
High-Throughput Sequencing Analysis Revealed a Preference for Animal-Based Food in Purple Sea Urchins



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
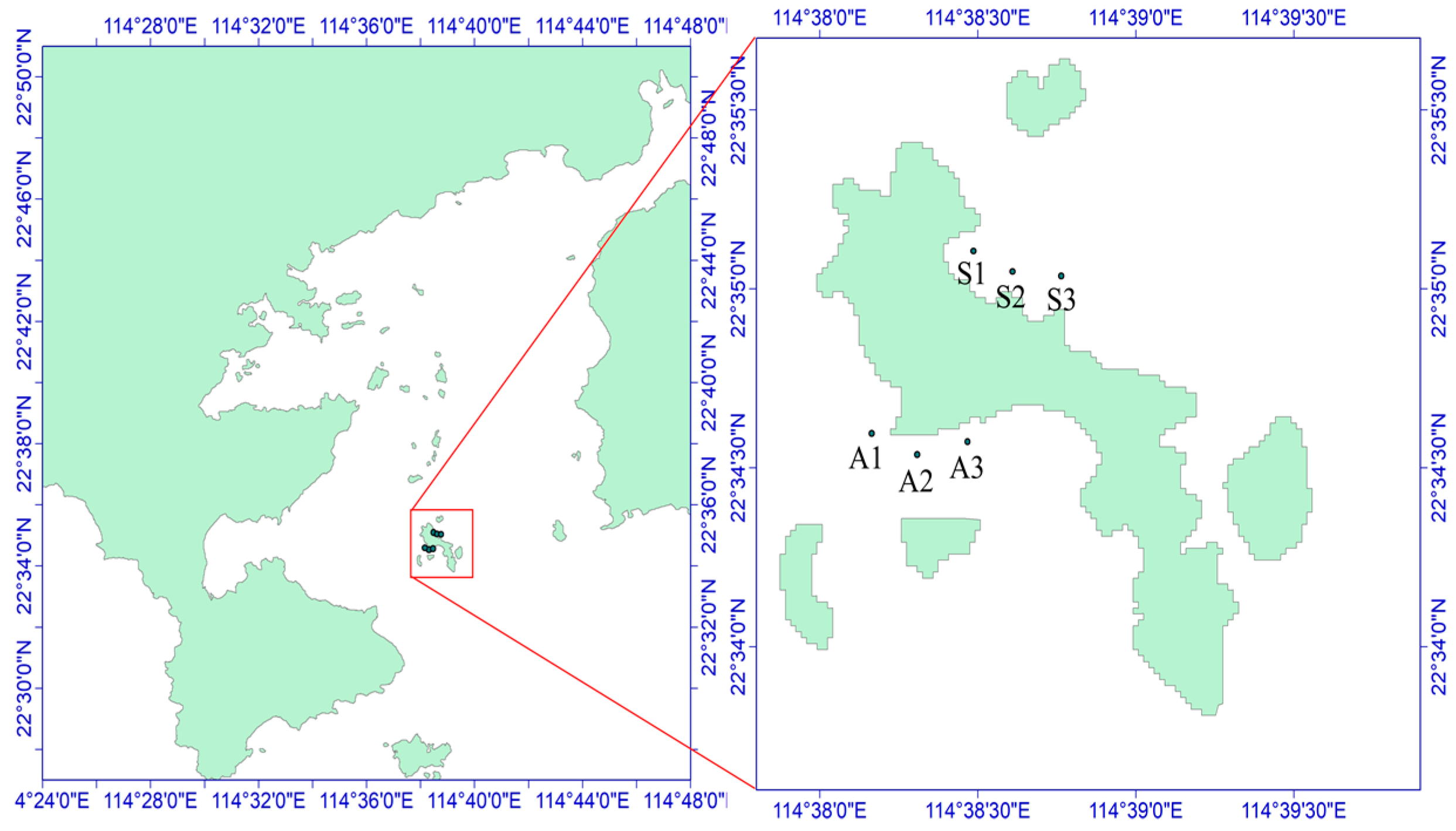
2.1. Sample Collection and Processing
2.2. Sample Preparation
2.3. Stable Isotope Analysis
2.4. High-Throughput Sequencing of 18S rDNA
2.5. 16S rDNA High-Throughput Sequencing
2.6. Statistical Analysis
3. Results and Analysis
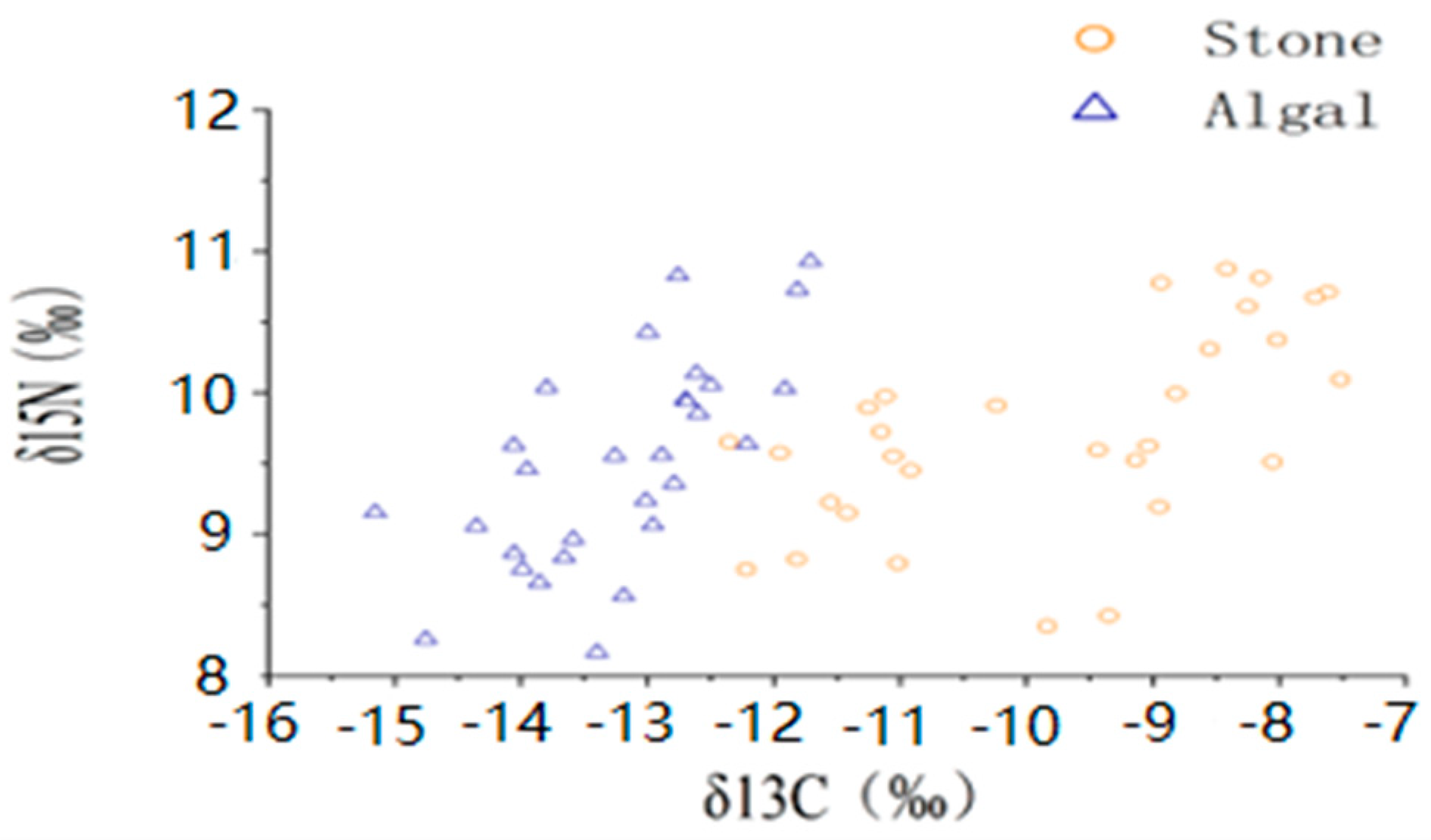
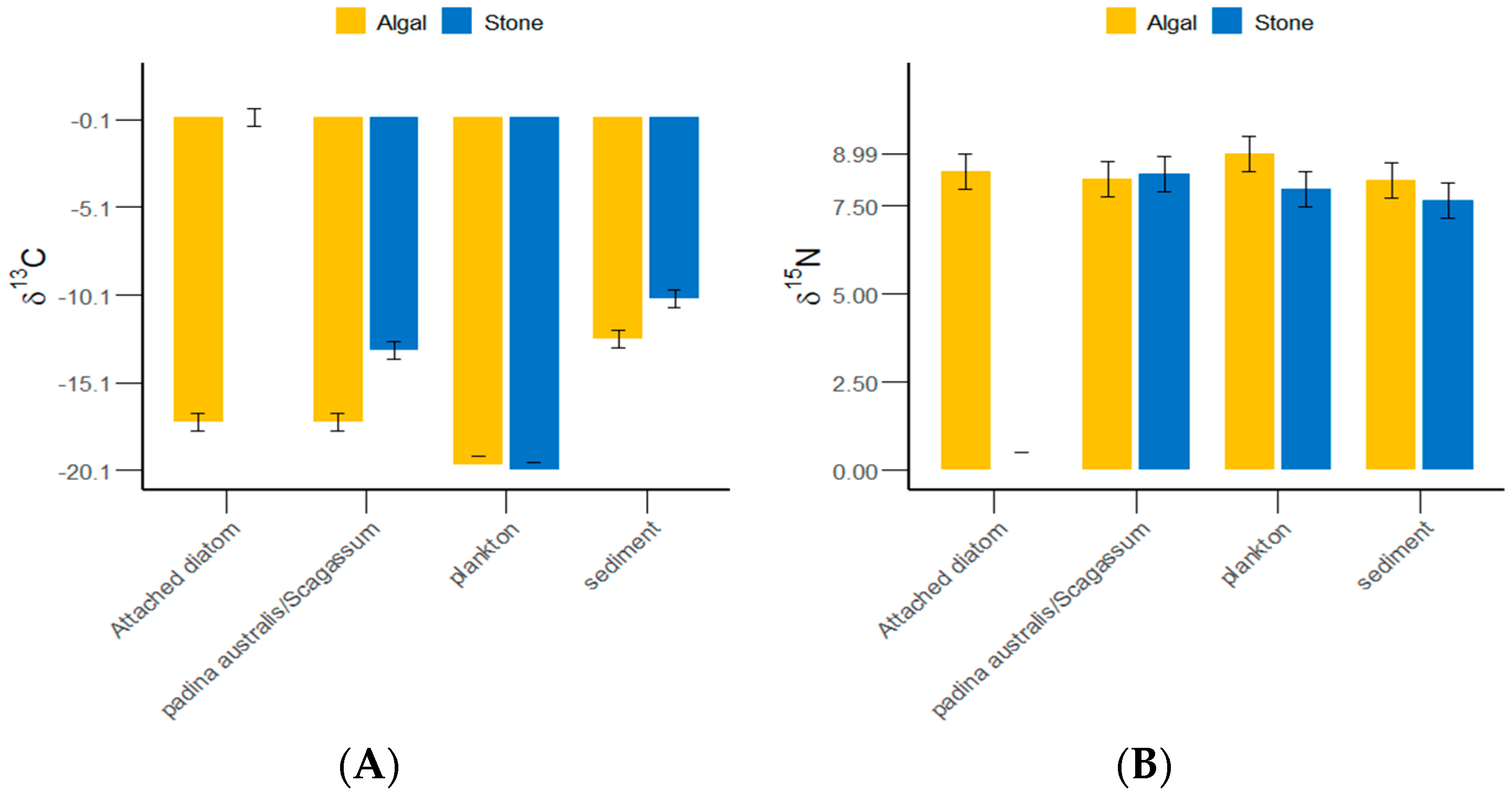
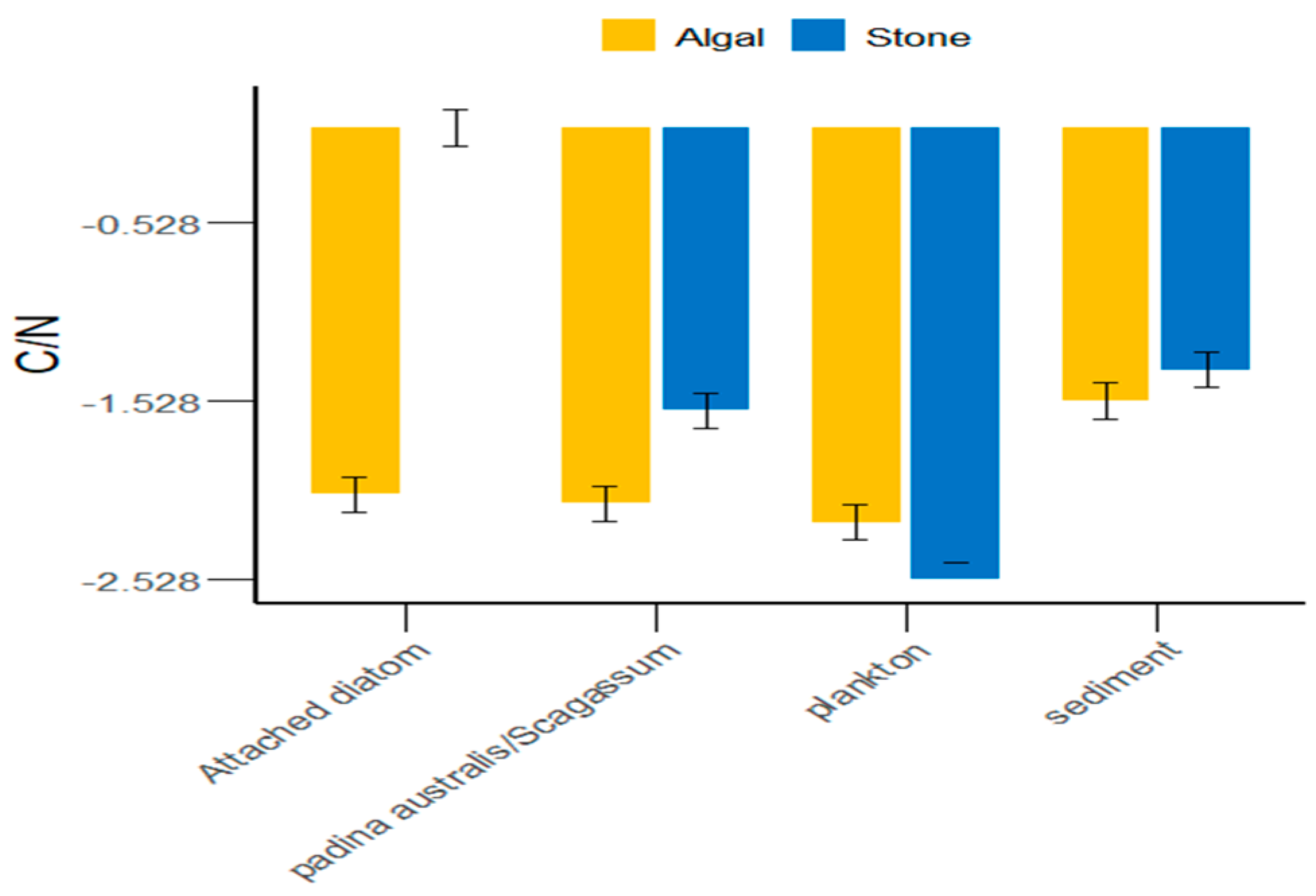
3.1. δ13C and δ15N Values and C/N Ratios of Potential Food Sources
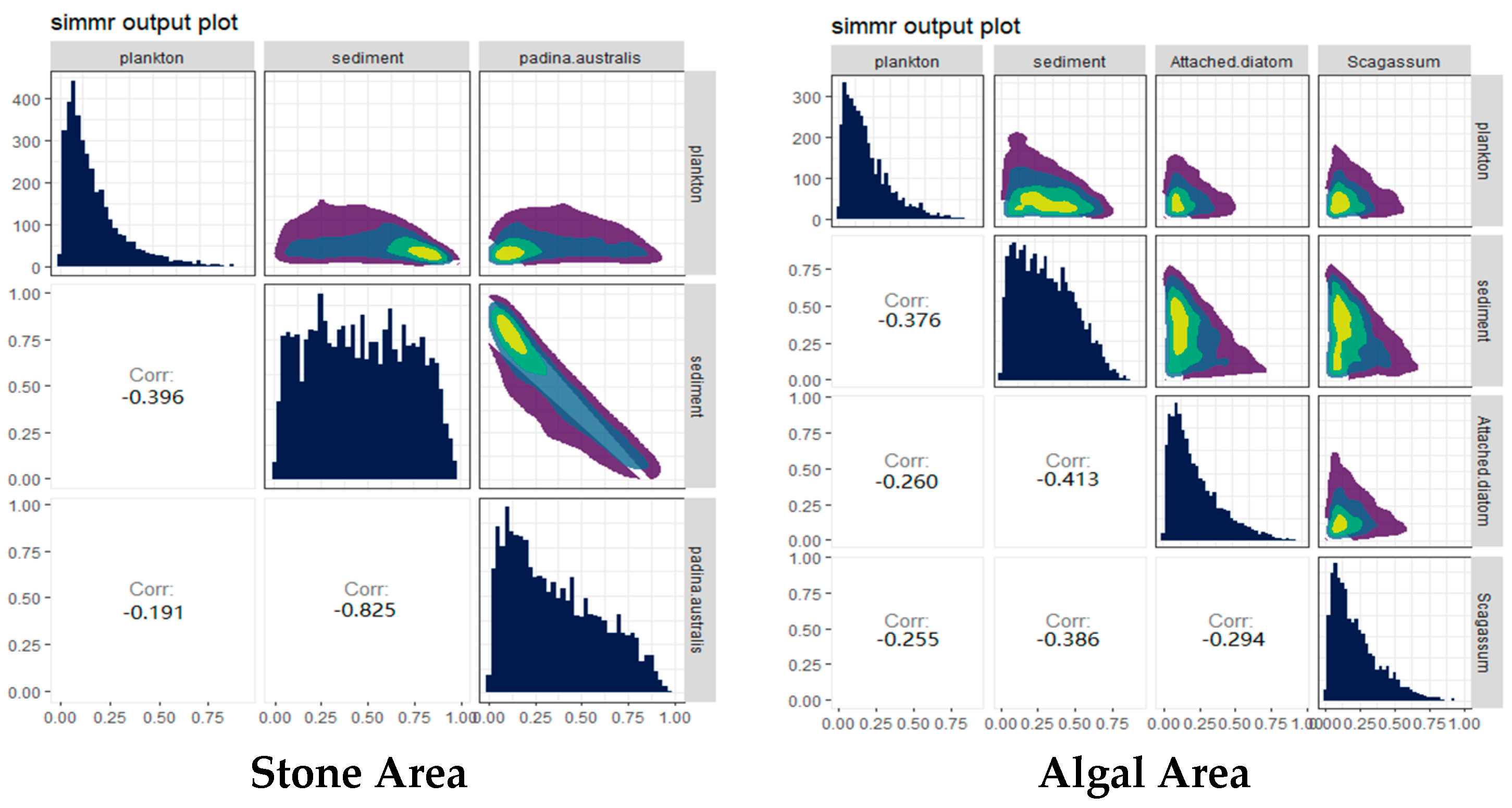
3.2. Stable-Isotope-Ratio-Based Analysis of the Diet of the Purple Sea Urchin
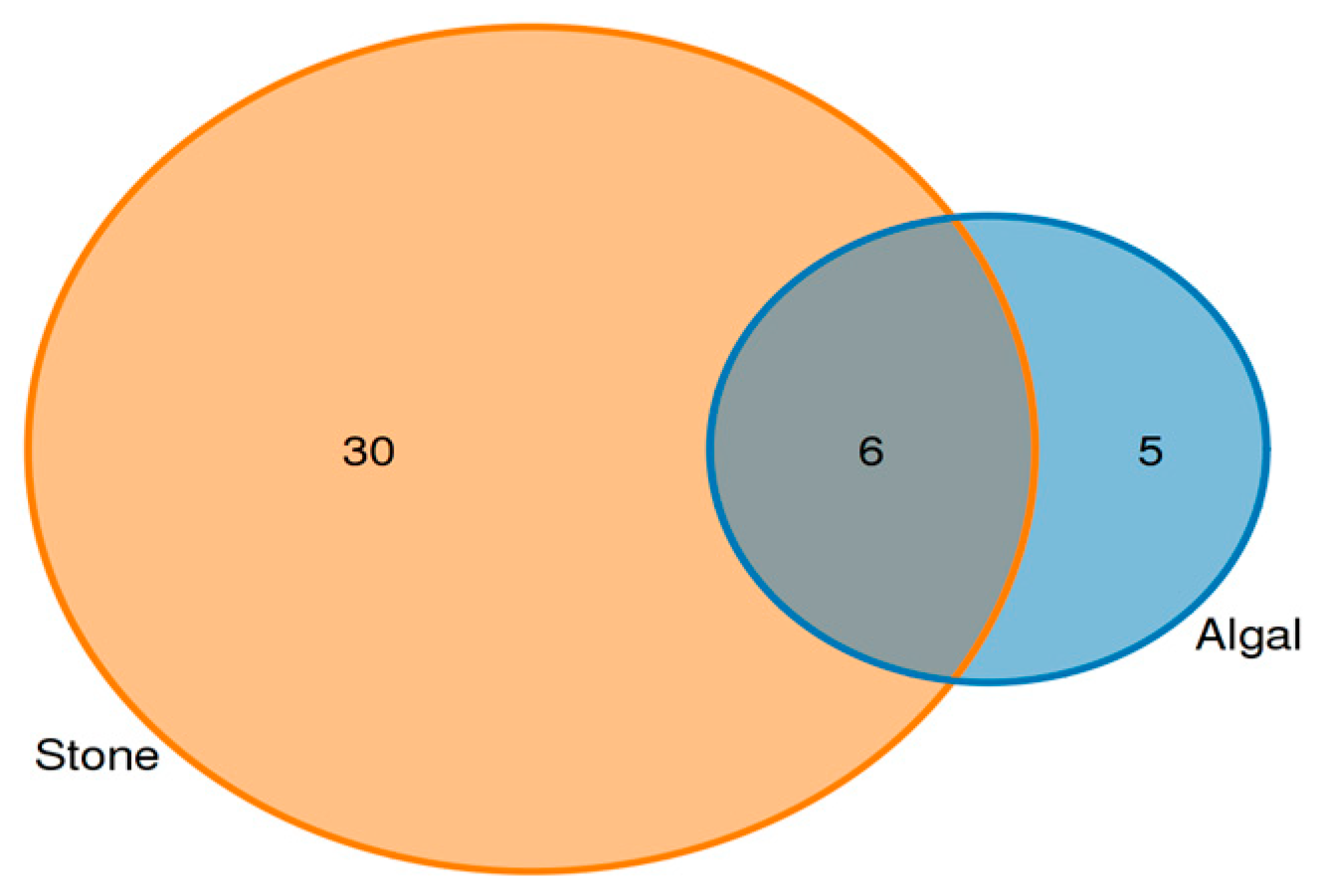
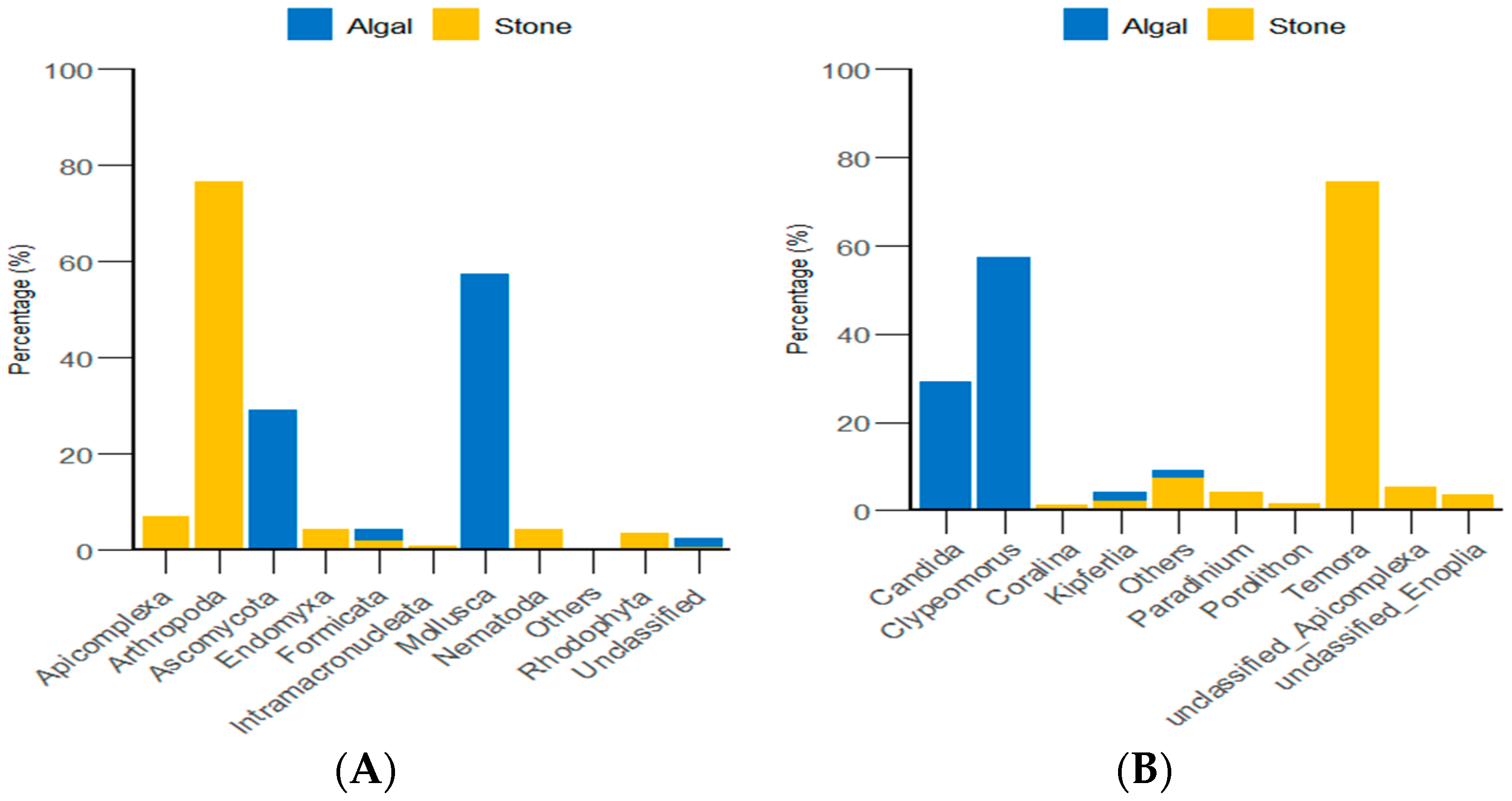
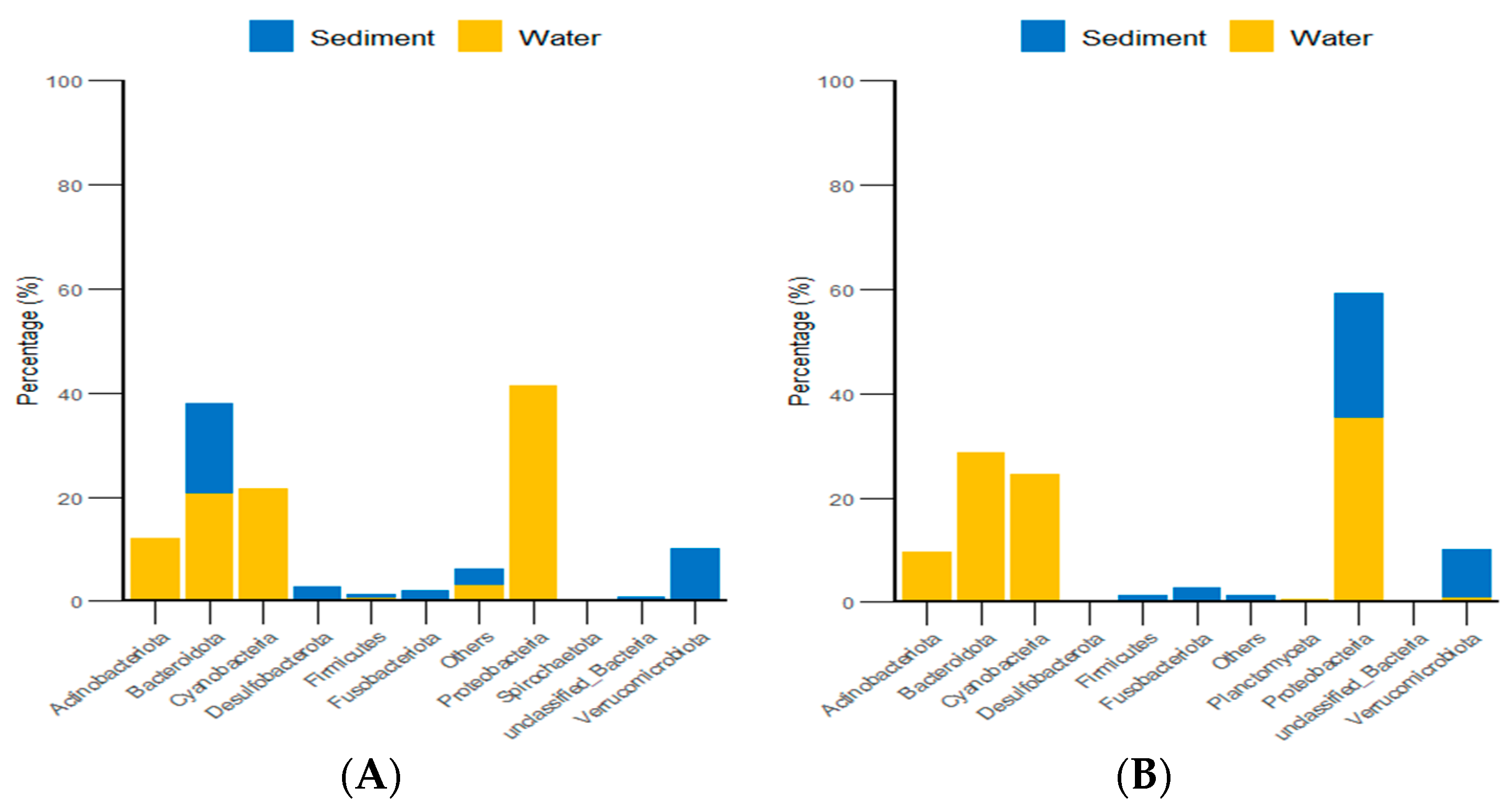
3.3. Analysis of the Diet Composition and Relative Abundance of Purple Sea Urchins Based on 18S
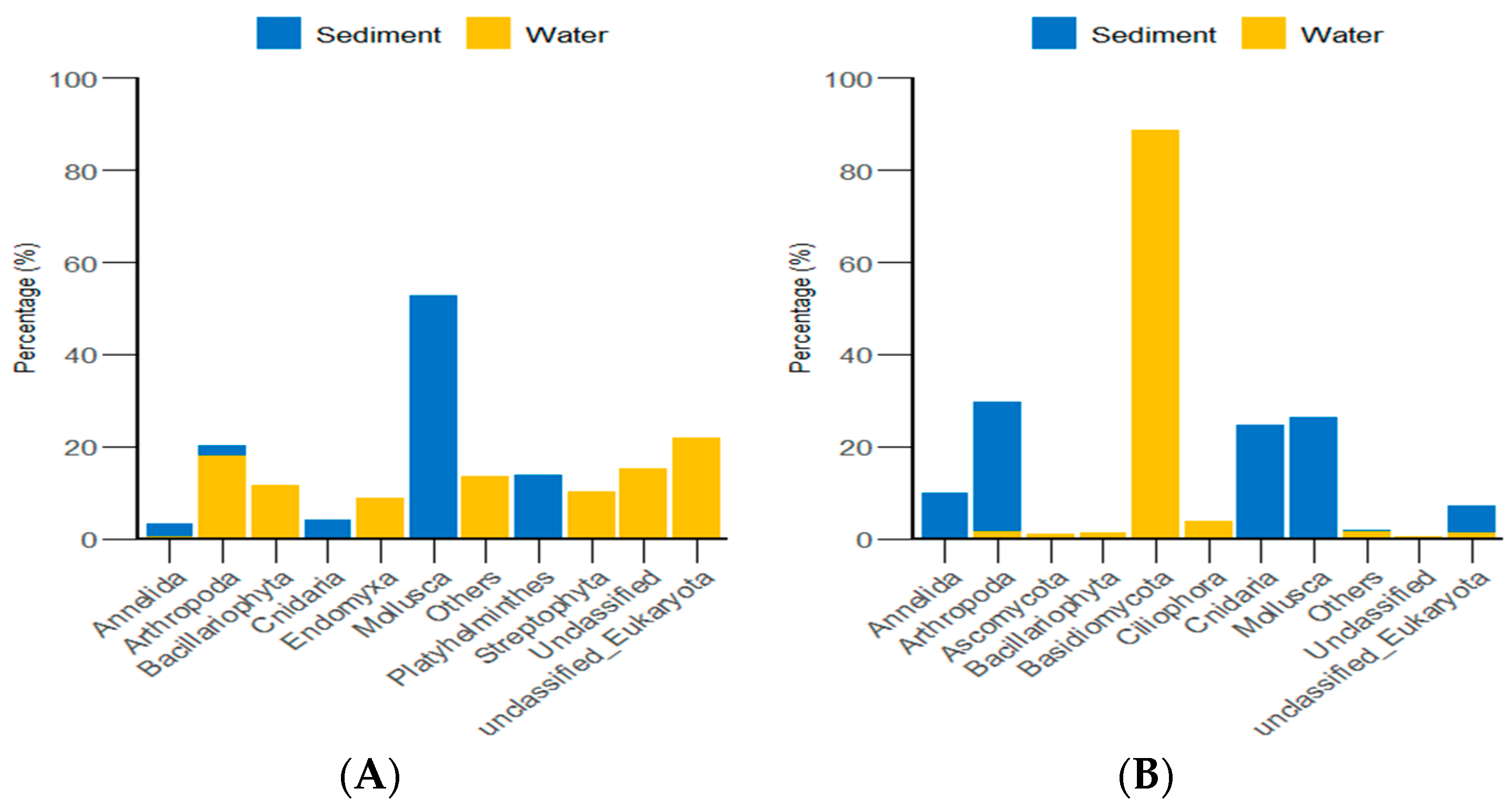
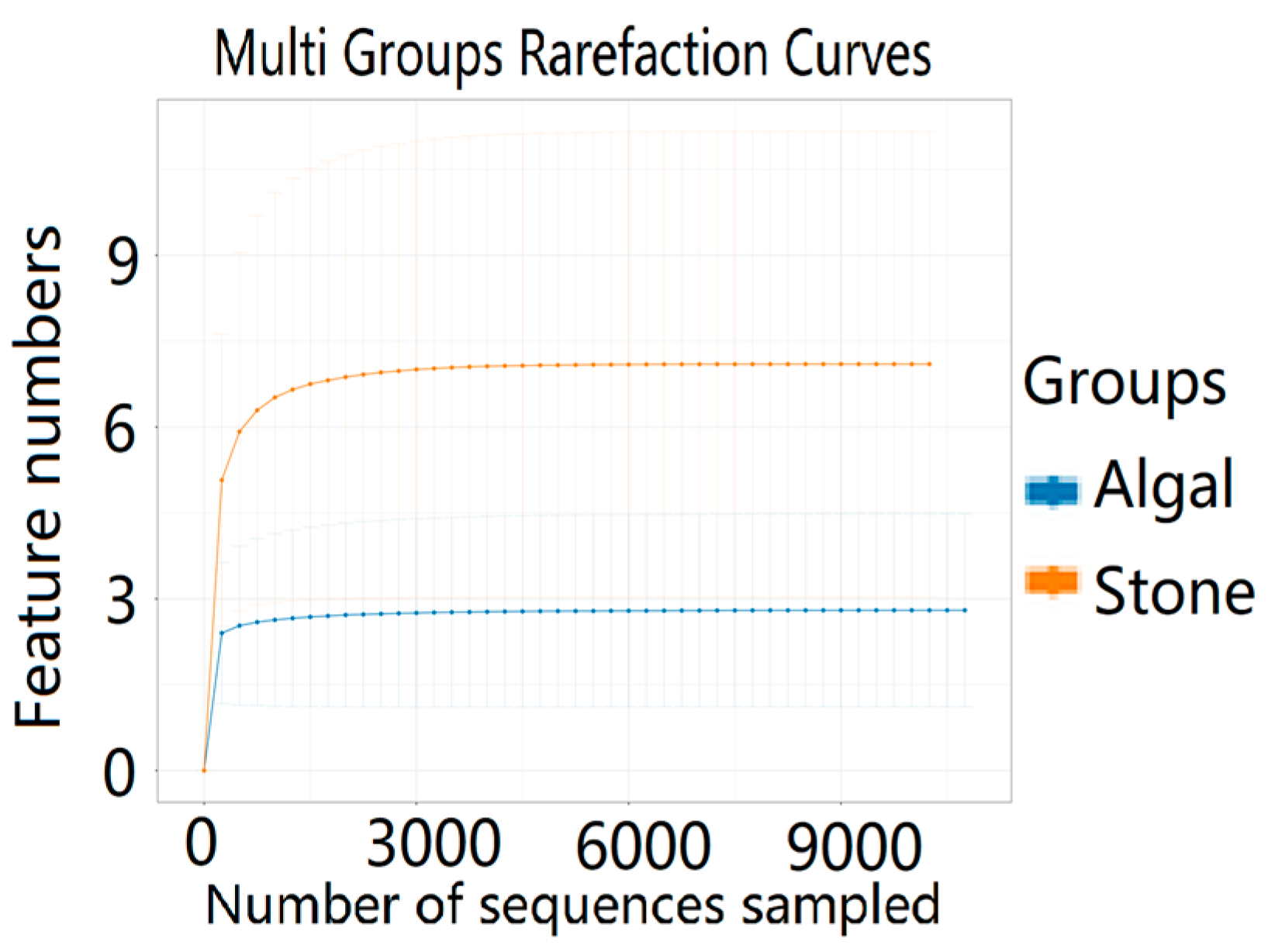
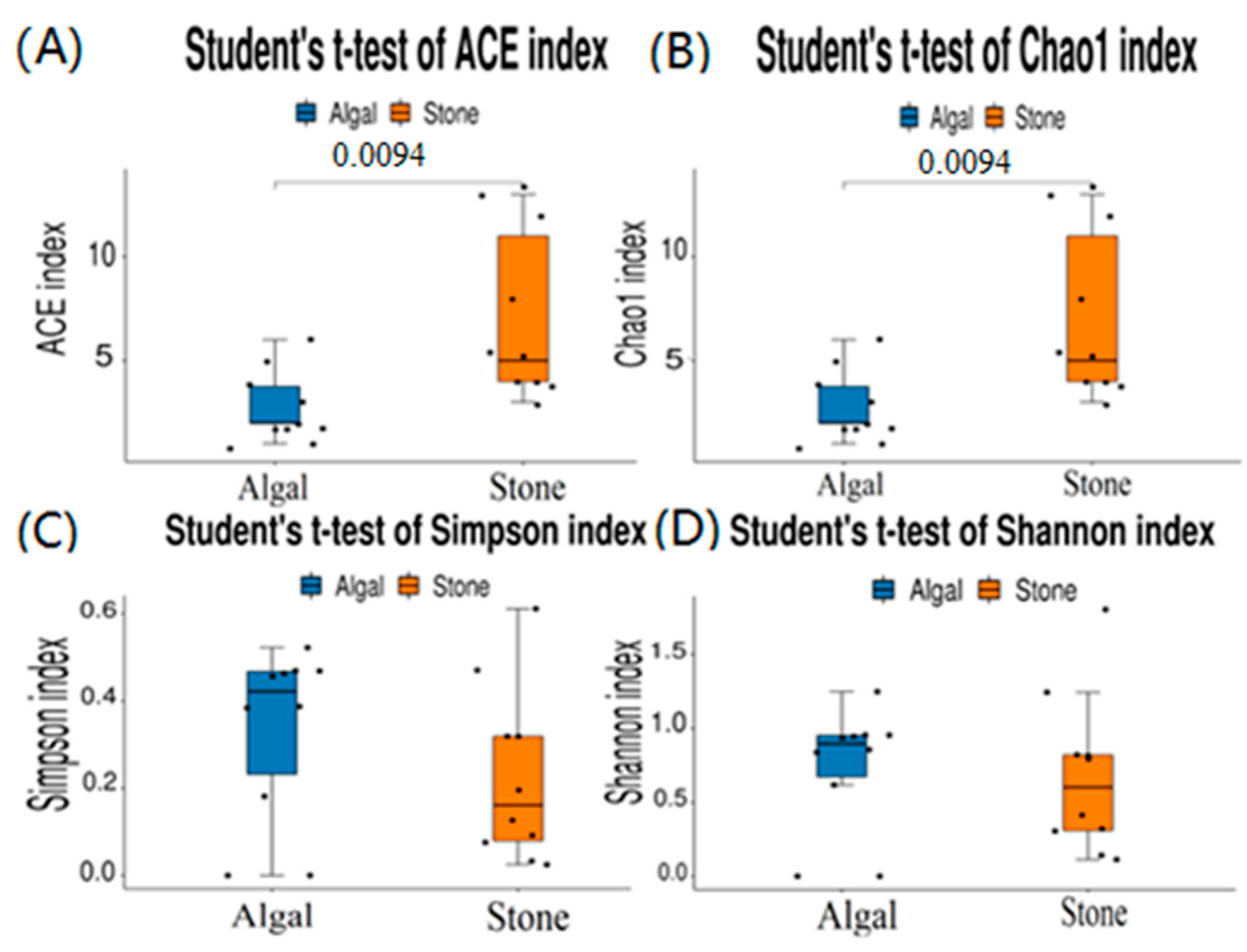
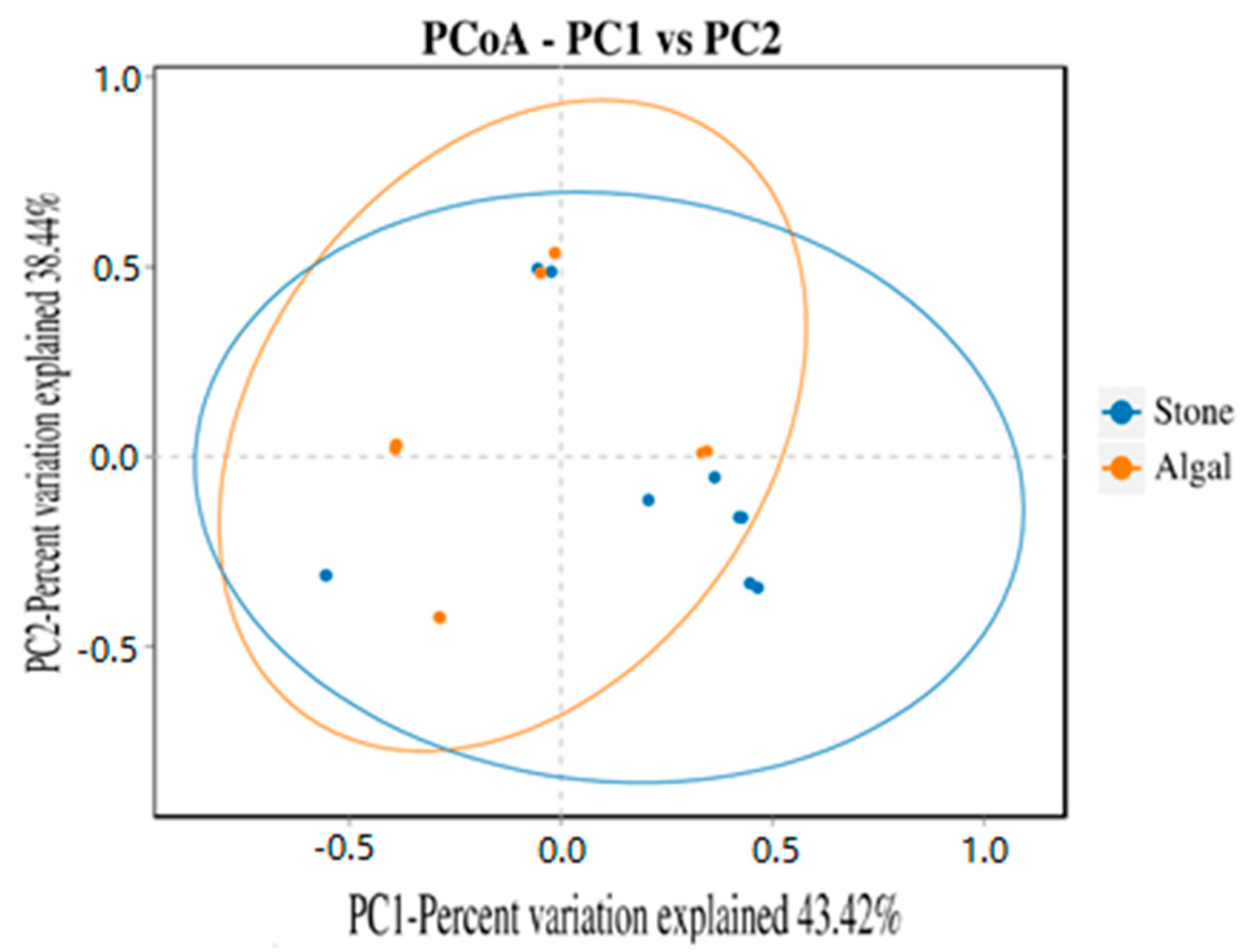
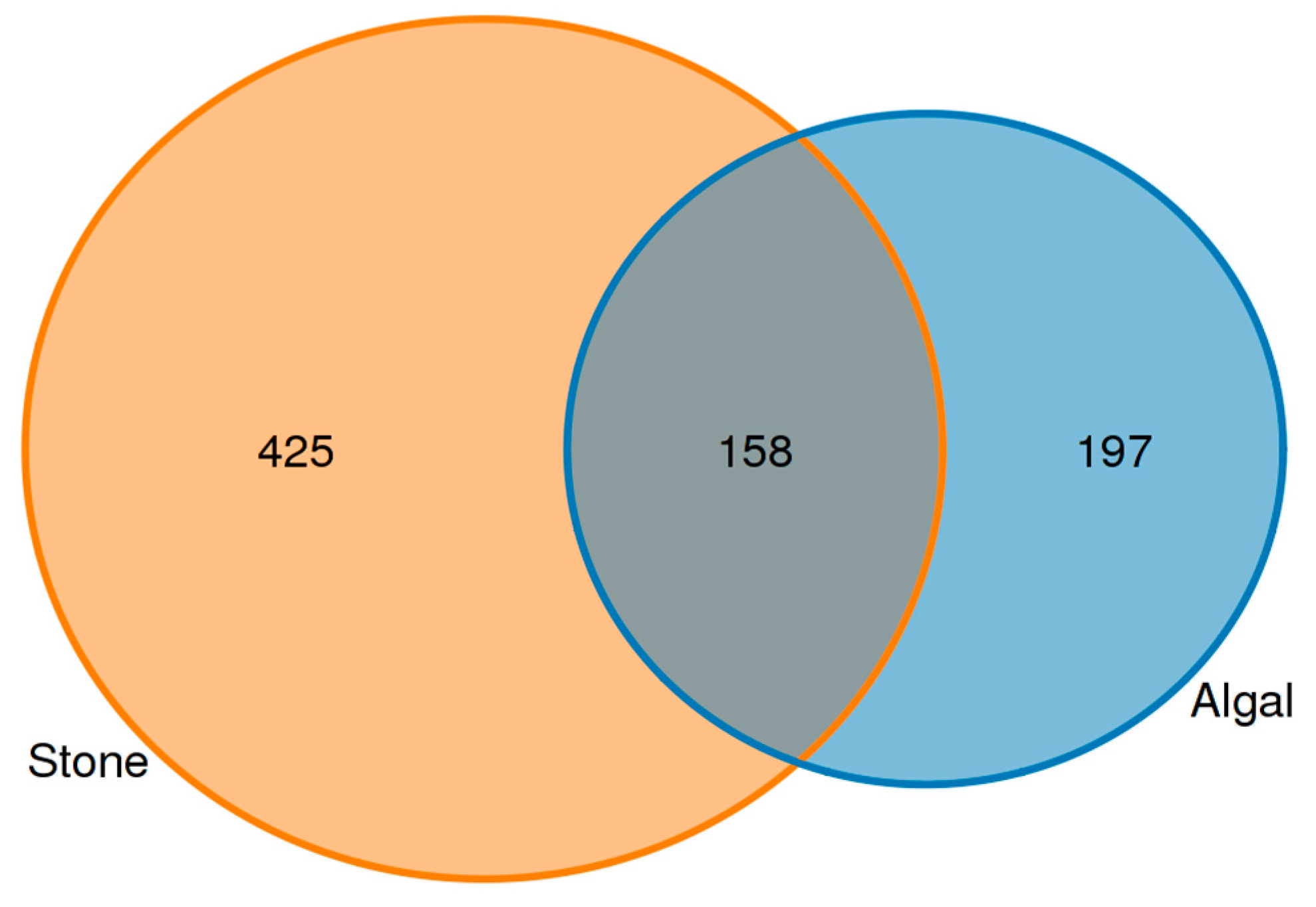
