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, J.; Hao, Z.; Sun, N.; Guo, J.; Zheng, X.; Sun, P.; Yin, W.; Fan, K.; Li, H. Baicalin ameliorates high fat diet-induced nonalcoholic fatty liver disease in mice via adenosine monophosphate-activated protein kinase-mediated regulation of SREBP1/Nrf2/NF-κB signaling pathways. Phytother. Res. 2023, 37, 2405–2418. [Google Scholar] [CrossRef]
- Liu, W.J.; Chen, W.W.; Chen, J.Y.; Sun, Y.B.; Chang, D.; Wang, C.X.; Xie, J.D.; Lin, W.; Li, S.H.; Xu, W.; et al. Baicalin attenuated metabolic dysfunction-associated fatty liver disease by suppressing oxidative stress and inflammation via the p62-Keap1-Nrf2 signalling pathway in db/db mice. Phytother. Res. 2023, 39, 1663–1678. [Google Scholar] [CrossRef]
- Nam, J.E.; Jo, S.Y.; Ahn, C.W.; Kim, Y.S. Baicalin attenuates fibrogenic process in human renal proximal tubular cells (HK-2) exposed to diabetic milieu. Life Sci. 2020, 254, 117742. [Google Scholar] [CrossRef]
- Hu, H.; Li, W.; Hao, Y.; Peng, Z.; Zou, Z.; Liang, W. Baicalin ameliorates renal fibrosis by upregulating CPT1α-mediated fatty acid oxidation in diabetic kidney disease. Phytomedicine 2024, 122, 155162. [Google Scholar] [CrossRef]
- Li, Z.; Xia, X.; Zhang, S.; Zhang, A.; Bo, W.; Zhou, R. Up-regulation of Toll-like receptor 4 was suppressed by emodin and baicalin in the setting of acute pancreatitis. Biomed. Pharmacother. 2009, 63, 120–128. [Google Scholar] [CrossRef]
- Ma, C.; Ma, Z.; Fu, Q.; Ma, S. Anti-asthmatic effects of baicalin in a mouse model of allergic asthma. Phytother. Res. 2014, 28, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Zhai, L.; Peng, J.; Wu, H.; Bian, Z.; Xiao, H. Phytochemicals as regulators of Th17/Treg balance in inflammatory bowel diseases. Biomed. Pharmacother. 2021, 141, 111931. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Lee, J.S.; Choi, J.S.; Nam, Y.J.; Han, J.H.; Byun, H.D.; Song, M.J.; Oh, J.S.; Kim, S.G.; Choi, Y. Identification and Characterization of Baicalin as a Phosphodiesterase 4 Inhibitor. Phytother. Res. 2016, 30, 144–151. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Wang, H.; Sui, H.H.; Li, L.; Zhou, C.L.; Huang, J.J. Inhibitory effect of baicalin on allergic response in ovalbumin-induced allergic rhinitis guinea pigs and lipopolysaccharide-stimulated human mast cells. Inflamm. Res. 2016, 65, 603–612. [Google Scholar] [CrossRef]
- Zhang, C.L.; Zhang, S.; He, W.X.; Lu, J.L.; Xu, Y.J.; Yang, J.Y.; Liu, D. Baicalin may alleviate inflammatory infiltration in dextran sodium sulfate-induced chronic ulcerative colitis via inhibiting IL-33 expression. Life Sci. 2017, 186, 125–132. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Q.; Gao, Z.; Yu, C.; Zhang, L. Baicalin alleviates IL-1β-induced inflammatory injury via down-regulating miR-126 in chondrocytes. Biomed. Pharmacother. 2018, 99, 184–190. [Google Scholar] [CrossRef]
- Ji, W.; Liang, K.; An, R.; Wang, X. Baicalin protects against ethanol-induced chronic gastritis in rats by inhibiting Akt/NF-κB pathway. Life Sci. 2019, 239, 117064. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yang, X.; Yang, J.; Li, M. Baicalin ameliorates lupus autoimmunity by inhibiting differentiation of Tfh cells and inducing expansion of Tfr cells. Cell Death Dis. 2019, 10, 140. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Song, S.; Wang, Y.; Zhu, J.; Li, X. Potential mechanism of oral baicalin treating psoriasis via suppressing Wnt signaling pathway and inhibiting Th17/IL-17 axis by activating PPARγ. Phytother. Res. 2022, 36, 3969–3987. [Google Scholar] [CrossRef]
- Picciolo, G.; Mannino, F.; Irrera, N.; Minutoli, L.; Altavilla, D.; Vaccaro, M.; Oteri, G.; Squadrito, F.; Pallio, G. Reduction of oxidative stress blunts the NLRP3 inflammatory cascade in LPS stimulated human gingival fibroblasts and oral mucosal epithelial cells. Biomed. Pharmacother. 2022, 146, 112525. [Google Scholar] [CrossRef]
- Braicu, C.; Zanoaga, O.; Zimta, A.-A.; Tigu, A.B.; Kilpatrick, K.L.; Bishayee, A.; Nabavi, S.M.; Berindan-Neagoe, I. Natural compounds modulate the crosstalk between apoptosis- and autophagy-regulated signaling pathways: Controlling the uncontrolled expansion of tumor cells. Semin. Cancer Biol. 2022, 80, 218–236. [Google Scholar] [CrossRef] [PubMed]
- Motoo, Y.; Sawabu, N. Antitumor effects of saikosaponins, baicalin and baicalein on human hepatoma cell lines. Cancer Lett. 1994, 86, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Yano, H.; Mizoguchi, A.; Fukuda, K.; Haramaki, M.; Ogasawara, S.; Momosaki, S.; Kojiro, M. The herbal medicine sho-saiko-to inhibits proliferation of cancer cell lines by inducing apoptosis and arrest at the G0/G1 phase. Cancer Res. 1994, 54, 448–454. [Google Scholar]
