Molecular and Morphological Characterization of the Entomopathogenic Nematode Oscheius cyrus (Nematoda: Rhabditidae) and Molecular Variability of Heterorhabditis bacteriophora from Georgia (Caucasus)

 , , ,
, , ,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Sampling and Nematode Recovery
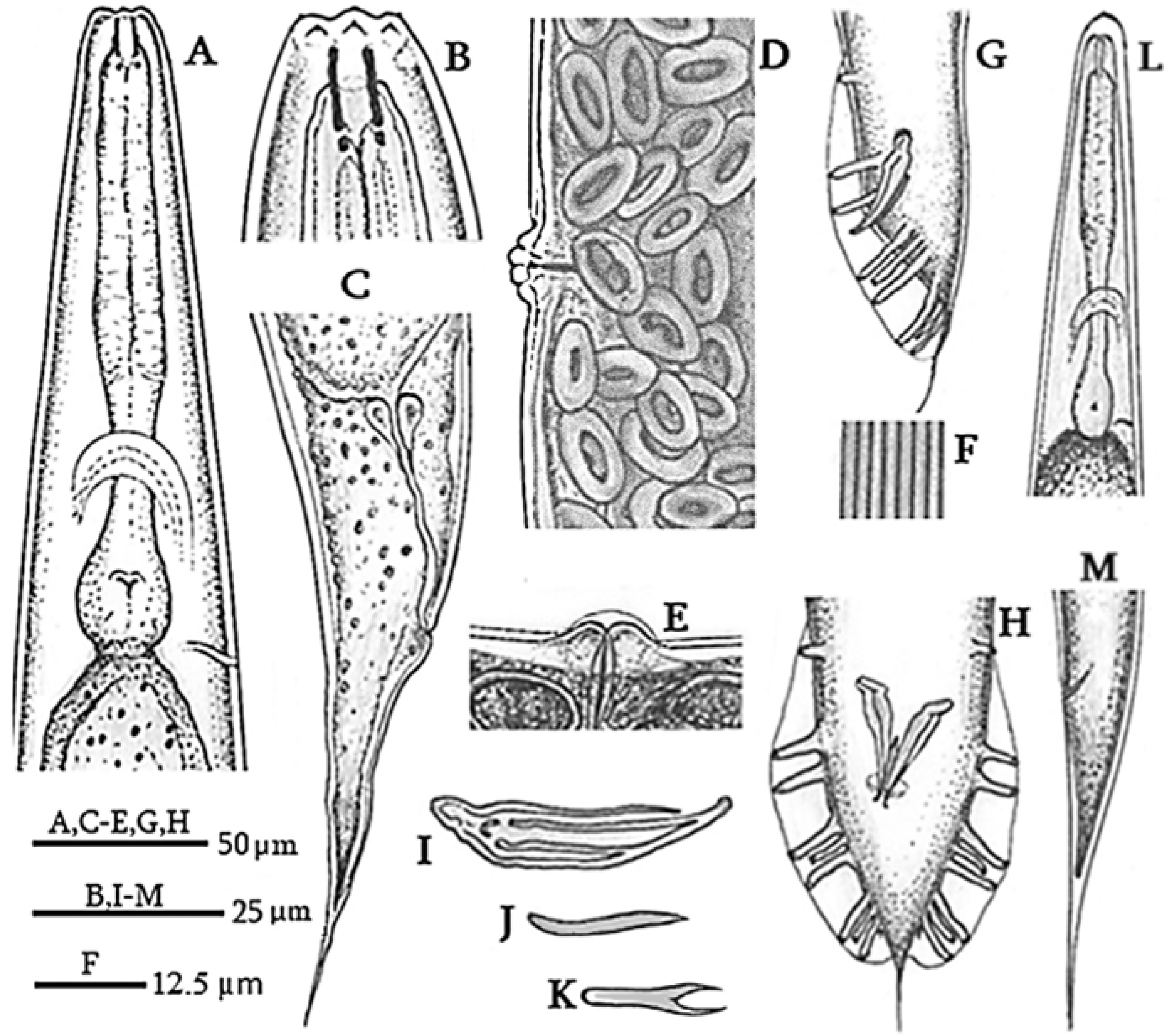
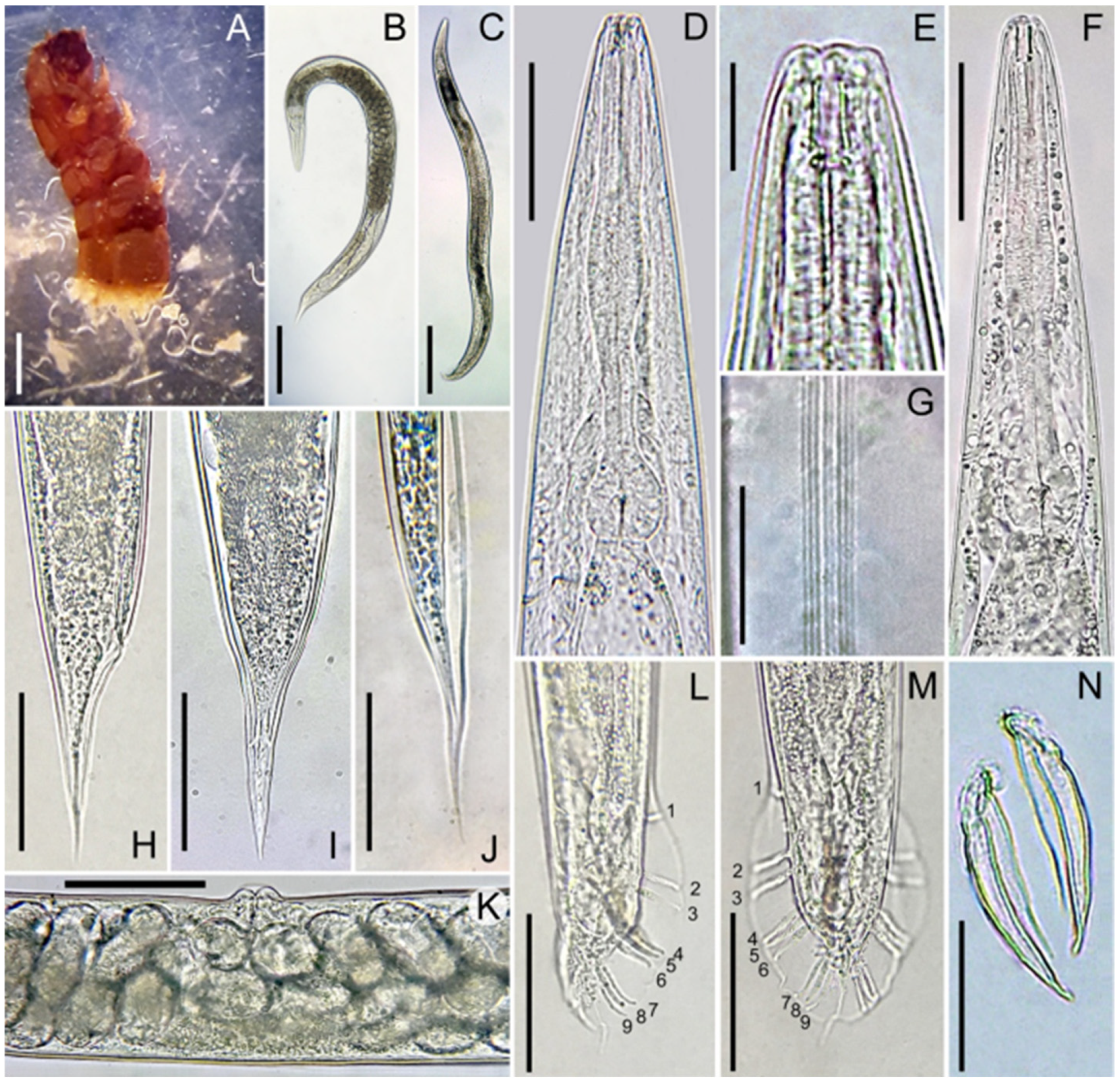
2.2. Morphological and Morphometric Identification of Oscheius cyrus
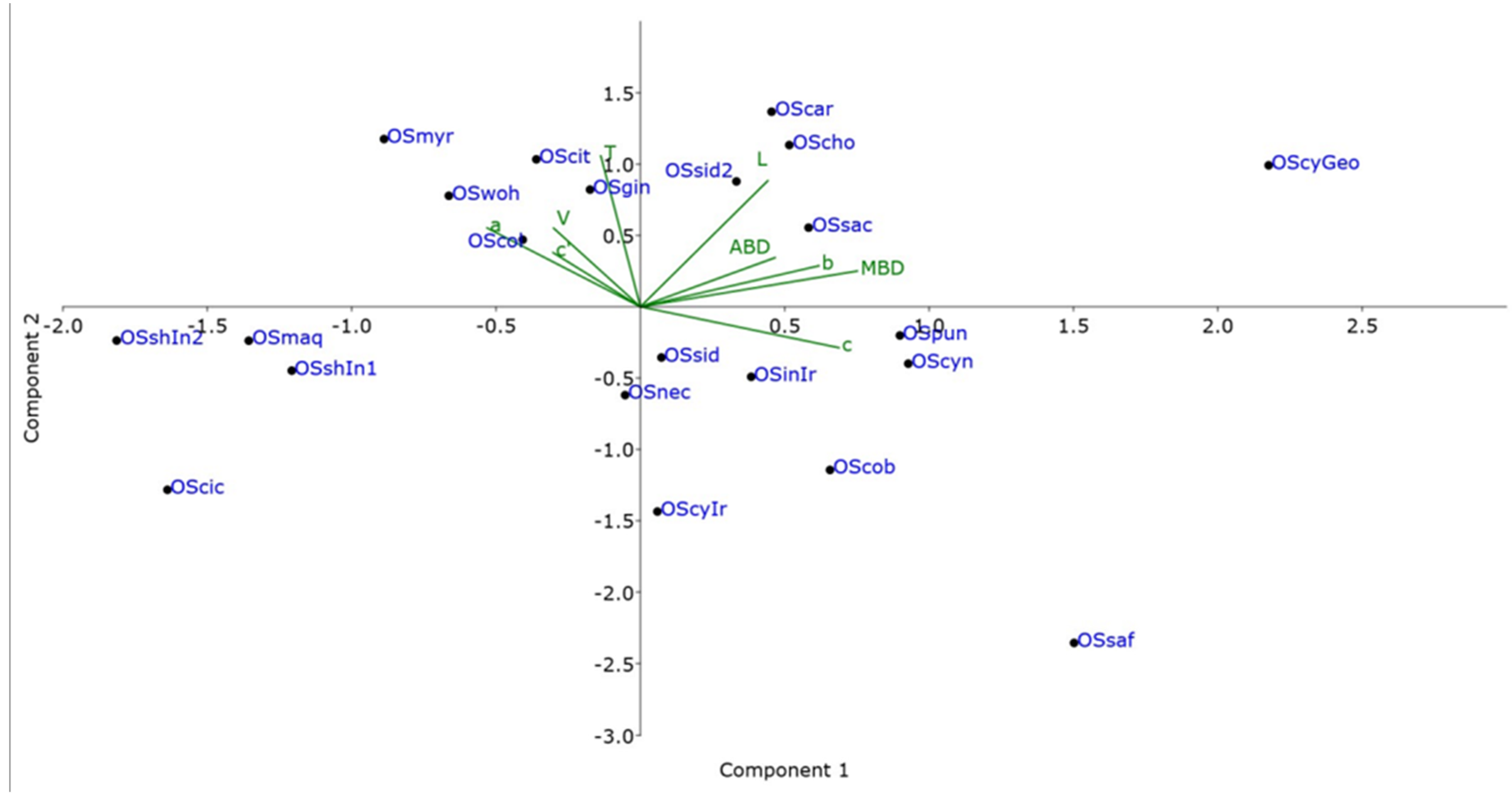
2.3. Multivariate Morphometric Analysis
2.4. Entomopathogenicity of Oscheius cyrus
2.5. Molecular Identification
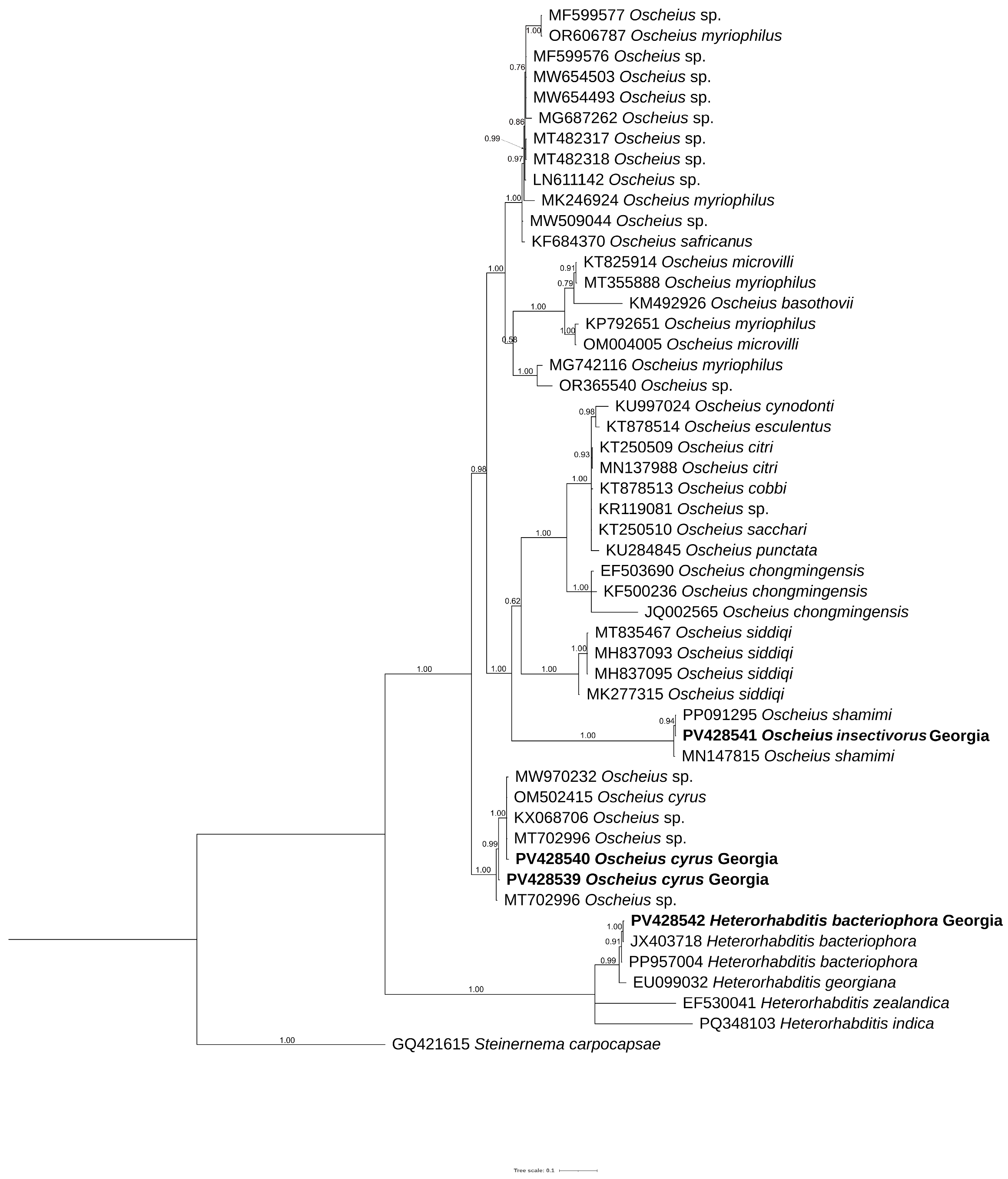
2.6. Phylogenetic Analysis
3. Results
3.1. Morphological and Morphometric Observations of Oscheius cyrus
3.2. PCA
3.3. Entomopathogenicity Test
3.4. Diagnosis and Relationships
3.5. Molecular Characterization
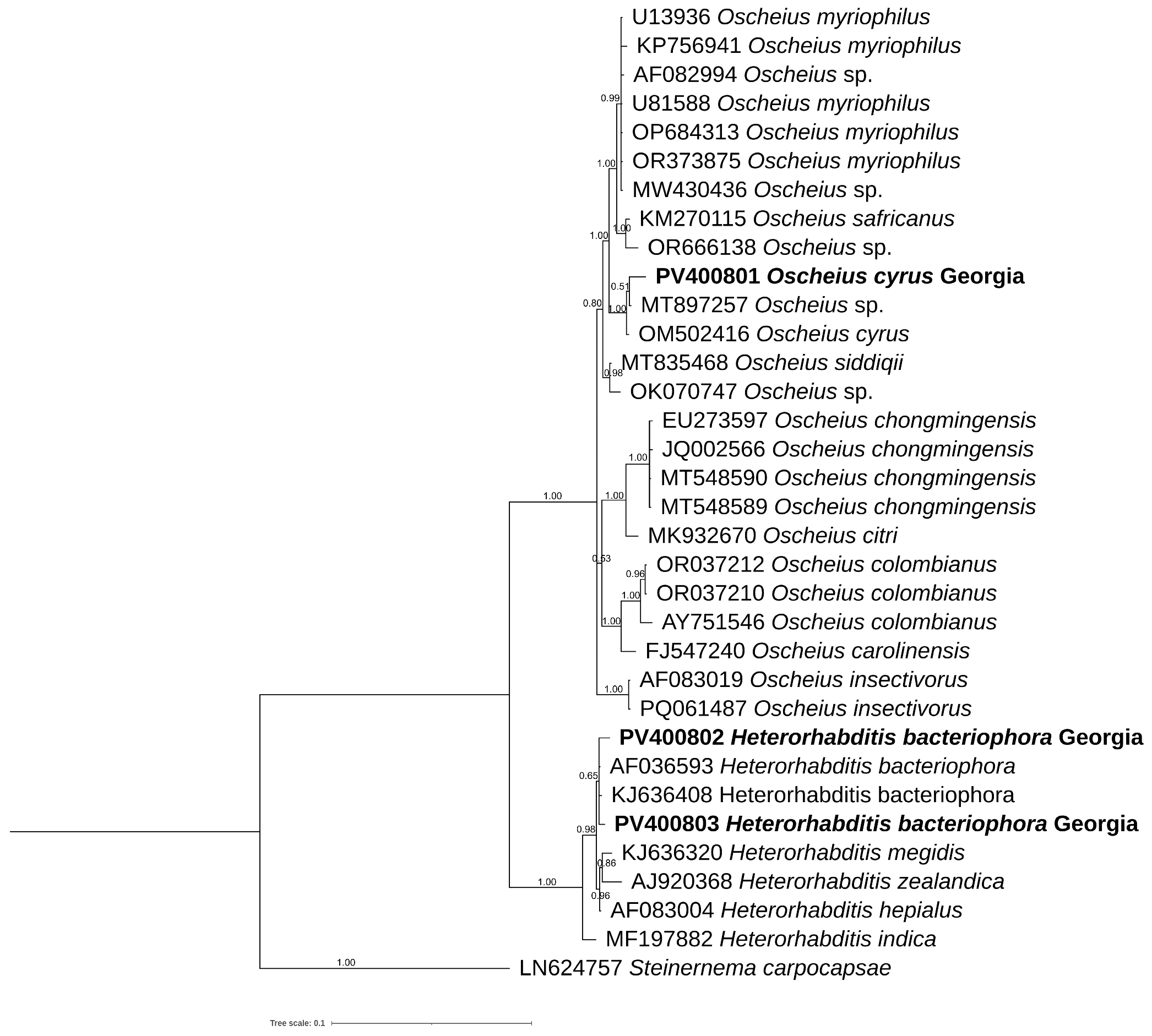
3.6. Phylogenetic Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaya, H.K.; Gaugler, R. Entomopathogenic nematodes. Ann. Rev. Entomol. 1993, 38, 181–206. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.; Han, R.; Dolinski, C. Entomopathogenic nematode production and application technology. J. Nematol. 2012, 44, 206–217. [Google Scholar] [PubMed]
- Adams, B.J.; Fodor, A.; Koppenhöfer, H.S.; Stackenbrandt, E.; Stock, S.P.; Klein, M.G. Biodiversity and systematic of nematode-bacterium entomopathogens. Biol. Control 2006, 38, 4–21. [Google Scholar] [CrossRef]
- Ye, W.; Torres-Barragan, A.; Cardoza, Y. Oscheius carolinensis n. sp. (Nematoda: Rhabditidae), a potential entomopathogenic nematode from vermicompost. Nematology 2010, 12, 121–135. [Google Scholar] [CrossRef]
- Liu, Q.; Mracek, Z.; Zhang, L.; Puza, V.; Dong, L. Re-description of Oscheius chongmingensis (Zhang et al. 2008) (Nematoda: Rhabditidae) and its entomopathogenicity. Nematology 2012, 14, 139–149. [Google Scholar] [CrossRef]
- Torrini, G.; Mazza, G.; Carletti, B.; Benvenuti, C.; Roversi, P.F.; Fanelli, E.; De Luca, F.; Troccoli, A.; Tarasco, E. Oscheius onirici sp. n. (Nematoda: Rhabditidae): A new entomopathogenic nematode from an Italian cave. Zootaxa 2015, 26, 533–548. [Google Scholar] [CrossRef]