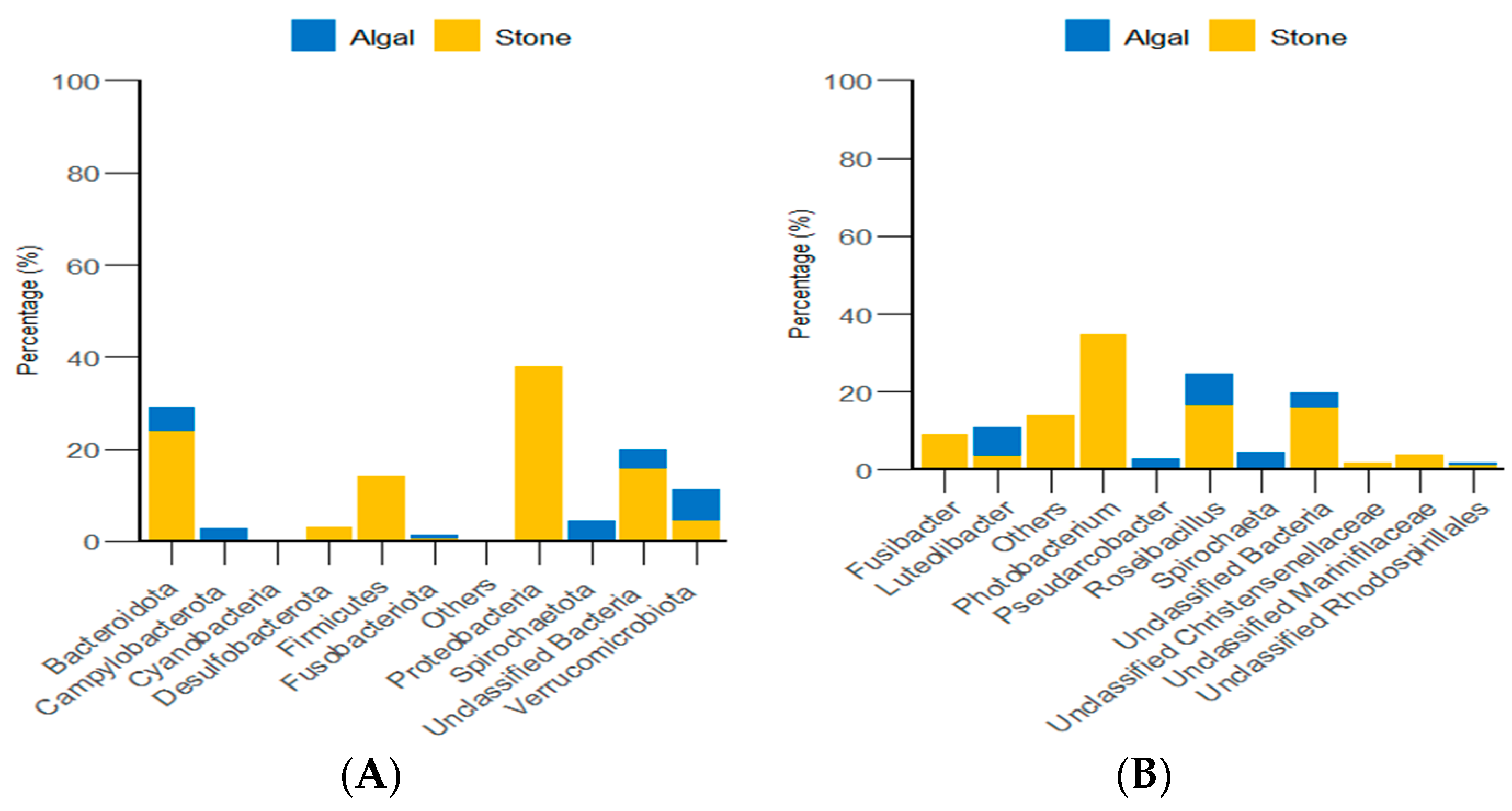
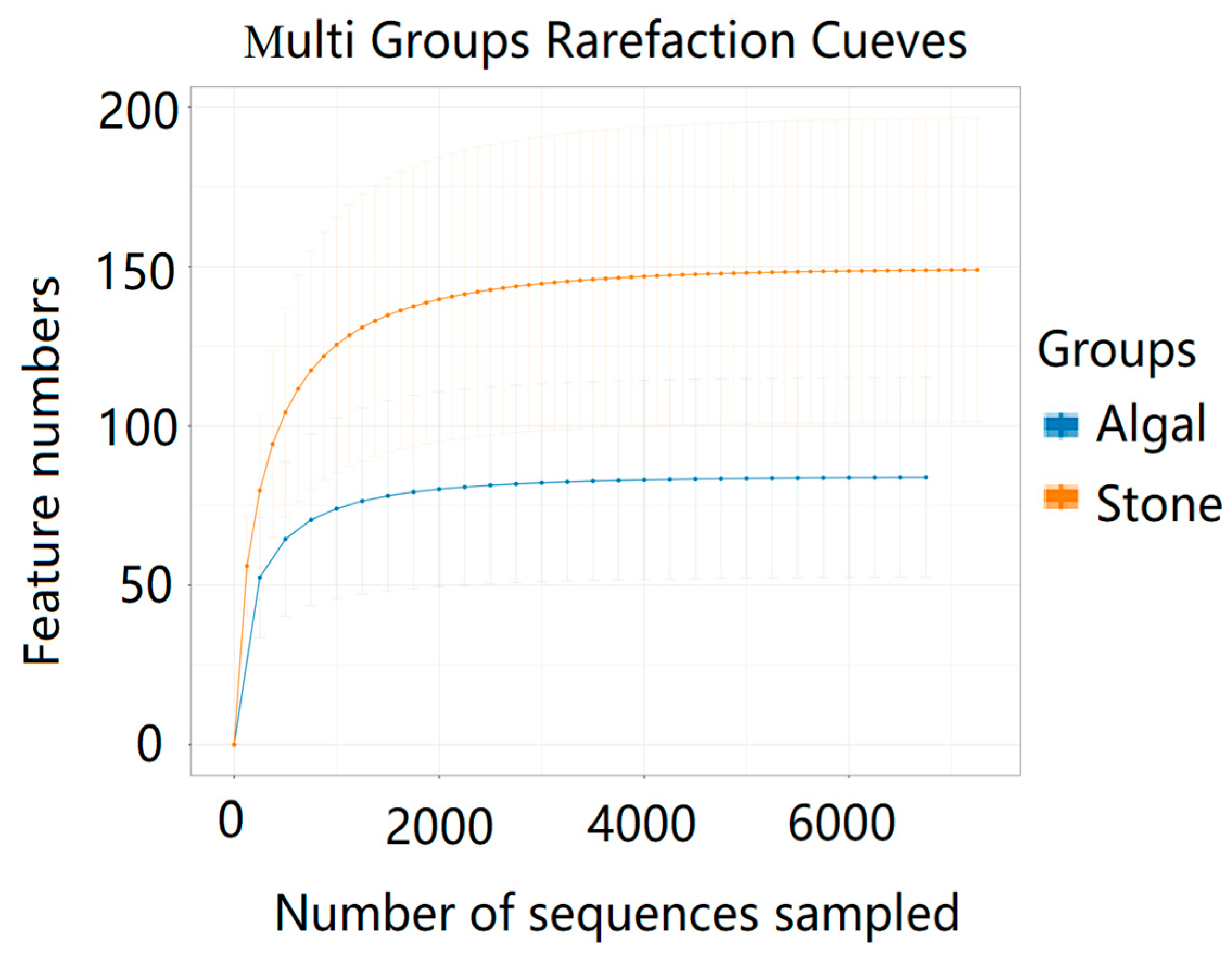
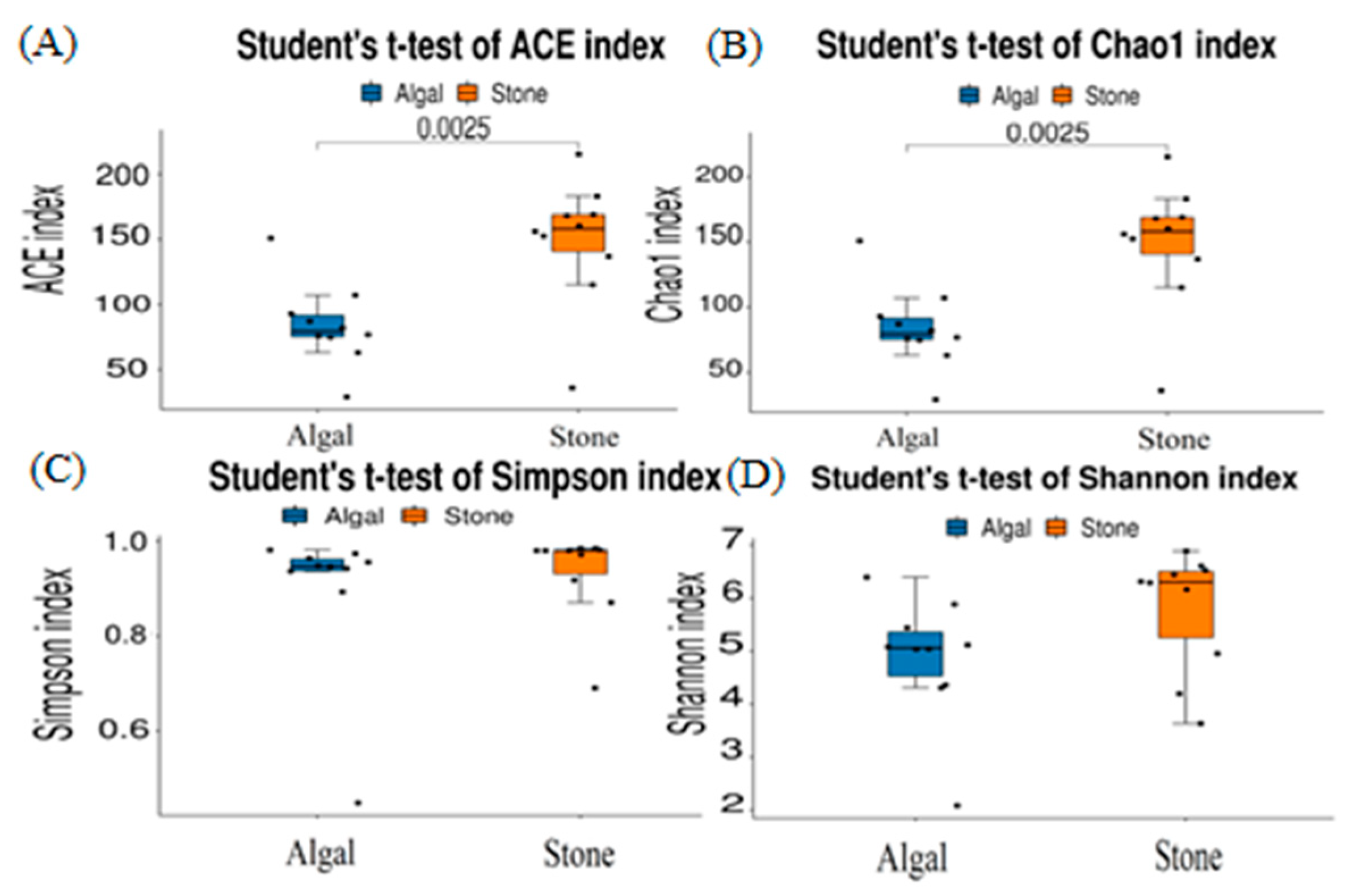
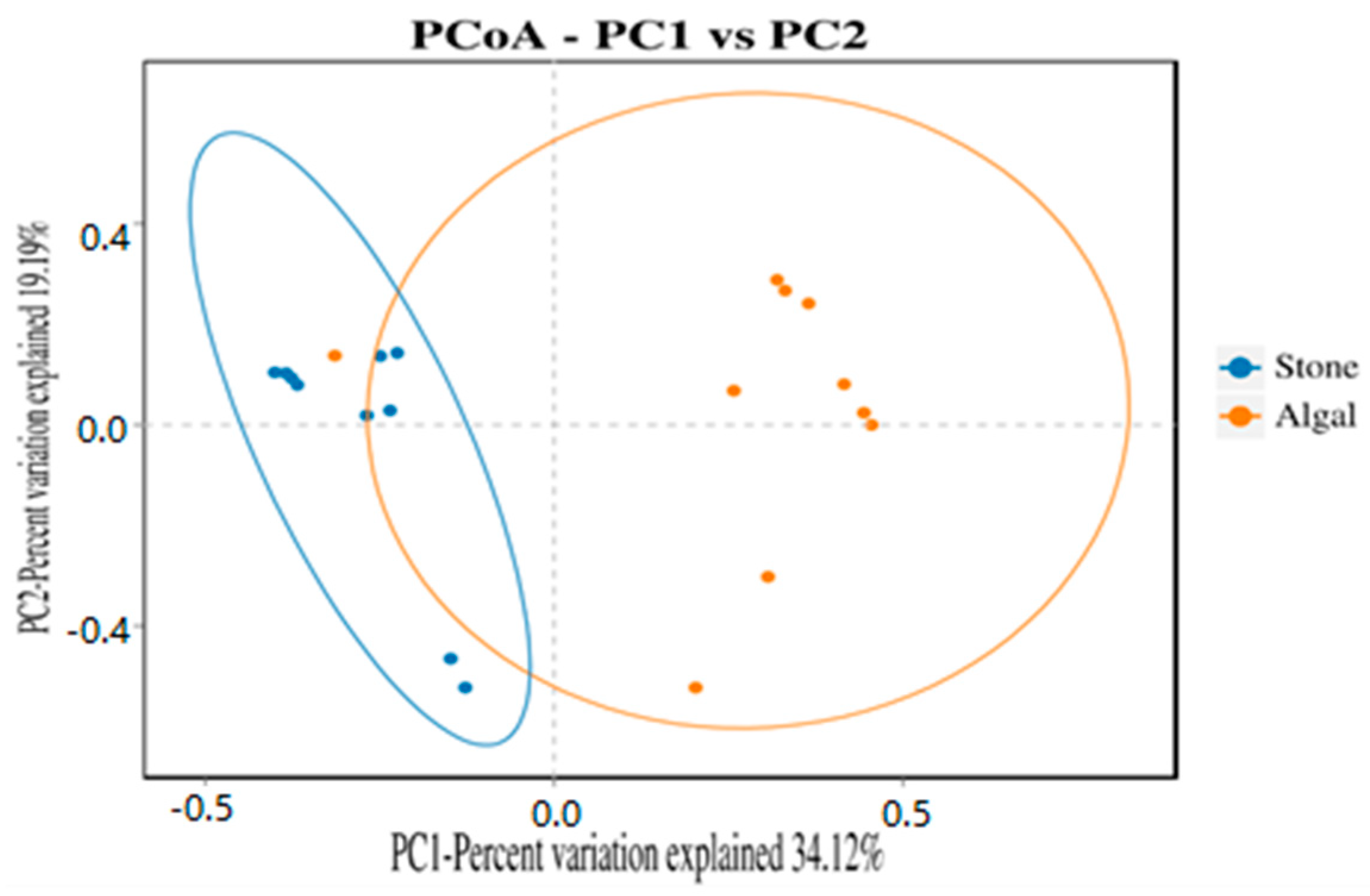
3.4. Diversity and Differential Analysis of the Sea Urchin Gut Microbiome
4. Discussion
4.1. Combining Stable Isotope Analysis with High-Throughput Sequencing Technology Can Provide a More Comprehensive Understanding of an Animal’s Feeding Habits
4.2. Habitat Environmental Characteristics Drive Changes in the Gut Food Composition of Purple Sea Urchins
4.3. Diet and Environment Can Influence the Composition and Diversity of the Gut Microbiota in Sea Urchins by Inducing Changes in Their Feeding Preferences
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, M.; Li, C. Current Progress on Identification of Virus Pathogens and the Antiviral Effectors in Echinoderms. Dev. Comp. Immunol. 2021, 116, 103912. [Google Scholar] [CrossRef] [PubMed]
- Uthicke, S.; Schaffelke, B.; Byrne, M. A Boom–Bust Phylum? Ecological and Evolutionary Consequences of Density Variations in Echinoderms. Ecol. Monogr. 2009, 79, 3–24. [Google Scholar] [CrossRef]
