
Diversity and Life History Traits of Native Weed Communities in Agricultural Areas: A Case Study in Eastern China

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
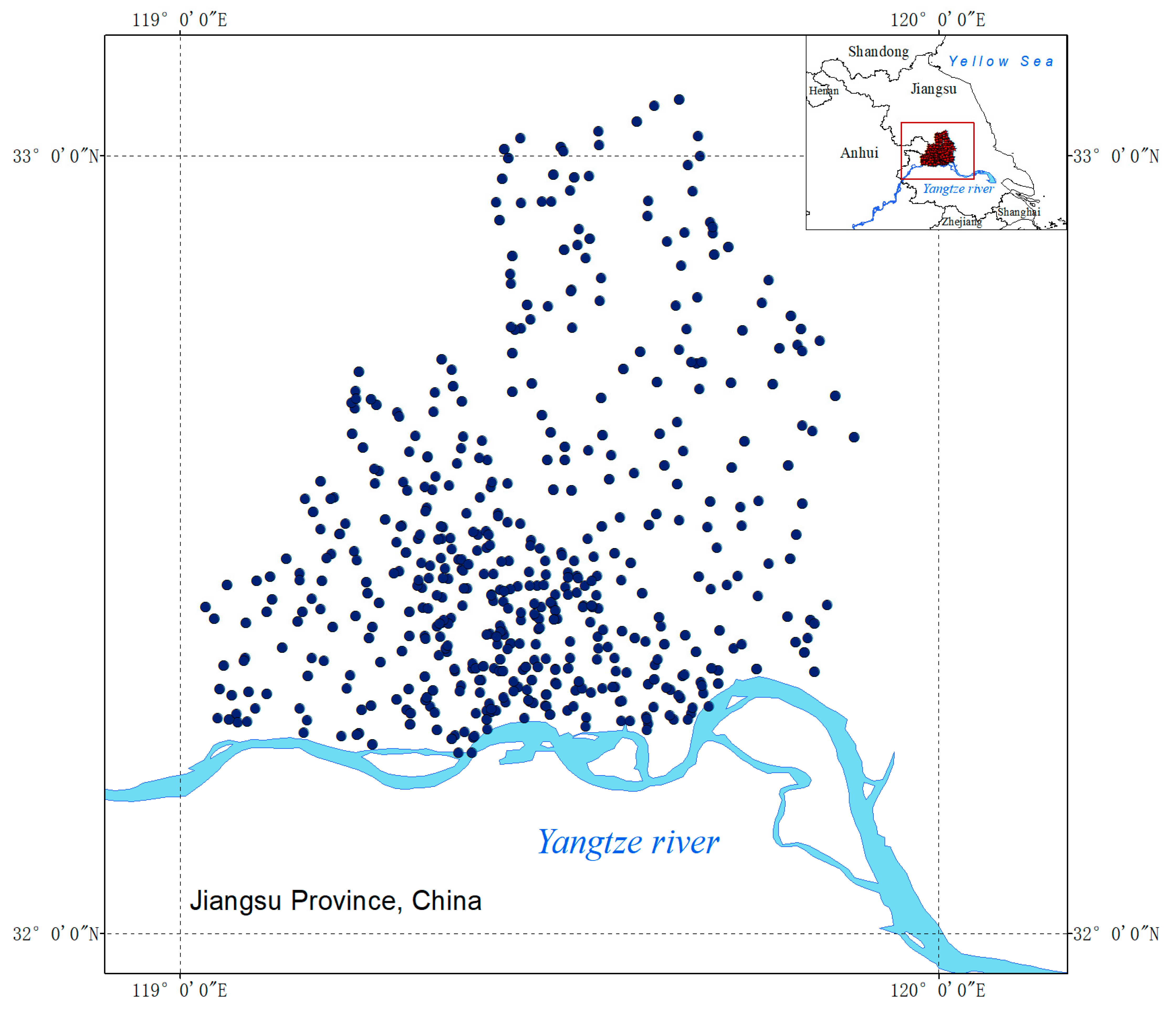
2.1. Investigating Area
2.2. Investigation Method
2.3. Traits of Weeds
2.4. Data Statistical Analysis
3. Result
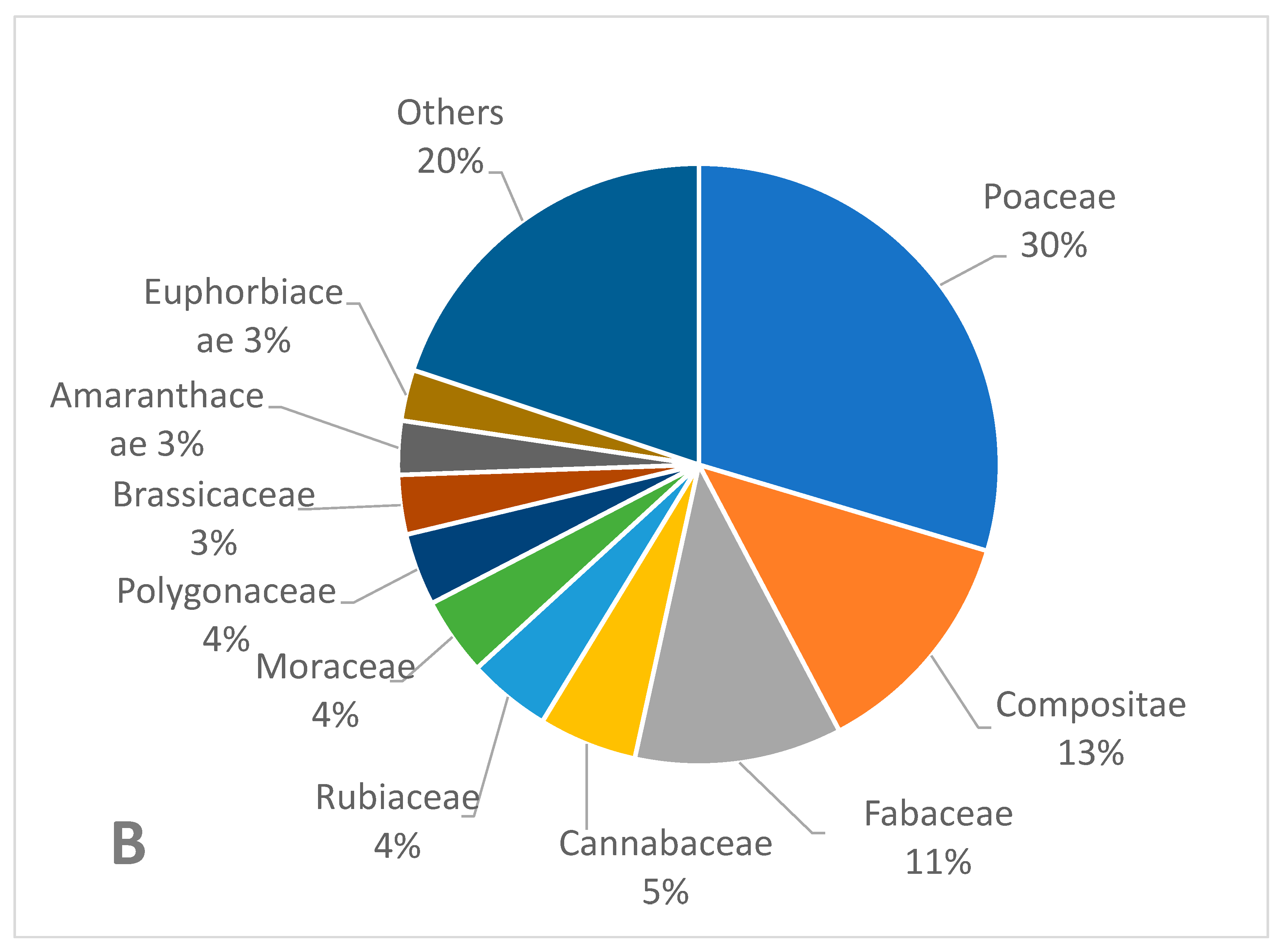
3.1. Weed Species Recorded in the Survey
3.2. Plant Height
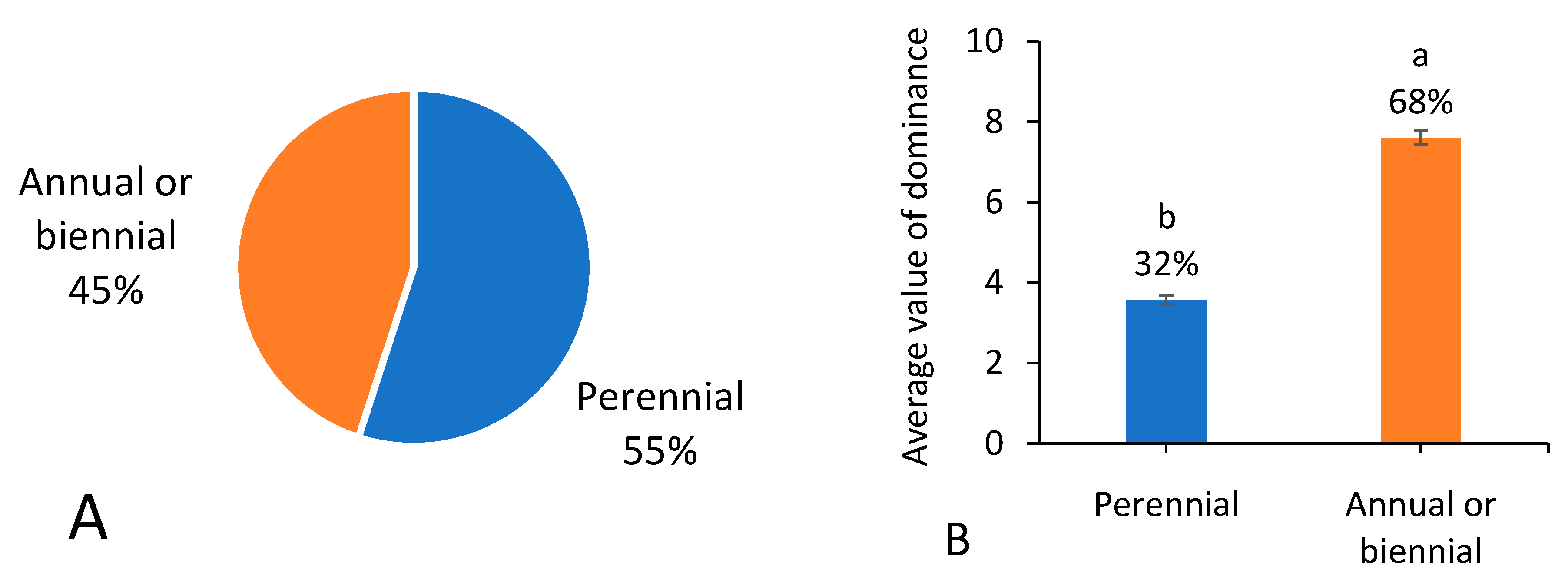
3.3. Life Type
3.4. Drought Resistance
3.5. Shade Tolerance
3.6. Life History Trait
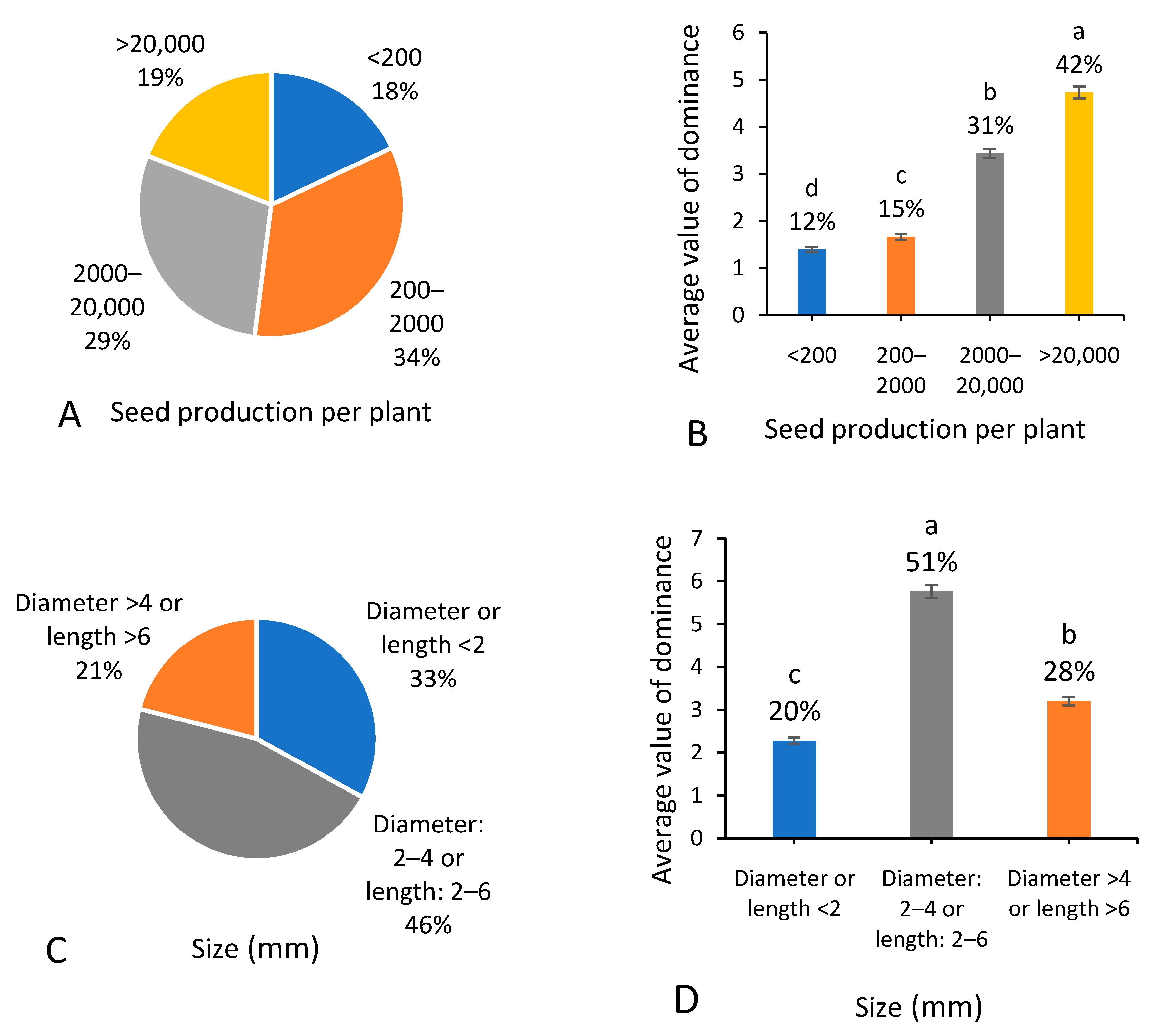
3.7. Reproductive Characteristics
3.8. Appendages of Fruits
3.9. Herbicide Resistance and Weediness
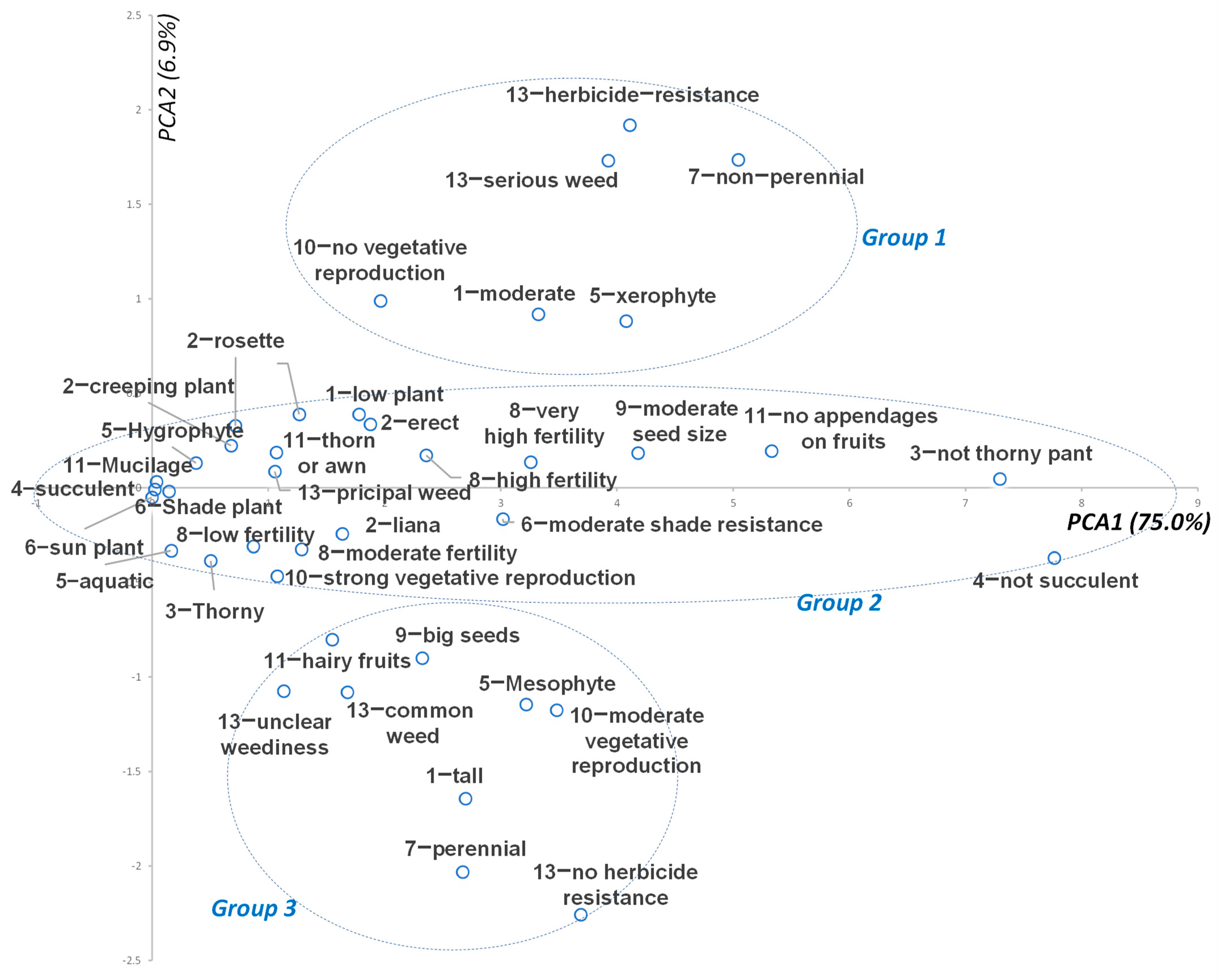
3.10. PCA of Weed Traits
4. Discussion
4.1. Native Weed Diversity in Agricultural Areas
4.2. Traits Closely Related to Weediness in Agricultural Areas
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baker, H.G. The Evolution of Weeds. Annu. Rev. Ecol. Syst. 1974, 5, 1–24. [Google Scholar] [CrossRef]
- Oerke, E.C. Crop Losses to Pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Spahillari, M.; Hammer, K.; Gladis, T.; Diederichsen, A. Weeds as Part of Agrobiodiversity. Outlook Agric. 1999, 4, 227–232. [Google Scholar] [CrossRef]
- Barroso, J.; Miller, Z.J.; Lehnhoff, E.A.; Hatfield, P.G.; Menalled, F.D. Impacts of Cropping System and Management Practices on the Assembly of Weed Communities. Weed Res. 2015, 4, 426–435. [Google Scholar] [CrossRef]
- Wang, F.; Huang, J.; Zhang, N.; Li, Y.; He, S.; Wen, J.; Yin, L.; Liang, Y. Exploring Plant Characteristics for Constructing a Pre-Border Weed Risk Assessment for China. Biol. Invasions 2024, 4, 909–933. [Google Scholar] [CrossRef]
- He, S.; Yin, L.; Wen, J.; Liang, Y. A Test of the Australian Weed Risk Assessment System in China. Biol. Invasions 2018, 8, 2061–2076. [Google Scholar] [CrossRef]
