

The Mutations in RcMYB114 Affect Anthocyanin Glycoside Accumulation in Rose
 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Cloning of the RcMYB114 Gene and Producing the Mutations by Site-Directed Mutagenesis
2.3. Gene Expression Analysis
2.4. Yeast Vector Construction and Yeast Two-Hybrid Assay
2.5. Construction of Transient Overexpression Vector and Tobacco Transformation
2.6. Determination of Anthocyanin Glycoside Content
3. Results
3.1. Anthocyanin Content of ‘Slater’s Crimson China’ Petals at Different Developmental Stages
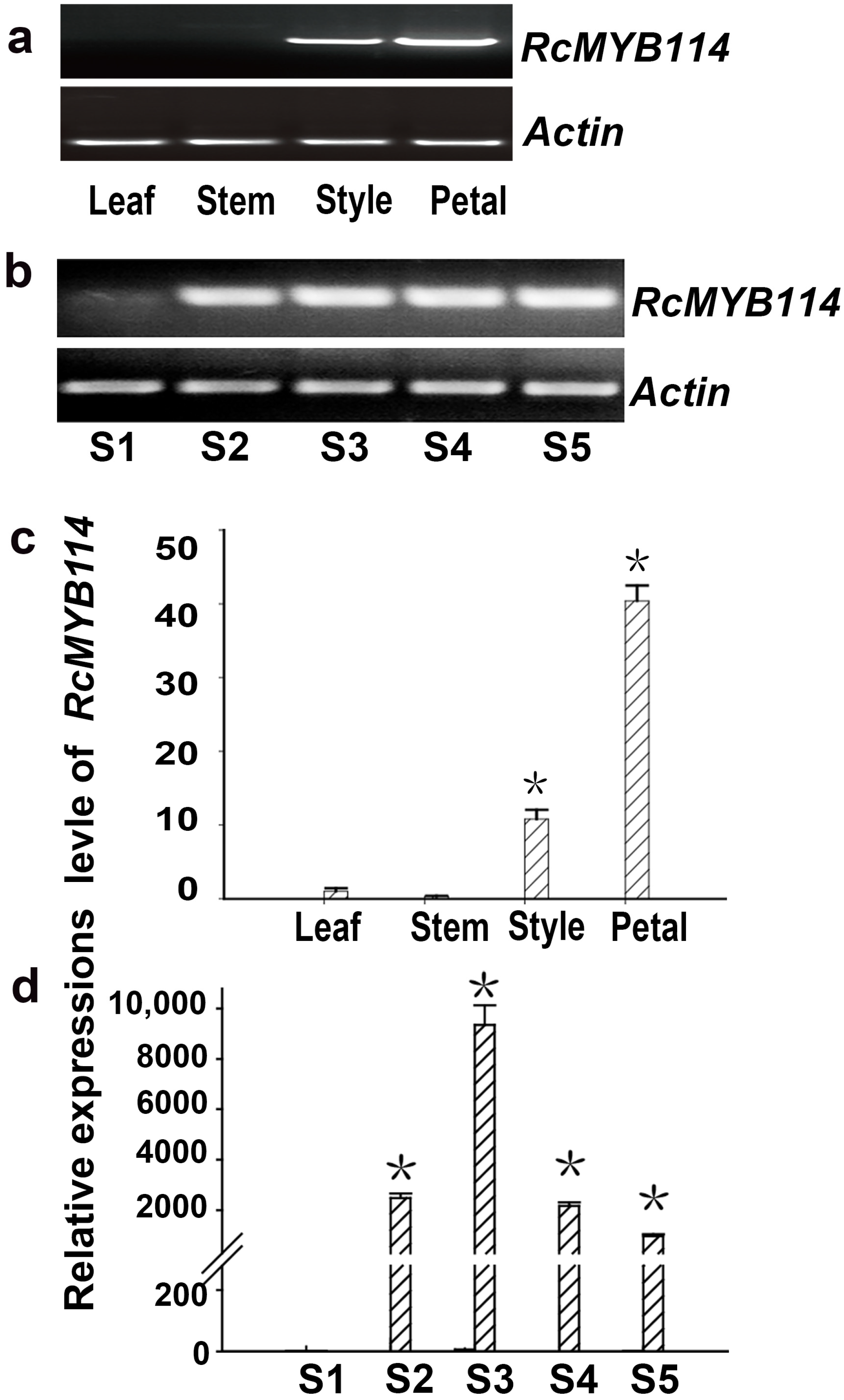
3.2. Expression Analysis of RcMYB114 in ‘Slater’s Crimson China’
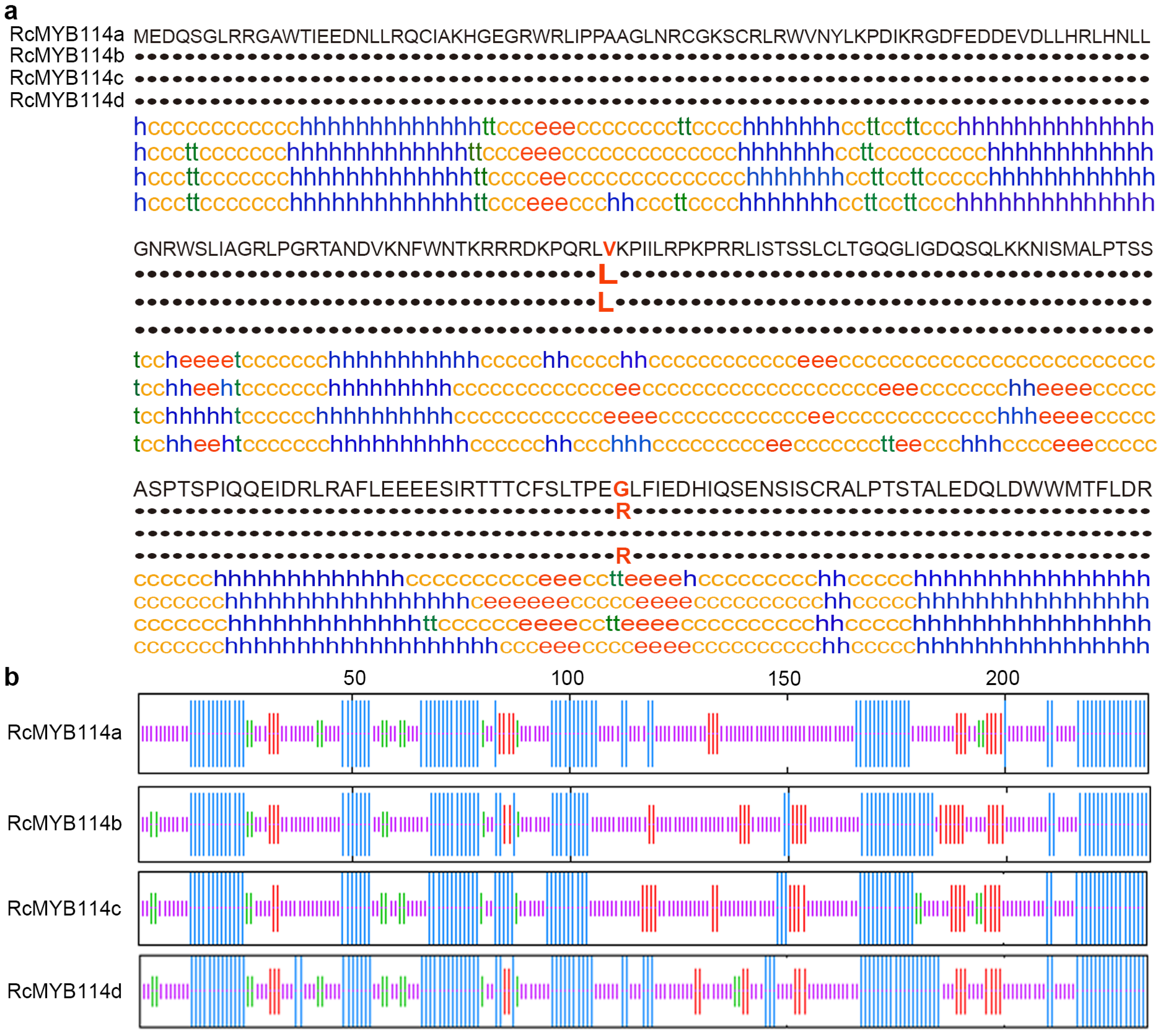
3.3. Cloning and Sequence Analysis of RcMYB114 in ‘Slater’s Crimson China’
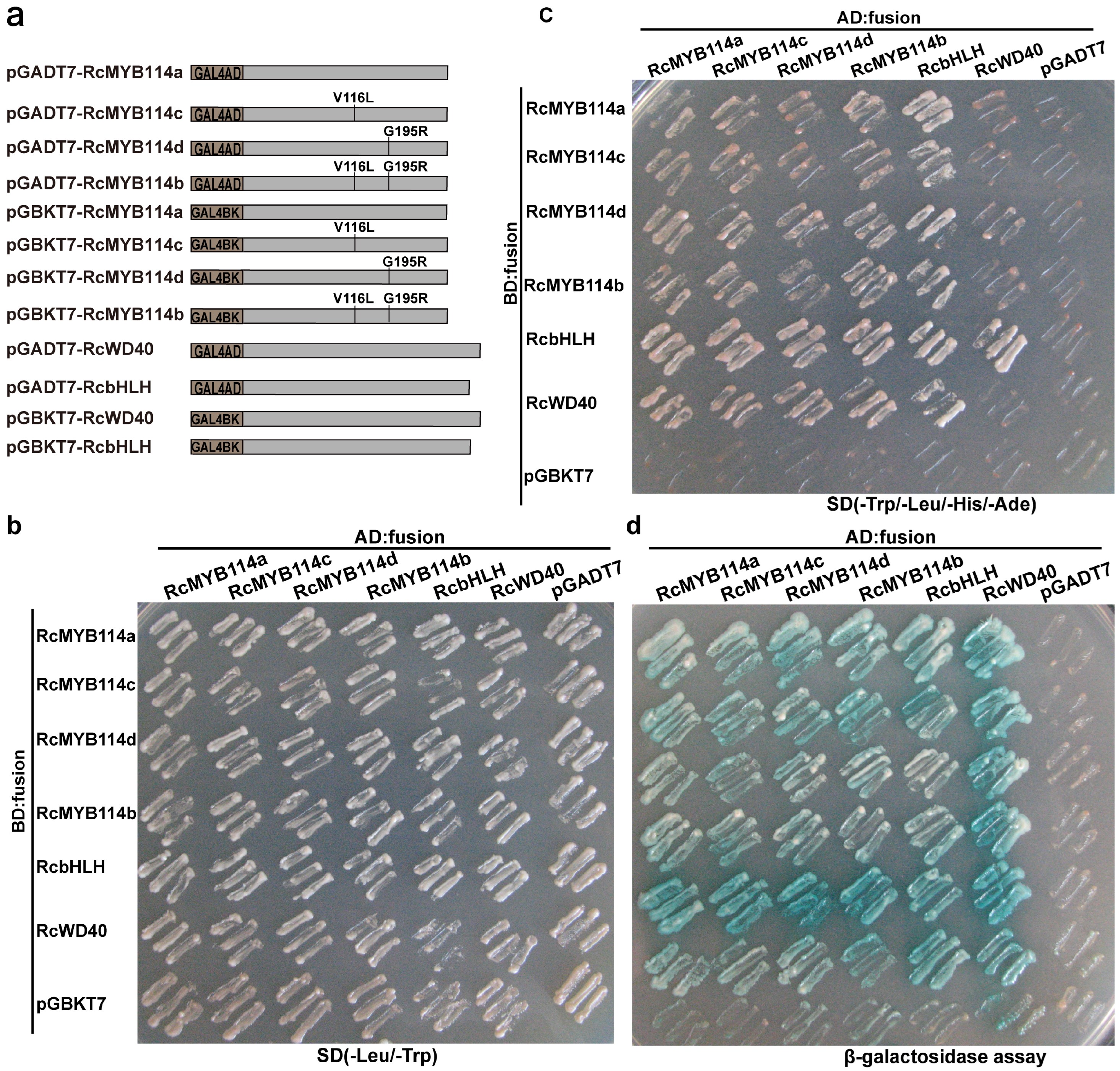
3.4. RcMYB114a and Its Mutations Can Interact with RcbHLH and RcWD40
3.5. Effect of RcMYB114 and Its Mutation on Anthocyanin Accumulation
4. Discussion
4.1. The RcMYB114 Gene Is a Key Regulator of Anthocyanin Accumulation in Rose
4.2. Differential Activity of Genes with Missense Mutations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin, C.; Gerats, T. Control of pigment biosynthesis genes during petal development. Plant Cell 1993, 5, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Stracke, R.; Holtgrawe, D.; Schneider, J.; Pucker, B.; Sorensen, T.R.; Weisshaar, B. Genome-wide identification and characterisation of R2R3-MYB genes in sugar beet (Beta vulgaris). BMC Plant Biol. 2014, 14, 249. [Google Scholar] [CrossRef] [PubMed]
- Matus, J.T.; Aquea, F.; Arce-Johnson, P. Analysis of the grape MYB R2R3 subfamily reveals expanded wine quality-related clades and conserved gene structure organization across Vitis and Arabidopsis genomes. BMC Plant Biol. 2008, 8, 83. [Google Scholar] [CrossRef] [PubMed]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr. Opin. Plant Biol. 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Pucker, B.; Pandey, A.; Weisshaar, B.; Stracke, R. The R2R3-MYB gene family in banana (Musa acuminata): Genome-wide identification, classification and expression patterns. PLoS ONE 2020, 15, e0239275. [Google Scholar] [CrossRef]