- Rodríguez-Barreras, R.; Godoy-Vitorino, F.; Præbel, K.; Wangensteen, O.S. DNA Metabarcoding Unveils Niche Overlapping and Competition among Caribbean Sea Urchins. Reg. Stud. Mar. Sci. 2020, 40, 101537. [Google Scholar] [CrossRef]
- Kelly, S.R.; Hamel, K.; Narvaez, C.A.; Armstrong, T.J.; Grace, S.P.; Feehan, C.J. Sea Urchin Arbacia Punctulata Feeding Preference for Algal Turf over Kelp in a Degraded Kelp Forest ecosystem. J. Exp. Mar. Biol. Ecol. 2024, 571, 151976. [Google Scholar] [CrossRef]
- Pulgar, J.; Moya, A.; Fernández, M.; Varas, O.; Guzmán-Rivas, F.; Urzúa, Á.; Quijón, P.A.; García-Huidobro, M.R.; Aldana, M.; Duarte, C. Upwelling Enhances Seaweed Nutrient Quality, Altering Feeding Behavior and Growth Rates in an Intertidal Sea Urchin, Loxechinus Albus. Sci. Total Environ. 2022, 851, 158307. [Google Scholar] [CrossRef]
- Yiu, S.K.F.; Chung, S.S.-W. Spatial Distribution and Habitat Relationship of Sea Urchin Assemblages (Echinodermata: Echinoidea) in Hong Kong Waters. Cont. Shelf Res. 2024, 273, 105170. [Google Scholar] [CrossRef]
- Ross, D.A.N.; Hamel, J.-F.; Mercier, A. Bathymetric and Interspecific Variability in Maternal Reproductive Investment and Diet of Eurybathic Echinoderms. Deep Sea Res. Part II Top. Stud. Oceanogr. 2013, 94, 333–342. [Google Scholar] [CrossRef]
