
Synergistic Toxicity of Combined Exposure to Acrylamide and Polystyrene Nanoplastics on the Gut–Liver Axis in Mice


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Animal Experimental Design
2.3. Biochemical Analyses of Serum, Liver, and Colon
2.4. Histopathological Analyses of the Liver and Colon
2.5. Real-Time Quantitative PCR (RT-qPCR) Analysis
2.6. Untargeted Metabolomics of Liver and Bioinformatics Analysis
2.7. 16S rRNA Amplicon Sequencing of Cecal Contents and Bioinformatics Analysis
2.8. Statistical Analysis
3. Results
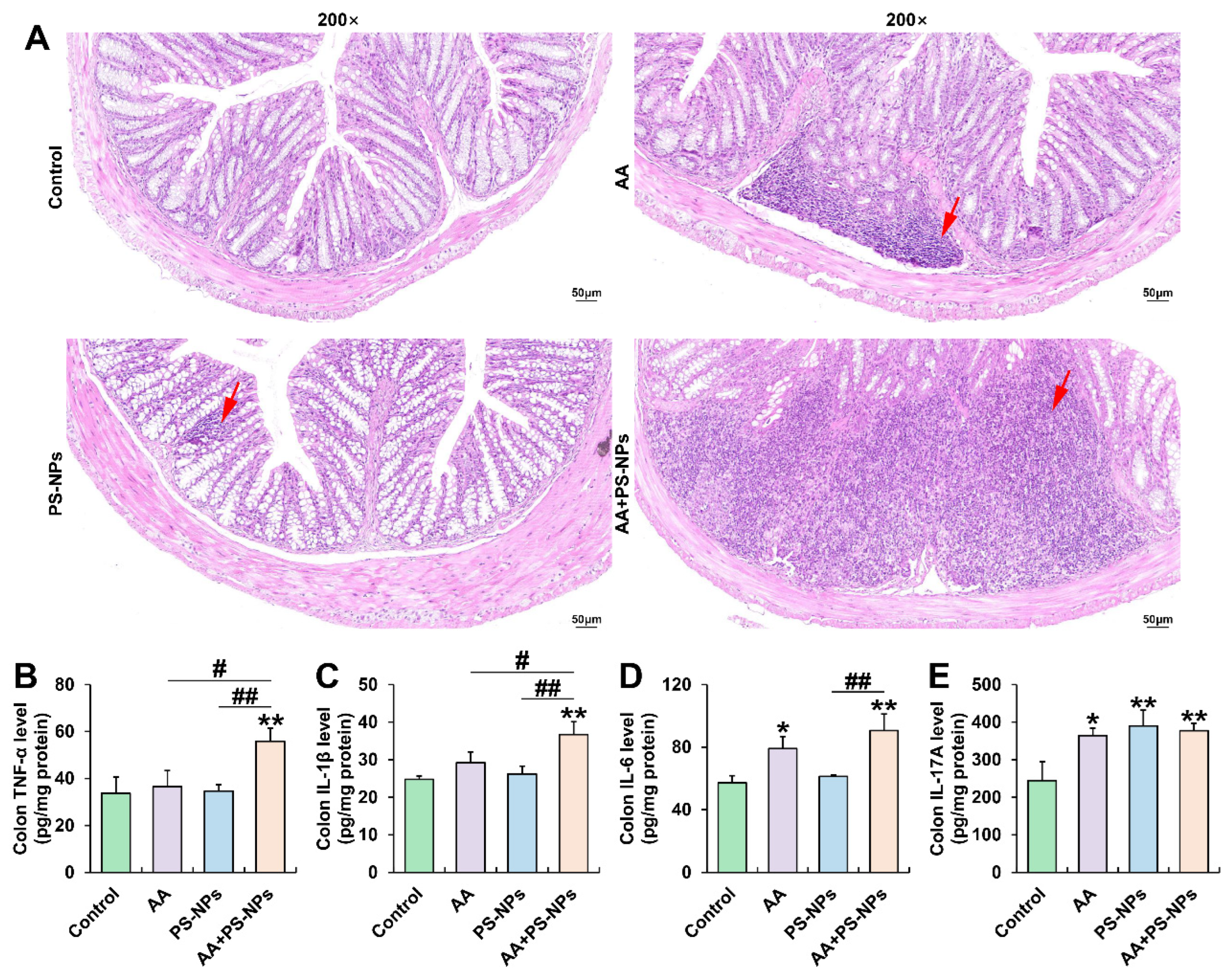
3.1. Co-Exposure to AA and PS-NPs Aggravates Colonic Inflammation in Mice
3.2. Co-Exposure to AA and PS-NPs Damages Colonic Barrier Function in Mice
3.3. Co-Exposure to AA and PS-NPs Exacerbates Liver Injury and Inflammation in Mice
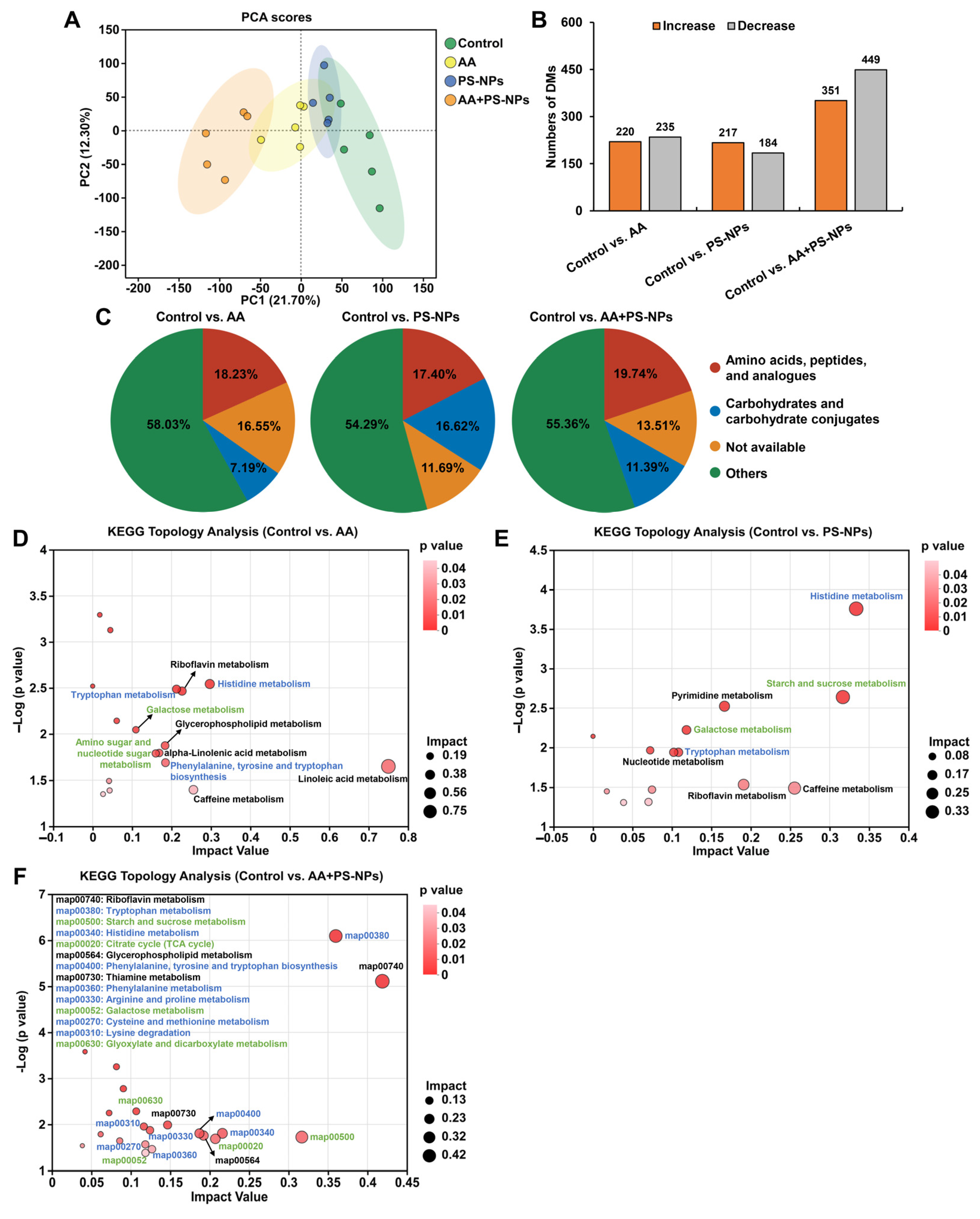
3.4. Co-Exposure to AA and PS-NPs Alters Liver Metabolism in Mice
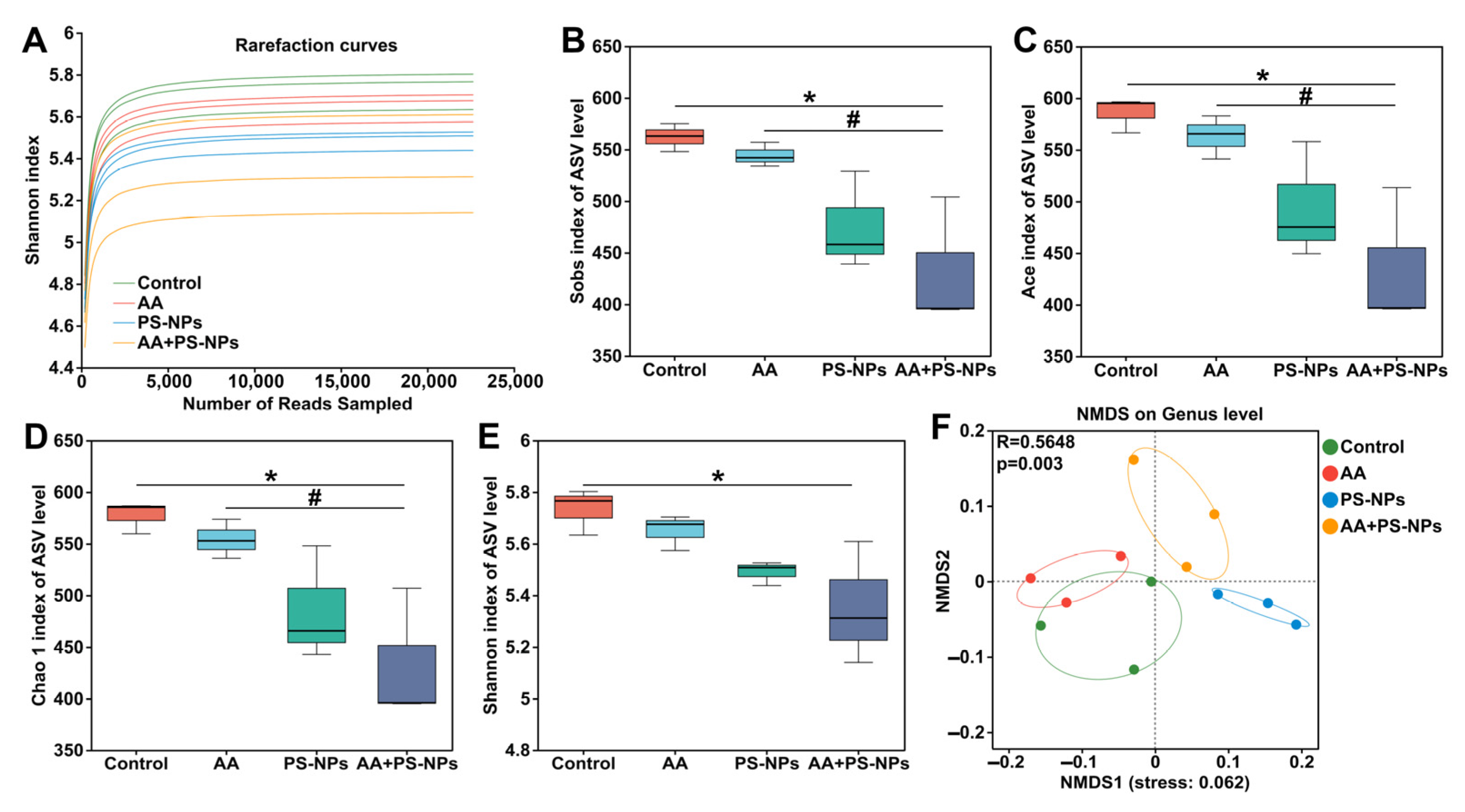
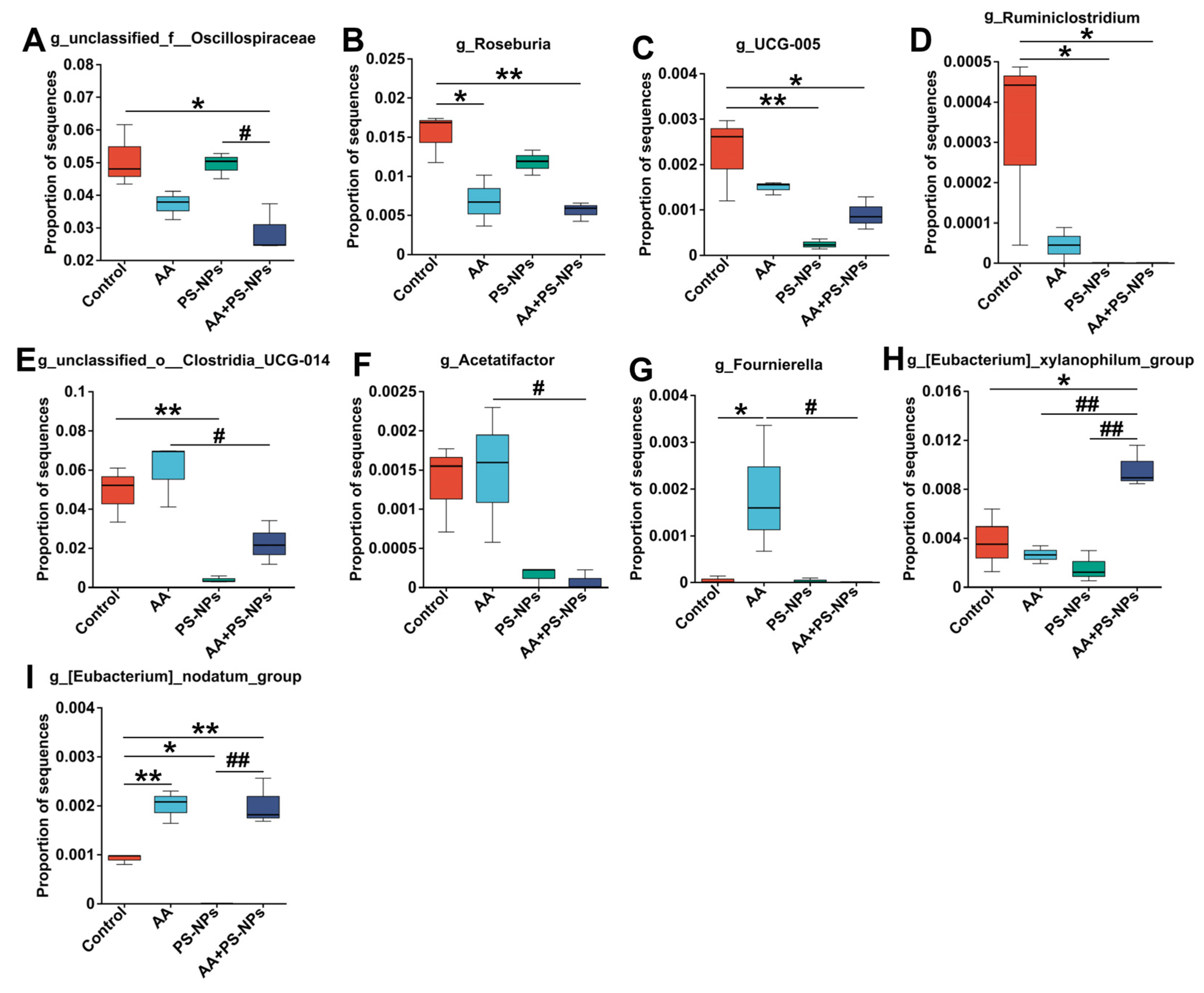
3.5. Co-Exposure to AA and PS-NPs Changes Cecal Microbiota in Mice
3.6. Correlation Analysis Between Gut Microbiota and Related Intestinal–Liver Indicators
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aktağ, I.G.; Hamzalıoğlu, A.; Kocadağlı, T.; Gökmen, V. Dietary exposure to acrylamide: A critical appraisal on the conversion of disregarded intermediates into acrylamide and possible reactions during digestion. Curr. Res. Food Sci. 2022, 5, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- European Union. Commission Regulation (EU) 2017/2158 of 20 November 2017 Establishing Mitigation Measures and Benchmark Levels for the Reduction of the Presence of Acrylamide in food. Off. J. Eur. Union. 2017, 304, 24–44. [Google Scholar]
- Žilić, S.; Nikolić, V.; Mogol, B.A.; Hamzalıoğlu, A.; Taş, N.G.; Kocadağlı, T.; Simić, M.; Gökmen, V. Acrylamide in corn-based thermally processed foods: A review. J. Agric. Food Chem. 2022, 70, 4165–4181. [Google Scholar] [CrossRef]
- Esposito, F.; Squillante, J.; Nolasco, A.; Montuori, P.; Macrì, P.G.; Cirillo, T. Acrylamide levels in smoke from conventional cigarettes and heated tobacco products and exposure assessment in habitual smokers. Environ. Res. 2022, 208, 112659. [Google Scholar] [CrossRef]
- Li, M.; Lin, Y.L.; Zhang, T.Y.; Xu, B.; Zhang, M.S.; Chu, W.H.; Tang, Y.L. Enhancing trace acrylamide analysis by bromine derivatization coupled with direct-immersion solid-phase microextraction in drinking water. Environ. Technol. 2021, 42, 2768–2775. [Google Scholar] [CrossRef]
- Bellicha, A.; Wendeu-Foyet, G.; Coumoul, X.; Koual, M.; Pierre, F.; Guéraud, F.; Zelek, L.; Debras, C.; Srour, B.; Sellem, L.; et al. Dietary exposure to acrylamide and breast cancer risk: Results from the NutriNet-Santé cohort. Am. J. Clin. Nutr. 2022, 116, 911–919. [Google Scholar] [CrossRef]
- Hosseini-Esfahani, F.; Beheshti, N.; Nematollahi, A.; Koochakpoor, G.; Verij-Kazemi, S.; Mirmiran, P.; Azizi, F. The association between dietary acrylamide intake and the risk of type 2 diabetes incidence in the Tehran lipid and glucose study. Sci. Rep. 2023, 13, 8235. [Google Scholar] [CrossRef]