- Kato, M.; Liu, W.; Yi, H.; Asai, N.; Hayakawa, A.; Kozaki, K.; Takahashi, M.; Nakashima, I. The herbal medicine Sho-saiko-to inhibits growth and metastasis of malignant melanoma primarily developed in ret-transgenic mice. J. Investig. Dermatol. 1998, 111, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Chan, F.L.; Choi, H.L.; Chen, Z.Y.; Chan, P.S.; Huang, Y. Induction of apoptosis in prostate cancer cell lines by a flavonoid, baicalin. Cancer Lett. 2000, 160, 219–228. [Google Scholar] [CrossRef]
- Chen, S.; Ruan, Q.; Bedner, E.; Deptala, A.; Wang, X.; Hsieh, T.C.; Traganos, F.; Darzynkiewicz, Z. Effects of the flavonoid baicalin and its metabolite baicalein on androgen receptor expression, cell cycle progression and apoptosis of prostate cancer cell lines. Cell Prolif. 2001, 34, 293–304. [Google Scholar] [CrossRef]
- Wu, Y.X.; Sato, E.; Kimura, W.; Miura, N. Baicalin and scutellarin are proteasome inhibitors that specifically target chymotrypsin-like catalytic activity. Phytother. Res. 2013, 27, 1362–1367. [Google Scholar] [CrossRef]
- Li-Weber, M. New therapeutic aspects of flavones: The anticancer properties of Scutellaria and its main active constituents Wogonin, Baicalein and Baicalin. Cancer Treat. Rev. 2009, 35, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Meena, A.; Luqman, S. Baicalin mediated regulation of key signaling pathways in cancer. Pharmacol. Res. 2021, 164, 105387. [Google Scholar] [CrossRef]
- Zhou, Q.M.; Wang, S.; Zhang, H.; Lu, Y.Y.; Wang, X.F.; Motoo, Y.; Su, S.B. The combination of baicalin and baicalein enhances apoptosis via the ERK/p38 MAPK pathway in human breast cancer cells. Acta Pharmacol. Sin. 2009, 30, 1648–1658. [Google Scholar] [CrossRef]
- Wang, C.Z.; Li, X.L.; Wang, Q.F.; Mehendale, S.R.; Yuan, C.S. Selective fraction of Scutellaria baicalensis and its chemopreventive effects on MCF-7 human breast cancer cells. Phytomedicine 2010, 17, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Liu, X.; Lin, D.; Zhang, L.; Wu, Y.; Chang, Y.; Jin, M.; Huang, G. Baicalin induces cell death of non-small cell lung cancer cells via MCOLN3-mediated lysosomal dysfunction and autophagy blockage. Phytomedicine 2024, 133, 155872. [Google Scholar] [CrossRef]
- Yang, K.; Zeng, L.; Ge, A.; Chen, Z.; Bao, T.; Long, Z.; Ge, J.; Huang, L. Investigating the regulation mechanism of baicalin on triple negative breast cancer’s biological network by a systematic biological strategy. Biomed. Pharmacother. 2019, 118, 109253. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Liu, B.; Sun, J.; Lu, L.; Liu, L.; Qiu, J.; Li, Q.; Yan, C.; Jiang, S.; Mohammadtursun, N.; et al. Scutellaria Flavonoids Effectively Inhibit the Malignant Phenotypes of Non-small Cell Lung Cancer in an Id1-dependent Manner. Int. J. Biol. Sci. 2019, 15, 1500–1513. [Google Scholar] [CrossRef]
- Jin, M.Z.; Jin, W.L. The updated landscape of tumor microenvironment and drug repurposing. Signal Transduct. Target. Ther. 2020, 5, 166. [Google Scholar] [CrossRef]
- Cassetta, L.; Pollard, J.W. A timeline of tumour-associated macrophage biology. Nat. Rev. Cancer 2023, 23, 238–257. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, L.; Zhu, J.; Zhang, Y.; Yang, R.; Yan, J.; Huang, R.; Zheng, C.; Xiao, W.; Huang, C.; et al. Predicting the herbal medicine triggering innate anti-tumor immunity from a system pharmacology perspective. Biomed. Pharmacother. 2021, 143, 112105. [Google Scholar] [CrossRef]
- Han, S.; Wang, W.; Wang, S.; Yang, T.; Zhang, G.; Wang, D.; Ju, R.; Lu, Y.; Wang, H.; Wang, L. Tumor microenvironment remodeling and tumor therapy based on M2-like tumor associated macrophage-targeting nano-complexes. Theranostics 2021, 11, 2892–2916. [Google Scholar] [CrossRef]
- Gong, S.Q.; Sun, W.; Wang, M.; Fu, Y.Y. Role of TLR4 and TCR or BCR against baicalin-induced responses in T and B cells. Int. Immunopharmacol. 2011, 11, 2176–2180. [Google Scholar] [CrossRef]
- Lin, M.Y.; Cheng, W.T.; Cheng, H.C.; Chou, W.C.; Chen, H.I.; Ou, H.C.; Tsai, K.L. Baicalin Enhances Chemosensitivity to Doxorubicin in Breast Cancer Cells via Upregulation of Oxidative Stress-Mediated Mitochondria-Dependent Apoptosis. Antioxidants 2021, 10, 1506. [Google Scholar] [CrossRef]
- Wang, G.; Liang, J.; Gao, L.R.; Si, Z.P.; Zhang, X.T.; Liang, G.; Yan, Y.; Li, K.; Cheng, X.; Bao, Y.; et al. Baicalin administration attenuates hyperglycemia-induced malformation of cardiovascular system. Cell Death Dis. 2018, 9, 234. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Wu, X.D.; Davey, A.K.; Wang, J. The anti-inflammatory effect of baicalin on hypoxia/reoxygenation and TNF-alpha induced injury in cultural rat cardiomyocytes. Phytother. Res. 2010, 24, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Wang, Q.; Qi, J.; Yu, H.; Wang, C.; Wang, X.; Ren, Y.; Yang, F. Promoting effect of baicalin on nitric oxide production in CMECs via activating the PI3K-AKT-eNOS pathway attenuates myocardial ischemia-reperfusion injury. Phytomedicine 2019, 63, 153035. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Jiang, L.; Yan, Q.; Zhou, C.; Guo, X.; Chen, T.; Ma, S.; Luo, Y.; Hu, C.; Yang, F.; et al. Evidence construction of baicalin for treating myocardial ischemia diseases: A preclinical meta-analysis. Phytomedicine 2022, 107, 154476. [Google Scholar] [CrossRef]
