
Neuroprotective, Antioxidant and Anti-Inflammatory Effect of Greek Pomegranate Seed Oil on N2a Neuroblastoma Cells and Mild Cognitive Impairment Patients

 , , ,
, , ,  and
and
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Pomegranate Seed Oil
2.2. Cell Culture
2.2.1. MTT Viability Assay
2.2.2. Assessment of the Antioxidant Potential of PSO with MDA Determination
2.2.3. Treatment of LPS-Induced N2a Cells with PSO
2.2.4. Western Blotting Analysis
2.2.5. Statistical Analysis
2.3. Evaluation of the Effect of PSO Consumption on MCI Patients
2.3.1. Subjects
2.3.2. Study Design
2.3.3. Processing of Blood Serum Samples
2.3.4. Analysis of Biomarkers Levels by ELISA
2.3.5. Statistical Analysis for PSO Effect on MCI Patients
3. Results
3.1. The Βeneficical Effect οf PSO on LPS-Stimulated N2a Murine Neuroblastoma Cells
3.1.1. Tolerability of N2a Cells to PSO
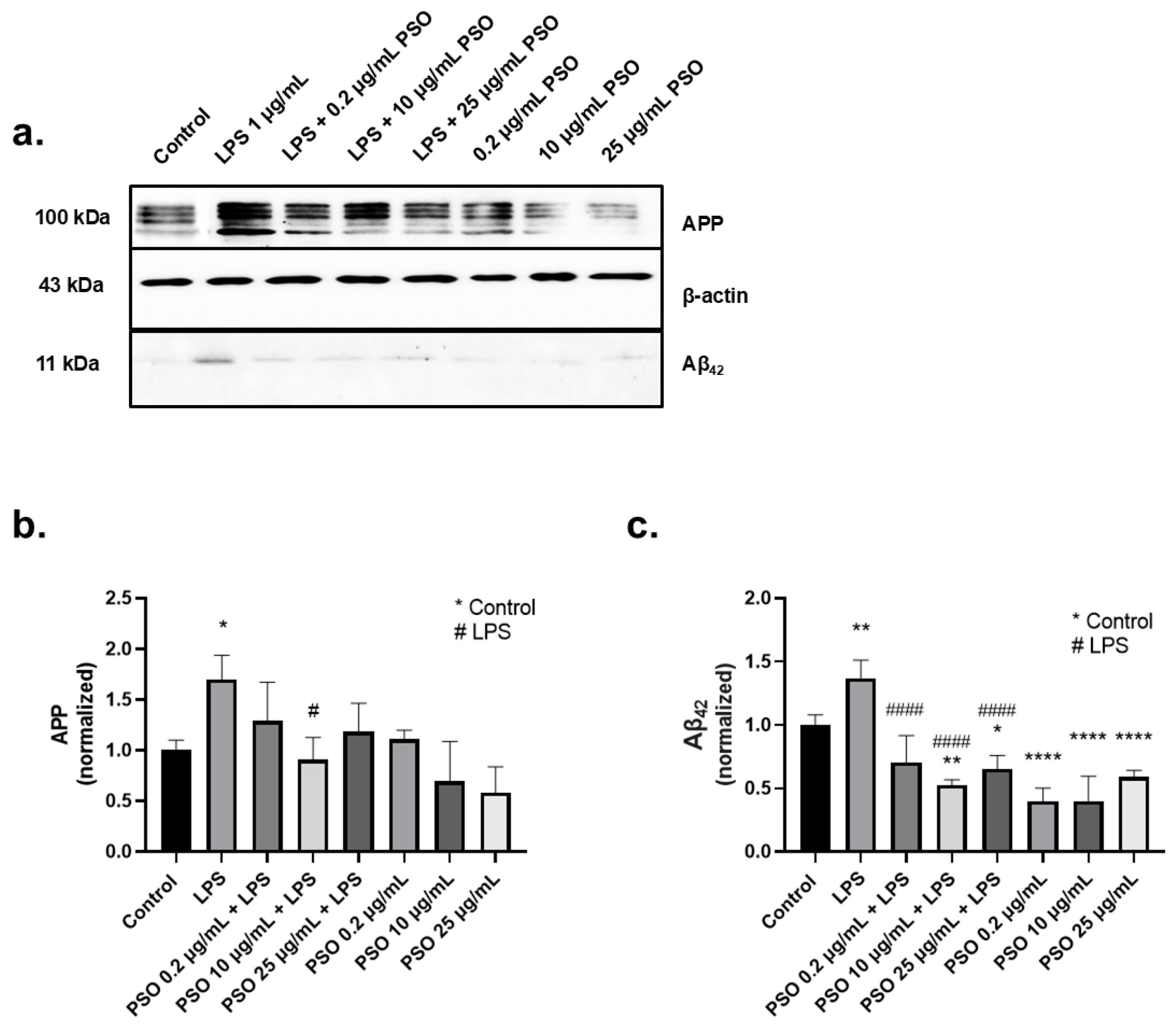
3.1.2. PSO Treatment Attenuated the Overexpression of Amyloid Triggered by LPS
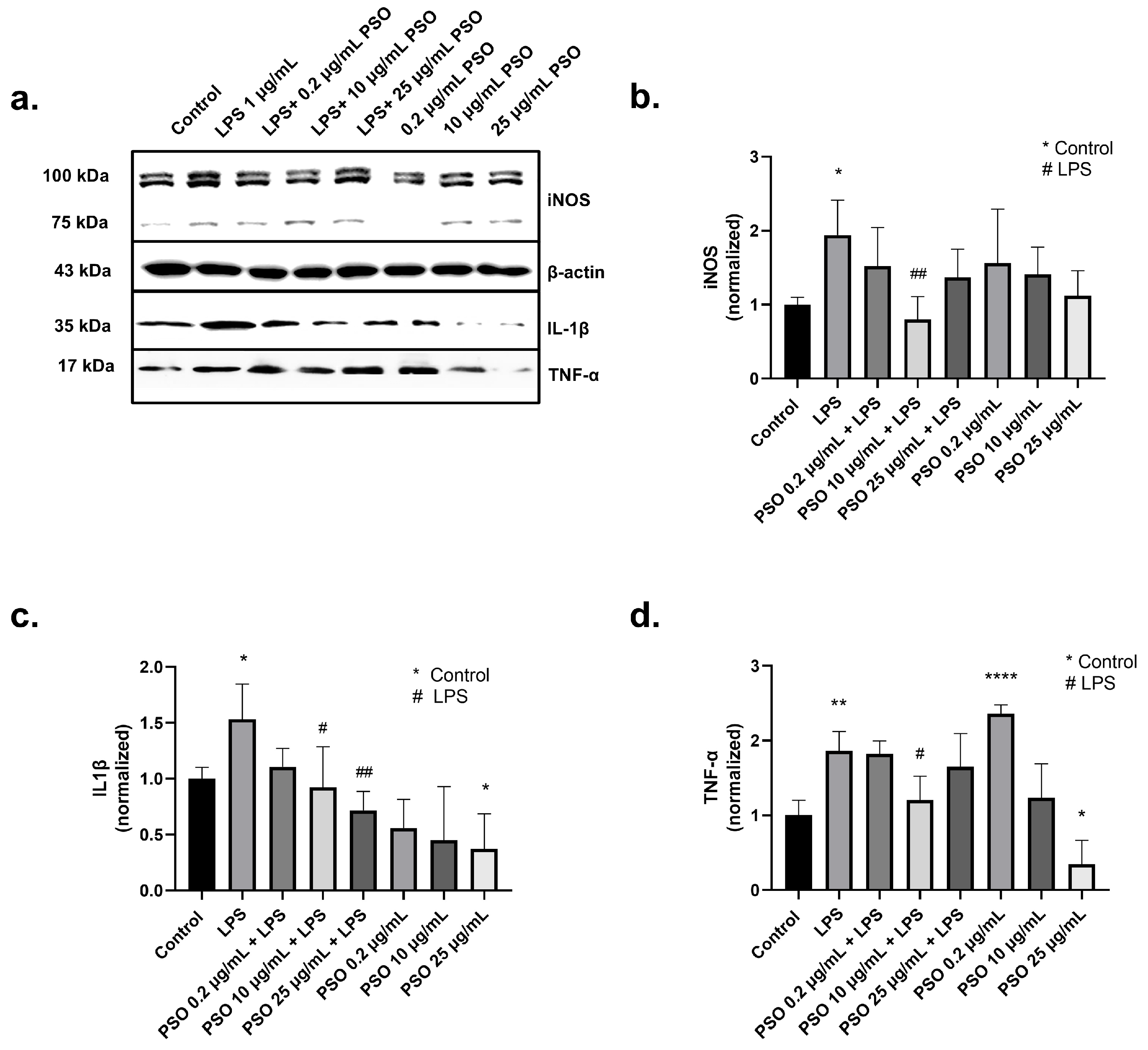
3.1.3. PSO Mediates a Potent Anti-Inflammatory Effect on N2a Cells
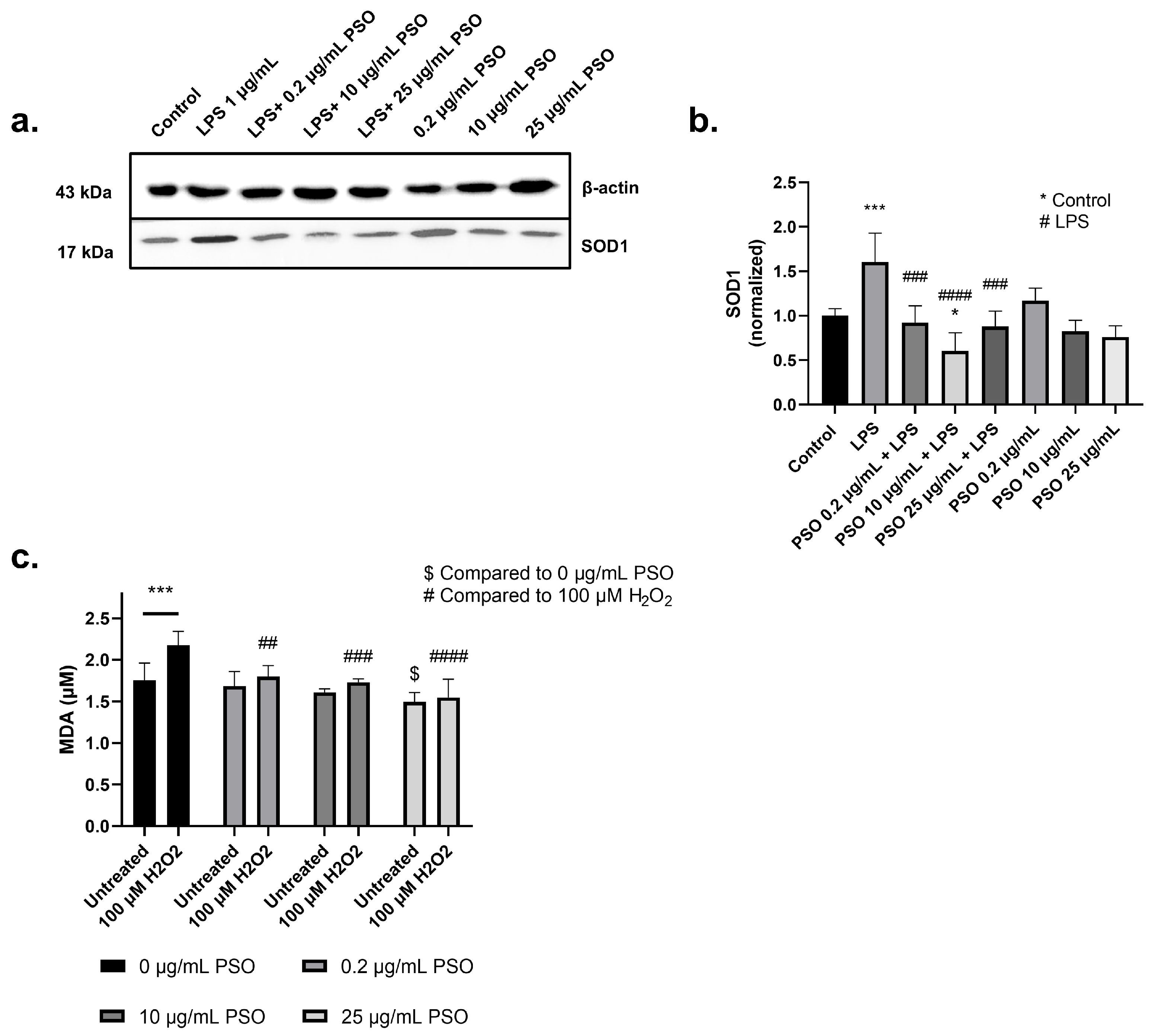
3.1.4. PSO Exerts an Antioxidant Activity in N2a Cells
3.2. The Impact of Pomegranate Seed Oil on MCI Patients: A Clinical Evaluation
3.2.1. Statistical Analysis of Subjects’ Demographics
3.2.2. Consumption of PSO for 12 Months Improved the Ratio of Aβ42/Aβ40
3.2.3. The Consumption of PSO for 12 Months Reduced p-tau181 Levels in MCI Patients
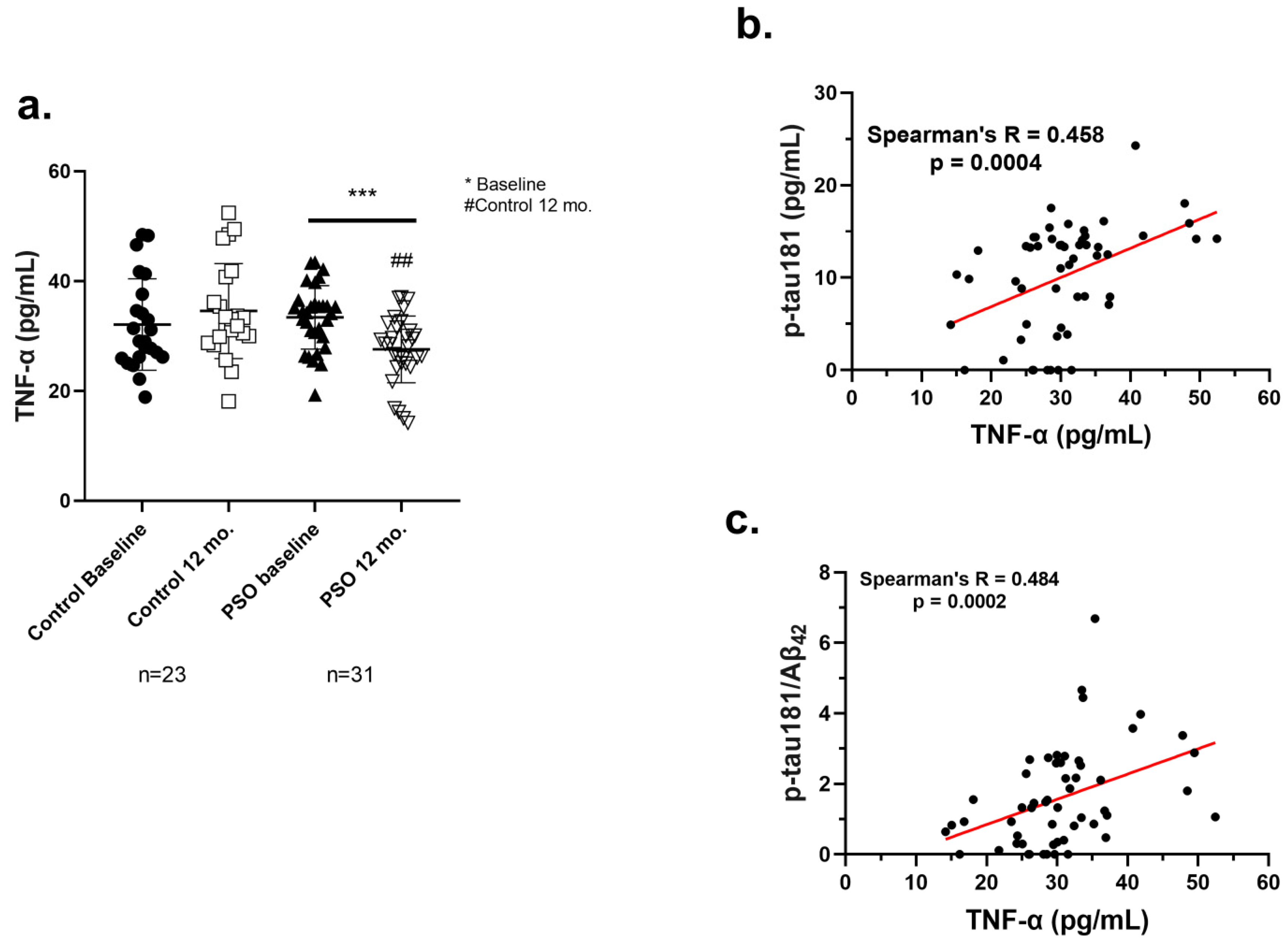
3.2.4. PSO Consumption Reduced TNF-α Levels in Sera of MCI Patients
3.2.5. Positive Correlation of TNF-α with AD-Related Biomarkers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- 2023 Alzheimer’s Disease Facts and Figures. Alzheimer’s Dement. 2023, 19, 1598–1695. [CrossRef] [PubMed]
- Gunes, S.; Aizawa, Y.; Sugashi, T.; Sugimoto, M.; Rodrigues, P.P. Biomarkers for Alzheimer’s Disease in the Current State: A Narrative Review. Int. J. Mol. Sci. 2022, 23, 4692. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.; Knopman, D.S. Classification and Epidemiology of MCI. Clin. Geriatr. Med. 2013, 29, 753–772. [Google Scholar] [CrossRef]
- Selkoe, D.J. Alzheimer’s Disease: A Central Role for Amyloid. J. Neuropathol. Exp. Neurol. 1994, 53, 438–447. [Google Scholar] [CrossRef]
- Hardy, J.; Selkoe, D.J. The Amyloid Hypothesis of Alzheimer’s Disease: Progress and Problems on the Road to Therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef]
- Prajapat, M.; Kaur, G.; Choudhary, G.; Pahwa, P.; Bansal, S.; Joshi, R.; Batra, G.; Mishra, A.; Singla, R.; Kaur, H.; et al. A Systematic Review for the Development of Alzheimer’s Disease in in Vitro Models: A Focus on Different Inducing Agents. Front. Aging Neurosci. 2023, 15, 1296919. [Google Scholar] [CrossRef]
- Biber, K.; Neumann, H.; Inoue, K.; Boddeke, H.W.G.M. Neuronal “On” and “Off” Signals Control Microglia. Trends Neurosci. 2007, 30, 596–602. [Google Scholar] [CrossRef]
- Wyss-Coray, T.; Rogers, J. Inflammation in Alzheimer Disease-A Brief Review of the Basic Science and Clinical Literature. Cold Spring Harb. Perspect. Med. 2012, 2, a006346. [Google Scholar] [CrossRef] [PubMed]
- Tejera, D.; Heneka, M.T. Microglia in Alzheimer’s Disease: The Good, the Bad and the Ugly. Curr. Alzheimer Res. 2016, 13, 370–380. [Google Scholar] [CrossRef]