- Wang, A.; Wan, X.; Zhuang, P.; Jia, W.; Ao, Y.; Liu, X.; Tian, Y.; Zhu, L.; Huang, Y.; Yao, J.; et al. High fried food consumption impacts anxiety and depression due to lipid metabolism disturbance and neuroinflammation. Proc. Natl. Acad. Sci. USA 2023, 120, e2221097120. [Google Scholar] [CrossRef]
- Wang, B.; Wang, X.; Yu, L.; Liu, W.; Song, J.; Fan, L.; Zhou, M.; Yang, M.; Ma, J.; Cheng, M.; et al. Acrylamide exposure increases cardiovascular risk of general adult population probably by inducing oxidative stress, inflammation, and TGF-β1: A prospective cohort study. Environ. Int. 2022, 164, 107261. [Google Scholar] [CrossRef]
- Govindaraju, I.; Sana, M.; Chakraborty, I.; Rahman, M.H.; Biswas, R.; Mazumder, N. Dietary acrylamide: A detailed review on formation, detection, mitigation, and its health impacts. Foods 2024, 13, 556. [Google Scholar] [CrossRef]
- Yue, Z.; Chen, Y.; Song, Y.; Zhang, J.; Yang, X.; Wang, J.; Li, L.; Sun, Z. Effect of acrylamide on glucose homeostasis in female rats and its mechanisms. Food Chem. Toxicol. 2020, 135, 110894. [Google Scholar] [CrossRef] [PubMed]
- Yue, Z.; Chen, Y.; Dong, Q.; Li, D.; Guo, M.; Zhang, L.; Shi, Y.; Wu, H.; Li, L.; Sun, Z. Acrylamide induced glucose metabolism disorder in rats involves gut microbiota dysbiosis and changed bile acids metabolism. Food Res. Int. 2022, 157, 111405. [Google Scholar] [CrossRef] [PubMed]
- Yue, Z.; Zhao, F.; Guo, Y.; Zhang, Y.; Chen, Y.; He, L.; Li, L. Lactobacillus reuteri JCM 1112 ameliorates chronic acrylamide-induced glucose metabolism disorder via the bile acid–TGR5–GLP-1 axis and modulates intestinal oxidative stress in mice. Food Func. 2024, 15, 6450–6458. [Google Scholar] [CrossRef] [PubMed]
- Peivasteh-Roudsari, L.; Karami, M.; Barzegar-Bafrouei, R.; Samiee, S.; Karami, H.; Tajdar-Oranj, B.; Mahdavi, V.; Alizadeh, A.M.; Sadighara, P.; Oliveri Conti, G.; et al. Toxicity, metabolism, and mitigation strategies of acrylamide: A comprehensive review. Int. J. Environ. Health Res. 2024, 34, 1–29. [Google Scholar] [CrossRef]
- Sewwandi, M.; Wijesekara, H.; Rajapaksha, A.U.; Soysa, S.; Vithanage, M. Microplastics and plastics-associated contaminants in food and beverages; Global trends, concentrations, and human exposure. Environ. Pollut. 2023, 317, 120747. [Google Scholar] [CrossRef]
- Hussain, K.A.; Romanova, S.; Okur, I.; Zhang, D.; Kuebler, J.; Huang, X.; Wang, B.; Fernandez-Ballester, L.; Lu, Y.; Schubert, M.; et al. Assessing the release of microplastics and nanoplastics from plastic containers and reusable food pouches: Implications for human health. Environ. Sci. Technol. 2023, 57, 9782–9792. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Bahmid, N.A.; Salman, S.H.M.; Nawaz, A.; Walayat, N.; Shekhawat, G.K.; Gvozdenko, A.A.; Blinov, A.V.; Nagdalian, A.A. Migration of microplastics from plastic packaging into foods and its potential threats on human health. Adv. Food Nutr. Res. 2023, 103, 313–359. [Google Scholar] [CrossRef]
- Son, J.W.; Nam, Y.; Kim, C. Nanoplastics from disposable paper cups and microwavable food containers. J. Hazard. Mater. 2024, 464, 133014. [Google Scholar] [CrossRef]
- Chen, Y.; Meng, Y.; Liu, G.; Huang, X.; Chai, G. Probabilistic estimation of airborne micro- and nanoplastic intake in humans. Environ. Sci. Technol. 2024, 58, 9071–9081. [Google Scholar] [CrossRef]