- Weber, C.; Noels, H. Atherosclerosis: Current pathogenesis and therapeutic options. Nat. Med. 2011, 17, 1410–1422. [Google Scholar] [CrossRef]
- Dong, L.H.; Wen, J.K.; Miao, S.B.; Jia, Z.; Hu, H.J.; Sun, R.H.; Wu, Y.; Han, M. Baicalin inhibits PDGF-BB-stimulated vascular smooth muscle cell proliferation through suppressing PDGFRβ-ERK signaling and increase in p27 accumulation and prevents injury-induced neointimal hyperplasia. Cell Res. 2010, 20, 1252–1262. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, F.; Fan, L.; Zhang, W.; Wang, T.; Du, Y.; Bai, X. Baicalin alleviates atherosclerosis by relieving oxidative stress and inflammatory responses via inactivating the NF-κB and p38 MAPK signaling pathways. Biomed. Pharmacother. 2018, 97, 1673–1679. [Google Scholar] [CrossRef]
- Paudel, K.R.; Kim, D.W. Microparticles-Mediated Vascular Inflammation and its Amelioration by Antioxidant Activity of Baicalin. Antioxidants 2020, 9, 890. [Google Scholar] [CrossRef] [PubMed]
- Mills, K.T.; Stefanescu, A.; He, J. The global epidemiology of hypertension. Nat. Rev. Nephrol. 2020, 16, 223–237. [Google Scholar] [CrossRef]
- Daghbouche-Rubio, N.; Lopez-Lopez, J.R.; Perez-Garcia, M.T.; Cidad, P. Vascular smooth muscle ion channels in essential hypertension. Front. Physiol. 2022, 13, 1016175. [Google Scholar] [CrossRef]
- Lin, Y.L.; Dai, Z.K.; Lin, R.J.; Chu, K.S.; Chen, I.J.; Wu, J.R.; Wu, B.N. Baicalin, a flavonoid from Scutellaria baicalensis Georgi, activates large-conductance Ca2+-activated K+ channels via cyclic nucleotide-dependent protein kinases in mesenteric artery. Phytomedicine 2010, 17, 760–770. [Google Scholar] [CrossRef]
- Ding, L.; Jia, C.; Zhang, Y.; Wang, W.; Zhu, W.; Chen, Y.; Zhang, T. Baicalin relaxes vascular smooth muscle and lowers blood pressure in spontaneously hypertensive rats. Biomed. Pharmacother. 2019, 111, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Zhang, S.; Li, C.; Liu, Y.; Zhao, J.; Wang, Y.; Yang, Y.; Zhang, L. 5-Lipoxygenase as an emerging target against age-related brain disorders. Ageing Res. Rev. 2021, 69, 101359. [Google Scholar] [CrossRef]
- Jiang, H.; Lv, P.; Li, J.; Wang, H.; Zhou, T.; Liu, Y.; Lin, W. Baicalin inhibits colistin sulfate-induced apoptosis of PC12 cells. Neural Regen. Res. 2013, 8, 2597–2604. [Google Scholar]
- Banks, W.A. From blood-brain barrier to blood-brain interface: New opportunities for CNS drug delivery. Nat. Rev. Drug Discov. 2016, 15, 275–292. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Huang, X.; Chen, W. The Effects of Baicalin and Baicalein on Cerebral Ischemia: A Review. Aging Dis. 2017, 8, 850–867. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, S.; Wan, J.; Yang, Q.; Xiang, Y.; Ni, L.; Long, Y.; Cui, M.; Ci, Z.; Tang, D.; et al. Preparation, Characterization and in vivo Study of Borneol-Baicalin-Liposomes for Treatment of Cerebral Ischemia-Reperfusion Injury. Int. J. Nanomed. 2020, 15, 5977–5989. [Google Scholar] [CrossRef]
- Long, Y.; Liu, S.; Wan, J.; Zhang, Y.; Li, D.; Yu, S.; Shi, A.; Li, N.; He, F. Brain targeted borneol-baicalin liposome improves blood-brain barrier integrity after cerebral ischemia-reperfusion injury via inhibiting HIF-1α/VEGF/eNOS/NO signal pathway. Biomed. Pharmacother. 2023, 160, 114240. [Google Scholar] [CrossRef]
- Long, Y.; Yang, Q.; Xiang, Y.; Zhang, Y.; Wan, J.; Liu, S.; Li, N.; Peng, W. Nose to brain drug delivery—A promising strategy for active components from herbal medicine for treating cerebral ischemia reperfusion. Pharmacol. Res. 2020, 159, 104795. [Google Scholar] [CrossRef]
- Li, X.; Li, S.; Ma, C.; Li, T.; Yang, L. Preparation of baicalin-loaded ligand-modified nanoparticles for nose-to-brain delivery for neuroprotection in cerebral ischemia. Drug Deliv. 2022, 29, 1282–1298. [Google Scholar] [CrossRef]
- Campbell, B.C.V.; De Silva, D.A.; Macleod, M.R.; Coutts, S.B.; Schwamm, L.H.; Davis, S.M.; Donnan, G.A. Ischaemic stroke. Nat. Rev. Dis. Primers 2019, 5, 70. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Wang, L.; Wang, L.P.; Wan, Q. Therapeutic targets of neuroprotection and neurorestoration in ischemic stroke: Applications for natural compounds from medicinal herbs. Biomed. Pharmacother. 2022, 148, 112719. [Google Scholar] [CrossRef]
- Dai, J.; Qiu, Y.M.; Ma, Z.W.; Yan, G.F.; Zhou, J.; Li, S.Q.; Wu, H.; Jin, Y.C.; Zhang, X.H. Neuroprotective effect of baicalin on focal cerebral ischemia in rats. Neural Regen. Res. 2018, 13, 2129–2133. [Google Scholar] [PubMed]