- Hulme, P.E. Weed Risk Assessment: A Way Forward or a Waste of Time? J. Appl. Ecol. 2012, 1, 10–19. [Google Scholar] [CrossRef]
- Pheloung, P.C.; Williams, P.A.; Halloy, S.R. A Weed Risk Assessment Model for Use as a Biosecurity Tool Evaluating Plant Introductions. J. Environ. Manag. 1999, 4, 239–251. [Google Scholar] [CrossRef]
- Koop, A.L.; Fowler, L.; Newton, L.P.; Caton, B.P. Development and Validation of a Weed Screening Tool for the United States. Biol. Invasions 2012, 2, 273–294. [Google Scholar] [CrossRef]
- Andreu, J.; Vilà, M. Risk Analysis of Potential Invasive Plants in Spain. J. Nat. Conserv. 2010, 1, 34–44. [Google Scholar] [CrossRef]
- Weber, E.; Gut, D. Assessing the Risk of Potentially Invasive Plant Species in Central Europe. J. Nat. Conserv. 2004, 3, 171–179. [Google Scholar] [CrossRef]
- Baker, H.G. Characteristics and Modes of Origin of Weeds. Agric. Food Sci. 1965, 147–168. [Google Scholar]
- Pyšek, P.; Richardson, D.M. Traits Associated with Invasiveness in Alien Plants: Where Do We Stand? In Biological Invasions; Springer: Berlin/Heidelberg, Germany, 2007; pp. 97–125. [Google Scholar] [CrossRef]
- Hamilton, M.A.; Murray, B.R.; Cadotte, M.W.; Hose, G.C.; Baker, A.C.; Harris, C.J.; Licari, D. Life-History Correlates of Plant Invasiveness at Regional and Continental Scales. Ecol. Lett. 2005, 10, 1066–1074. [Google Scholar] [CrossRef]
- Sutherland, S. What Makes a Weed a Weed: Life History Traits of Native and Exotic Plants in the USA. Oecologia 2004, 1, 24–39. [Google Scholar] [CrossRef]
- Khattak, W.A.; Sun, J.; Hameed, R.; Zaman, F.; Abbas, A.; Khan, K.A.; Elboughdiri, N.; Akbar, R.; He, F.; Ullah, M.W.; et al. Unveiling the Resistance of Native Weed Communities: Insights for Managing Invasive Weed Species in Disturbed Environments. Biol. Rev. 2024, 3, 753–777. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wen, J.; Luo, D.; Zhu, C. Response Characteristics of Soil Moisture to Rainfall for a Single Grass Type in an Urban Area. Hydrol. Res. 2023, 2, 99–115. [Google Scholar] [CrossRef]