- Han, Y.; Yu, J.; Zhao, T.; Cheng, T.; Wang, J.; Yang, W.; Pan, H.; Zhang, Q. Dissecting the genome-wide evolution and function of R2R3-MYB transcription factor family in Rosa chinensis. Genes 2019, 10, 823. [Google Scholar] [CrossRef]
- Schwinn, K.; Venail, J.; Shang, Y.; Mackay, S.; Alm, V.; Butelli, E.; Oyama, R.; Bailey, P.; Davies, K.; Martin, C. A small family of MYB-regulatory genes controls floral pigmentation intensity and patterning in the genus Antirrhinum. Plant Cell 2006, 18, 831–851. [Google Scholar] [CrossRef]
- Gu, Z.; Zhu, J.; Hao, Q.; Yuan, Y.W.; Duan, Y.W.; Men, S.; Wang, Q.; Hou, Q.; Liu, Z.A.; Shu, Q.; et al. A novel R2R3-MYB transcription factor contributes to petal bblotch formation by regulating organ-specific expression of PsCHS in tree peony (Paeonia suffruticosa). Plant Cell Physiol. 2019, 60, 599–611. [Google Scholar] [CrossRef]
- Butelli, E.; Garcia-Lor, A.; Licciardello, C.; Las Casas, G.; Hill, L.; Recupero, G.R.; Keremane, M.L.; Ramadugu, C.; Krueger, R.; Xu, Q.; et al. Changes in anthocyanin production during domestication of Citrus. Plant Physiol. 2017, 173, 2225–2242. [Google Scholar] [CrossRef]
- Zheng, X.; Om, K.; Stanton, K.A.; Thomas, D.; Cheng, P.A.; Eggert, A.; Simmons, E.; Yuan, Y.W.; Conradi Smith, G.D.; Puzey, J.R.; et al. The regulatory network for petal anthocyanin pigmentation is shaped by the MYB5a/NEGAN transcription factor in Mimulus. Genetics 2021, 217, 36. [Google Scholar] [CrossRef]
- Yadav, R.; Gorathoki, S.; Dhakal, S.; Purnima, B.; Shah, A.; Poudel, S. A Review on overview role of mutation in plant breeding. Rev. Food Agric. 2021, 2, 39–42. [Google Scholar] [CrossRef]
