
New Insights into the Geometry and Topology of DNA Replication Intermediates


 , ,
, ,
Simple Summary
Abstract
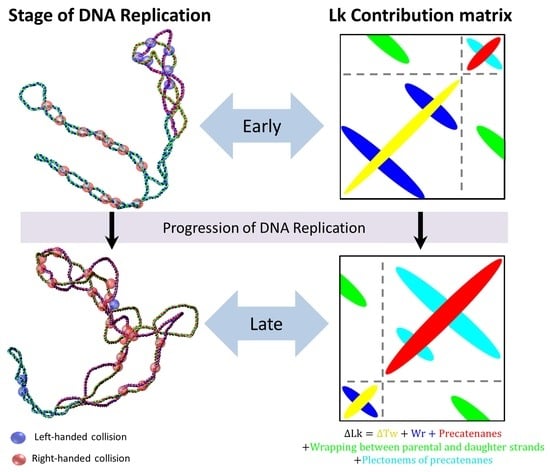
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Modeling of Molecules
2.2. Molecular Dynamics Simulation
2.3. Topological Properties
3. Results
3.1. Distribution of Superhelical Stress After Deproteinization
3.2. Temporal Evolution of the Topological Components of ∆LK in Deproteinized Replication Intermediates
3.3. Dynamics of Collision Events in Deproteinized Replication Intermediates
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CCC | Covalently closed circular |
| RI | Replication intermediate |
| Lk | Linking number |
| ∆Lk | Linking number variation |
| Tw | Twist |
| ∆Tw | Twist variation |
| Wr | Writhe |
| ΔWr | Writhe variation |
References
- Postow, L.; Hardy, C.D.; Arsuaga, J.; Cozzarelli, N.R. Topological domain structure of the Escherichia coli chromosome. Genes Dev. 2004, 18, 1766–1779. [Google Scholar] [CrossRef] [PubMed]
- Schvartzman, J.B.; Hernández, P.; Krimer, D.B.; Dorier, J.; Stasiak, A. Closing the DNA replication cycle: From simple circular molecules to supercoiled and knotted DNA catenanes. Nucleic Acids Res. 2019, 47, 7182–7198. [Google Scholar] [CrossRef]
- Bauer, W.R.; Crick, F.H.; White, J.H. Supercoiled DNA. Sci. Am. 1980, 243, 100–113. [Google Scholar] [PubMed]
- Boles, T.C.; White, J.H.; Cozzarelli, N.R. Structure of plectonemically supercoiled DNA. J. Mol. Biol. 1990, 213, 931–951. [Google Scholar] [CrossRef]
- Keszthelyi, A.; Minchell, N.; Baxter, J. The Causes and Consequences of Topological Stress during DNA Replication. Genes 2016, 7, 134. [Google Scholar] [CrossRef]
- Cebrián, J.; Castán, A.; Martínez, V.; Kadomatsu-Hermosa, M.J.; Parra, C.; Fernández-Nestosa, M.J.; Schaerer, C.; Hernández, P.; Krimer, D.B.; Schvartzman, J.B. Direct Evidence for the Formation of Precatenanes during DNA Replication. J. Biol. Chem. 2015, 290, 13725–13735. [Google Scholar] [CrossRef]
- McKie, S.J.; Desai, P.R.; Seol, Y.; Allen, A.M.; Maxwell, A.; Neuman, K.C. Topoisomerase VI is a chirally-selective, preferential DNA decatenase. eLife 2022, 11, e67021. [Google Scholar] [CrossRef]
- Champoux, J.J.; Been, M.D. Topoisomerases and the Swivel Problem. In Mechanistic Studies of DNA Replication and Genetic Recombination; Elsevier: Amsterdam, The Netherlands, 1980; pp. 809–815. [Google Scholar]
- Champoux, J.J. DNA Topoisomerases: Structure, Function, and Mechanism. Annu. Rev. Biochem. 2001, 70, 369–413. [Google Scholar] [CrossRef] [PubMed]
- Sundin, O.; Varshavsky, A. Terminal stages of SV40 DNA replication proceed via multiply intertwined catenated dimers. Cell 1980, 21, 103–114. [Google Scholar] [CrossRef]
- Sundin, O.; Varshavsky, A. Arrest of segregation leads to accumulation of highly intertwined catenated dimers: Dissection of the final stages of SV40 DNA replication. Cell 1981, 25, 659–669. [Google Scholar] [CrossRef]