- Chen, C.Y.; Lin, Z. Exploring the potential and challenges of developing physiologically-based toxicokinetic models to support human health risk assessment of microplastic and nanoplastic particles. Environ. Int. 2024, 186, 108617. [Google Scholar] [CrossRef]
- Wu, P.; Lin, S.; Cao, G.; Wu, J.; Jin, H.; Wang, C.; Wong, M.H.; Yang, Z.; Cai, Z. Absorption, distribution, metabolism, excretion and toxicity of microplastics in the human body and health implications. J. Hazard. Mater. 2022, 437, 129361. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Liu, Y.; Zhang, T.; Zhang, F.; Ren, H.; Zhang, Y. Analysis of Microplastics in Human Feces Reveals a Correlation between Fecal Microplastics and Inflammatory Bowel Disease Status. Environ. Sci. Technol. 2022, 56, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K.; Ma, X.; Lichtfouse, E.; Robert, D. Nanoplastics are potentially more dangerous than microplastics. Environ. Chem. Lett. 2023, 21, 1933–1936. [Google Scholar] [CrossRef]
- Yang, H.; Kong, L.; Chen, Z.; Wu, J. Effect of functional groups of polystyrene nanoplastics on the neurodevelopmental toxicity of acrylamide in the early life stage of zebrafish. Aquat. Toxicol. 2025, 278, 107177. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Schnabl, B. The gut-liver axis and gut microbiota in health and liver disease. Nat. Rev. Microbiol. 2023, 21, 719–733. [Google Scholar] [CrossRef]
- Amedei, A.; Gitto, S.; Campani, C.; Marra, F. Probiotics and the gut-liver axis. In Probiotics: Advanced Food and Health Applications; Brandelli, A., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 467–481. [Google Scholar]
- Wen, S.; Zhao, Y.; Liu, S.; Chen, Y.; Yuan, H.; Xu, H. Polystyrene microplastics exacerbated liver injury from cyclophosphamide in mice: Insight into gut microbiota. Sci. Total Environ. 2022, 840, 156668. [Google Scholar] [CrossRef]
- Sánchez, J.; Cabrer, J.M.; Rosselló, C.A.; Palou, A.; Picó, C. Formation of hemoglobin adducts of acrylamide after its ingestion in rats is dependent on age and sex. J. Agric. Food Chem. 2008, 56, 5096–5101. [Google Scholar] [CrossRef]
- Senathirajah, K.; Attwood, S.; Bhagwat, G.; Carbery, M.; Wilson, S.; Palanisami, T. Estimation of the mass of microplastics ingested—A pivotal first step towards human health risk assessment. J. Hazard. Mater. 2021, 404, 124004. [Google Scholar] [CrossRef]
- Wang, Y.L.; Lin, S.X.; Wang, Y.; Liang, T.; Jiang, T.; Liu, P.; Li, X.Y.; Lang, D.Q.; Liu, Q.; Shen, C.Y. p-Synephrine ameliorates alloxan-induced diabetes mellitus through inhibiting oxidative stress and inflammation via suppressing the NF-kappa B and MAPK pathways. Food Funct. 2023, 14, 1971–1988. [Google Scholar] [CrossRef]
- Guo, J.; Han, X.; Tan, H.; Huang, W.; You, Y.; Zhan, J. Blueberry extract improves obesity through regulation of the gut microbiota and bile acids via pathways involving FXR and TGR5. iScience 2019, 19, 676–690. [Google Scholar] [CrossRef]
- Tang, K.H.D.; Li, R.; Li, Z.; Wang, D. Health risk of human exposure to microplastics: A review. Environ. Chem. Lett. 2024, 22, 1155–1183. [Google Scholar] [CrossRef]