- Serepa-Dlamini, M.H.; Gray, V.M. A new species of entomopathogenic nematode Oscheius safricana n. sp. (Nematoda: Rhabditidae) from South Africa. Arch. Phytopathol. Plant Prot. 2018, 51, 309–321. [Google Scholar] [CrossRef]
- Karimi, J.; Rezaee, N.; Shokoohi, E. Addition a new insect parasitic nematode, Oscheius tipulae to Iranian fauna. Nematropica 2018, 48, 45–54. [Google Scholar]
- Santoiemma, G.; Glazer, I.; Battisti, A.; Bianchi, A.; Fanelli, E.; Mori, N.; Sacchi, S.; Tarasco, E.; Troccoli, A.; De Luca, F. Occurrence and diversity of the entomopathogenic nematode Oscheius myriophilus (Nematoda: Rhabditida) in the Italian outbreak area of Popillia japonica (Coleoptera: Scarabaeidae). Nematology 2024, 26, 817–831. [Google Scholar] [CrossRef]
- Andrássy, I. Evolution as a Basis for the Systematization of Nematodes; Pitman Publishing: London, UK, 1976. [Google Scholar]
- Foye, S.; Steffan, S.A.; Rare, A. Recently discovered nematode, Oscheius onirici (Rhabditida: Rhabditidae), kills Drosophila suzukii (Diptera: Drosophilidae) within fruit. J. Econ. Entomol. 2020, 113, 1047–1051. [Google Scholar] [CrossRef]
- Campos-Herrera, R.; Půža, V.; Jaffuel, G.; Blanco-Pérez, R.; Čepulytė-Rakauskienė, R.; Turlings, T.C.J. Unraveling the intraguild competition between Oscheius spp. nematodes and entomopathogenic nematodes: Implications for their natural distribution in Swiss agricultural soils. J. Invert. Pathol. 2015, 132, 216–227. [Google Scholar] [CrossRef]
- Zhang, K.; Baiocchi, T.; Lu, D.; Chang, D.Z.; Dillman, A.R. Differentiating between scavengers and entomopathogenic nematodes: Which is Oscheius chongmingensis? J. Invert. Pathol. 2019, 167, 107245. [Google Scholar] [CrossRef]
- Tabassum, K.A.; Shahina, F.N.; Erum, Y.L. Description of six new species of Oscheius andrassy, 1976 (Nematoda: Rhabditida) from Pakistan with a key and diagnostic compendium to species of the genus. Pak. J. Nematol. 2016, 34, 109–161. [Google Scholar]
- Zhou, G.; Yang, H.; Wang, F.; Bao, H.; Wang, G.; Hou, X.; Lin, J.; Yedid, G.; Zhang, K. Oscheius microvilli n. sp. (Nematoda: Rhabditidae): A facultatively pathogenic nematode from Chongming Island, China. J. Nematol. 2017, 49, 33–41. [Google Scholar] [CrossRef]