- Kouzine, F.; Sanford, S.; Elisha-Feil, Z.; Levens, D. The functional response of upstream DNA to dynamic supercoiling in vivo. Nat. Struct. Mol. Biol. 2008, 15, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Funnell, B.E.; Baker, T.A.; Kornberg, A. In vitro assembly of a prepriming complex at the origin of the Escherichia coli chromosome. J. Biol. Chem. 1987, 262, 10327–10334. [Google Scholar] [CrossRef]
- Marians, K.J.; Minden, J.S.; Parada, C. Replication of superhelical DNAs in vitro. Prog. Nucleic Acid Res. Mol. Biol. 1986, 33, 111–140. [Google Scholar]
- Gellert, M.; Mizuuchi, K.; O’Dea, M.H.; Nash, H.A. DNA gyrase: An enzyme that introduces superhelical turns into DNA. Proc. Natl. Acad. Sci. USA 1976, 73, 3872–3876. [Google Scholar] [CrossRef]
- López, V.; Martínez-Robles, M.L.; Hernández, P.; Krimer, D.B.; Schvartzman, J.B. Topo IV is the topoisomerase that knots and unknots sister duplexes during DNA replication. Nucleic Acids Res. 2012, 40, 3563–3573. [Google Scholar] [CrossRef] [PubMed]
- Drlica, K.; Zhao, X. DNA Gyrase, Topoisomerase IV, and the 4-Quinolones. Microbiol. Mol. Biol. Rev. 1997, 61, 377–392. [Google Scholar] [PubMed]
- Schvartzman, J.B.; Martínez, V.; Hernández, P.; Krimer, D.B.; Fernández-Nestosa, M. Changes in the topology of DNA replication intermediates: Important discrepancies between in vitro and in vivo. BioEssays 2021, 43, 2000309. [Google Scholar] [CrossRef]
- Bates, A.D.; Maxwell, A. DNA Topology; Oxford Academic: Oxford, UK, 2005. [Google Scholar]
- Stone, M.D.; Bryant, Z.; Crisona, N.J.; Smith, S.B.; Vologodskii, A.; Bustamante, C.; Cozzarelli, N.R. Chirality sensing by Escherichia coli topoisomerase IV and the mechanism of type II topoisomerases. Proc. Natl. Acad. Sci. USA 2003, 100, 8654–8659. [Google Scholar] [CrossRef]
- Fogg, J.M.; Judge, A.K.; Stricker, E.; Chan, H.L.; Zechiedrich, L. Supercoiling and looping promote DNA base accessibility and coordination among distant sites. Nat. Commun. 2021, 12, 5683. [Google Scholar] [CrossRef]
- Wang, Q.; Irobalieva, R.N.; Chiu, W.; Schmid, M.F.; Fogg, J.M.; Zechiedrich, L.; Pettitt, B.M. Influence of DNA sequence on the structure of minicircles under torsional stress. Nucleic Acids Res. 2017, 45, 7633–7642. [Google Scholar] [CrossRef]
- Peter, B.J.; Ullsperger, C.; Hiasa, H.; Marians, K.J.; Cozzarelli, N.R. The Structure of Supercoiled Intermediates in DNA Replication. Cell 1998, 94, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Martínez, V.; Schaerer, C.; Hernández, P.; Krimer, D.B.; Schvartzman, J.B.; Fernández-Nestosa, M.J. Distribution of torsional stress between the un-replicated and replicated regions in partially replicated molecules. J. Biomol. Struct. Dyn. 2021, 39, 2266–2277. [Google Scholar] [CrossRef]
- Olavarrieta, L. Supercoiling, knotting and replication fork reversal in partially replicated plasmids. Nucleic Acids Res. 2002, 30, 656–666. [Google Scholar] [CrossRef]