- Qiang, S. Multivariate Analysis, Description, and Ecological Interpretation of Weed Vegetation in the Summer Crop Fields of Anhui Province, China. J. Integr. Plant Biol. 2005, 10, 1193–1210. [Google Scholar] [CrossRef]
- Chen, G.; He, Y.; Qiang, S. Increasing Seriousness of Plant Invasions in Croplands of Eastern China in Relation to Changing Farming Practices: A Case Study. PLoS ONE 2013, 8, e74136. [Google Scholar] [CrossRef]
- Beckie, H.J.; Owen, M.J.; Borger, C.P.D.; Gill, G.S.; Widderick, M.J. Agricultural Weed Assessment Calculator: An Australian Evaluation. Plants 2020, 9, 1737. [Google Scholar] [CrossRef]
- Ou, H.; Lu, C.; O’Toole, D.K. A Risk Assessment System for Alien Plant Bio-Invasion in Xiamen, China. J. Environ. Sci. 2008, 8, 989–997. [Google Scholar] [CrossRef]
- Barkley, T.M. A Geographical Atlas of World Weeds. Brittonia 1980, 32, 127. [Google Scholar] [CrossRef]
- Kraehmer, H. Specialization of Weeds and Biodiversity. In Atlas of Weed Mapping; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016. [Google Scholar]
- Li, C. Weed Flora of China; Science Press: Beijing, China, 1998. [Google Scholar]
- Ma, J. Alien Invasive Flora of China; Shanghai Jiao Tong University Press: Shanghai, China, 2020. [Google Scholar]
- Liu, X.; Weng, X.; Edwards, J.; Wang, L.; Zhang, C.; Qiu, J.; Li, Z. Editorial: Adaptive Evolution of Grasses. Front. Plant Sci. 2023, 14, 1105320. [Google Scholar] [CrossRef] [PubMed]
- Edwards, E.J.; Smith, S.A. Phylogenetic Analyses Reveal the Shady History of Grasses. Proc. Natl. Acad. Sci. USA 2010, 6, 2532–2537. [Google Scholar] [CrossRef] [PubMed]