- Mo, B.; Qin, C.; Chen, C.; Li, X.; Feng, X.; Tong, F.; Yuan, H. Dietary analysis of purple sea urchin in Daya Bay based on carbon and nitrogen stable isotope techniques. J. Fish. Sci. China 2017, 24, 566–575. [Google Scholar]
- Lau, D.C.C.; Dumont, C.P.; Lui, G.C.S.; Qiu, J.-W. Effectiveness of a Small Marine Reserve in Southern China in Protecting the Harvested Sea Urchin Anthocidaris Crassispina: A Mark-and-Recapture Study. Biol. Conserv. 2011, 144, 2674–2683. [Google Scholar] [CrossRef]
- Leung, P.T.Y.; Yan, M.; Yiu, S.K.F.; Lam, V.T.T.; Ip, J.C.H.; Au, M.W.Y.; Chen, C.-Y.; Wai, T.-C.; Lam, P.K.S. Molecular Phylogeny and Toxicity of Harmful Benthic Dinoflagellates Coolia (Ostreopsidaceae, Dinophyceae) in a Sub-Tropical Marine Ecosystem: The First Record from Hong Kong. Mar. Pollut. Bull. 2017, 124, 878–889. [Google Scholar] [CrossRef]
- Chen, G.; Xiang, W.-Z.; Lau, C.-C.; Peng, J.; Qiu, J.-W.; Chen, F.; Jiang, Y. A Comparative Analysis of Lipid and Carotenoid Composition of the Gonads of Anthocidaris Crassispina, Diadema Setosum and Salmacis Sphaeroides. Food Chem. 2010, 120, 973–977. [Google Scholar] [CrossRef]