- Su, W.; Tao, R.; Liu, W.; Yu, C.; Yue, Z.; He, S.; Lavelle, D.; Zhang, W.; Zhang, L.; An, G.; et al. Characterization of four polymorphic genes controlling red leaf colour in lettuce that have undergone disruptive selection since domestication. Plant Biotechnol. J. 2020, 18, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, H.; Yang, Y.; Wang, H.; Xue, Z.; Fan, Y.; Sun, P.; Zhang, H.; Zhang, X.; Jin, W. Rosa1, a transposable element-like insertion, produces red petal coloration in Rose through altering RcMYB114 transcription. Front. Plant Sci. 2022, 13, 857684. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.N.; Hunt, H.D.; Horton, R.M.; Pullen, J.K.; Pease, L.R. Site-directed mutagenesis by overlap extension using the polymerase chain reaction. Gene 1989, 77, 51–59. [Google Scholar] [CrossRef]
- Li, M.; Yang, Y.; Wang, H.; Fan, Y.; Sun, P.; Jin, W. Identification and analysis of self incompatibility S-RNase in Rose. Acta Hortic. Sin. 2022, 49, 157–165. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, H.; Yang, Y.; Li, M.F.; Zhang, Y.T.; Liu, J.; Dong, J.; Li, J.; Butelli, E.; Xue, Z.; et al. The control of red colour by a family of MYB transcription factors in octoploid strawberry (Fragaria × ananassa) fruits. Plant Biotechnol. J. 2020, 18, 1169–1184. [Google Scholar] [CrossRef]
- Jin, W.; Wang, H.; Li, M.; Wang, J.; Yang, Y.; Zhang, X.; Yan, G.; Zhang, H.; Liu, J.; Zhang, K. The R2R3 MYB transcription factor PavMYB10.1 involves in anthocyanin biosynthesis and determines fruit skin colour in sweet cherry (Prunus avium L.). Plant Biotechnol. J. 2016, 14, 2120–2133. [Google Scholar] [CrossRef]
- Andersen, O.M.; Chen, K.; Du, L.; Liu, H.; Liu, Y. A novel R2R3-MYB from grape hyacinth, MaMybA, which is different from MaAN2, confers intense and magenta anthocyanin pigmentation in tobacco. Proc. Natl. Acad. Sci. USA 2019, 19, 20232–20239. [Google Scholar]