- Tang, W.; Sun, X.; Fang, J.S.; Zhang, M.; Sucher, N.J. Flavonoids from Radix Scutellariae as potential stroke therapeutic agents by targeting the second postsynaptic density 95 (PSD-95)/disc large/zonula occludens-1 (PDZ) domain of PSD-95. Phytomedicine 2004, 11, 277–284. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Wang, Z.; Zhang, X.Y.; Ying, K.; Liu, J.X.; Wang, Y.Y. Gene expression profile induced by oral administration of baicalin and gardenin after focal brain ischemia in rats. Acta Pharmacol. Sin. 2005, 26, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.Q.; Liu, Q.; Xu, W.J.; Yu, Y.N.; Zhang, Y.Y.; Li, B.; Liu, J.; Wang, Z. Pure mechanistic analysis of additive neuroprotective effects between baicalin and jasminoidin in ischemic stroke mice. Acta Pharmacol. Sin. 2018, 39, 961–974. [Google Scholar] [CrossRef]
- Kostandy, B.B. The role of glutamate in neuronal ischemic injury: The role of spark in fire. Neurol. Sci. 2012, 33, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Shigeri, Y.; Seal, R.P.; Shimamoto, K. Molecular pharmacology of glutamate transporters, EAATs and VGLUTs. Brain Res. Brain Res. Rev. 2004, 45, 250–265. [Google Scholar] [CrossRef]
- Zhou, Z.Q.; Li, Y.L.; Ao, Z.B.; Wen, Z.L.; Chen, Q.W.; Huang, Z.G.; Xiao, B.; Yan, X.H. Baicalin protects neonatal rat brains against hypoxic-ischemic injury by upregulating glutamate transporter 1 via the phosphoinositide 3-kinase/protein kinase B signaling pathway. Neural Regen. Res. 2017, 12, 1625–1631. [Google Scholar]
- Song, X.; Gong, Z.; Liu, K.; Kou, J.; Liu, B.; Liu, K. Baicalin combats glutamate excitotoxicity via protecting glutamine synthetase from ROS-induced 20S proteasomal degradation. Redox Biol. 2020, 34, 101559. [Google Scholar] [CrossRef]
- Ge, Q.F.; Hu, X.; Ma, Z.Q.; Liu, J.R.; Zhang, W.P.; Chen, Z.; Wei, E.Q. Baicalin attenuates oxygen-glucose deprivation-induced injury via inhibiting NMDA receptor-mediated 5-lipoxygenase activation in rat cortical neurons. Pharmacol. Res. 2007, 55, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Li, C.T.; Zhang, W.P.; Fang, S.H.; Lu, Y.B.; Zhang, L.H.; Qi, L.L.; Huang, X.Q.; Huang, X.J.; Wei, E.Q. Baicalin attenuates oxygen-glucose deprivation-induced injury by inhibiting oxidative stress-mediated 5-lipoxygenase activation in PC12 cells. Acta Pharmacol. Sin. 2010, 31, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Tu, X.K.; Yang, W.Z.; Shi, S.S.; Wang, C.H.; Chen, C.M. Neuroprotective effect of baicalin in a rat model of permanent focal cerebral ischemia. Neurochem. Res. 2009, 34, 1626–1634. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Yuan, Z.Y.; Wang, Y.G.; Wan, H.J.; Hu, J.; Chai, Y.S.; Lei, F.; Xing, D.M.; Du, L.J. Role of baicalin in regulating Toll-like receptor 2/4 after ischemic neuronal injury. Chin. Med. J. 2012, 125, 1586–1593. [Google Scholar]
- Chen, H.; Guan, B.; Chen, X.; Chen, X.; Li, C.; Qiu, J.; Yang, D.; Liu, K.J.; Qi, S.; Shen, J. Baicalin Attenuates Blood-Brain Barrier Disruption and Hemorrhagic Transformation and Improves Neurological Outcome in Ischemic Stroke Rats with Delayed t-PA Treatment: Involvement of ONOO(-)-MMP-9 Pathway. Transl. Stroke Res. 2018, 9, 515–529. [Google Scholar] [CrossRef]
- Chen, H.S.; Chen, X.; Li, W.T.; Shen, J.G. Targeting RNS/caveolin-1/MMP signaling cascades to protect against cerebral ischemia-reperfusion injuries: Potential application for drug discovery. Acta Pharmacol. Sin. 2018, 39, 669–682. [Google Scholar] [CrossRef]
- Wareham, L.K.; Liddelow, S.A.; Temple, S.; Benowitz, L.I.; Di Polo, A.; Wellington, C.; Goldberg, J.L.; He, Z.; Duan, X.; Bu, G.; et al. Solving neurodegeneration: Common mechanisms and strategies for new treatments. Mol. Neurodegener. 2022, 17, 23. [Google Scholar] [CrossRef]
- Guo, C.; Chen, X.; Xiong, P. Baicalin suppresses iron accumulation after substantia nigra injury: Relationship between iron concentration and transferrin expression. Neural Regen. Res. 2014, 9, 630–636. [Google Scholar]
- Kumar, H.; Ropper, A.E.; Lee, S.H.; Han, I. Propitious Therapeutic Modulators to Prevent Blood-Spinal Cord Barrier Disruption in Spinal Cord Injury. Mol. Neurobiol. 2017, 54, 3578–3590. [Google Scholar] [CrossRef]
- Herrman, H.; Kieling, C.; Mcgorry, P.; Horton, R.; Sargent, J.; Patel, V. Reducing the global burden of depression: A Lancet-World Psychiatric Association Commission. Lancet 2019, 393, e42–e43. [Google Scholar] [CrossRef]
- Marwaha, S.; Palmer, E.; Suppes, T.; Cons, E.; Young, A.H.; Upthegrove, R. Novel and emerging treatments for major depression. Lancet 2023, 401, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Xu, J.; Zou, D.; Yang, L.; Wang, Y. Baicalin influences the dendritic morphology of newborn neurons in the hippocampus of chronically stressed rats. Neural Regen. Res. 2013, 8, 496–505. [Google Scholar] [PubMed]
- Yu, H.Y.; Yin, Z.J.; Yang, S.J.; Ma, S.P.; Qu, R. Baicalin Reverses Depressive-Like Behaviours and Regulates Apoptotic Signalling Induced by Olfactory Bulbectomy. Phytother. Res. 2016, 30, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Ma, Z.; Liu, K.; Li, Y.; Liu, D.; Xu, L.; Deng, X.; Qu, R.; Ma, Z.; Ma, S. Baicalin exerts antidepressant effects through Akt/FOXG1 pathway promoting neuronal differentiation and survival. Life Sci. 2019, 221, 241–248. [Google Scholar] [CrossRef]