- Dincer, T.; Cakli, S. Chemical Composition and Biometrical Measurements of the Turkish Sea Urchin (Paracentrotus Lividus, Lamarck, 1816). Crit. Rev. Food Sci. Nutr. 2007, 47, 21–26. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable Products from Biotechnology of Microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, S.; Mendes, S.; Raposo, A.; Santos, P.M.; Gomes, A.S.; Ganhão, R.; Costa, J.L.; Gil, M.M.; Pombo, A. Motivation and Preferences of Portuguese Consumers’ towards Sea Urchin Roe. Int. J. Gastron. Food Sci. 2021, 24, 100312. [Google Scholar] [CrossRef]
- Kanamoto, Y.; Nakamura, K. Age and Growth Analyses of the Purple Sea Urchin Heliocidaris Crassispina Inhabiting Different Feeding Environments in the Shimane Peninsula, Japan. Reg. Stud. Mar. Sci. 2023, 65, 103096. [Google Scholar] [CrossRef]
- Rocha, A.C.; Ressurreição, M.; Baeta, A.; Veríssimo, H.; Camarão, B.; Fernández-Boo, S.; Pombo, A.; Lourenço, S.; Gomes, A.S.; Santos, P.M.; et al. Temporal and Spatial Variability in the Isotopic Composition of Sea Urchins along Portuguese Coast. Mar. Environ. Res. 2023, 192, 106236. [Google Scholar] [CrossRef]