- Carey, C.C.; Strahle, J.T.; Selinger, D.A.; Chandler, V.L. Mutations in the pale aleurone color1 regulatory gene of the Zea mays anthocyanin pathway have distinct phenotypes relative to the functionally similar TRANSPARENT TESTA GLABRA1 gene in Arabidopsis thaliana. Plant Cell 2004, 16, 450–464. [Google Scholar] [CrossRef]
- Li, S. Transcriptional control of flavonoid biosynthesis: Fine-tuning of the MYB-bHLH-WD40 (MBW) complex. Plant Signal. Behav. 2014, 9, e27522. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, P.; Chen, G.; Wu, J.; Liu, Z.; Lian, H. FvbHLH9, functions as a positive regulator of anthocyanin biosynthesis, by forming HY5-bHLH9 transcription complex in strawberry fruits. Plant Cell Physiol. 2020, 16, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Liu, X.; Li, H.; Yin, X.; Grierson, D.; Li, F.; Chen, K. CmMYB#7, an R3 MYB transcription factor, acts as a negative regulator of anthocyanin biosynthesis in chrysanthemum. J. Exp. Bot. 2019, 70, 3111–3123. [Google Scholar] [PubMed]
- Xu, J.; Xu, H.; Liu, Y.; Wang, X.; Xu, Q.; Deng, X. Genome-wide identification of sweet orange (Citrus sinensis) histone modification gene families and their expression analysis during the fruit development and fruit-blue mold infection process. Front. Plant Sci. 2015, 6, 607. [Google Scholar] [CrossRef]
- Falcone Ferreyra, M.L.; Rius, S.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef]
- Yin, X.; Zhang, Y.; Zhang, L.; Wang, B.; Zhao, Y.; Irfan, M.; Chen, L.; Feng, Y. Regulation of MYB transcription factors of anthocyanin synthesis in Lily flowers. Front. Plant Sci. 2021, 12, 761668. [Google Scholar] [CrossRef]
- Luo, Y.; Xu, X.; Yang, L.; Zhu, X.; Du, Y.; Fang, Z. A R2R3-MYB transcription factor, FeR2R3-MYB, positively regulates anthocyanin biosynthesis and drought tolerance in common buckwheat (Fagopyrum esculentum). Plant Physiol. Biochem. 2024, 217, 109254. [Google Scholar] [CrossRef]