- Stroemberg, C.A.E. Evolution of Grasses and Grassland Ecosystems. Annu. Rev. Earth Planet. Sci. 2011, 39, 517–544. [Google Scholar] [CrossRef]
- Xu, K.; Shu, L.; Xie, Q.; Song, M.; Zhu, Y.; Cao, W.; Ni, J. Precision Weed Detection in Wheat Fields for Agriculture 4.0, A Survey of Enabling Technologies, Methods, and Research Challenges. Comput. Electron. Agric. 2023, 212, 108106. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, J.; DiTommaso, A.; Zhang, C.; Zheng, G.; Liang, W.; Islam, F.; Yang, C.; Chen, X.; Zhou, W. Weed Research Status, Challenges, and Opportunities in China. Crop Prot. 2020, 134, 104449. [Google Scholar] [CrossRef]
- Zhang, C.; Huang, C.H.; Liu, M.; Hu, Y.; Panero, J.L.; Luebert, F.; Gao, T.; Ma, H. Phylotranscriptomic Insights into Asteraceae Diversity, Polyploidy, and Morphological Innovation. J. Integr. Plant Biol. 2021, 7, 1273–1293. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seed Dormancy in Asteraceae: A Global Vegetation Zone and Taxonomic/Phylogenetic Assessment. Seed Sci. Res. 2023, 2, 135–169. [Google Scholar] [CrossRef]
- Funk, V.A.; Bayer, R.J.; Keeley, S.; Chan, R.; Watson, L.; Gemeinholzer, B.; Schilling, E.; Panero, J.L.; Baldwin, B.G.; Garcia-Jacas, N.; et al. but Antarctica: Using a Supertree to Understand the Diversity and Distribution of the Compositae. Biol. Skr. K. Dan. Vidensk. Selsk. 2005, 55, 343–374. [Google Scholar]
- Abdel Bar, F.M.; Abdel Fatah, N.H.; Amen, Y.; Halim, A.F.; Saad, H.E. Genus Lactuca (Asteraceae): A Comprehensive Review. Rec. Nat. Prod. 2023, 2, 201–231. [Google Scholar] [CrossRef]