- Lu, Y.; Sun, G.; Yang, F.; Guan, Z.; Zhang, Z.; Zhao, J.; Liu, Y.; Chu, L.; Pei, L. Baicalin regulates depression behavior in mice exposed to chronic mild stress via the Rac/LIMK/cofilin pathway. Biomed. Pharmacother. 2019, 116, 109054. [Google Scholar] [CrossRef]
- Guo, L.T.; Wang, S.Q.; Su, J.; Xu, L.X.; Ji, Z.Y.; Zhang, R.Y.; Zhao, Q.W.; Ma, Z.Q.; Deng, X.Y.; Ma, S.P. Baicalin ameliorates neuroinflammation-induced depressive-like behavior through inhibition of toll-like receptor 4 expression via the PI3K/AKT/FoxO1 pathway. J. Neuroinflam. 2019, 16, 95. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, F.; Guan, X. Baicalin reverse depressive-like behaviors through regulation SIRT1-NF-kB signaling pathway in olfactory bulbectomized rats. Phytother. Res. 2019, 33, 1480–1489. [Google Scholar] [CrossRef]
- Zhou, R.; Wang, J.; Han, X.; Ma, B.; Yuan, H.; Song, Y. Baicalin regulates the dopamine system to control the core symptoms of ADHD. Mol. Brain 2019, 12, 11. [Google Scholar] [CrossRef]
- Wu, J.; Cao, M.; Peng, Y.; Dong, B.; Jiang, Y.; Hu, C.; Zhu, P.; Xing, W.; Yu, L.; Xu, R.; et al. Research progress on the treatment of epilepsy with traditional Chinese medicine. Phytomedicine 2023, 120, 155022. [Google Scholar] [CrossRef]
- Gao, Z.; Huang, K.; Xu, H. Protective effects of flavonoids in the roots of Scutellaria baicalensis Georgi against hydrogen peroxide-induced oxidative stress in HS-SY5Y cells. Pharmacol. Res. 2001, 43, 173–178. [Google Scholar] [CrossRef]
- Chew, N.W.S.; Ng, C.H.; Tan, D.J.H.; Kong, G.; Lin, C.; Chin, Y.H.; Lim, W.H.; Huang, D.Q.; Quek, J.; Fu, C.E.; et al. The global burden of metabolic disease: Data from 2000 to 2019. Cell Metab. 2023, 35, 414–428.e3. [Google Scholar] [CrossRef]
- Fang, P.; Sun, Y.; Gu, X.; Shi, M.; Bo, P.; Zhang, Z.; Bu, L. Baicalin ameliorates hepatic insulin resistance and gluconeogenic activity through inhibition of p38 MAPK/PGC-1α pathway. Phytomedicine 2019, 64, 153074. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Y.; Lou, M.; Xia, W.; Liu, Q.; Xie, G.; Liu, L.; Liu, B.; Yang, J.; Qin, M. Baicalin regulates SirT1/STAT3 pathway and restrains excessive hepatic glucose production. Pharmacol. Res. 2018, 136, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Han, S.; Wang, M.; Han, L.; Huang, Y.; Bo, P.; Fang, P.; Zhang, Z. Baicalin protects against insulin resistance and metabolic dysfunction through activation of GALR2/GLUT4 signaling. Phytomedicine 2022, 95, 153869. [Google Scholar] [CrossRef]
- Szkudelski, T.; Szkudelska, K. The Anti-Diabetic Potential of Baicalin: Evidence from Rodent Studies. Int. J. Mol. Sci. 2023, 25, 431. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- Pal, M.; Febbraio, M.A.; Lancaster, G.I. The roles of c-Jun NH2-terminal kinases (JNKs) in obesity and insulin resistance. J. Physiol. 2016, 594, 267–279. [Google Scholar] [CrossRef]
- Baradaran Rahimi, V.; Askari, V.R.; Hosseinzadeh, H. Promising influences of Scutellaria baicalensis and its two active constituents, baicalin, and baicalein, against metabolic syndrome: A review. Phytother. Res. 2021, 35, 3558–3574. [Google Scholar] [CrossRef]
- Fan, L.; Zhang, W.; Guo, D.; Tan, Z.R.; Xu, P.; Li, Q.; Liu, Y.Z.; Zhang, L.; He, T.Y.; Hu, D.L.; et al. The effect of herbal medicine baicalin on pharmacokinetics of rosuvastatin, substrate of organic anion-transporting polypeptide 1B1. Clin. Pharmacol. Ther. 2008, 83, 471–476. [Google Scholar] [CrossRef]
- Li, J.Z.; Chen, N.; Ma, N.; Li, M.R. Mechanism and Progress of Natural Products in the Treatment of NAFLD-Related Fibrosis. Molecules 2023, 28, 7936. [Google Scholar] [CrossRef]
- Li, H.T.; Wu, X.D.; Davey, A.K.; Wang, J. Antihyperglycemic effects of baicalin on streptozotocin—Nicotinamide induced diabetic rats. Phytother. Res. 2011, 25, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jiang, H.; Cao, S.; Chen, Q.; Cui, M.; Wang, Z.; Li, D.; Zhou, J.; Wang, T.; Qiu, F.; et al. Baicalin and its metabolites suppresses gluconeogenesis through activation of AMPK or AKT in insulin resistant HepG-2 cells. Eur. J. Med. Chem. 2017, 141, 92–100. [Google Scholar] [CrossRef]
- Yang, L.L.; Xiao, N.; Liu, J.; Liu, K.; Liu, B.; Li, P.; Qi, L.W. Differential regulation of baicalin and scutellarin on AMPK and Akt in promoting adipose cell glucose disposal. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Yu, M.; Min, W.; Wan, D.; Han, S.; Shan, Y.; Wang, R.; Shi, M.; Zhang, Z.; Bo, P. Effect of baicalin on GLUT4 expression and glucose uptake in myotubes of rats. Life Sci. 2018, 196, 156–161. [Google Scholar] [CrossRef]
- Miao, L.; Zhang, X.; Zhang, H.; Cheong, M.S.; Chen, X.; Farag, M.A.; Cheang, W.S.; Xiao, J. Baicalin ameliorates insulin resistance and regulates hepatic glucose metabolism via activating insulin signaling pathway in obese pre-diabetic mice. Phytomedicine 2023, 124, 155296. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. The pathobiology of diabetic complications—A unified mechanism. Diabetes 2005, 54, 1615–1625. [Google Scholar] [CrossRef]