- Bridgeman, L.; Juan, C.; Juan-García, A.; Berrada, H. Individual and combined effect of acrylamide, fumitremorgin C and penitrem A on human neuroblastoma SH-SY5Y cells. Food Chem. Toxicol. 2023, 182, 114114. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Lu, J.; Nie, C.; Guo, Z.; Li, C.; Yu, Q.; Xie, J.; Chen, Y. Combined effects of acrylamide and ochratoxin A on the intestinal barrier in Caco-2 cells. Foods 2023, 12, 1318. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Qiu, W.; Pang, X.; Su, Y.; Zhang, X.; Huang, J.; Xie, H.; Liao, J.; Tang, Z.; Chen, Z.; et al. Polystyrene nanoplastics and cadmium co-exposure aggravated cardiomyocyte damage in mice by regulating PANoptosis pathway. Environ. Pollut. 2024, 347, 123713. [Google Scholar] [CrossRef]
- Zhang, K.K.; Wan, J.Y.; Chen, Y.C.; Cheng, C.H.; Zhou, H.Q.; Zheng, D.K.; Lan, Z.X.; You, Q.H.; Sun, J. Polystyrene nanoplastics exacerbate aflatoxin B1-induced hepatic injuries by modulating the gut-liver axis. Sci. Total Environ. 2024, 935, 173285. [Google Scholar] [CrossRef]
- Choi, Y.J.; Park, J.W.; Lim, Y.; Seo, S.; Hwang, D.Y. In vivo impact assessment of orally administered polystyrene nanoplastics: Biodistribution, toxicity, and inflammatory response in mice. Nanotoxicology 2021, 15, 1180–1198. [Google Scholar] [CrossRef]
- Yuan, Y.; Li, L.; Nan, B.; Yang, C.; Yan, H. Allicin ameliorates intestinal barrier damage via microbiota-regulated short-chain fatty acids-TLR4/MyD88/NF-κB cascade response in acrylamide-induced rats. J. Agric. Food Chem. 2021, 69, 12837–12852. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert Rev. Gastroent. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef]
- Yang, D.; Jacobson, A.; Meerschaert, K.A.; Sifakis, J.J.; Wu, M.; Chen, X.; Yang, T.; Zhou, Y.; Anekal, P.V.; Rucker, R.A.; et al. Nociceptor neurons direct goblet cells via a CGRP-RAMP1 axis to drive mucus production and gut barrier protection. Cell 2022, 185, 4190–4205. [Google Scholar] [CrossRef]
- Zheng, D.; Liao, H.; Chen, S.; Liu, X.; Mao, C.; Zhang, C.; Meng, M.; Wang, Z.; Wang, Y.; Jiang, Q.; et al. Elevated levels of circulating biomarkers related to leaky gut syndrome and bacterial translocation are associated with Graves’ disease. Front. Endocrinol. 2021, 12, 796212. [Google Scholar] [CrossRef] [PubMed]
- Slifer, Z.M.; Blikslager, A.T. The integral role of tight junction proteins in the repair of injured intestinal epithelium. Int. J. Mol. Sci. 2020, 21, 972. [Google Scholar] [CrossRef] [PubMed]
- An, L.; Wirth, U.; Koch, D.; Schirren, M.; Drefs, M.; Koliogiannis, D.; Nieß, H.; Andrassy, J.; Guba, M.; Bazhin, A.V.; et al. The role of gut-derived lipopolysaccharides and the intestinal barrier in fatty liver diseases. J. Gastrointest. Surg. 2022, 26, 671–683. [Google Scholar] [CrossRef]
- Xiong, X.; Ren, Y.; Cui, Y.; Li, R.; Wang, C.; Zhang, Y. Obeticholic acid protects mice against lipopolysaccharide-induced liver injury and inflammation. Biomed. Pharmacother. 2017, 96, 1292–1298. [Google Scholar] [CrossRef]
- Ozer, J.; Ratner, M.; Shaw, M.; Bailey, W.; Schomaker, S. The current state of serum biomarkers of hepatotoxicity. Toxicology 2008, 245, 194–205. [Google Scholar] [CrossRef]