- Li, C.; Qiu, J.; Huang, S.; Yin, J.; Yang, G. AaMYB3 interacts with AabHLH1 to regulate proanthocyanidin accumulation in Anthurium andraeanum (Hort.)-another strategy to modulate pigmentation. Mol. Genet. Genom. 2019, 294, 469–478. [Google Scholar] [CrossRef]
- Jiu, S.; Guan, L.; Leng, X.; Zhang, K.; Haider, M.S.; Yu, X.; Zhu, X.; Zheng, T.; Ge, M.; Wang, C.; et al. The role of VvMYBA2r and VvMYBA2w alleles of the MYBA2 locus in the regulation of anthocyanin biosynthesis for molecular breeding of grape (Vitis spp.) skin coloration. Plant Biotechnol. J. 2021, 19, 1216–1239. [Google Scholar] [CrossRef]
- Hichri, I.; Barrieu, F.; Bogs, J.; Kappel, C.; Delrot, S.; Lauvergeat, V. Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. J. Exp. Bot. 2011, 62, 2465–2483. [Google Scholar] [CrossRef]
- Quattrocchio, F.; Wing, J.; van der Woude, K.; Souer, E.; de Vetten, N.; Mol, J.; Koes, R. Molecular analysis of the anthocyanin2 gene of petunia and its role in the evolution of flower color. Plant Cell 1999, 11, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, L.-J.; Wang, Y.; Geng, Z.; Liu, S.; Chen, C.; Chen, S.; Jiang, J.; Chen, F. CmMYB9a activates floral coloration by positively regulating anthocyanin biosynthesis in chrysanthemum. Plant Mol. Biol. 2022, 108, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.C.; Rausher, M.D. R2R3-MYB genes control petal pigmentation patterning in Clarkia gracilis ssp. sonomensis (Onagraceae). New Phytol. 2021, 229, 1147–1162. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, M.; Shimoyamada, Y.; Nakatsuka, T.; Masuda, K. Two R2R3-MYB genes, homologs of Petunia AN2, regulate anthocyanin biosyntheses in flower tepals, tepal spots and leaves of asiatic hybrid lily. Plant Cell Physiol. 2010, 51, 463–474. [Google Scholar] [CrossRef]
- Yamagishi, M. Oriental hybrid lily Sorbonne homologue of LhMYB12 regulates anthocyanin biosyntheses in flower tepals and tepal spots. Mol. Breed. 2011, 28, 381–389. [Google Scholar] [CrossRef]