- Minter, M.R.; Taylor, J.M.; Crack, P.J. The Contribution of Neuroinflammation to Amyloid Toxicity in Alzheimer’s Disease. J. Neurochem. 2016, 136, 457–474. [Google Scholar] [CrossRef]
- Ansari, M.A.; Scheff, S.W. Oxidative Stress in the Progression of Alzheimer Disease in the Frontal Cortex. J. Neuropathol. Exp. Neurol. 2010, 69, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.J.; Zhang, X.; Chen, W.W. Role of Oxidative Stress in Alzheimer’s Disease (Review). Biomed. Rep. 2016, 4, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Zabel, M.; Nackenoff, A.; Kirsch, W.M.; Harrison, F.E.; Perry, G.; Schrag, M. Markers of Oxidative Damage to Lipids, Nucleic Acids and Proteins and Antioxidant Enzymes Activities in Alzheimer’s Disease Brain: A Meta-Analysis in Human Pathological Specimens. Free Radic. Biol. Med. 2018, 115, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Tzekaki, E.E.; Tsolaki, M.; Pantazaki, A.A.; Geromichalos, G.; Lazarou, E.; Kozori, M.; Sinakos, Z. The Pleiotropic Beneficial Intervention of Olive Oil Intake on the Alzheimer’s Disease Onset via Fibrinolytic System. Exp. Gerontol. 2021, 150, 111344. [Google Scholar] [CrossRef]
- Okado-Matsumoto, A.; Fridovich, I. Subcellular Distribution of Superoxide Dismutases (SOD) in Rat Liver. Cu,Zn-SOD in Mitochondria. J. Biol. Chem. 2001, 276, 38388–38393. [Google Scholar] [CrossRef]
- Youssef, P.; Chami, B.; Lim, J.; Middleton, T.; Sutherland, G.T.; Witting, P.K. Evidence Supporting Oxidative Stress in a Moderately Affected Area of the Brain in Alzheimer’s Disease. Sci. Rep. 2018, 8, 11553. [Google Scholar] [CrossRef]
- Wu, W.; Ji, Y.; Wang, Z.; Wu, X.; Li, J.; Gu, F.; Chen, Z.; Wang, Z. The FDA-Approved Anti-Amyloid-β Monoclonal Antibodies for the Treatment of Alzheimer’s Disease: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Eur. J. Med. Res. 2023, 28, 544. [Google Scholar] [CrossRef]
- Hassan, M.; Raza, H.; Abbasi, M.A.; Moustafa, A.A.; Seo, S.Y. The Exploration of Novel Alzheimer’s Therapeutic Agents from the Pool of FDA Approved Medicines Using Drug Repositioning, Enzyme Inhibition and Kinetic Mechanism Approaches. Biomed. Pharmacother. 2019, 109, 2513–2526. [Google Scholar] [CrossRef]
- Gómez-Pinilla, F. Brain Foods: The Effects of Nutrients on Brain Function. Nat. Rev. Neurosci. 2008, 9, 568–578. [Google Scholar] [CrossRef]
- Francis, S.T.; Head, K.; Morris, P.G.; Macdonald, I.A. The Effect of Flavanol-Rich Cocoa on the FMRI Response to a Cognitive Task in Healthy Young People. J. Cardiovasc. Pharmacol. 2006, 47, S215–S220. [Google Scholar] [CrossRef]
- Mehdi, A.; Lamiae, B.; Samira, B.; Ramchoun, M.; Abdelouahed, K.; Tamas, F.; Hicham, B. Pomegranate (Punica granatum L.) Attenuates Neuroinflammation Involved in Neurodegenerative Diseases. Foods 2022, 11, 2570. [Google Scholar] [CrossRef]
- Khan, A.; Ali, S.; Murad, W.; Hayat, K.; Siraj, S.; Jawad, M.; Khan, R.A.; Uddin, J.; Al-Harrasi, A.; Khan, A. Phytochemical and Pharmacological Uses of Medicinal Plants to Treat Cancer: A Case Study from Khyber Pakhtunkhwa, North Pakistan. J. Ethnopharmacol. 2021, 281, 114437. [Google Scholar] [CrossRef] [PubMed]
- Nazil, N.; Begum, H.A.; Hamayun, M.; Khan, A.; Yaseem, T.; Bussmann, R.W.; Murad, W. Quantitative Ethnobotanical Appraisal of Medicinal Plants Used by Indigenous Communities of District Malakand, Pakistan. Ethnobot. Res. Appl. 2022, 24, 1–14. [Google Scholar] [CrossRef]