- Kakulia, G.; Veremtchuk, G. A new species of nematode of Amphimallon solstitialis—Neoaplectana georgica sp. n. (Nematoda, Steinernematidae). Bull. Georgian Acad. Sci. 1965, 15, 713–718. [Google Scholar]
- Gorgadze, O. The entomopathogenic nematode new species Steinernema (Neoaplectana) disparica sp. n. (Rhabditidae, Steinernematidae) from Georgia. Bull. Georgian Acad. Sci. 2001, 164, 161–163. [Google Scholar]
- Gorgadze, O.; Lordkipanidze, M. Nematoda Steinernema gurgistana sp. n. (Rhabditida: Steinernematidae) from Agriotes gurgistana. Proc. Georgian Acad. Sci. Biol. Ser. 2006, 174, 117–122. [Google Scholar]
- Gorgadze, O.A.; Lortkhipanidze, M.G.; Ogier, J.C.; Tailliez, P.; Burjanadze, M.C. Steinernema tbilisiensis sp. n. (Nematoda: Steinernematidae), a new species of entomopathogenic nematode from Georgia. J. Agric. Sci. Technol. 2015, 5, 264–276. [Google Scholar] [CrossRef]
- Gorgadze, O.A.; Ivanova, E.S.; Lortkhipanidze, M.G.; Spiridonov, S.E. Redescription of Steinernema thesami Gorgadze, 1988 (Rhabditida: Steinernematidae) from Georgia. Russian J. Nematol. 2016, 24, 17–31. [Google Scholar]
- Gorgadze, O.; Fanelli, E.; Lortkipanidze, M.; Troccoli, A.; Burjanadze, M.; Tarasco, E.; De Luca, F. Steinernema borjomiense n. sp. (Rhabditida: Steinernematidae), a new entomopathogenic nematode from Georgia. Nematology 2018, 20, 653–669. [Google Scholar] [CrossRef]
- Gorgadze, O.; Troccoli, A.; Fanelli, E.; Tarasco, E.; De Luca, F. Characterization of a population of Pelodera strongyloides (Nematoda: Rhabditidae) associated with the beetle Lucanus ibericus (Coleoptera: Lucanidae) from Georgia. J. Nematol. 2020, 52, 1–12. [Google Scholar] [CrossRef]
- Gorgadze, O.; Troccoli, A.; Fanelli, E.; Tarasco, E.; De Luca, F. Phasmarhabditis thesamica n. sp. (Nematoda: Rhabditidae), a new slug nematode from southern slope of Caucasus, Georgia. Nematology 2022, 24, 617–629. [Google Scholar] [CrossRef]
- Kuhestani, K.; Karimi, J.; Shokoohi, E.; Makhdoumi, A. Description of Oscheius cyrus n. sp. (Nematoda: Rhabditidae) as new entomopathogenic nematode from Iran. J. Helminthol. 2022, 96, e69. [Google Scholar] [CrossRef] [PubMed]
