
Temporal and Spatial Population Genetic Variation in Chilean Jack Mackerel (Trachurus murphyi)


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection Areas
2.2. Laboratory Procedures
2.3. Analysis Procedures
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SPO | South Pacific Ocean |
| PCR | Polymerase Chain Reaction |
| DNA | Deoxyribonucleic Acid |
| AMOVA | Analysis of Molecular Variance |
References
- Serra, R. Important Life History Aspects of the Chilean Jack Mackerel, Trachurus symmetricus murphyi. Investig. Pesq. 1991, 36, 67–83. [Google Scholar]
- Bailey, K. Description and Surface Distribution of Juvenile Peruvian Jack Mackerel, Trachurus murphyi, Nichols from the Subtropical Convergence Zone of Central South Pacific. Fish. Bull. 1989, 87, 273–278. [Google Scholar]
- Evseenko, S. On the Reproduction of the Peruvian Jack Mackerel, Trachurus symmetricus murphyi (Nichols), in the Southern Part of the Pacific Ocean. Vopr. Ikhtiologii 1987, 27, 264–273. [Google Scholar]
- Canales-Aguirre, C.B.; Ferrada, S.; Galleguillos, R. Isolation and Characterization of Microsatellite Loci for the Jack Mackerel (Trachurus murphyi Nichols, 1920). Conserv. Genet. 2010, 11, 1235–1237. [Google Scholar] [CrossRef]
- Cárdenas, L.; Silva, A.X.; Magoulas, A.; Cabezas, J.; Poulin, E.; Ojeda, F.P. Genetic Population Structure in the Chilean Jack Mackerel, Trachurus murphyi (Nichols) across the South-Eastern Pacific Ocean. Fish. Res. 2009, 100, 109–115. [Google Scholar] [CrossRef]
- Cubillos, L.A.; Paramo, J.; Ruiz, P.; Núñez, S.; Sepúlveda, A. The Spatial Structure of the Oceanic Spawning of Jack Mackerel (Trachurus murphyi) off Central Chile (1998–2001). Fish. Res. 2008, 90, 261–270. [Google Scholar] [CrossRef]
- Ferrada Fuentes, S.; Galleguillos, R.; Herrera-Yáñez, V.; Canales-Aguirre, C.B. Population Genetics of Chilean Jack Mackerel, Trachurus murphyi Nichols, 1920, (Pisces, Carangidae), in Waters of the South Pacific Ocean. Fishes 2023, 8, 162. [Google Scholar] [CrossRef]
- George-Nascimento, M. Geographical Variations in the Jack Mackerel Trachurus symmetricus murphyi Populations in the Southeastern Pacific Ocean as Evidenced from the Associated Parasite Communities. J. Parasitol. 2000, 86, 929–932. [Google Scholar] [CrossRef]