- Bliska, J.B.; Cozzarelli, N.R. Use of site-specific recombination as a probe of DNA structure and metabolism in vivo. J. Mol. Biol. 1987, 194, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zechiedrich, L.; Chan, H.S. Action at Hooked or Twisted–Hooked DNA Juxtapositions Rationalizes Unlinking Preference of Type-2 Topoisomerases. J. Mol. Biol. 2010, 400, 963–982. [Google Scholar] [CrossRef] [PubMed]
- Rawdon, E.J.; Dorier, J.; Racko, D.; Millett, K.C.; Stasiak, A. How topoisomerase IV can efficiently unknot and decatenate negatively supercoiled DNA molecules without causing their torsional relaxation. Nucleic Acids Res. 2016, 44, 4528–4538. [Google Scholar] [CrossRef]
- Huang, J.; Schlick, T.; Vologodskii, A. Dynamics of site juxtaposition in supercoiled DNA. Proc. Natl. Acad. Sci. USA 2001, 98, 968–973. [Google Scholar] [CrossRef]
- Arnoldi, E.; Pan, X.S.; Fisher, L.M. Functional determinants of gate-DNA selection and cleavage by bacterial type II topoisomerases. Nucleic Acids Res. 2013, 41, 9411–9423. [Google Scholar] [CrossRef]
- Morgan, I.L.; McKie, S.J.; Kim, R.; Seol, Y.; Xu, J.; Harami, G.M.; Maxwell, A.; Neuman, K.C. Highly sensitive mapping of in vitro type II topoisomerase DNA cleavage sites with SHAN-seq. Nucleic Acids Res. 2024, 52, 9777–9787. [Google Scholar] [CrossRef]
- Benjamin, H.W.; Cozzarelli, N.R. Genetic Chemistry: The Molecular Basis of Heredity; Robert A. Welch Foundation: Houston, TX, USA, 1986; pp. 107–126. [Google Scholar]
- Corless, S.; Gilbert, N. Effects of DNA supercoiling on chromatin architecture. Biophys. Rev. 2016, 8, 51–64. [Google Scholar] [CrossRef]
- Cebrián, J.; Martínez-Robles, M.L.; Martínez, V.; Hernández, P.; Krimer, D.B.; Schvartzman, J.B.; Fernández-Nestosa, M.-J. Role of Supercoiling and Topoisomerases in DNA Knotting. DNA 2024, 4, 170–179. [Google Scholar] [CrossRef]
- Neuman, K.C.; Charvin, G.; Bensimon, D.; Croquette, V. Mechanisms of chiral discrimination by topoisomerase IV. Proc. Natl. Acad. Sci. USA 2009, 106, 6986–6991. [Google Scholar] [CrossRef]
- Ouldridge, T.E. Coarse-Grained Modelling of DNA and DNA Self-Assembly; Springer: Heidelberg, Germany, 2012. [Google Scholar]
- Ouldridge, T.E.; Louis, A.A.; Doye, J.P.K. Structural, mechanical, and thermodynamic properties of a coarse-grained DNA model. J. Chem. Phys. 2011, 134, 085101. [Google Scholar] [CrossRef] [PubMed]
- Šulc, P.; Romano, F.; Ouldridge, T.E.; Rovigatti, L.; Doye, J.P.K.; Louis, A.A. Sequence-dependent thermodynamics of a coarse-grained DNA model. J. Chem. Phys. 2012, 137, 135101. [Google Scholar] [CrossRef] [PubMed]
- Snodin, B.E.K.; Randisi, F.; Mosayebi, M.; Šulc, P.; Schreck, J.S.; Romano, F.; Ouldridge, T.E.; Tsukanov, R.; Nir, E.; Louis, A.A.; et al. Introducing improved structural properties and salt dependence into a coarse-grained model of DNA. J. Chem. Phys. 2015, 142, 234901. [Google Scholar] [CrossRef]
- Sengar, A.; Ouldridge, T.E.; Henrich, O.; Rovigatti, L.; Šulc, P. A Primer on the oxDNA Model of DNA: When to Use it, How to Simulate it and How to Interpret the Results. Front. Mol. Biosci. 2021, 8, 693710. [Google Scholar] [CrossRef]