- Bonab, A.M.; Atighi, M.R.; Ghavamabad, G.R.; Pedram, M. First report of Oscheius insectivorus (Körner, 1954) Andrássy, 1976 (Nematoda: Rhabditidae) from Iran with its efficacy against Chrysomela populi L. (Col.: Chrysomelidae) and Galleria mellonella L. (Lep.: Pyralidae). J. Helminthol. 2025, 99, e36. [Google Scholar] [CrossRef]
- Bedding, R.A.; Akhurst, R.J. A simple technique for the detection of insect parasitic rhabditid nematodes in soil. Nematologica 1975, 21, 109–110. [Google Scholar] [CrossRef]
- Kaya, H.K.; Stock, S.P. Techniques in Insect Nematology. In Techniques in Insect Pathology; Lacey, L.A., Ed.; Academic Press: London, UK, 1997; pp. 281–324. [Google Scholar]
- Esser, R.P. A water agar en face technique. Proc. Helminthol. Soc. Wash. 1986, 53, 254–255. [Google Scholar]
- Troccoli, A. Aspetti pratici e comparativi di alcune metodologie tradizionali applicate all’identificazione dei nematodi da quarantena. Nematol. Mediterr. 2002, 30, 107–110. [Google Scholar]
- Courtney, W.D.; Polley, D.; Miller, V.L. TAF, an improved fixative in nematode technique. Plant Dis. Report. 1955, 39, 570–571. [Google Scholar]
- Seinhorst, J.W. A rapid method for the transfer of nematodes from fixative to anhydrous glycerin. Nematologica 1959, 4, 67–69. [Google Scholar] [CrossRef]
- Hominick, W.M.; Reid, A.P.; Bohan, D.A.; Briscoe, B.R. Entomopathogenic nematodes: Biodiversity, geographical distribution and the convention on biological diversity. Biocontrol Sci. Technol. 1996, 6, 317–331. [Google Scholar] [CrossRef]
- Archidona-Yuste, A.; Navas-Cortés, J.A.; Cantalapiedra-Navarrete, C.; Palomares-Rius, J.E.; Castillo, P. Cryptic diversity and species delimitation in the Xiphinema americanum-group complex (Nematoda: Longidoridae) as inferred from morphometrics and molecular markers. Zool. J. Linn. Soc. 2016, 176, 231–265. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; p. 24. [Google Scholar]
- Addinsoft (2007) XLSTAT 2007. Data Analysis and Statistics Software for Microsoft Excel; Addinsoft: Paris, France, 2015. [Google Scholar]