- Hernández, A.; Sepúlveda, A.; Miranda, L. Morfometría y Merística Del Jurel (Trachurus symmetricus murphyi) a Lo Largo de Chile. In Biologίa y Ecologίa del Jurel en Aguas Chilenas, 1st ed.; Arcos, D., Ed.; Instituto de Investigación Pesquera: Talcahuano, Chile, 1998; pp. 47–63. [Google Scholar]
- Horn, P.L.; Ó Maolagáin, C. The Growth and Age Structure of Chilean Jack Mackerel (Trachurus murphyi) Following Its Influx to New Zealand Waters. J. Fish Biol. 2021, 98, 1144–1154. [Google Scholar] [CrossRef]
- Oliva, M. Metazoan Parasites of the Jack Mackerel Trachurus murphyi (Teleostei, Carangidae) in a Latitudinal Gradient from South America (Chile and Peru). Parasite 1999, 6, 223–230. [Google Scholar] [CrossRef]
- Taylor, P. Stock Structure and Population Biology of the Peruvian Jack Mackerel, Trachurus symmetricus murphyi; National Institute of Water and Atmospheric Research: Auckland, New Zealand, 2002; p. 21. ISSN 1175-1584. [Google Scholar]
- Vásquez, S.; Correa-Ramírez, M.; Parada, C.; Sepúlveda, A. The Influence of Oceanographic Processes on Jack Mackerel (Trachurus murphyi) Larval Distribution and Population Structure in the Southeastern Pacific Ocean. ICES J. Mar. Sci. 2013, 70, 1097–1107. [Google Scholar] [CrossRef]
- Glover, K.A.; Skaala, Ø.; Limborg, M.; Kvamme, C.; Torstensen, E. Microsatellite DNA Reveals Population Genetic Differentiation among Sprat (Sprattus sprattus) Sampled throughout the Northeast Atlantic, Including Norwegian Fjords. ICES J. Mar. Sci. 2011, 68, 2145–2151. [Google Scholar] [CrossRef]
- Waples, R. Separating the Wheat from the Chaff: Patterns of Genetic Differentiation in High Gene Flow Species. J. Hered. 1998, 89, 438–450. [Google Scholar] [CrossRef]
- Jowett, T. Preparation of Nucleic Acids. In Drosophila: A Practical Approach; IRL Press: Oxford, UK, 1986; pp. 275–286. [Google Scholar]
- Kasapidis, P.; Magoulas, A. Development and Application of Microsatellite Markers to Address the Population Structure of the Horse Mackerel Trachurus trachurus. Fish. Res. 2008, 89, 132–135. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research—An Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef]