- Suma, A.; Poppleton, E.; Matthies, M.; Šulc, P.; Romano, F.; Louis, A.A.; Doye, J.P.K.; Micheletti, C.; Rovigatti, L. TacoxDNA: A user-friendly web server for simulations of complex DNA structures, from single strands to origami. J. Comput. Chem. 2019, 40, 2586–2595. [Google Scholar] [CrossRef] [PubMed]
- Henrich, O.; Gutiérrez Fosado, Y.A.; Curk, T.; Ouldridge, T.E. Coarse-grained simulation of DNA using LAMMPS: An implementation of the oxDNA model and its applications. Eur. Phys. J. E 2018, 41, 57. [Google Scholar] [CrossRef]
- Poppleton, E.; Romero, R.; Mallya, A.; Rovigatti, L.; Šulc, P. OxDNA.org: A public webserver for coarse-grained simulations of DNA and RNA nanostructures. Nucleic Acids Res. 2021, 49, W491–W498. [Google Scholar] [CrossRef]
- Poppleton, E.; Bohlin, J.; Matthies, M.; Sharma, S.; Zhang, F.; Šulc, P. Design, optimization and analysis of large DNA and RNA nanostructures through interactive visualization, editing and molecular simulation. Nucleic Acids Res. 2020, 48, e72. [Google Scholar] [CrossRef]
- Bohlin, J.; Matthies, M.; Poppleton, E.; Procyk, J.; Mallya, A.; Yan, H.; Šulc, P. Design and simulation of DNA, RNA and hybrid protein–nucleic acid nanostructures with oxView. Nat. Protoc. 2022, 17, 1762–1788. [Google Scholar] [CrossRef] [PubMed]
- Crick, F.H. Linking numbers and nucleosomes. Proc. Natl. Acad. Sci. USA 1976, 73, 2639–2643. [Google Scholar] [CrossRef] [PubMed]
- Dennis, M.R.; Hannay, J.H. Geometry of Călugăreanu’s theorem. Proc. R. Soc. A 2005, 461, 3245–3254. [Google Scholar] [CrossRef]
- Scovell, W. Supercoiled DNA. In Concepts in Biochemistry; Bowling Green State University: Bowling Green, OH, USA, 1986; Volume 63, pp. 562–585. [Google Scholar]
- Klenin, K.; Langowski, J. Computation of writhe in modeling of supercoiled DNA. Biopolymers 2000, 54, 307–317. [Google Scholar] [CrossRef]
- Fuller, F.B. Decomposition of the linking number of a closed ribbon: A problem from molecular biology. Proc. Natl. Acad. Sci. USA 1978, 75, 3557–3561. [Google Scholar] [CrossRef]
- Sierzega, Z.; Wereszczynski, J.; Prior, C. WASP: A software package for correctly characterizing the topological development of ribbon structures. Sci. Rep. 2021, 11, 1527. [Google Scholar] [CrossRef]
- Coronel, L.; Suma, A.; Micheletti, C. Dynamics of supercoiled DNA with complex knots: Large-scale rearrangements and persistent multi-strand interlocking. Nucleic Acids Res. 2018, 46, 7533–7541. [Google Scholar] [CrossRef]
- Araki, S.; Nakai, T.; Hizume, K.; Takeyasu, K.; Yoshikawa, K. Hydrodynamic radius of circular DNA is larger than that of linear DNA. Chem. Phys. Lett. 2006, 418, 255–259. [Google Scholar] [CrossRef]
- Huang, J.; Schlick, T. Macroscopic modeling and simulations of supercoiled DNA with bound proteins. J. Chem. Phys. 2002, 117, 8573–8586. [Google Scholar] [CrossRef]