- Kostić, A.Ž.; Janaćković, P.; Kolašinac, S.M.; Dajić Stevanović, Z.P. Balkans’ Asteraceae Species as a Source of Biologically Active Compounds for the Pharmaceutical and Food Industry. Chem. Biodivers. 2020, 17, e2000097. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Pergl, J.; Essl, F.; Lenzner, B.; Dawson, W.; Kreft, H.; Weigelt, P.; Winter, M.; Kartesz, J.; Nishino, M.; et al. Naturalized Alien Flora of the World: Species Diversity, Taxonomic and Phylogenetic Patterns, Geographic Distribution and Global Hotspots of Plant Invasion. Preslia 2017, 3, 203–274. [Google Scholar] [CrossRef]
- Christenhusz, M.J.; Byng, J.W. The Number of Known Plants Species in the World and Its Annual Increase. Phytotaxa 2016, 3, 201–217. [Google Scholar] [CrossRef]
- Azani, N.; Babineau, M.; Bailey, C.D.; Banks, H.; Barbosa, A.R.; Pinto, R.B.; Boatwright, J.S.; Borges, L.M.; Brown, G.K.; Bruneau, A.; et al. A New Subfamily Classification of the Leguminosae Based on a Taxonomically Comprehensive Phylogeny. Taxon 2017, 1, 44–77. [Google Scholar] [CrossRef]
- Maroyi, A. Medicinal Uses of the Fabaceae Family in Zimbabwe: A Review. Plants 2023, 12, 1255. [Google Scholar] [CrossRef]
- Usman, M.; Khan, W.R.; Yousaf, N.; Akram, S.; Murtaza, G.; Kudus, K.A.; Ditta, A.; Rosli, Z.; Rajpar, M.N.; Nazre, M. Exploring the Phytochemicals and Anti-Cancer Potential of the Members of Fabaceae Family: A Comprehensive Review. Molecules 2022, 27, 3863. [Google Scholar] [CrossRef]
- Gulzar, R.; Wani, S.A.; Hassan, T.; Reddy, C.S.; Shrestha, B.B.; Mukul, S.A.; Shabbir, A.; Iqbal, I.M.; Ranwala, S.M.; Dorjee Sujanapal, P. Looking Beyond the Political Boundaries: An Integrated Inventory of Invasive Alien Flora of South Asia. Biol. Invasions 2023, 26, 57–78. [Google Scholar] [CrossRef]
- Arianoutsou, M.; Bazos, I.; Christopoulou, A.; Kokkoris, Y.; Zikos, A.; Zervou, S.; Delipetrou, P.; Cardoso, A.C.; Deriu, I.; Gervasini, E.; et al. Alien Plants of Europe: Introduction Pathways, Gateways and Time Trends. Peerj 2021, 9, e11270. [Google Scholar] [CrossRef]
- Qiang, H.; Ma, J. Invasive Alien Plants in China: An Update. Plant Divers. 2023, 1, 117–121. [Google Scholar] [CrossRef]