- Waisundara, V.Y.; Siu, S.Y.; Hsu, A.; Huang, D.; Tan, B.K. Baicalin upregulates the genetic expression of antioxidant enzymes in Type-2 diabetic Goto-Kakizaki rats. Life Sci. 2011, 88, 1016–1025. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, C.; Liu, W.; Chen, J.; Sun, Y.; Chang, D.; Wang, H.; Xu, W.; Lu, J.J.; Zhou, X.; et al. Upregulation of Nrf2 signaling: A key molecular mechanism of Baicalin’s neuroprotective action against diabetes-induced cognitive impairment. Biomed. Pharmacother. 2024, 174, 116579. [Google Scholar] [CrossRef]
- Jiang, H.; Yao, Q.; An, Y.; Fan, L.; Wang, J.; Li, H. Baicalin suppresses the progression of Type 2 diabetes-induced liver tumor through regulating METTL3/m6A/HKDC1 axis and downstream p-JAK2/STAT1/clevaged Capase3 pathway. Phytomedicine 2022, 94, 153823. [Google Scholar] [CrossRef]
- Qin, W.; Shang, Q.; Shen, G.; Li, B.; Zhang, P.; Zhang, Y.; Zhao, W.; Chen, H.; Liu, H.; Xie, B.; et al. Restoring bone-fat equilibrium: Baicalin’s impact on P38 MAPK pathway for treating diabetic osteoporosis. Biomed. Pharmacother. 2024, 175, 116571. [Google Scholar] [CrossRef]
- Yang, R.; Wang, R.; Xu, A.; Zhang, J.; Ma, J. Mitigating neurodegenerative diseases: The protective influence of baicalin and baicalein through neuroinflammation regulation. Front. Pharmacol. 2024, 15, 1425731. [Google Scholar] [CrossRef] [PubMed]
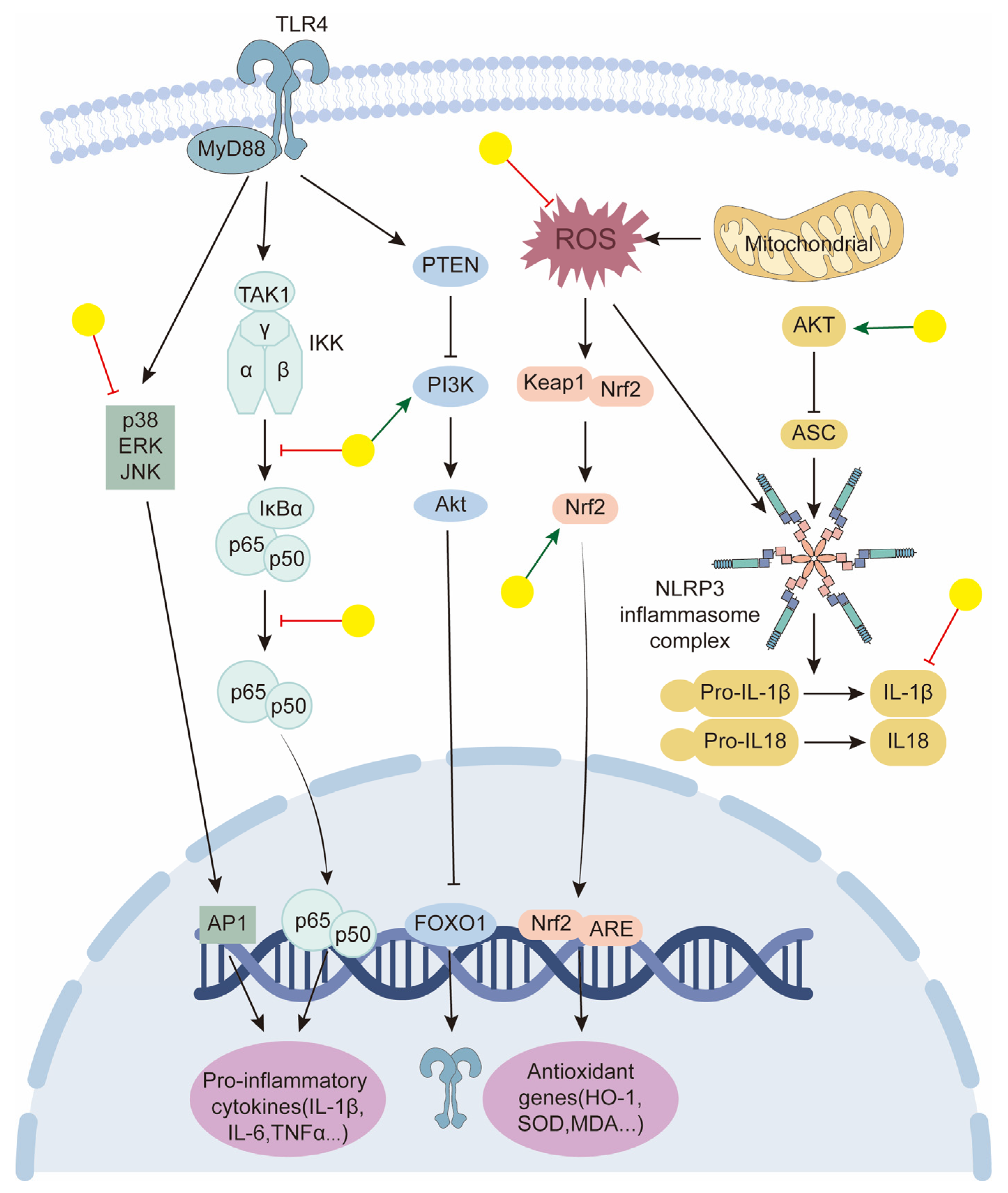
- Felix, F.B.; Vago, J.P.; Beltrami, V.A.; Araújo, J.M.D.; Grespan, R.; Teixeira, M.M.; Pinho, V. Biochanin A as a modulator of the inflammatory response: An updated overview and therapeutic potential. Pharmacol. Res. 2022, 180, 106246. [Google Scholar] [CrossRef]
- Liao, H.; Ye, J.; Gao, L.; Liu, Y. The main bioactive compounds of Scutellaria baicalensis Georgi. for alleviation of inflammatory cytokines: A comprehensive review. Biomed. Pharmacother. 2021, 133, 110917. [Google Scholar] [CrossRef] [PubMed]
- Živanović, N.; Lesjak, M.; Simin, N.; Srai, S.K.S. Beyond Mortality: Exploring the Influence of Plant Phenolics on Modulating Ferroptosis-A Systematic Review. Antioxidants 2024, 13, 334. [Google Scholar] [CrossRef]
- Gao, Z.; Huang, K.; Yang, X.; Xu, H. Free radical scavenging and antioxidant activities of flavonoids extracted from the radix of Scutellaria baicalensis Georgi. Biochim Biophys Acta 1999, 1472, 643–650. [Google Scholar] [CrossRef]
- Ng, T.B.; Liu, F.; Wang, Z.T. Antioxidative activity of natural products from plants. Life Sci. 2000, 66, 709–723. [Google Scholar] [CrossRef]
- Chen, J.W.; Zhu, Z.Q.; Hu, T.X.; Zhu, D.Y. Structure-activity relationship of natural flavonoids in hydroxyl radical-scavenging effects. Acta Pharmacol. Sin. 2002, 23, 667–672. [Google Scholar] [PubMed]
- Altavilla, D.; Squadrito, F.; Bitto, A.; Polito, F.; Burnett, B.P.; Di Stefano, V.; Minutoli, L. Flavocoxid, a dual inhibitor of cyclooxygenase and 5-lipoxygenase, blunts pro-inflammatory phenotype activation in endotoxin-stimulated macrophages. Br. J. Pharmacol. 2009, 157, 1410–1418. [Google Scholar] [CrossRef]
- Dai, S.X.; Zou, Y.; Feng, Y.L.; Liu, H.B.; Zheng, X.B. Baicalin down-regulates the expression of macrophage migration inhibitory factor (MIF) effectively for rats with ulcerative colitis. Phytother. Res. 2012, 26, 498–504. [Google Scholar] [CrossRef]
- An, H.J.; Lee, J.Y.; Park, W. Baicalin Modulates Inflammatory Response of Macrophages Activated by LPS via Calcium-CHOP Pathway. Cells 2022, 11, 3076. [Google Scholar] [CrossRef]
- Zou, Y.; Dai, S.X.; Chi, H.G.; Li, T.; He, Z.W.; Wang, J.; Ye, C.G.; Huang, G.L.; Zhao, B.; Li, W.Y.; et al. Baicalin attenuates TNBS-induced colitis in rats by modulating the Th17/Treg paradigm. Arch. Pharm. Res. 2015, 38, 1873–1887. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Xiao, Z.; Yan, H.; Lu, X.; Zhang, X.; Luo, L.; Long, C.; Zhu, Y. Baicalin suppresses Th1 and Th17 responses and promotes Treg response to ameliorate sepsis-associated pancreatic injury via the RhoA-ROCK pathway. Int. Immunopharmacol. 2020, 86, 106685. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Qu, F.; Burrows, E.; Yu, Y.; Nan, R. Acupuncture can improve absorption of baicalin from extracts of Scutellaria baicalensis Georgi in rats. Phytother. Res. 2009, 23, 1415–1420. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, X.; Li, J.; Liu, R.; Shu, L.; Jin, J. Effects of Labrasol on the corneal drug delivery of baicalin. Drug Deliv. 2009, 16, 399–404. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, D.; Liu, H.; Yang, Q.; Yao, K.; Wang, X.; Wang, L.; Yang, X. Effect of Low Molecular Weight Chitosans on Drug Permeation through Mouse Skin: 1. Transdermal Delivery of Baicalin. J. Pharm. Sci. 2010, 99, 2991–2998. [Google Scholar] [CrossRef]