- Valdés, A.; Coronel, L.; Martínez-García, B.; Segura, J.; Dyson, S.; Díaz-Ingelmo, O.; Micheletti, C.; Roca, J. Transcriptional supercoiling boosts topoisomerase II-mediated knotting of intracellular DNA. Nucleic Acids Res. 2019, 47, 6946–6955. [Google Scholar] [CrossRef]
- Robertson, R.M.; Laib, S.; Smith, D.E. Diffusion of isolated DNA molecules: Dependence on length and topology. Proc. Natl. Acad. Sci. USA 2006, 103, 7310–7314. [Google Scholar] [CrossRef] [PubMed]
- Bruand, C.; Ehrlich, S.D.; Jannière, L. Unidirectional theta replication of the structurally stable Enterococcus faecalis plasmid pAM beta 1. EMBO J. 1991, 10, 2171–2177. [Google Scholar] [CrossRef] [PubMed]
- Barańska, S.; Konopa, G.; Wegrzyn, G. Directionality of lambda plasmid DNA replication carried out by the heritable replication complex. Nucleic Acids Res. 2002, 30, 1176–1181. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vologodskii, A. Unlinking of Supercoiled DNA Catenanes by Type IIA Topoisomerases. Biophys. J. 2011, 101, 1403–1411. [Google Scholar] [CrossRef][Green Version]
- Dalvie, E.D.; Stacy, J.C.; Neuman, K.C.; Osheroff, N. Recognition of DNA Supercoil Handedness during Catenation Catalyzed by Type II Topoisomerases. Biochemistry 2022, 61, 2148–2158. [Google Scholar] [CrossRef]
- Schalbetter, S.A.; Mansoubi, S.; Chambers, A.L.; Downs, J.A.; Baxter, J. Fork rotation and DNA precatenation are restricted during DNA replication to prevent chromosomal instability. Proc. Natl. Acad. Sci. USA 2015, 112, E4565–E4570. [Google Scholar] [CrossRef]
- Seol, Y.; Neuman, K.C. The dynamic interplay between DNA topoisomerases and DNA topology. Biophys. Rev. 2016, 8, 101–111. [Google Scholar] [CrossRef]
- Brown, P.O.; Cozzarelli, N.R. A Sign Inversion Mechanism for Enzymatic Supercoiling of DNA. Science 1979, 206, 1081–1083. [Google Scholar] [CrossRef]
- Nöllmann, M.; Stone, M.D.; Bryant, Z.; Gore, J.; Crisona, N.J.; Hong, S.C.; Mitelheiser, S.; Maxwell, A.; Bustamante, C.; Cozzarelli, N.R. Multiple modes of Escherichia coli DNA gyrase activity revealed by force and torque. Nat. Struct. Mol. Biol. 2007, 14, 264–271. [Google Scholar] [CrossRef]
- Gellert, M.; Mizuuchi, K.; O’Dea, M.H.; Itoh, T.; Tomizawa, J.I. Nalidixic acid resistance: A second genetic character involved in DNA gyrase activity. Proc. Natl. Acad. Sci. USA 1977, 74, 4772–4776. [Google Scholar] [CrossRef]
- Higgins, N.P.; Peebles, C.L.; Sugino, A.; Cozzarelli, N.R. Purification of subunits of Escherichia coli DNA gyrase and reconstitution of enzymatic activity. Proc. Natl. Acad. Sci. USA 1978, 75, 1773–1777. [Google Scholar] [CrossRef] [PubMed]
- Kreuzer, K.N.; Cozzarelli, N.R. Formation and resolution of DNA catenanes by DNA gyrase. Cell 1980, 20, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Ashley, R.E.; Dittmore, A.; McPherson, S.A.; Turnbough, C.L.; Neuman, K.C.; Osheroff, N. Activities of gyrase and topoisomerase IV on positively supercoiled DNA. Nucleic Acids Res. 2017, 45, 9611–9624. [Google Scholar] [CrossRef] [PubMed]