- Pembleton, L.W.; Cogan, N.O.I.; Forster, J.W. StAMPP: An R Package for Calculation of Genetic Differentiation and Structure of Mixed-Ploidy Level Populations. Mol. Ecol. Resour. 2013, 13, 946–952. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Cham, Switzerland, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Rice, W.R. Analyzing Tables of Statistical Test. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Ritland, K. Estimators for Pairwise Relatedness and Individual Inbreeding Coefficients. Genet. Res. 1996, 67, 175–185. [Google Scholar] [CrossRef]
- Ward, R.D.; Woodwark, M.; Skibinski, D.O.F. A Comparison of Genetic Diversity Levels in Marine, Freshwater, and Anadromous Fishes. J. Fish Biol. 1994, 44, 213–232. [Google Scholar] [CrossRef]
- Sepúlveda, A.; Cubillos, L.; Grechina, A.; Peña, H.; Vilugron, L.; Hernández, A.; Miranda, L.; Sobarzo, P.; Serra, R.; Braun, M.; et al. Migración de Jurel Desde y Hacia La ZEE de Chile Central; Informes Técnicos; FIP/IT No96-15; FIP: Valparaiso, Chile, 1996; p. 273. [Google Scholar]
- Canales-Aguirre, C.B.; Ferrada-Fuentes, S.; Galleguillos, R.; Hernández, C.E. Genetic Structure in a Small Pelagic Fish Coincides with a Marine Protected Area: Seascape Genetics in Patagonian Fjords. PLoS ONE 2016, 11, e0160670. [Google Scholar] [CrossRef] [PubMed]
- Canales-Aguirre, C.B.; Ferrada, S.; Hernandez, C.E.; Galleguillos, R. Population Structure and Demographic History of Genypterus blacodes Using Microsatellite Loci. Fish. Res. 2010, 106, 102–106. [Google Scholar] [CrossRef]
- Canales-Aguirre, C.B.; Ferrada-Fuentes, S.; Galleguillos, R.; Oyarzun, F.X.; Hernández, C.E. Population Genetic Structure of Patagonian Toothfish (Dissostichus eleginoides) in the Southeast Pacific and Southwest Atlantic Ocean. PeerJ 2018, 6, e4173. [Google Scholar] [CrossRef]
- Canales-Aguirre, C.B.; Ferrada-Fuentes, S.; Galleguillos, R.; Oyarzun, F.X.; Buratti, C.C.; Hernández, C.E. High Genetic Diversity and Low-Population Differentiation in the Patagonian Sprat (Sprattus fuegensis) Based on Mitochondrial DNA. Mitochondrial DNA Part DNA Mapp. Seq. Anal. 2018, 29, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Wirgin, I.I.; Waldman, J.R. Use of Nuclear DNA in Stock Identification: Single-Copy and Repetitive Sequence Markers; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Gorbunova, N.; Evseenko, S.; Garetovsky, S. Distribution of Ichthyoplankton in the Frontal Zones of the Peruvian Waters. J. Ichthyol. 