- Chalasani, N.; Vuppalanchi, R.; Navarro, V.; Fontana, R.; Bonkovsky, H.; Barnhart, H.; Kleiner, D.E.; Hoofnagle, J.H. Acute liver injury due to flavocoxid (Limbrel), a medical food for osteoarthritis: A case series. Ann. Intern. Med. 2012, 156, 857–860. [Google Scholar] [CrossRef]
- Kim, M.S.; Ham, S.H.; Kim, J.H.; Shin, J.E.; Oh, J.; Kim, T.W.; Yun, H.I.; Lim, J.H.; Jang, B.S.; Cho, J.H. Single-dose oral toxicity of fermented Scutellariae Radix extract in rats and dogs. Toxicol. Res. 2012, 28, 263–268. [Google Scholar] [CrossRef]
- Hwang, J.M.; Wang, C.J.; Chou, F.P.; Tseng, T.H.; Hsieh, Y.S.; Hsu, J.D.; Chu, C.Y. Protective effect of baicalin on tert-butyl hydroperoxide-induced rat hepatotoxicity. Arch. Toxicol. 2005, 79, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.L.; Gong, L.K.; Wang, H.; Xiao, Y.; Wu, X.F.; Zhang, Y.H.; Xue, X.; Qi, X.M.; Ren, J. Baicalin protects mouse from Concanavalin A-induced liver injury through inhibition of cytokine production and hepatocyte apoptosis. Liver Int. 2007, 27, 582–591. [Google Scholar] [CrossRef]
- Shi, L.; Zhang, S.; Huang, Z.; Hu, F.; Zhang, T.; Wei, M.; Bai, Q.; Lu, B.; Ji, L. Baicalin promotes liver regeneration after acetaminophen-induced liver injury by inducing NLRP3 inflammasome activation. Free Radic. Biol. Med. 2020, 160, 163–177. [Google Scholar] [CrossRef]
- Yang, M.D.; Chiang, Y.M.; Higashiyama, R.; Asahina, K.; Mann, D.A.; Mann, J.; Wang, C.C.; Tsukamoto, H. Rosmarinic acid and baicalin epigenetically derepress peroxisomal proliferator-activated receptor γ in hepatic stellate cells for their antifibrotic effect. Hepatology 2012, 55, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Yin, X.; Yuan, J.; Liang, Z.; Song, J.; Li, Y.; Peng, C.; Hylands, P.J.; Zhao, Z.; Xu, Q. Antifibrotic activities of Scutellariae Radix extracts and flavonoids: Comparative proteomics reveals distinct and shared mechanisms. Phytomedicine 2022, 100, 154049. [Google Scholar] [CrossRef] [PubMed]
- Zaghloul, R.A.; Zaghloul, A.M.; El-Kashef, D.H. Hepatoprotective effect of Baicalin against thioacetamide-induced cirrhosis in rats: Targeting NOX4/NF-κB/NLRP3 inflammasome signaling pathways. Life Sci. 2022, 295, 120410. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Active Components or Major Compounds | Application | Reference |
|---|---|---|---|
| Hange-Shashin-To | Baicalin, glycyrrhizin, isoliquiritin, berberine, coptisine, palmitine, and saponins | Diarrhoeal | [2] |
| Soshiho-tang | Homogentisic acid, baicalin, glycyrrhizin, saikosaponin A, 6-gingerol, and ginsenoside Rg3 | Chronic liver disease | [3] |
| Angong Niuhuang sticker | Curcuma, berberine hydrochloride, baicalin, geniposide, borneol, and musk | Cerebral ischemia | [4] |
| Huang-Lian-Jie-Du-Tang | Berberine, palmatine, baicalin, baicalein, and gardenoside | Cerebrovascular disease | [5] |
| Huaijiao pill | Ophoricoside, baicalin, naringin, genistein, rutin, quercetin, and 5-O-methylvisammioside | Hematochezia, edema, and carbuncle | [6] |
| Gegen Qinlian decoction | Puerarin, liquiritin, berberine, and baicalin | Diarrhea and inflammation symptoms | [7] |
| Shuanghuanglian preparation | Chlorogenic acid, phillyrin, baicalin, and baicalein | Respiratory tract infection | [8] |
| Chaiqin chengqi decoction | Emodin, baicalin, rhein, and chrysin | Acute pancreatitis | [9] |
| Qingda granules | Baicalin | Spontaneous hypertension | [10] |
| Jinzhen granules | Gallic acid, baicalin, glycyrrhizic acid, hyodeoxycholic acid, and cholic acid | Viral-induced diseases | [11] |
| Liang-Ge-San | Geniposide, liquiritin, forsythenside A, forsythin, baicalin, baicalein, rhein, and emodin | Virus-induced diseases | [12] |
| Xiaoer Chaige Tuire oral liquid | Puerarin, daidzein, benzoic acid, baicalin, baicalein, wogonoside, wogonin, oroxylin A, 3′-methoxypuerarin, paeoniflorin, scopoletin, and liquiritigenin | Anti-inflammation and antivirus | [13] |
| Lanqin oral solution | Geniposide, berberine, palmatine, and baicalin | Pharyngitis | [14] |
| Qingkailing | Hyodeoxycholic acid, geniposide, baicalin, and cholic acid | Ischemic stroke | [15] |
| Bu-Shen-Ning-Xin decoction | Berberine, paeoniflorin, morroniside, gallic acid, loganin, and baicalin | Premature ovarian insufficiency | [16] |
| Wenqingyin | Baicalin, coptisine, and paeoniflorin | Sepsis-induced acute lung injury | [17] |
| Qing-Yi recipe | Baicalin, wogonoside, geniposide, rhein, costunolide, and paeoniflorin | Acute diseases of the abdomen | [18] |
| Sanfeng Tongqiao dripping pills | L-Menthone, pulegone, hesperetin, baicalin, wogonin, pulegone, and luteolin | Allergic rhinitis | [19] |
| Disease | Biological Effects | Mechanisms of Action | Reference | |
|---|---|---|---|---|
| Cancer | ||||
| Lymphoma | Induces apoptosis | ↓ PI3K/Akt pathway | [32] | |
| Hepatocellular carcinoma | Repolarization of TAM toward the M1 phenotype | ↑ RelB/p52 pathway | [33] | |