- Yamagishi, M. A novel R2R3-MYB transcription factor regulates light-mediated floral and vegetative anthocyanin pigmentation patterns in Lilium regale. Mol. Breed. 2015, 36, 3. [Google Scholar] [CrossRef]
- Yamagishi, M.; Ihara, H.; Arakawa, K.; Toda, S.; Suzuki, K. The origin of the LhMYB12 gene, which regulates anthocyanin pigmentation of tepals, in oriental and Asiatic hybrid lilies (Lilium spp.). Sci. Hortic. 2014, 174, 119–125. [Google Scholar] [CrossRef]
- Yamagishi, M.; Yoshida, Y.; Nakayama, M. The transcription factor LhMYB12 determines anthocyanin pigmentation in the tepals of Asiatic hybrid lilies (Lilium spp.) and regulates pigment quantity. Mol. Breed. 2012, 30, 913–925. [Google Scholar] [CrossRef]
- Yamagishi, M.; Toda, S.; Tasaki, K. The novel allele of the LhMYB12 gene is involved in splatter-type spot formation on the flower tepals of Asiatic hybrid lilies (Lilium spp.). New Phytol. 2014, 201, 1009–1020. [Google Scholar] [CrossRef]
- Ben-Simhon, Z.; Judeinstein, S.; Nadler-Hassar, T.; Trainin, T.; Bar-Ya’akov, I.; Borochov-Neori, H.; Holland, D. A pomegranate (Punica granatum L.) WD40-repeat gene is a functional homologue of Arabidopsis TTG1 and is involved in the regulation of anthocyanin biosynthesis during pomegranate fruit development. Planta 2011, 234, 865–881. [Google Scholar] [CrossRef]
- Lim, S.H.; Kim, D.H.; Kim, J.K.; Lee, J.Y.; Ha, S.H. A Radish basic helix-loop-helix transcription factor, RsTT8 acts a positive regulator for anthocyanin biosynthesis. Front. Plant Sci. 2017, 8, 1917. [Google Scholar] [CrossRef] [PubMed]
- Lai, B.; Du, L.N.; Liu, R.; Hu, B.; Su, W.B.; Qin, Y.H.; Zhao, J.T.; Wang, H.C.; Hu, G.B. Two LcbHLH transcription factors interacting with LcMYB1 in regulating late structural genes of anthocyanin biosynthesis in Nicotiana and Litchi chinensis during anthocyanin accumulation. Front. Plant Sci. 2016, 7, 166. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-F.; Yin, X.-R.; Allan, A.C.; Lin-Wang, K.; Shi, Y.