- Ali, S.S.; Ahmad, R.; Hussain, M.A.; Pervez, R. Pest Management Through Entomopathogenic Nematodes; Indian Institute of Pulses Research: Kanpur, India, 2005. [Google Scholar]
- Abbot, W.S. Method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–276. [Google Scholar] [CrossRef]
- De Luca, F.; Fanelli, E.; Di Vito, M.; Reyes, A.; De Giorgi, C. Comparisons of the sequences of the D3 expansion of the 26S ribosomal genes reveals different degrees of heterogeneity in different populations and species of Pratylenchus from the Mediterranean region. Eur. J. Plant Pathol. 2004, 110, 949–957. [Google Scholar] [CrossRef]
- Joyce, S.A.; Reid, A.; Driver, F.; Curran, J. Application of polymerase chain reaction (PCR) methods to identification of entomopathogenic nematodes. In COST 812 Biotechnology: Genetics of Entomopathogenic Nematode-Bacterium Complexes: Proceedings of Symposium and Workshop, Phoenix, AZ, USA, 6–8 March 1994; Burnell, A.M., Ehlers, R.U., Masson, J.P., Eds.; St. Patrick’s College: Co. Kildare, Ireland; European Commission, DG XII: Kirchberg, Luxembourg, 1994; pp. 178–187. [Google Scholar]
- Nunn, G.B. Nematode Molecular Evolution: An Investigation of Evolutionary Patterns Among Nematodes Based upon DNA Sequences. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 1992. [Google Scholar]
- Kanzaki, N.; Futai, K. A PCR primer set for determination of phylogenetic relationships of Bursaphelenchus species within the xylophilus-group. Nematology 2002, 4, 35–41. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Darriba, D.; Taboada, G.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.; Huelsenbeck, J. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Larget, B.; Simon, D.L. Markov chains Monte Carlo algorithms for the Bayesian analysis of phylogenetic trees. Mol. Biol. Evol. 1999, 16, 750. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1.4:Tree Figure Drawing Tool 2009; University of Edinburgh: Edinburgh, UK, 2009. [Google Scholar]