- Shi, C.; Han, X.; Guo, W.; Wu, Q.; Yang, X.; Wang, Y.; Tang, G.; Wang, S.; Wang, Z.; Liu, Y.; et al. Disturbed Gut-Liver axis indicating oral exposure to polystyrene microplastic potentially increases the risk of insulin resistance. Environ. Int. 2022, 164, 107273. [Google Scholar] [CrossRef]
- Sun, W.; Yan, S.; Meng, Z.; Tian, S.; Jia, M.; Huang, S.; Wang, Y.; Zhou, Z.; Diao, J.; Zhu, W. Combined ingestion of polystyrene microplastics and epoxiconazole increases health risk to mice: Based on their synergistic bioaccumulation in vivo. Environ. Int. 2022, 166, 107391. [Google Scholar] [CrossRef]
- Xue, C.; Li, G.; Zheng, Q.; Gu, X.; Shi, Q.; Su, Y.; Chu, Q.; Yuan, X.; Bao, Z.; Lu, J.; et al. Tryptophan metabolism in health and disease. Cell Metab. 2023, 35, 1304–1326. [Google Scholar] [CrossRef]
- Holeček, M. Histidine in health and disease: Metabolism, physiological importance, and use as a supplement. Nutrients 2020, 12, 848. [Google Scholar] [CrossRef]
- Branco, A.C.C.C.; Yoshikawa, F.S.Y.; Pietrobon, A.J.; Sato, M.N. Role of histamine in modulating the immune response and inflammation. Mediat. Inflamm. 2018, 2018, 9524075. [Google Scholar] [CrossRef]
- Fernstrom, J.D.; Fernstrom, M.H. Tyrosine, phenylalanine, and catecholamine synthesis and function in the brain. J. Nutr. 2007, 137, 1539S–1548S. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, Y.; Arisaka, H.; Yoshida, S.; Mori, M.; Takahashi, M. Proline protects liver from D-galactosamine hepatitis by activating the IL-6/STAT3 survival signaling pathway. Amino Acids 2012, 43, 2371–2380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tan, W.; Wang, X.; Zheng, X.; Huang, Y.; Li, B.; Meng, Z.; Gao, Y.; Qian, Z.; Liu, F.; et al. Chronic Liver Failure Consortium (Ch-CLIF.C). Metabolic biomarkers significantly enhance the prediction of HBV-related ACLF occurrence and outcomes. J. Hepatol. 2023, 79, 1159–1171. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, X.; Wang, M.; Chang, Y.; Zhang, F.; Ban, Z.; Tang, R.; Gan, Q.; Wu, S.; Guo, Y.; et al. The lysine catabolite saccharopine impairs development by disrupting mitochondrial homeostasis. J. Cell Biol. 2019, 218, 580–597. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Wei, X.; Hu, H.; Zhang, B.; Yang, D.; Du, H.; Zhu, R.; Sun, X.; Oh, Y.; Gu, N. Effects of oral administration of polystyrene nanoplastics on plasma glucose metabolism in mice. Chemosphere 2022, 288, 132607. [Google Scholar] [CrossRef]
- Forouzanfar, F.; Guest, P.C.; Jamialahmadi, T.; Sahebkar, A. Hepatoprotective effect of trehalose: Insight into its mechanisms of action. In Natural Products and Human Diseases; Sahebkar, A., Sathyapalan, T., Eds.; Springer: Cham, Switzerland, 2021; pp. 489–500. [Google Scholar]
- Conte, F.; van Buuringen, N.; Voermans, N.C.; Lefeber, D.J. Galactose in human metabolism, glycosylation and congenital metabolic diseases: Time for a closer look. Bba-Gen. Subjects 2021, 1865, 129898. [Google Scholar] [CrossRef]
- Succoio, M.; Sacchettini, R.; Rossi, A.; Parenti, G.; Ruoppolo, M. Galactosemia: Biochemistry, molecular genetics, newborn screening, and treatment. Biomolecules 2022, 12, 968. [Google Scholar] [CrossRef]