- Qin, C.; Chen, P.; Sarà, G.; Mo, B.; Zhang, A.; Li, X. Ecological Implications of Purple Sea Urchin (Heliocidaris crassispina, Agassiz, 1864) Enhancement on the Coastal Benthic Food Web: Evidence from Stable Isotope Analysis. Mar. Environ. Res. 2020, 158, 104957. [Google Scholar] [CrossRef]
- Magierecka, A.; Lind, Å.J.; Aristeidou, A.; Sloman, K.A.; Metcalfe, N.B. Chronic Exposure to Stressors Has a Persistent Effect on Feeding Behaviour but Not Cortisol Levels in Sticklebacks. Anim. Behav. 2021, 181, 71–81. [Google Scholar] [CrossRef]
- Cicala, D.; Sbrana, A.; Valente, T.; Berto, D.; Rampazzo, F.; Gravina, M.F.; Maiello, G.; Russo, T. Trophic Niche Overlap of Deep-Sea Fish Species Revealed by the Combined Approach of Stomach Contents and Stable Isotopes Analysis in the Central Tyrrhenian Sea. Deep Sea Res. Part Oceanogr. Res. Pap. 2024, 206, 104281. [Google Scholar] [CrossRef]
- Deniro, M.J.; Epstein, S. Influence of Diet on the Distribution of Nitrogen Isotopes in Animals. Geochim. Cosmochim. Acta 1981, 45, 341–351. [Google Scholar] [CrossRef]