1985, 25, 67–79. [Google Scholar]
- Feng, Z.; Yu, W.; Zhang, Y.; Li, Y.; Chen, X. Habitat Variations of Two Commercially Valuable Species Along the Chilean Waters Under Different-Intensity El Niño Events. Front. Mar. Sci. 2022, 9, 919620. [Google Scholar] [CrossRef]
- Hirao, A.S.; Imoto, J.; Fujiwara, A.; Watanabe, C.; Yoda, M.; Matsuura, A.; Akita, T. Genome-Wide SNP Analysis Coupled with Geographic and Reproductive-Phenological Information Reveals Panmixia in a Classical Marine Species, the Japanese Jack Mackerel (Trachurus japonicus). Fish. Res. 2024, 279, 107146. [Google Scholar] [CrossRef]
- Mota, K.G.; Perazza, C.A.; Velho, F.V.; Sardinha, M.d.L.; Morais, L.; Vaini, J.O.; Hallerman, E.; Hilsdorf, A.W.S. Population Genetic Diversity of Cunene Horse Mackerel Trachurus trecae on the Angolan Coast: Implications for Management and Conservation. Fish. Res. 2024, 277, 107069. [Google Scholar] [CrossRef]
- DeWoody, J.A.; Avise, J.C. Microsatellite Variation in Marine, Freshwater and Anadromous Fishes Compared with Other Animals. J. Fish Biol. 2000, 56, 461–473. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Phelps, S.R. Use of Allelic Frequencies to Describe Population Structure. Can. J. Fish. Aquat. Sci. 1981, 38, 1507–1514. [Google Scholar] [CrossRef]
- Meirmans, P.G.; Hedrick, P.W. Assessing Population Structure: FST and Related Measures. Mol. Ecol. Resour. 2011, 11, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, G.; Jueterbock, A.; Kraemer, P.; Deppermann, J.; Harmand, P. Calculations of Population Differentiation Based on GST and D: Forget GST but Not All of Statistics! Mol. Ecol. 2010, 19, 3845–3852. [Google Scholar] [CrossRef] [PubMed]
- Whitlock, M.C. G’ST and D Do Not Replace FST. Mol. Ecol. 2011, 20, 1083–1091. [Google Scholar] [CrossRef]
- Campbell, E.O.; Brunet, B.M.T.; Dupuis, J.R.; Sperling, F.A.H. Would an RRS by Any Other Name Sound as RAD? Methods Ecol. Evol. 2018, 9, 1920–1927. [Google Scholar] [CrossRef]