- Martínez-Reyes, I.; Chandel, N.S. Mitochondrial TCA cycle metabolites control physiology and disease. Nat. Commun. 2020, 11, 102. [Google Scholar] [CrossRef]
- Song, Y.; Hu, T.; Gao, H.; Zhai, J.; Gong, J.; Zhang, Y.; Tao, L.; Sun, J.; Li, Z.; Qu, X. Altered metabolic profiles and biomarkers associated with astragaloside IV-mediated protection against cisplatin-induced acute kidney injury in rats: An HPLC-TOF/MS-based untargeted metabolomics study. Biochem. Pharmacol. 2021, 183, 114299. [Google Scholar] [CrossRef]
- Dabke, K.; Hendrick, G.; Devkota, S. The gut microbiome and metabolic syndrome. J. Clin. Investig. 2019, 129, 4050–4057. [Google Scholar] [CrossRef]
- Qiu, P.; Ishimoto, T.; Fu, L.; Zhang, J.; Zhang, Z.; Liu, Y. The Gut microbiota in inflammatory bowel disease. Front. Cell. Infect. Microbiol. 2022, 12, 733992. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Sun, G.; Duan, J.; Luo, C.; Yangji, C.; Zhong, R.; Chen, L.; Zhu, Y.; Wangdui, B.; Zhang, H. Alterations in gut microbiota improve SCFA production and fiber utilization in Tibetan pigs fed alfalfa diet. Front. Microbiol. 2022, 13, 969524. [Google Scholar] [CrossRef]
- Hu, W.; Huang, L.; Zhou, Z.; Yin, L.; Tang, J. Diallyl disulfide (DADS) ameliorates intestinal candida albicans infection by modulating the gut microbiota and metabolites and providing intestinal protection in mice. Front. Cell. Infect. Microbiol. 2022, 11, 743454. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Pan, Y.; Liu, L.; Liang, J.; Wu, L.; Zhu, H.; Zhang, P. High salinity slowed organic acid production from acidogenic fermentation of kitchen wastewater by shaping functional bacterial community. J. Environ. Manag. 2022, 310, 114765. [Google Scholar] [CrossRef]
- Pfeiffer, N.; Desmarchelier, C.; Blaut, M.; Daniel, H.; Haller, D.; Clavel, T. Acetatifactor muris gen. nov., sp nov., a novel bacterium isolated from the intestine of an obese mouse. Arch. Microbiol. 2012, 194, 901–907. [Google Scholar] [CrossRef]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia spp.: A marker of health? Future Microbiol. 2017, 12, 157–170. [Google Scholar] [CrossRef]
- Yao, Y.; Cai, X.; Fei, W.; Ye, Y.; Zhao, M.; Zheng, C. The role of short-chain fatty acids in immunity, inflammation and metabolism. Crit. Rev. Food Sci. 2022, 62, 1–12. [Google Scholar] [CrossRef]
- Duan, Y.; Guo, F.; Li, C.; Xiang, D.; Gong, M.; Yi, H.; Chen, L.; Yan, L.; Zhang, D.; Dai, L.; et al. Aqueous extract of fermented Eucommia ulmoides leaves alleviates hyperlipidemia by maintaining gut homeostasis and modulating metabolism in high-fat diet fed rats. Phytomedicine 2024, 128, 155291. [Google Scholar] [CrossRef]
- Liu, Y.; Feng, Y.; Yang, X.; Lv, Z.; Li, P.; Zhang, M.; Wei, F.; Jin, X.; Hu, Y.; Guo, Y.; et al. Mining chicken ileal microbiota for immunomodulatory microorganisms. ISME J. 2023, 17, 758–774. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, M.; Yang, M.; Jin, C.; Song, Y.; Chen, J.; Gao, M.; Ai, Z.; Su, D. Pulsatilla chinensis saponins ameliorate inflammation and DSS-induced ulcerative colitis in rats by regulating the composition and diversity of intestinal flora. Front. Cell. Infect. Microbiol. 2021, 11, 728929. [Google Scholar] [CrossRef]