- Boecklen, W.J.; Yarnes, C.T.; Cook, B.A.; James, A.C. On the Use of Stable Isotopes in Trophic Ecology. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 411–440. [Google Scholar] [CrossRef]
- McCormack, S.A.; Trebilco, R.; Melbourne-Thomas, J.; Blanchard, J.L.; Fulton, E.A.; Constable, A. Using Stable Isotope Data to Advance Marine Food Web Modelling. Rev. Fish Biol. Fish. 2019, 29, 277–296. [Google Scholar] [CrossRef]
- Layman, C.A.; Giery, S.T.; Buhler, S.; Rossi, R.; Penland, T.; Henson, M.N.; Bogdanoff, A.K.; Cove, M.V.; Irizarry, A.D.; Schalk, C.M.; et al. A Primer on the History of Food Web Ecology: Fundamental Contributions of Fourteen Researchers. Food Webs 2015, 4, 14–24. [Google Scholar] [CrossRef]
- Phillips, D.L.; Gregg, J.W. Source Partitioning Using Stable Isotopes: Coping with Too Many Sources. Oecologia 2003, 136, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.L.; Inger, R.; Bearhop, S.; Jackson, A.L.; Moore, J.W.; Parnell, A.C.; Semmens, B.X.; Ward, E.J. Best Practices for Use of Stable Isotope Mixing Models in Food-Web Studies. Can. J. Zool. 2014, 92, 823–835. [Google Scholar] [CrossRef]