- Ullah, A.; Khan, A.; Ahmed, S.; Irfan, H.M.; Hafiz, A.A.; Jabeen, K.; Alruwaili, M.; Alotaibi, M.; Al Naeem, W.; Khan, A.; et al. A Review of Pomegranate Supplementation: A Promising Remedial Avenue for Alzheimer’s Disease. Heliyon 2023, 9, e22483. [Google Scholar] [CrossRef]
- Hernández, F.; Melgarejo, P.; Tomás-Barberán, F.A.; Artés, F. Evolution of Juice Anthocyanins during Ripening of New Selected Pomegranate (Punica granatum) Clones. Eur. Food Res. Technol. 1999, 210, 39–42. [Google Scholar] [CrossRef]
- Saparbekova, A.A.; Kantureyeva, G.O.; Kudasova, D.E.; Konarbayeva, Z.K.; Latif, A.S. Potential of Phenolic Compounds from Pomegranate (Punica granatum L.) by-Product with Significant Antioxidant and Therapeutic Effects: A Narrative Review. Saudi J. Biol. Sci. 2023, 30, 103553. [Google Scholar] [CrossRef]
- Singh, R.P.; Chidambara Murthy, K.N.; Jayaprakasha, G.K. Studies on the Antioxidant Activity of Pomegranate (Punica granatum) Peel and Seed Extracts Using in Vitro Models. J. Agric. Food Chem. 2002, 50, 81–86. [Google Scholar] [CrossRef]
- Mastrodi Salgado, J.; Ferreira, T.R.B.; de Oliveira Biazotto, F.; dos Santos Dias, C.T. Increased Antioxidant Content in Juice Enriched with Dried Extract of Pomegranate (Punica granatum) Peel. Plant Foods Hum. Nutr. 2012, 67, 39–43. [Google Scholar] [CrossRef]
- Derakhshan, Z.; Ferrante, M.; Tadi, M.; Ansari, F.; Heydari, A.; Hosseini, M.S.; Conti, G.O.; Sadrabad, E.K. Antioxidant Activity and Total Phenolic Content of Ethanolic Extract of Pomegranate Peels, Juice and Seeds. Food Chem. Toxicol. 2018, 114, 108–111. [Google Scholar] [CrossRef]
- Chidambara Murthy, K.N.; Jayaprakasha, G.K.; Singh, R.P. Studies on Antioxidant Activity of Pomegranate (Punica granatum) Peel Extract Using in Vivo Models. J. Agric. Food Chem. 2002, 50, 4791–4795. [Google Scholar] [CrossRef]
- Ciccone, L.; Nencetti, S.; Rossello, A.; Orlandini, E. Pomegranate: A Source of Multifunctional Bioactive Compounds Potentially Beneficial in Alzheimer’s Disease. Pharmaceuticals 2023, 16, 1036. [Google Scholar] [CrossRef] [PubMed]
- Chatzikostopoulos, T.; Tsolaki, M. The effects of pomegranate seed oil on mild cognitive impairment. J. Prev. Alzheimer’s Dis. 2022, 9, 1961–1970. [Google Scholar] [CrossRef]
- Siddarth, P.; Li, Z.; Miller, K.J.; Ercoli, L.M.; Merril, D.A.; Henning, S.M.; Heber, D.; Small, G.W. Randomized Placebo-Controlled Study of the Memory Effects of Pomegranate Juice in Middle-Aged and Older Adults. Am. J. Clin. Nutr. 2020, 111, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Hartman, R.E.; Shah, A.; Fagan, A.M.; Schwetye, K.E.; Parsadanian, M.; Schulman, R.N.; Finn, M.B.; Holtzman, D.M. Pomegranate Juice Decreases Amyloid Load and Improves Behavior in a Mouse Model of Alzheimer’s Disease. Neurobiol. Dis. 2006, 24, 506–515. [Google Scholar] [CrossRef]
- Rojanathammanee, L.; Puig, K.L.; Combs, C.K. Pomegranate Polyphenols and Extract Inhibit Nuclear Factor of Activated T-Cell Activity and Microglial Activation Invitroand Inatransgenic Mouse Model of Alzheimer Disease. J. Nutr. 2013, 143, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, N.; Periyanayagam, K.; Sugithra, B. Pomegranate (Punicagranatum.Linn. var. Ganesh) Leaf Extracts Ameliorates Neurotoxicity In Transgenic Drosophila Expressing Human Amyloid Pathology. Indian J. Pharm. Biol. Res. 2021, 9, 1–7. [Google Scholar] [CrossRef]