- Kraehmer, H.; Jabran, K.; Mennan, H.; Chauhan, B.S. Global Distribution of Rice Weeds a Review. Crop Prot. 2016, 80, 73–86. [Google Scholar] [CrossRef]
- Zhu, J. Investigation and Risk Assessment of Invasive Alien Plants in Ningbo City. Master’s Thesis, University of Ningbo, Ningbo, China, 2019. [Google Scholar]
- Barreto, L.S. The Reconciliation of the R-K, and C-S-R-Models for Life-History Strategies. Silva Lusit. 2008, 1, 97–103. [Google Scholar]
- Liu, Y.; Wu, H.; Wang, C.; Cheng, J.; Qiang, S. A Comparative Study Reveals the Key Biological Traits Causing Bioinvasion Differences among Four Alien Species of Genus Veronica in China. J. Plant Ecol. 2023, 16, rtac068. [Google Scholar] [CrossRef]
- Cavero, J.; Zaragoza, C.; Suso, M.L.; Pardo, A. Competition between Maize and Datura Stramonium in an Irrigated Field under Semi-Arid Conditions. Weed Res. 1999, 3, 225–240. [Google Scholar] [CrossRef]
- Gaba, S.; Perronne, R.; Fried, G.; Gardarin, A.; Bretagnolle, F.; Biju-Duval, L.; Colbach, N.; Cordeau, S.; Fernández-Aparicio, M.; Gauvrit, C.; et al. Response and Effect Traits of Arable Weeds in Agro-Ecosystems: A Review of Current Knowledge. Weed Res. 2017, 3, 123–147. [Google Scholar] [CrossRef]
- Daehler, C.C. The Taxonomic Distribution of Invasive Angiosperm Plants: Ecological Insights and Comparison to Agricultural Weeds. Biol. Conserv. 1998, 2, 167–180. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, B.; Qiang, S. Analyses of Species Composition and Diversity of Weed Seed Bank of Main Crop Fields in Jiangsu Province and Its Correlation with Environmental Factors. J. Plant Resour. Environ. 2012, 1, 1–13. [Google Scholar]