-N.; Huang, Y.-J.; Ferguson, I.B.; Xu, C.-J.; Chen, K.-S. The role of MrbHLH1 and MrMYB1 in regulating anthocyanin biosynthetic genes in tobacco and Chinese bayberry (Myrica rubra) during anthocyanin biosynthesis. Plant Cell Tissue Organ Cult. (PCTOC) 2013, 115, 285–298. [Google Scholar] [CrossRef]
- Chioti, V.; Zeliou, K.; Bakogianni, A. Nutritional value of eggplant cultivars and association with sequence variation in genes coding for major phenolics. Plants 2022, 11, 2267. [Google Scholar] [CrossRef]
- Wang, X.; Olsen, O.; Knudsen, S. Expression of the dihydroflavonol reductase gene in an anthocyanin-free barley mutant. Hereditas 1993, 119, 67–75. [Google Scholar] [CrossRef]
- Xiang, F.; Liu, W.C. Direct balancing of lipid mobilization and ROS production by the epoxidation of fatty acid catalyzed by a cytochrome P450 protein during seed germination. New Phytol. 2022, 237, 2104–2117. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Purpose | Primer Sequence (5′→3′) |
|---|---|---|
| RcMYB114a | Gene clone | F: CAACGTCATTAACTGTGGGATC |
| R: GCCGTGAGCAGTGGCTTTC | ||
| RcMYB114a, RcMYB114b, RcMYB114c, and RcMYB114d | Transient expression | F:GAAATTtctagaATGGAGGACCAGTCGGGTTTGAG |
| R: GAAATTgagctcTCATTATCGATCTAAGAATGTCATCC | ||
| RcbHLH | F:GAAATTtcgcgaATGGCTACACCGCCACCGAGTAGTAGC | |
| R: GAAATTaccggtTTAAGAGTCAGATTGGGGTATCAC | ||
| RcMYB114c | site-directed mutagenesis | F: CCACAAAGATTGTTGAAGCCCATCATAC |
| R: AGTATGATGGGCTTCAACAATCTTTGTG | ||
| RcMYB114d | F: TTGACCCCTGAAAGATTATTTATTGAAG | |
| R: ATCTTCAATAAATAATCTTTCAGGGGTC | ||
| RcMYB114a:AD, RcMYB114b:AD, RcMYB114c:AD, and RcMYB114d:AD | Yeast two-hybrid assays | F: GAAATTcatatgATGGAGGACCAGTCGGGTTTGAG |
| R: GAAATTgaattcTCATTATCGATCTAAGAATGTCATCC | ||
| RcMYB114a:BK, RcMYB114b:BK, RcMYB114c:BK, and RcMYB114d:BK | F: GAAATTcatatgATGGAGGACCAGTCGGGTTTGAG | |
| R: GAAATTgaattcTCATTATCGATCTAAGAATGTCATCC | ||