- Sander, H.; Wallace, S.; Plouse, R.; Tiwari, S.; Gomes, A.V. Ponceau S Waste: Ponceau S Staining for Total Protein Normalization. Anal. Biochem. 2019, 575, 44–53. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Human Experimentation. Code of Ethics of the World Medical Association. Br. Med. J. 1964, 2, 177. [CrossRef]
- Skrzypczak-Wiercioch, A.; Sałat, K. Lipopolysaccharide-Induced Model of Neuroinflammation: Mechanisms of Action, Research Application and Future Directions for Its Use. Molecules 2022, 27, 5481. [Google Scholar] [CrossRef]
- Feng, S.; Achoute, L.; Margolskee, R.F.; Jiang, P.; Wang, H. Lipopolysaccharide-Induced Inflammatory Cytokine Expression in Taste Organoids. Chem. Senses 2020, 45, 187–194. [Google Scholar] [CrossRef]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 Signal Transduction Pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Hewett, S.J.; Jackman, N.A.; Claycomb, R.J.; Neuroscience, P.I.; Haven, N. Interleukin-1β in Central Nervous System Injury and Repair Sandra. Eur. J. Neurodegener. Dis. 2012, 1, 195–211. [Google Scholar] [PubMed]
- Halle, A.; Hornung, V.; Petzold, G.C.; Stewart, C.R.; Monks, B.G.; Reinheckel, T.; Fitzgerald, K.A.; Latz, E.; Moore, K.J.; Golenbock, D.T. The NALP3 Inflammasome Is Involved in the Innate Immune Response to Amyloid-β. Nat. Immunol. 2008, 9, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Tzekaki, E.E.; Papaspyropoulos, A.; Tsolaki, M.; Lazarou, E.; Kozori, M.; Pantazaki, A.A. Restoration of BMI1 Levels after the Administration of Early Harvest Extra Virgin Olive Oil as a Therapeutic Strategy against Alzheimer’s Disease. Exp. Gerontol. 2021, 144, 111178. [Google Scholar] [CrossRef]
- Janelidze, S.; Stomrud, E.; Palmqvist, S.; Zetterberg, H.; Van Westen, D.; Jeromin, A.; Song, L.; Hanlon, D.; Tan Hehir, C.A.; Baker, D.; et al. Plasma β-Amyloid in Alzheimer’s Disease and Vascular Disease. Sci. Rep. 2016, 6, 26801. [Google Scholar] [CrossRef]
- Klafki, H.W.; Morgado, B.; Wirths, O.; Jahn, O.; Bauer, C.; Esselmann, H.; Schuchhardt, J.; Wiltfang, J. Is Plasma Amyloid-β 1–42/1–40 a Better Biomarker for Alzheimer’s Disease than AβX—42/X—40? Fluids Barriers CNS 2022, 19, 96. [Google Scholar] [CrossRef]
- Meng, J. Plasma PTau181 as a Biomarker for Alzheimer’s Disease. Medcomm 2020, 1, 74–76. [Google Scholar] [CrossRef]
- Chong, J.R.; Ashton, N.J.; Karikari, T.K.; Tanaka, T.; Saridin, F.N.; Reilhac, A.; Robins, E.G.; Nai, Y.H.; Vrooman, H.; Hilal, S.; et al. Plasma P-Tau181 to Aβ42 Ratio Is Associated with Brain Amyloid Burden and Hippocampal Atrophy in an Asian Cohort of Alzheimer’s Disease Patients with Concomitant Cerebrovascular Disease. Alzheimer’s Dement. 2021, 17, 1649–1662. [Google Scholar] [CrossRef]
- Tzekaki, E.E.; Katsipis, G.; Andreadou, E.G.; Mouzakidis, C.; Baldimtsi, E.N.; Karathanasi, E.M.; Hassandra, M.; Galanis, E.; Hatzigeorgiadis, A.; Goudas, M.; et al. VRADA Training System as a Non-Pharmacological Dual Intervention to Alleviate Symptoms of the Pathophysiology of Mild Cognitive Impairment. Brain Disord. 2023, 11, 100090. [Google Scholar] [CrossRef]
- Katsipis, G.; Tzekaki, E.E.; Andreadou, E.G.; Mouzakidis, C.; Baldimtsi, E.N.; Karathanasi, E.M.; Hassandra, M.; Galanis, E.; Hatzigeorgiadis, A.; Goudas, M.; et al. The Effect of Physical Exercise with Cognitive Training on Inflammation and Alzheimer’s Disease Biomarkers of Mild Cognitive Impairment Patients. Neurosci. Appl. 2024, 3, 104085. [Google Scholar] [CrossRef]