- Barres, B.; Corio-Costet, M.F.; Debieu, D.D.; Délye, C.; Fillinger-David, S.; Grosman, J.; Micoud, A.; Siegwart, M.; Walker, A.S. Trends and Challenges in Pesticide Resistance Detection. Trends Plant Sci. 2016, 10, 834–853. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in Action: Plants Resistant to Herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef]
- Matshidze, M.M.; Ndou, V. Herbicide Resistance Cases in South Africa: A Review of the Current State of Knowledge. S. Afr. J. Sci. 2023, 119, 11–12. [Google Scholar] [CrossRef]
- Ruzmi, R.; Ahmad-Hamdani, M.S.; Bakar, B.B. Prevalence of Herbicide-Resistant Weed Species in Malaysian Rice Fields: A Review. Weed Biol. Manag. 2017, 1, 3–16. [Google Scholar] [CrossRef]

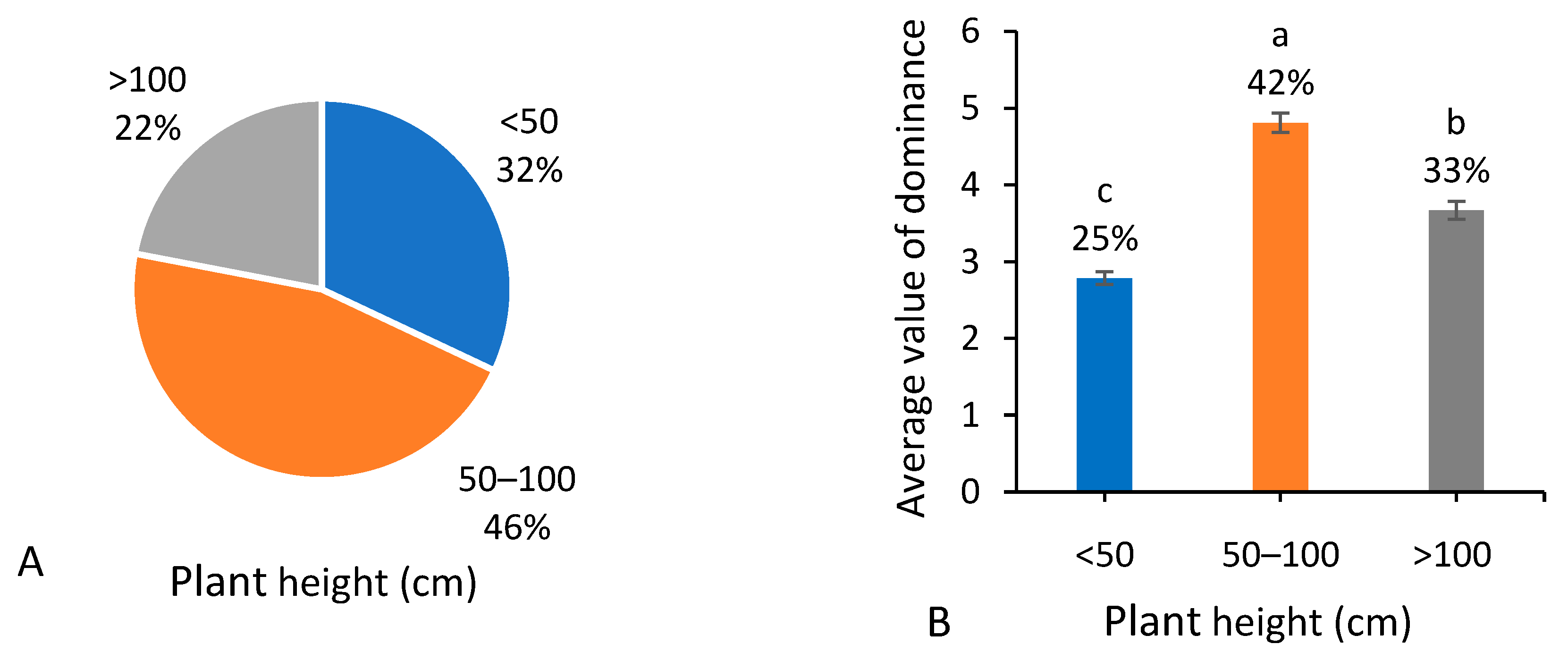




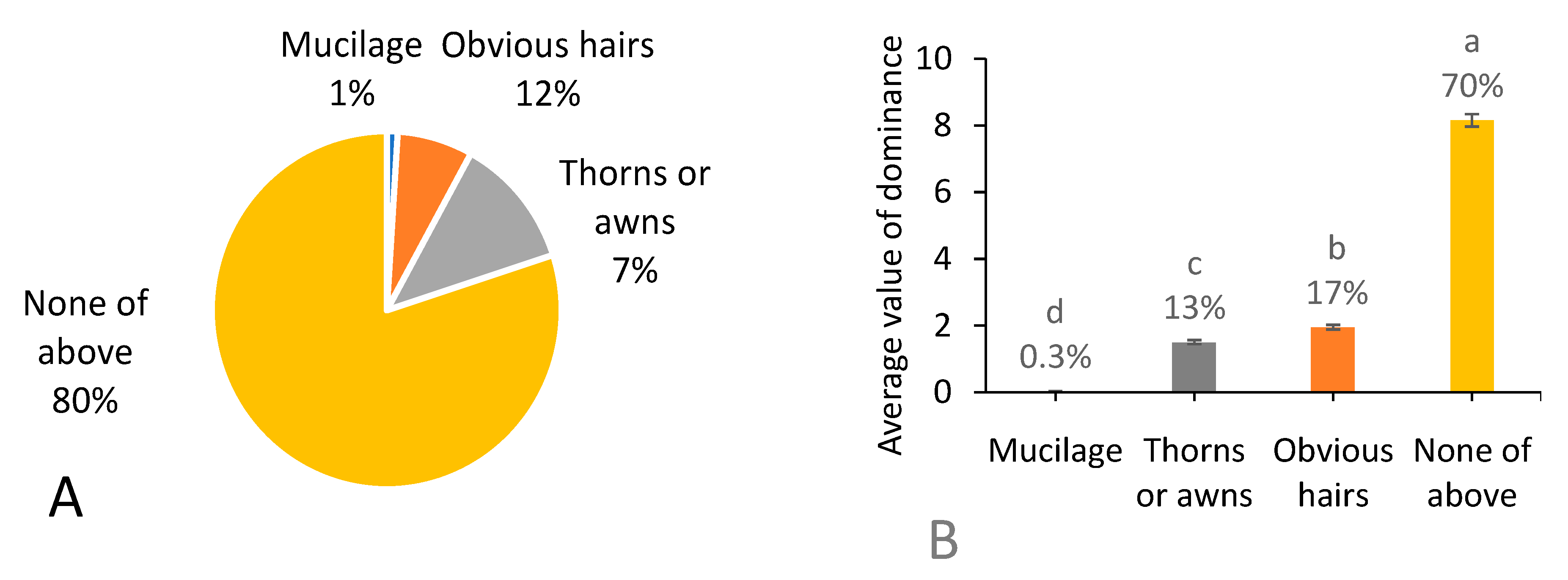
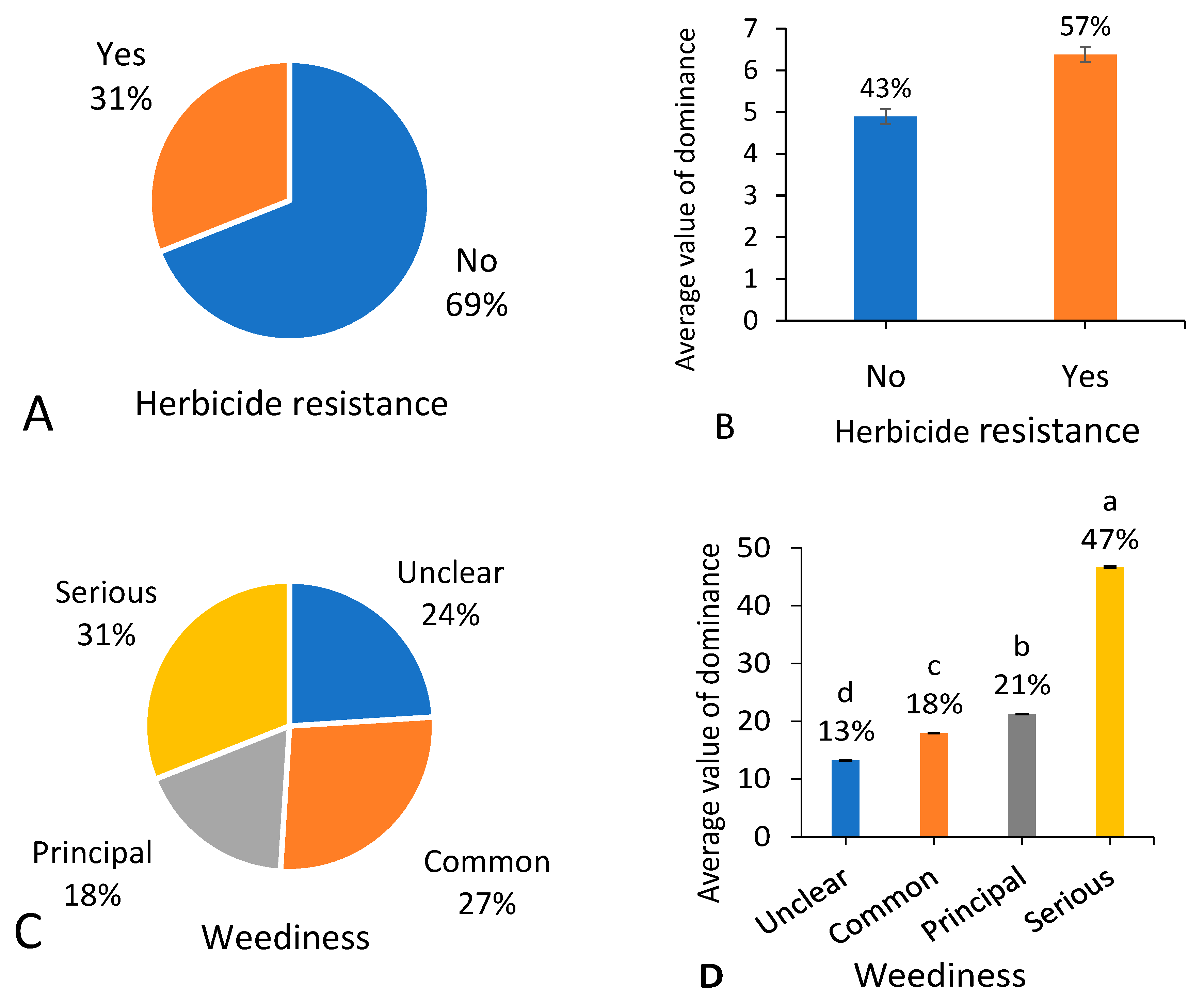

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Maximum Height of Weeds in the Field | ||
|---|---|---|---|
| >80 cm a | 20 cm–80 cm b | <20 cm c | |
| 0.1 | 1–3 stems or total coverage <0.1% | <10 stems or total coverage <1% | <15 stems or total coverage <2% |
| 0.5 | 4–10 stems or total coverage 0.2–0.9% | 11–15 stems or total coverage 1–2% | 16–30 stems or total coverage 3–5% |
| 1 | 11–15 stems or total coverage 1–2% | 16–30 stems or total coverage 3–5% | 31–60 stems or total coverage 6–10% |
| 2 | 16–30 stems or total coverage 3–5% | 31–60 stems or total coverage 6–10% | 61–100 stems or total coverage 11–25% |
| 3 | 31–60 stems or total coverage 6–10% | 61–100 stems or total coverage 11–25% | 101–200 stems or total coverage 25–50% |
| 4 | 61–100 stems or total coverage 11–25% | 101–200 stems or total coverage 25–50% | 201–500 stems or total coverage 50–90% |
| 5 | >100 stems or total coverage >25% | >200 stems or total coverage >50% | >500 stems or total coverage >90% |
| Code | Indicator | Traits |
|---|---|---|
| 1 | Plant height | >100 cm (tall), 50–100 cm (moderate), or <50 cm (low) |