- Mahboob, M.; Tahseen, Q. Oscheius shamimi Tahseen and Nisa 2006 revisited with supplementary molecular data, SEM observations and the report of its association with a ground beetle, Scarites indus Olivier 1795. Arch. Phytopathol. Plant Prot. 2021, 55, 355–372. [Google Scholar] [CrossRef]
- Stock, S.P.; Caicedo, A.M.; Calatayud, P.A. Rhabditis (Oscheius) colombiana n. sp. (Nematoda: Rhabditidae), a necromenic associate of the subterranean burrower bug Cyrtomenus bergi (Hemiptera: Cydnidae) from the Cauca Valley, Colombia. Nematology 2005, 7, 363–373. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, J.; Xu, M.; Sun, J.; Yang, S.; An, X.; Gao, G.; Lin, M.; Lai, R.; He, Z.; et al. Heterorhabditidoides chongmingensis gen. nov., sp. nov. (Rhabditida: Rhabditidae) a novel member of the entomopathogenic nematodes. J. Invert. Pathol. 2008, 98, 153–168. [Google Scholar] [CrossRef]
- Bhat, A.H.; Gautum, S.; Rana, A.; Chaubey, A.K.; Abolafia, J.; Půža, V. Morphological, Morphometrical and molecular characterization of Oscheius siddiqii Tabassum and Shahina, 2010 (Rhabditida, Rhabditidae) from India with its taxonomic consequences for the subgenus Oscheius Andrássy, 1976. Biology 2021, 10, 1239. [Google Scholar] [CrossRef]
- Rana, A.; Bhat, A.H.; Chaubey, A.K.; Půža, V.; Abolafia, J. Redescription and synonymization of Oscheius citri Tabassum, Ahahina, Nasira and Erum, 2016 (Rhabditida, Rhabditidae) from India and its taxonomical consequences. J. Helminthol. 2021, 95, 1–14. [Google Scholar] [CrossRef]
- Valadas, V.; Laranjo, M.; Mota, M.; Oliveira, S. A survey of entomopathogenic nematode species in continental Portugal. J. Helminthol. 2014, 88, 327–341. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ratios and Character | Oscheius cyrus | O. cyrus | O. shamimi | O. carolinensis | O. colombiana | O. chongmingensis | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| This Work | [24] | [48] | [4] | [49] | [5] | ||||||
| Georgia | Iran | India | USA | Colombia | China | ||||||
| MALE | FEMALE | FEMALE | MALE | FEMALE | MALE | FEMALE | MALE | FEMALE | MALE | FEMALE | |
| n | 20 | 20 | 10 | 10 | 10 | 20 | 20 | 20 | 20 | 20 | 20 |