- Ou, W.; Yang, J.; Simanauskaite, J.; Choi, M.; Castellanos, D.M.; Chang, R.; Sun, J.; Jagadeesan, N.; Parfitt, K.D.; Cribbs, D.H.; et al. Biologic TNF—α Inhibitors Reduce Microgliosis, Neuronal Loss, and Tau Phosphorylation in a Transgenic Mouse Model of Tauopathy. J. Neuroinflamm. 2021, 18, 312. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lian, S.; Zhang, Y.; Zhao, Q. Efficacy and Safety of Donepezil for Mild Cognitive Impairment: A Systematic Review and Meta-Analysis. Clin. Neurol. Neurosurg. 2022, 213, 107134. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Pacheco, N.; Sarmiento-Blanco, S.; Vergara-Cadavid, G.; Castro-Leones, M.; Contreras-Puentes, N. Monoclonal Therapy with Lecanemab in the Treatment of Mild Alzheimer’s Disease: A Systematic Review and Meta-Analysis. Ageing Res. Rev. 2025, 104, 102620. [Google Scholar] [CrossRef]
- Heidebrink, J.L.; Paulson, H.L. Lessons Learned from Approval of Aducanumab for Alzheimer’s Disease. Annu. Rev. Med. 2024, 75, 99–111. [Google Scholar] [CrossRef]
- Cordiano, R.; Gammeri, L.; Di Salvo, E.; Gangemi, S.; Minciullo, P.L. Pomegranate (Punica granatum L.) Extract Effects on Inflammaging. Molecules 2024, 29, 4174. [Google Scholar] [CrossRef] [PubMed]
- Boroushaki, M.T.; Mollazadeh, H.; Afshari, A.R. Pomegranate Seed Oil: A Comprehensive Review on Its Therapeutic Effects. Int. J. Pharm. Sci. Res. 2016, 7, 430–442. [Google Scholar] [CrossRef]
- Fahmy, H.; Hegazi, N.; El-Shamy, S.; Farag, M.A. Pomegranate Juice as a Functional Food: A Comprehensive Review of Its Polyphenols, Therapeutic Merits, and Recent Patents. Food Funct. 2020, 11, 5768–5781. [Google Scholar] [CrossRef]
- Caruso, A.; Barbarossa, A.; Tassone, A.; Ceramella, J.; Carocci, A.; Catalano, A.; Basile, G.; Fazio, A.; Iacopetta, D.; Franchini, C.; et al. Pomegranate: Nutraceutical with Promising Benefits on Human Health. Appl. Sci. 2020, 10, 6915. [Google Scholar] [CrossRef]
- Tastan, O.; Baysal, T. Adulteration Analysis of Pomegranate Juice. In Frontiers in Drug Safety; Bentham Science: Sharjah, United Arab Emirates, 2018. [Google Scholar]
- Ahles, S.; Cuijpers, I.; Hartgens, F.; Troost, F.J. The Effect of a Citrus and Pomegranate Complex on Physical Fitness and Mental Well-Being in Healthy Elderly: A Randomized Placebo-Controlled Trial. J. Nutr. Heal. Aging 2022, 26, 839–846. [Google Scholar] [CrossRef]
- Petrou, B.; Ginzberg, A.; Binyamin, O.; Karussis, D. Beneficial Effects of a Nano Formulation of Pomegranate Seed Oil, GranaGard, on the Cognitive Function of Multiple Sclerosis Patients. Mult. Scler. Relat. Disord. 2021, 54, 103103. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.L.; Rodriguez-Ortiz, C.J.; Kitazawa, M. Infection, Systemic Inflammation, and Alzheimer’s Disease. Microbes Infect. 2015, 17, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Itzhaki, R.F.; Golde, T.E.; Heneka, M.T.; Readhead, B. Do Infections Have a Role in the Pathogenesis of Alzheimer Disease? Nat. Rev. Neurol. 2020, 16, 193–197. [Google Scholar] [CrossRef]
- Nava Catorce, M.; Gevorkian, G. LPS-Induced Murine Neuroinflammation Model: Main Features and Suitability for Pre-Clinical Assessment of Nutraceuticals. Curr. Neuropharmacol. 2016, 14, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; McGeer, E.; McGeer, P.L. Activated Human Microglia Stimulate Neuroblastoma Cells to Upregulate Production of Beta Amyloid Protein and Tau: Implications for Alzheimer’s Disease Pathogenesis. Neurobiol. Aging 2015, 36, 42–52. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, Y.K.; Yuk, D.Y.; Choi, D.Y.; Ban, S.B.; Oh, K.W.; Hong, J.T. Neuro-Inflammation Induced by Lipopolysaccharide Causes Cognitive Impairment through Enhancement of Beta-Amyloid Generation. J. Neuroinflamm. 2008, 5, 37. [Google Scholar] [CrossRef]