| RcWD40:AD and RcWD40:BK | F: GAAATTcatatgATGGAGAACTCGACCCAAGAATC | |
| R: GAAATTgaattcTCAAACCTTCAACAGCTGCATCTTA | ||
| RcbHLH:AD and RcbHLH:BK | F: GAAATTggatccATGGCTACACCGCCACCGAGTAGTAGC | |
| R: GAAATTgaattcTCATTAAGAGTCAGATTGGGGTATCAC | ||
| RcMYB114a | RT-qPCR | F:ACCAAGCGGCGTCGGGACAAAC |
| R: CCCGTCAAACAGAGTGAACTGGTCG | ||
| RcC4H | F:ATGTTCGACAGGAGATTTGAAAGCG | |
| R:ATTATACTCGAAGCTCTGCGCCAAC | ||
| RcCHI | F:TTTCCTCCCGCCGTCAAGCC | |
| R:CCAAGTAGACTCCAATCGCCGTGAA | ||
| RcCHS | F:CTACTTTCGTATCACCAACAGCG | |
| R:TTCAGTCAAATACATATAACGCTTC | ||
| RcF3H | F:GCTCCAGGACCAAGTCGGTGGACT | |
| R:TGATCGGCGTTCTTGAACCTCCC | ||
| RcANS | F:GGAACTTGCCCTCGGCGTGG | |
| R:ATGACGATGGAGTTGGGCACGC | ||
| RcPAL | F:AAGATTTTGCGAGAAGGATTTGC | |
| R:GCATTCTTCTCATTCTCACCATTTGT | ||
| RcUFGT | F:GCCCCAAACACCCTCTTCTCA | |
| R:CCTGAGGCTTACCCACAAAAACA | ||
| RcDFR | F:CACCGTGCGAGACCCTGCTAA | |
| R:TCAAAATCCATAGGAGTGGCGACA | ||
| RcActin | F: GGCTGTTCTTTCCCTCTATGC | |
| R:GCGTTTCAGATGCCCAGAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Yang, Y.; Wang, H.; Sun, P.; Zhou, S.; Kang, Y.; Sun, X.; Jin, M.; Jin, W. The Mutations in RcMYB114 Affect Anthocyanin Glycoside Accumulation in Rose. Biology 2025, 14, 258. https://doi.org/10.3390/biology14030258
Li M, Yang Y, Wang H, Sun P, Zhou S, Kang Y, Sun X, Jin M, Jin W. The Mutations in RcMYB114 Affect Anthocyanin Glycoside Accumulation in Rose. Biology. 2025; 14(3):258. https://doi.org/10.3390/biology14030258
Chicago/Turabian StyleLi, Maofu, Yuan Yang, Hua Wang, Pei Sun, Shuting Zhou, Yanhui Kang, Xiangyi Sun, Min Jin, and Wanmei Jin. 2025. "The Mutations in RcMYB114 Affect Anthocyanin Glycoside Accumulation in Rose" Biology 14, no. 3: 258. https://doi.org/10.3390/biology14030258
APA StyleLi, M., Yang, Y., Wang, H., Sun, P., Zhou, S., Kang, Y., Sun, X., Jin, M., & Jin, W. (2025). The Mutations in RcMYB114 Affect Anthocyanin Glycoside Accumulation in Rose. Biology, 14(3), 258. https://doi.org/10.3390/biology14030258