- Levine, C.; Hiasa, H.; Marians, K.J. DNA gyrase and topoisomerase IV: Biochemical activities, physiological roles during chromosome replication, and drug sensitivities. Biochim. Et Biophys. Acta (BBA)-Gene Struct. Expr. 1998, 1400, 29–43. [Google Scholar] [CrossRef]
- Cebrián, J.; Martínez, V.; Hernández, P.; Krimer, D.B.; Fernández-Nestosa, M.J.; Schvartzman, J.B. Two-Dimensional Gel Electrophoresis to Study the Activity of Type IIA Topoisomerases on Plasmid Replication Intermediates. Biology 2021, 10, 1195. [Google Scholar] [CrossRef]
- Peng, H.; Marians, K.J. Decatenation activity of topoisomerase IV during oriC and pBR322 DNA replication in vitro. Proc. Natl. Acad. Sci. USA 1993, 90, 8571–8575. [Google Scholar] [CrossRef]
- Zechiedrich, E.L.; Cozzarelli, N.R. Roles of topoisomerase IV and DNA gyrase in DNA unlinking during replication in Escherichia coli. Genes Dev. 1995, 9, 2859–2869. [Google Scholar] [CrossRef]
- Crisona, N.J.; Strick, T.R.; Bensimon, D.; Croquette, V.; Cozzarelli, N.R. Preferential relaxation of positively supercoiled DNA by E. coli topoisomerase IV in single-molecule and ensemble measurements. Genes Dev. 2000, 14, 2881–2892. [Google Scholar] [CrossRef]
- Helgesen, E.; Sætre, F.; Skarstad, K. Topoisomerase IV tracks behind the replication fork and the SeqA complex during DNA replication in Escherichia coli. Sci. Rep. 2021, 11, 474. [Google Scholar] [CrossRef]
- Charvin, G.; Bensimon, D.; Croquette, V. Single-molecule study of DNA unlinking by eukaryotic and prokaryotic type-II topoisomerases. Proc. Natl. Acad. Sci. USA 2003, 100, 9820–9825. [Google Scholar] [CrossRef]
- Marko, J.F. Supercoiled and braided DNA under tension. Phys. Rev. E 1997, 55, 1758–1772. [Google Scholar] [CrossRef]
- Junier, I.; Ghobadpour, E.; Espeli, O.; Everaers, R. DNA supercoiling in bacteria: State of play and challenges from a viewpoint of physics based modeling. Front. Microbiol. 2023, 14, 1192831. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez, V.; Ruiz-Díaz, E.; Cardozo, D.; Cappo, C.; Schaerer, C.E.; Cebrián, J.; Krimer, D.B.; Fernández-Nestosa, M.J. New Insights into the Geometry and Topology of DNA Replication Intermediates. Biology 2025, 14, 478. https://doi.org/10.3390/biology14050478
Martínez V, Ruiz-Díaz E, Cardozo D, Cappo C, Schaerer CE, Cebrián J, Krimer DB, Fernández-Nestosa MJ. New Insights into the Geometry and Topology of DNA Replication Intermediates. Biology. 2025; 14(5):478. https://doi.org/10.3390/biology14050478
Chicago/Turabian StyleMartínez, Victor, Edith Ruiz-Díaz, Delia Cardozo, Cristian Cappo, Christian E. Schaerer, Jorge Cebrián, Dora B. Krimer, and María José Fernández-Nestosa. 2025. "New Insights into the Geometry and Topology of DNA Replication Intermediates" Biology 14, no. 5: 478. https://doi.org/10.3390/biology14050478
APA StyleMartínez, V., Ruiz-Díaz, E., Cardozo, D., Cappo, C., Schaerer, C. E., Cebrián, J., Krimer, D. B., & Fernández-Nestosa, M. J. (2025). New Insights into the Geometry and Topology of DNA Replication Intermediates. Biology, 14(5), 478. https://doi.org/10.3390/biology14050478