- Kilian, A.; Wenzl, P.; Huttner, E.; Carling, J.; Xia, L.; Blois, H.; Caig, V.; Heller-Uszynska, K.; Jaccoud, D.; Hopper, C.; et al. Diversity Arrays Technology: A Generic Genome Profiling Technology on Open Platforms. In Data Production and Analysis in Population Genomics: Methods and Protocols; Pompanon, F., Bonin, A., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; pp. 67–89. ISBN 978-1-61779-870-2. [Google Scholar]
- Miller, M.R.; Dunham, J.P.; Amores, A.; Cresko, W.A.; Johnson, E.A. Rapid and Cost-Effective Polymorphism Identification and Genotyping Using Restriction Site Associated DNA (RAD) Markers. Genome Res. 2007, 17, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Peterson, B.K.; Weber, J.N.; Kay, E.H.; Fisher, H.S.; Hoekstra, H.E. Double Digest RADseq: An Inexpensive Method for de Novo SNP Discovery and Genotyping in Model and Non-Model Species. PLoS ONE 2012, 7, e37135. [Google Scholar] [CrossRef] [PubMed]
- Canales-Aguirre, C.B.; Larson, W.A.; McKinney, G.J.; Claure, C.E.; Rocha, J.D.; Ceballos, S.G.; Cádiz, M.I.; Yáñez, J.M.; Gomez-Uchida, D. Neutral and Adaptive Loci Reveal Fine-Scale Population Structure in Eleginops maclovinus from North Patagonia. Ecol. Evol. 2022, 12, e9343. [Google Scholar] [CrossRef]
- Drinan, D.P.; Gruenthal, K.M.; Canino, M.F.; Lowry, D.; Fisher, M.C.; Hauser, L. Population Assignment and Local Adaptation along an Isolation-by-Distance Gradient in Pacific Cod (Gadus macrocephalus). Evol. Appl. 2018, 11, 1448–1464. [Google Scholar] [CrossRef]
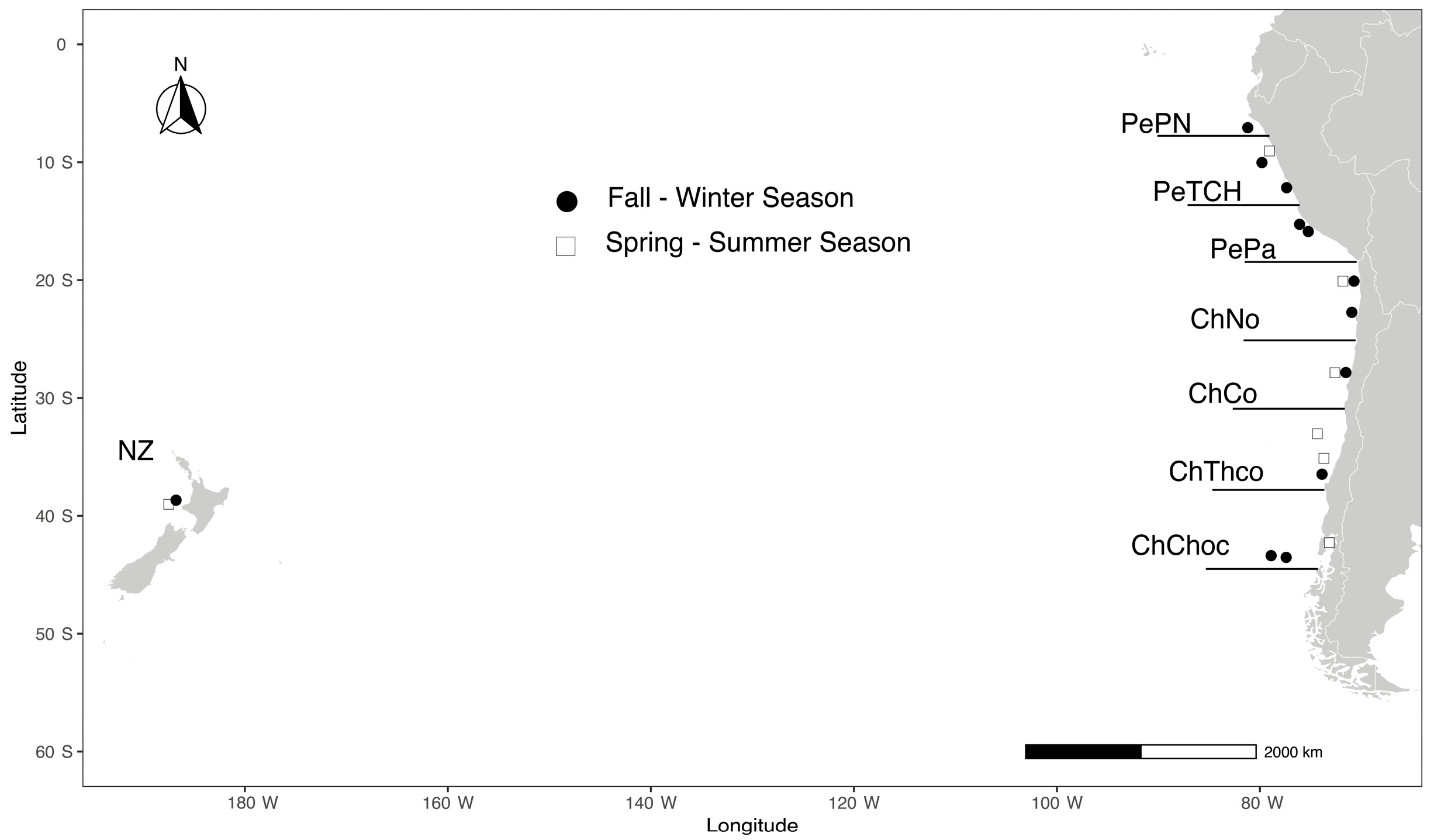
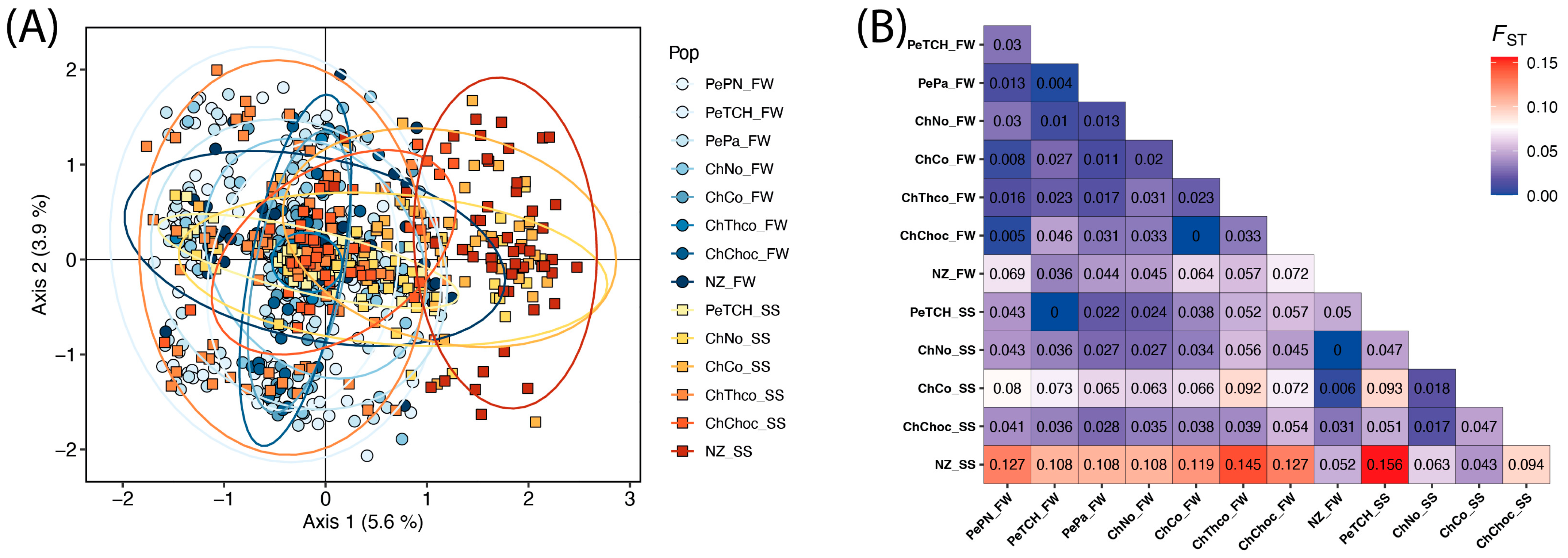

{kind=link}
{kind=link}
| Fall–Winter Season | Spring–Summer Season | |||||||
|---|---|---|---|---|---|---|---|---|
| Area | N | NA | HO | HE | N | NA | HO | HE |
| PePN | 40 | 12.1 | 0.68 | 0.751 | - | - | - | - |
| PeTCH | 101 | 16.6 | 0.697 | 0.824 | 48 | 11.3 | 0.529 | 0.633 |
| PePa | 135 | 20.1 | 0.764 | 0.839 | - | - | - | - |
| ChNo | 96 | 15.7 | 0.73 | 0.825 | 48 | 16.3 | 0.74 | 0.818 |
| ChCo | 38 | 11.6 | 0.615 | 0.775 | 48 | 15.2 | 0.658 | 0.77 |
| ChThco | 40 | 8.2 | 0.572 | 0.701 | 72 | 15.3 | 0.637 | 0.739 |