- Fu, R.; Niu, R.; Li, R.; Yue, B.; Zhang, X.; Cao, Q.; Wang, J.; Sun, Z. Fluoride-induced alteration in the diversity and composition of bacterial microbiota in mice colon. Biol. Trace Elem. Res. 2020, 196, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhao, X.; Liu, T.; Wang, Y.; Wang, J.; Kong, L.; Zhao, Q.; Chen, Y.; Chen, L.; Zhang, H. Xuanfei Baidu decoction attenuates intestinal disorders by modulating NF-κB pathway, regulating T cell immunity and improving intestinal flora. Phytomedicine 2022, 101, 154100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wan, Y.; Zhou, X.; Zhang, H.; Zhao, H.; Ma, L.; Dong, X.; Yan, M.; Zhao, T.; Li, P. Characteristics of serum metabolites and gut microbiota in diabetic kidney disease. Front. Pharmacol. 2022, 13, 872988. [Google Scholar] [CrossRef]
- He, F.; Shi, H.; Liu, R.; Tian, G.; Qi, Y.; Wang, T. Randomly-shaped nanoplastics induced stronger biotoxicity targeted to earthworm Eisenia fetida species: Differential effects and the underlying mechanisms of realistic and commercial polystyrene nanoplastics. Sci. Total Environ. 2023, 877, 162854. [Google Scholar] [CrossRef] [PubMed]
- The Joint FAO/WHO Expert Committee on Food Additives (JECFA). Evaluation of Certain Food Additives and Contaminants: Seventy-Second Report of the Joint FAO/WHO Expert Committee on Food Additive; WHO Technical Report Series; WHO: Geneva, Switzerland, 2011; p. 959. [Google Scholar]
- Tardiff, R.G.; Gargas, M.L.; Kirman, C.R.; Carson, M.L.; Sweeney, L.M. Estimation of safe dietary intake levels of acrylamide for humans. Food Chem. Toxicol. 2010, 48, 658–667. [Google Scholar] [CrossRef]
- The Joint FAO/WHO Expert Committee on Food Additives (JECFA). Evaluation of Certain Food Additives and Contaminants: Sixty-Fourth Report of the Joint FAO/WHO Expert Committee on Food Additive; WHO Technical Report Series; WHO: Geneva, Switzerland, 2005; p. 930. [Google Scholar]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef]
- Chen, S.N.; Tan, Y.; Xiao, X.C.; Li, Q.; Wu, Q.; Peng, Y.Y.; Ren, J.; Dong, M.L. Deletion of TLR4 attenuates lipopolysaccharide-induced acute liver injury by inhibiting inflammation and apoptosis. Acta Pharmacol. Sin. 2021, 42, 1610–1619. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Li, W.; Zhang, B.; Yin, J.; Liuqi, S.; Wang, J.; Peng, B.; Wang, S. Fucoidan ameliorated dextran sulfate sodium-induced ulcerative colitis by modulating gut microbiota and bile acid metabolism. J. Agric. Food Chem. 2022, 70, 14864–14876. [Google Scholar] [CrossRef]
- Zhao, Z.; Ning, J.; Bao, X.Q.; Shang, M.; Ma, J.; Li, G.; Zhang, D. Fecal microbiota transplantation protects rotenone-induced Parkinson’s disease mice via suppressing inflammation mediated by the lipopolysaccharide-TLR4 signaling pathway through the microbiota-gut-brain axis. Microbiome 2021, 9, 226. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | M/Z | Control vs. AA | Control vs. PS-NPs | Control vs. AA + PS-NPs | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| VIP | FC | Regulate | VIP | FC | Regulate | VIP | FC | Regulate | ||
| Amino acids metabolism | ||||||||||
| Kynurenine | 207.079 | 1.280 | 0.958 | ↓ | 1.280 | 0.958 | ↓ | 1.310 | 0.925 | ↓ |