- Wang, L.-M.; Wu, Q.; Kirk, R.A.; Horn, K.P.; Ebada Salem, A.H.; Hoffman, J.M.; Yap, J.T.; Sonnen, J.A.; Towner, R.A.; Bozza, F.A.; et al. Lipopolysaccharide Endotoxemia Induces Amyloid-β and p-Tau Formation in the Rat Brain. Am. J. Nucl. Med. Mol. Imaging 2018, 8, 86–99. [Google Scholar]
- Lee, Y.J.; Choi, D.Y.; Choi, I.S.; Kim, K.H.; Kim, Y.H.; Kim, H.M.; Lee, K.; Cho, W.G.; Jung, J.K.; Han, S.B.; et al. Inhibitory Effect of 4-O-Methylhonokiol on Lipopolysaccharide-Induced Neuroinflammation, Amyloidogenesis and Memory Impairment via Inhibition of Nuclear Factor-KappaB in Vitro and in Vivo Models. J. Neuroinflamm. 2012, 9, 35. [Google Scholar] [CrossRef]
- Liu, J.; Wang, D.; Li, S.Q.; Yu, Y.; Ye, R.D. Suppression of LPS-Induced Tau Hyperphosphorylation by Serum Amyloid A. J. Neuroinflamm. 2016, 13, 28. [Google Scholar] [CrossRef]
- Meratan, A.A.; Hassani, V.; Mahdavi, A.; Nikfarjam, N. Pomegranate Seed Polyphenol-Based Nanosheets as an Efficient Inhibitor of Amyloid Fibril AsSDbly and Cytotoxicity of HEWL. RSC Adv. 2022, 12, 8719–8730. [Google Scholar] [CrossRef]
- Kwak, H.M.; Jeon, S.Y.; Sohng, B.H.; Kim, J.G.; Lee, J.M.; Lee, K.B.; Jeong, H.H.; Hur, J.M.; Kang, Y.H.; Song, K.S. β-Secretase (BACE1) Inhibitors from Pomegranate (Punica granatum) Husk. Arch. Pharm. Res. 2005, 28, 1328–1332. [Google Scholar] [CrossRef] [PubMed]
- Khokar, R.; Hachani, K.; Hasan, M.; Othmani, F.; Essam, M.; Al Mamari, A.; UM, D.; Khan, S.A. Anti-Alzheimer Potential of a Waste by-Product (Peel) of Omani Pomegranate Fruits: Quantification of Phenolic Compounds, in-Vitro Antioxidant, Anti-Cholinesterase and in-Silico Studies. Biocatal. Agric. Biotechnol. 2021, 38, 102223. [Google Scholar] [CrossRef]
- Yousof Ali, M.; Zaib, S.; Jannat, S.; Khan, I. Discovery of Potent and Selective Dual Cholinesterases and β-Secretase Inhibitors in Pomegranate as a Treatment for Alzheimer’s Disease. Bioorg. Chem. 2022, 129, 106137. [Google Scholar] [CrossRef]
- Alami, M.; Boumezough, K.; Zerif, E.; Zoubdane, N.; Khalil, A.; Bunt, T.; Laurent, B.; Witkowski, J.M.; Ramassamy, C.; Boulbaroud, S.; et al. In Vitro Assessment of the Neuroprotective Effects Of Pomegranate (Punica granatum L.) Polyphenols Against Tau Phosphorylation, Neuroinflammation, and Oxidative Stress. Nutrients 2024, 16, 3667. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Li, J.; Zhang, X.; Wang, L.; Zhang, W.; Yang, M.; Hou, C. Pomegranate Peel Polyphenols Inhibits Inflammation in LPS-Induced RAW264.7 Macrophages via the Suppression of TLR4/NF-ΚB Pathway Activation. Food Nutr. Res. 2019, 63. [Google Scholar] [CrossRef]
- Cao, Y.; Chen, J.; Ren, G.; Zhang, Y.; Tan, X.; Yang, L. Punicalagin Prevents Inflammation in Lps-Induced Raw264.7 Macrophages by Inhibiting Foxo3a/Autophagy Signaling Pathway. Nutrients 2019, 11, 2794. [Google Scholar] [CrossRef]
- Racková, L.; Ergin, V.; Bali, E.B.; Kuniaková, M.; Karasu, Ç. Pomegranate Seed Oil Modulates Functions and Survival of BV-2 Microglial Cells in Vitro. Int. J. Vitam. Nutr. Res. 2014, 84, 295–309. [Google Scholar] [CrossRef]
- Mattson, M.P. Hormesis Defined. Ageing Res. Rev. 2008, 7, 1–7. [Google Scholar] [CrossRef]