| 2 | Life type | Rosette, erect, creeping, or liana |
| 3 | Thorns or hooks | Yes or no |
| 4 | Succulent | Yes or no |
| 5 | Drought resistance | Aquatic, hygrophyte, mesophyte, or xerophyte |
| 6 | Shade tolerance | Shade plant, moderate, sun plant |
| 7 | Life span | Annual or biennial or perennial |
| 8 | Seed production per plant | <200 (low fertility), 200–2000 (moderate fertility), 2000–20,000 (high fertility), or >20,000 (very high fertility) |
| 9 | Size (mm) | Diameter or length < 2 (tiny), 2–4 in diameter or 2–6 in length (moderate), or diameter > 4 or length > 6 (big) |
| 10 | Vegetative reproduction | No, moderate, or strong |
| 11 | Appendages of fruits | Mucilage, obvious hairs, thorns or awns, or none of the above |
| 12 | Herbicide resistance a | Yes or no |
| 13 | Weediness b | Unclear, common, principal, or serious |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, G.; Huang, Z.; An, K.; Chen, Y.; Xue, J. Diversity and Life History Traits of Native Weed Communities in Agricultural Areas: A Case Study in Eastern China. Biology 2024, 13, 704. https://doi.org/10.3390/biology13090704
Chen G, Huang Z, An K, Chen Y, Xue J. Diversity and Life History Traits of Native Weed Communities in Agricultural Areas: A Case Study in Eastern China. Biology. 2024; 13(9):704. https://doi.org/10.3390/biology13090704
Chicago/Turabian StyleChen, Guoqi, Zeyue Huang, Kai An, Yang Chen, and Jiahao Xue. 2024. "Diversity and Life History Traits of Native Weed Communities in Agricultural Areas: A Case Study in Eastern China" Biology 13, no. 9: 704. https://doi.org/10.3390/biology13090704
APA StyleChen, G., Huang, Z., An, K., Chen, Y., & Xue, J. (2024). Diversity and Life History Traits of Native Weed Communities in Agricultural Areas: A Case Study in Eastern China. Biology, 13(9), 704. https://doi.org/10.3390/biology13090704