| Anthranilic acid | 136.041 | 1.702 | 0.936 | ↓ | 1.702 | 0.936 | ↓ | 1.854 | 0.854 | ↓ |
| Kynurenic acid | 190.049 | 1.886 | 0.931 | ↓ | 2.246 | 0.919 | ↓ | 1.864 | 0.879 | ↓ |
| L-Histidine | 156.076 | 1.549 | 1.036 | ↑ | 1.549 | 1.036 | ↑ | 1.327 | 1.053 | ↑ |
| Histamine | 110.073 | 1.222 | 1.051 | ↑ | 1.222 | 1.051 | ↑ | |||
| Urocanic acid | 137.036 | 1.284 | 1.046 | ↑ | 1.284 | 1.046 | ↑ | |||
| Serotonin | 177.101 | 1.526 | 1.057 | ↑ | 1.914 | 1.142 | ↑ | |||
| 5-Hydroxytryptophan | 219.079 | 1.199 | 1.059 | ↑ | ||||||
| Indole-3-acetamide | 175.086 | 1.888 | 1.106 | ↑ | ||||||
| Indole-3-acetaldoxime | 175.085 | 2.405 | 1.301 | ↑ | ||||||
| Indole-3-acetic acid | 158.059 | 2.052 | 1.142 | ↑ | ||||||
| Ergothioneine | 230.094 | 1.638 | 1.095 | ↑ | ||||||
| Methionine sulfoxide | 166.052 | 1.487 | 1.071 | ↑ | ||||||
| Saccharopine | 318.163 | 1.182 | 1.057 | ↑ | ||||||
| Phenylalanine | 331.163 | 1.525 | 0.911 | ↓ | ||||||
| L-proline | 116.070 | 1.017 | 0.977 | ↓ | ||||||
| Carbohydrate metabolism | ||||||||||
| Lactose | 377.087 | 2.033 | 1.126 | ↑ | 2.642 | 1.175 | ↑ | 2.279 | 1.233 | ↑ |
| D-Glucose | 225.063 | 1.050 | 1.038 | ↑ | 1.700 | 1.073 | ↑ | 1.265 | 1.081 | ↑ |
| UDP-D-galactose | 565.050 | 1.850 | 1.080 | ↑ | ||||||
| UDP-glucose | 565.050 | 1.792 | 1.099 | ↑ | ||||||
| UDP-N-acetylglucosamine | 606.077 | 1.535 | 1.059 | ↑ | ||||||
| Glucosamine | 162.075 | 1.092 | 1.024 | ↑ | ||||||
| Stachyose | 665.217 | 4.615 | 1.640 | ↑ | 3.423 | 1.686 | ↑ | |||
| Dextran | 549.169 | 4.026 | 1.335 | ↑ | 3.156 | 1.393 | ↑ | |||
| Levan | 487.163 | 3.875 | 1.243 | ↑ | 3.075 | 1.324 | ↑ | |||
| Trehalose | 325.111 | 3.726 | 1.199 | ↑ | 2.612 | 1.218 | ↑ | |||
| Galactinol | 387.116 | 2.110 | 1.108 | ↑ | 1.620 | 1.118 | ↑ | |||
| Citric acid | 191.021 | 1.429 | 0.909 | ↓ | ||||||
| Isocitric acid | 191.021 | 1.345 | 0.924 | ↓ | ||||||
| Glyceric acid | 105.020 | 1.297 | 0.929 | ↓ | ||||||
| 2-Ketoglutaric acid | 145.015 | 1.139 | 0.918 | ↓ | ||||||
| Glycolic acid | 75.009 | 1.069 | 0.955 | ↓ | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Luo, R.; Sun, Z.; Zhang, Y.; Guo, Y.; Chen, Y.; Li, L.; Yue, Z. Synergistic Toxicity of Combined Exposure to Acrylamide and Polystyrene Nanoplastics on the Gut–Liver Axis in Mice. Biology 2025, 14, 523. https://doi.org/10.3390/biology14050523
Liu Y, Luo R, Sun Z, Zhang Y, Guo Y, Chen Y, Li L, Yue Z. Synergistic Toxicity of Combined Exposure to Acrylamide and Polystyrene Nanoplastics on the Gut–Liver Axis in Mice. Biology. 2025; 14(5):523. https://doi.org/10.3390/biology14050523
Chicago/Turabian StyleLiu, Yongchuang, Ruiping Luo, Zhongke Sun, Yidan Zhang, Yuqi Guo, Yanjuan Chen, Lili Li, and Zonghao Yue. 2025. "Synergistic Toxicity of Combined Exposure to Acrylamide and Polystyrene Nanoplastics on the Gut–Liver Axis in Mice" Biology 14, no. 5: 523. https://doi.org/10.3390/biology14050523
APA StyleLiu, Y., Luo, R., Sun, Z., Zhang, Y., Guo, Y., Chen, Y., Li, L., & Yue, Z. (2025). Synergistic Toxicity of Combined Exposure to Acrylamide and Polystyrene Nanoplastics on the Gut–Liver Axis in Mice. Biology, 14(5), 523. https://doi.org/10.3390/biology14050523