- Lansky, E.P.; Newman, R.A. Punica granatum (Pomegranate) and Its Potential for Prevention and Treatment of Inflammation and Cancer. J. Ethnopharmacol. 2007, 109, 177–206. [Google Scholar] [CrossRef]
- Butterfield, D.A. Amyloid β-Peptide (1-42)-Induced Oxidative Stress and Neurotoxicity: Implications for Neurodegeneration in Alzheimer’s Disease Brain. A Review. Free Radic. Res. 2002, 36, 1307–1313. [Google Scholar] [CrossRef]
- Bisht, K.; Sharma, K.; Tremblay, M.È. Chronic Stress as a Risk Factor for Alzheimer’s Disease: Roles of Microglia-Mediated Synaptic Remodeling, Inflammation, and Oxidative Stress. Neurobiol. Stress 2018, 9, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Subash, S.; Essa, M.; Al-Asmi, A.; Al-Adawi, S.; Vaishnav, R.; Braidy, N.; Manivasagam, T.; Guillemin, G. Pomegranate from Oman Alleviates the Brain Oxidative Damage in Transgenic Mouse Model of Alzheimer′s Disease. J. Tradit. Complement. Med. 2014, 4, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Harakeh, S.; Ramadan, W.; Al Muhayawi, M.; Al Jaouni, S.; Mousa, S.; Hakeem, K. Pomegranate Peel Extract Lessens Histopathologic Changes and Restores Antioxidant Homeostasis in the Hippocampus of Rats with Aluminium Chloride-Induced Alzheimer’s Disease. Asian Pac. J. Trop. Med. 2020, 13, 456–463. [Google Scholar] [CrossRef]
- Molani-Gol, R.; Foroumandi, E.; Alizadeh, M.; Kheirouri, S. Pomegranate and Cognitive Performance: A Systematic Review. Curr. Pharm. Des. 2023, 29, 928–939. [Google Scholar] [CrossRef]
- Bookheimer, S.Y.; Renner, B.A.; Ekstrom, A.; Li, Z.; Henning, S.M.; Brown, J.A.; Jones, M.; Moody, T.; Small, G.W. Pomegranate Juice Augments Memory and FMRI Activity in Middle-Aged and Older Adults with Mild Memory Complaints. Evid.-Based Complement. Altern. Med. 2013, 2013, 946298. [Google Scholar] [CrossRef]
- Fowler, C.J.; Stoops, E.; Rainey-Smith, S.R.; Vanmechelen, E.; Vanbrabant, J.; Dewit, N.; Mauroo, K.; Maruff, P.; Rowe, C.C.; Fripp, J.; et al. Plasma P-Tau181/Aβ1-42 Ratio Predicts Aβ-PET Status and Correlates with CSF-p-Tau181/Aβ1-42 and Future Cognitive Decline. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2022, 14, e12375. [Google Scholar] [CrossRef]
- Graff-Radford, N.R.; Crook, J.E.; Lucas, J.; Boeve, B.F.; Knopman, D.S.; Ivnik, R.J.; Smith, G.E.; Younkin, L.H.; Petersen, R.C.; Younkin, S.G. Association of Low Plasma Abeta42/Abeta40 Ratios with Increased Imminent Risk for Mild Cognitive Impairment and Alzheimer Disease. Arch. Neurol. 2007, 64, 354–362. [Google Scholar] [CrossRef]
- Akbarian, M.; Mirzavi, F.; Amirahmadi, S.; Hosseini, M.; Alipour, M.; Feizi, H.; Rajabian, A. Amelioration of Oxidative Stress, Cholinergic Dysfunction, and Neuroinflammation in Scopolamine-Induced Amnesic Rats Fed with Pomegranate Seed. Inflammopharmacology 2022, 30, 1021–1035. [Google Scholar] [CrossRef]
- Xu, P.; Xu, L.; Huang, S.; Li, D.; Liu, Y.; Guo, H.; Dai, N.; Hong, Z.; Zhong, S. Analysis of the Molecular Mechanism of Punicalagin in the Treatment of Alzheimer’s Disease by Computer-Aided Drug Research Technology. ACS Omega 2022, 7, 6121–6132. [Google Scholar] [CrossRef]
- Chen, P.; Chen, F.; Lei, J.; Zhou, B. Pomegranate Polyphenol Punicalagin Improves Learning Memory Deficits, Redox Homeostasis, and Neuroinflammation in Aging Mice. Phyther. Res. 2023, 37, 3655–3674. [Google Scholar] [CrossRef]