| L | 1149 ± 91 (1019–1341) | 1816 ± 184 (1591–2329) | 1193 ± 64 (1097–1293) | 994.3 ± 56 (913–1083) | 1092 ± 104 (951–1255) | 1499 ± 283 (1000–2000) | 1728 ± 265 (1360–2420) | 915 ± 186 (665–1163) | 1288 ± 309 (923–1805) | 1015 ± 143 (848–1414) | 1684 ± 276 (1313–2182) |
| a | 17.2 ± 2.3 (13.1–22.6) | 15.5 ± 1 (12.9–18.5) | 15.3 ± 3.2 (10.5–19.6) | 20.3 ± 1.1 (19–22) | 19.7 ± 2.0 (15.4–24.4) | 20.4 ± 2.5 (15.8–24.3) | 18.2 ± 2.4 (14.9–23.2) | 18 ± 2.2 (16–29) | 17 ± 1.9 (15–19) | 20 ± 1.8 (17–25) | 18 ± 1.7 (14–21) |
| b | 6.2 ± 0.4 (5.4–7.0) | 8.5 ± 0.9 (7.0–10.2) | 6.8 ± 1.0 (5.8–8.9) | 5.91 ± 0.3 (5.4–6.3) | 5.7 ± 0.4 (5.1–6.5) | 7.0 ± 1.2 (4.9–8.9) | 7.0 ± 1.0 (5.4–9.5) | 4.9 ± 0.9 (3.9–5.4) | 6.5 ± 1.4 (5.2–8.0) | 5 ± 0.5 (4.3–6.6) | 6.9 ± 0.9 (5.6–9.0) |
| c | 19.9 ± 2 (17.0–22.9) | 16.0 ± 2 (13.0–20.5) | 13.3 ± 1.0 (12.0–14.5) | 34.7 ± 3.2 (30.9–39.6) | 8.8 ± 0.6 (7.9–9.8) | 32.3 ± 8.4 (18.9–50) | 11.0 ± 2.6 (8.4–17.8) | 14.5 ± 1.5 (13–16) | 9.2 ± 1.1 (8.3–10.0) | 23 ± 2.7 (18–29) | 11 ± 1.8 (8–13) |
| c’ | 1.5 ± 0.4 (1.2–1.7) | 4.7 ± 0.3 (4–5.8) | 4.7 ± 0.9 (3.8–5.9) | 1.1 ± 0.2 (0.9–1.5) | 5.7 ± 0.76 (4.4–6.9) | 1.4 ± 0.3 (1.0–2.3) | 4.4 ± 0.9 (3.2–6.1) | 2.4 ± 0.1 (2.1–3.5) | 4.5 ± 0.1 (3.5–5) | 1.5 ± 0.1 (1.2–1.8) | 4.2 ± 0.5 (3.1–4.8) |
| V% | - | 49.0 ±1 (47.1–51.0) | 49.8 ± 1.0 (49–50) | - | 48.2 ± 1.3 (46.9–51.2) | - | 50.3 ± 1.8 (47.6–55.6) | - | 51 ± 3 (47–57) | - | 49 ± 2.2 (44–52) |
| Maximum body diameter | 67 ± 8 (52–78) | 136 ± 9 (106–143) | 80.4 ± 20.1 (42–115) | 48.1 ± 3.5 (42–54) | 57.4 ± 8.7 (47–80) | 73 ± 9.8 (52–89) | 95.7 ± 13.6 (67–123) | 49 ± 16 (23–72) | 81.5 ± 19.5 (49–106) | 50 ± 7.9 (43–74) | 94 ± 16 (74–141) |
| Tail lenght | 57 ± 5 (49–67) | 132 ± 14 (96–153) | 77.6 ± 19.0 (45–100) | 27.7 ± 3.9 (23–35) | 121.5 ± 6.6 (110–130) | 48 ± 8.6 (32–64) | 160 ± 26.6 (108–206) | 63.5 ± 7 (51–70.5) | 140 ± 18 (110–167) | 45 ± 3.7 (38–56) | 157 ± 20 (121–188) |
| Anal Body width | 30 ± 5 (23–39) | 44 ± 6 (41–54) | 19.4 ± 3.1 (15–23) | 24.8 ± 0.9 (24–27) | 21 ± 3.8 (17–28) | 34 ± 4.6 (25–40) | 36 ± 3.6 (28–43) | 26 ± 5 (15–32) | 31 ± 5 (22–38) | 31 ± 2.6 (25–38) | 37 ± 2.9 (31–43) |
| Spicule length | 48 ± 4 (39–54) | - | - | 46.1 ± 2.1 (43–50) | - | 65 ± 8.9 (50–81) | - | 56 ± 9 (42–68) | - | 47 ± 5.1 (40–59) | - |