| ChChoc | 40 | 13.6 | 0.703 | 0.764 | 48 | 16.9 | 0.723 | 0.832 |
| NZ | 48 | 12.4 | 0.612 | 0.728 | 50 | 14.6 | 0.591 | 0.689 |
| Average | 13.8 | 0.672 | 0.776 | 14.9 | 0.646 | 0.747 | ||
| Fall–Winter Season | Spring–Summer Season | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Nº Groups | Aggrupation | FST | p-Value | FCT | p-Value | FST | p-Value | FCT | p-Value |
| Three | Peru v/s Chile v/s NZ | −0.103 | 1 | 0 | 0.689 | −0.007 | 1 | 0.05 | 0.133 |
| Two | Peru + ChNo v/s Chile | −0.14 | 1 | −0.074 | 0.973 | −0.031 | 1 | −0.005 | 0.604 |
| One | Panmixis | −0.103 | 1 | - | - | −0.030 | 1 | - | - |
| Expected Heterozygosity | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Season/Previous Studies | Year | TmurB6 | TmurA104 | TmurA115 | TmurC4 | TmurA101 | TmurB104 | TmurB2 | Tt29 | Tt62 | Tt133 |
| Ferrada Fuentes et al. [7] | 2007 | - | 0.943 | 0.95 | - | 0.927 | - | - | 0.747 | 0.855 | 0.861 |
| Cárdenas et al. [5] | 2009 | - | - | - | - | - | - | - | 0.76 | 0.83 | 0.87 |
| Fall–Winter | 2011 | 0.782 | 0.93 | 0.916 | 0.82 | 0.871 | 0.521 | 0.552 | 0.758 | 0.851 | 0.758 |
| Spring–Summer | 2011–2012 | 0.444 | 0.942 | 0.945 | 0.819 | 0.779 | 0.648 | 0.704 | 0.634 | 0.865 | 0.687 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canales-Aguirre, C.B.; Ferrada Fuentes, S.; Galleguillos, R. Temporal and Spatial Population Genetic Variation in Chilean Jack Mackerel (Trachurus murphyi). Biology 2025, 14, 510. https://doi.org/10.3390/biology14050510
Canales-Aguirre CB, Ferrada Fuentes S, Galleguillos R. Temporal and Spatial Population Genetic Variation in Chilean Jack Mackerel (Trachurus murphyi). Biology. 2025; 14(5):510. https://doi.org/10.3390/biology14050510
Chicago/Turabian StyleCanales-Aguirre, Cristian B., Sandra Ferrada Fuentes, and Ricardo Galleguillos. 2025. "Temporal and Spatial Population Genetic Variation in Chilean Jack Mackerel (Trachurus murphyi)" Biology 14, no. 5: 510. https://doi.org/10.3390/biology14050510
APA StyleCanales-Aguirre, C. B., Ferrada Fuentes, S., & Galleguillos, R. (2025). Temporal and Spatial Population Genetic Variation in Chilean Jack Mackerel (Trachurus murphyi). Biology, 14(5), 510. https://doi.org/10.3390/biology14050510