| Colon cancer | Induces senescence | ↑ DEPP and Ras/Raf/MEK/ ERK signaling | [34] | |
| Leukemia | Enhances apoptosis and reduces viability | ↓ Akt pathway | [35] | |
| Bladder cancer | Induces ferroptosis | ↓ FTH1 | [36] | |
| Non-small cell lung cancer | Promotes macrophage polarization to the M1 phenotype | ↑ JAK2-STAT1 pathway in macrophages | [37] | |
| Breast cancer | Triggers apoptosis and reduces inflammation and angiogenesis | ↓ NF-κB, Bcl-2, VEGF ↑ p53, Bax, and caspase-3 | [38] | |
| Colorectal cancer | Induces apoptosis | ↓ CDK/RB | [39] | |
| Hepatocellular carcinoma | Inhibits proliferation, migration, and invasion and induces cell cycle arrest and apoptosis | ↓ ROCK1 signaling | [40] | |
| Osteosarcomas | Suppresses cell proliferation and induces apoptosis and ferroptosis | ↓ Nrf2/ xCT/GPX4 regulatory axis | [41] | |
| Gastric cancer | Promotes ferroptosis | ↑ ROS | [42] | |
| Cardiovascular diseases | ||||
| Atherosclerosis (vascular inflammatory disorders) | Promotes the efflux of cholesterol from macrophages and delays the formation of foam cells | ↑ PPARγ-ABCA1/ABCG1 pathway | [43] | |
| Angiogenesis | - | ↑ ERRα pathway. | [44] | |
| Cardiac hypertrophy and heart failure | - | ↑ SIRT3/LKB1/AMPK signaling pathway | [45] | |
| PAH | - | ↑ A (2A) R activity ↓ PI3K/AKT signaling | [46] | |
| Hypertension | Reduces constriction and enhances vasodilation of abdominal aortic rings | ↓ MLCK/p-MLC pathway | [47] | |
| Neurological diseases | ||||
| Parkinson’s disease | Protects dopaminergic neurons | ↓ Iron accumulation | [48] | |
| Alzheimer’s disease | - | ↓ COX1-2/5-LOX | [49] | |
| Demyelinating diseases | Promotes myelin production and regeneration | ↑ PPARγ signaling pathway | [50] | |
| Spinal cord injury | - | ↑ PI3K/Akt | [51] | |
| Metabolic diseases | ||||
| Obesity | Modulates the expression of genes in the adipogenesis pathway | ↑ Antiadipogenic regulators, including KLF2, C/EBPγ, and CHOP ↓ The proadipogenic regulator KLF15 | [52] | |
| Hepatic steatosis | Decreases serum cholesterol, free fatty acid, and insulinconcentrations↓ Systemic inflammation | ↑ AMPK | [53] | |
| Diet-induced obesity and hepatic steatosis | Antisteatosis | ↑ CPT1 | [54] | |
| Non-alcoholic steatohepatitis | Decreases lipid accumulation | ↓ SREBP-1c and fatty acid synthase ↑ Fatty acid oxidation enzymes, includingPPARα and CPT1a | [55] | |
| NAFLD | Decreases lipid accumulation | ↑ AMPK and Nrf2 ↓ SREBP1 and NF-κB | [56] | |
| MAFLD | Oxidative stress and inflammation | ↑ p62-Keap1-Nrf2 signaling cascade | [57] | |
| Diabetic nephropathy | Anti-inflammatory effects | ↓ IκB and JAK2 phosphorylation | [58] | |
| Diabetic kidney disease | Ameliorates renal fibrosis | ↑ CPT1α | [59] | |
| Inflammatory diseases | ||||
| Acute pancreatitis (emodinand baicalin) | Anti-inflammatory effects | ↓ Serum TNF-a and IL-6 ↓ TLR4 | [60] | |
| Ulcerative colitis | - | |||
| Anti-asthmatic effects | - | ↓ Th17 cells | [61] | |
| Colitis | Reduces inflammatory mediators | ↓ Th17 ↑ Treg cells | [62] | |
| Asthma | Reduces inflammatory cell infiltration | ↓ Phosphodiesterase 4 (PDE4) | [63] | |
| Allergic rhinitis | Improves allergic rhinitis symptoms and | ↓ JAK2-STAT5 and NF-κB signaling | [64] | |
| Chronic ulcerative colitis | Reduces MPO, NO, and inflammatory cytokine levels | ↓ IL-33 expression ↓ NF-κB | [65] | |
| OA | Alleviates inflammatory injury, increases cell viability, and decreases cell apoptosis | ↓ miR-126↓NF-κB | [66] | |
| Chronic gastritis | Reduces IL-8, IL-1β, TNF-α, PGE2, NO, and ET-1 levels | ↓ Akt/NF-κB | [67] | |
| Lupus | Reduces urine protein levels and ameliorates lupus nephritis | ↓ mTOR activation ↓ differentiation of Tfh cells ↑ Expansion of Tfr cells | [68] | |
| Psoriasis | Decreases the level of inflammatory factors and inhibits Th1/Th17 cell differentiation | ↑ PPARγ ↓ Wnt signaling pathway and Th17/IL-17 axis | [69] | |
| Oral mucositis | Reduces inflammatory storm | ↓ oxidative stress and NLRP3 | [70] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, X.; Huang, R.; Xie, J.; Luo, S.; Cheng, X.; Cui, J.; Hu, D. Recent Advances in the Therapeutic Effects and Molecular Mechanisms of Baicalin. Biology 2025, 14, 637. https://doi.org/10.3390/biology14060637
Qiu X, Huang R, Xie J, Luo S, Cheng X, Cui J, Hu D. Recent Advances in the Therapeutic Effects and Molecular Mechanisms of Baicalin. Biology. 2025; 14(6):637. https://doi.org/10.3390/biology14060637
Chicago/Turabian StyleQiu, Xiaoyuan, Renyin Huang, Junke Xie, Shanshan Luo, Xiang Cheng, Jing Cui, and Desheng Hu. 2025. "Recent Advances in the Therapeutic Effects and Molecular Mechanisms of Baicalin" Biology 14, no. 6: 637. https://doi.org/10.3390/biology14060637
APA StyleQiu, X., Huang, R., Xie, J., Luo, S., Cheng, X., Cui, J., & Hu, D. (2025). Recent Advances in the Therapeutic Effects and Molecular Mechanisms of Baicalin. Biology, 14(6), 637. https://doi.org/10.3390/biology14060637