| Gubernaculum length | 22 ± 3 (18–28) | - | - | 16.6 ± 1.2 (15–18) | – | 29.4 ± 4.8 (20–35) | - | 20 ± 2 (16–24) | - | 25 ± 4.2 (19–32) | - |
| Lateral field (incisures) | 8 | 6 | 6 | 4 | 4 | 6 | |||||
| Bursa | Leptoderan | - | - | Leptoderan | - | Leptoderan | - | Leptoderan | - | Pseudo-peloderan | - |
| Papillae | 1 ± 1 ± 1/3 ± 3 ± ph | - | - | 1 ± 1 ± 1/3 ± 3 ± ph | - | 1 ± 1 ± 1/3 ± 3 ± ph | - | 1 ± 1 ± 1/3 ± 3 ± ph | - | 1 ± 2 ± 3/3 | |
| host (plant) | Corylus avellana | Forest | Vermicompost | Soil | |||||||
| host (insect) | Scarites indus (Coleoptera: Carabidae) Olivier 1795. | Cyrtomenus bergi (Hemiptera: Cydnidae) Fröschner | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorgadze, O.; Fanelli, E.; Vovlas, A.; Troccoli, A.; Tarasco, E.; De Luca, F. Molecular and Morphological Characterization of the Entomopathogenic Nematode Oscheius cyrus (Nematoda: Rhabditidae) and Molecular Variability of Heterorhabditis bacteriophora from Georgia (Caucasus). Biology 2025, 14, 512. https://doi.org/10.3390/biology14050512
Gorgadze O, Fanelli E, Vovlas A, Troccoli A, Tarasco E, De Luca F. Molecular and Morphological Characterization of the Entomopathogenic Nematode Oscheius cyrus (Nematoda: Rhabditidae) and Molecular Variability of Heterorhabditis bacteriophora from Georgia (Caucasus). Biology. 2025; 14(5):512. https://doi.org/10.3390/biology14050512
Chicago/Turabian StyleGorgadze, Oleg, Elena Fanelli, Alessio Vovlas, Alberto Troccoli, Eustachio Tarasco, and Francesca De Luca. 2025. "Molecular and Morphological Characterization of the Entomopathogenic Nematode Oscheius cyrus (Nematoda: Rhabditidae) and Molecular Variability of Heterorhabditis bacteriophora from Georgia (Caucasus)" Biology 14, no. 5: 512. https://doi.org/10.3390/biology14050512
APA StyleGorgadze, O., Fanelli, E., Vovlas, A., Troccoli, A., Tarasco, E., & De Luca, F. (2025). Molecular and Morphological Characterization of the Entomopathogenic Nematode Oscheius cyrus (Nematoda: Rhabditidae) and Molecular Variability of Heterorhabditis bacteriophora from Georgia (Caucasus). Biology, 14(5), 512. https://doi.org/10.3390/biology14050512