- Essa, M.M.; Subash, S.; Akbar, M.; Al-Adawi, S.; Guillemin, G.J. Long-Term Dietary Supplementation of Pomegranates, Figures and Dates Alleviate Neuroinflammation in a Transgenic Mouse Model of Alzheimer’s Disease. PLoS ONE 2015, 10, e0120964. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.E.; Hwang, C.J.; Lee, H.P.; Kim, C.S.; Son, D.J.; Ham, Y.W.; Hellström, M.; Han, S.B.; Kim, H.S.; Park, E.K.; et al. Inhibitory Effect of Punicalagin on Lipopolysaccharide-Induced Neuroinflammation, Oxidative Stress and Memory Impairment via Inhibition of Nuclear Factor-KappaB. Neuropharmacology 2017, 117, 21–32. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood Donor Demographics | |||
| Demographics | Control | PSO | p Values |
| Participants Number (N) | 23 | 31 | |
| Gender (Female/Male) | 21/2 | 19/12 | >0.999 |
| Age (years) | 75.09 ± 7.2 | 72.81 ± 7.4 | 0.2637 |
| Education (years) | 11.83 ± 3.35 | 11.4 ± 4.1 | 0.8442 |
| MMSE | |||
| Control Baseline | Control After | PSO Baseline | PSO After |
| 27.35 ± 1.555 | 26.96 ± 1.665 | 27.65 ± 1.380 | 28.45 ± 1.877 a,b |
| Biomarker | Biomarker Analysis of Study Groups | ||||
|---|---|---|---|---|---|
| Control (N = 23) | PSO (N = 31) | p-Values | |||
| Baseline (B) | 12 mo.(A) | Baseline (B) | 12 mo.(A) | ||
| Aβ42 (pg/mL) | 8.197 ± 2.343 | 6.157 ± 2.671 | 8.999 ± 3.280 | 10.22 ± 2.984 | CB-CA:0.0025 PSOB-PSOA: 0.0729 CA-PSOA: <0.0001 |
| Aβ40 (pg/mL) | 21.70 ± 3.123 | 24.15 ± 3.395 | 20.48 ± 5.628 | 16.22 ± 3.144 | CB-CA: 0.0002 PSOB-PSOA: 0.0014 CA-PSOA: <0.0001 |
| Aβ42/Aβ40 | 0.3842 ± 0.1140 | 0.2596 ± 0.1256 | 0.4602 ± 0.1961 | 0.5834 ± 0.2285 | CB-CA: 0.0005 PSOB-PSOA: 0.0146 CA-PSOA: <0.0001 |
| p-tau181 (pg/mL) | 11.41 ± 2.515 | 14.36 ± 2.862 | 12.13 ± 3.247 | 7.106 ± 5.485 | CB-CA: <0.0001 PSOB-PSOA: 0.0001 CA-PSOA: <0.0001 |
| ptau-181/Aβ42 | 1.512 ± 0.6242 | 2.766 ± 1.291 | 1.504 ± 0.6499 | 0.7394 ± 0.6782 | CB-CA: 0.0004 PSOB-PSOA:0.0004 CA-PSOA: <0.0001 |
| TNF-α (pg/mL) | 32.09 ± 8.332 | 34.56 ± 8.651 | 33.41 ± 5.771 | 27.57 ± 6.098 | CB-CA: 0.2747 PSOB-PSOA: 0.0008 CA-PSOA: 0.0010 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tzekaki, E.E.; Katsipis, G.; Chatzikostopoulos, A.; Koutoupa, A.; Lavrentiadou, S.N.; Tsolaki, M.; Pantazaki, A.A. Neuroprotective, Antioxidant and Anti-Inflammatory Effect of Greek Pomegranate Seed Oil on N2a Neuroblastoma Cells and Mild Cognitive Impairment Patients. Biology 2025, 14, 548. https://doi.org/10.3390/biology14050548
Tzekaki EE, Katsipis G, Chatzikostopoulos A, Koutoupa A, Lavrentiadou SN, Tsolaki M, Pantazaki AA. Neuroprotective, Antioxidant and Anti-Inflammatory Effect of Greek Pomegranate Seed Oil on N2a Neuroblastoma Cells and Mild Cognitive Impairment Patients. Biology. 2025; 14(5):548. https://doi.org/10.3390/biology14050548
Chicago/Turabian StyleTzekaki, Eleni E., Georgios Katsipis, Athanasios Chatzikostopoulos, Anna Koutoupa, Sophia N. Lavrentiadou, Magda Tsolaki, and Anastasia A. Pantazaki. 2025. "Neuroprotective, Antioxidant and Anti-Inflammatory Effect of Greek Pomegranate Seed Oil on N2a Neuroblastoma Cells and Mild Cognitive Impairment Patients" Biology 14, no. 5: 548. https://doi.org/10.3390/biology14050548
APA StyleTzekaki, E. E., Katsipis, G., Chatzikostopoulos, A., Koutoupa, A., Lavrentiadou, S. N., Tsolaki, M., & Pantazaki, A. A. (2025). Neuroprotective, Antioxidant and Anti-Inflammatory Effect of Greek Pomegranate Seed Oil on N2a Neuroblastoma Cells and Mild Cognitive Impairment Patients. Biology, 14(5), 548. https://doi.org/10.3390/biology14050548