- Nie, S.; Li, L.; Wu, Y.; Xiang, H.; Li, C.; Chen, S.; Zhao, Y.; Cen, J.; Yang, S.; Wang, Y. Exploring the Roles of Microorganisms and Metabolites in the Fermentation of Sea Bass (Lateolabrax japonicas) Based on High-Throughput Sequencing and Untargeted Metabolomics. LWT 2022, 167, 113795. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, Y.; Wu, Y.; Li, C.; Li, L.; Zhao, Y.; Hu, X.; Wei, Y.; Huang, H. Comparison of the Microbial Community and Flavor Compounds in Fermented Mandarin Fish (Siniperca chuatsi): Three Typical Types of Chinese Fermented Mandarin Fish Products. Food Res. Int. 2021, 144, 110365. [Google Scholar] [CrossRef]
- Shen, Y.; Wu, Y.; Wang, Y.; Li, L.; Li, C.; Zhao, Y.; Yang, S. Contribution of Autochthonous Microbiota Succession to Flavor Formation during Chinese Fermented Mandarin Fish (Siniperca chuatsi). Food Chem. 2021, 348, 129107. [Google Scholar] [CrossRef]
- Zhang, C.; Hu, S.; Lin, X.; Zhou, T.; Huang, H.; Liu, S. Diet and trophic level analysis of triggerfish (Balistapus undulatuse) in coral reefs of Nansha. J. Trop. Oceanogr. 2022, 41, 7–14. [Google Scholar]
- Lin, X.; Zhou, Y.; Lin, H.; Hu, S.; Huang, H.; Zhang, L.; Liu, S. Diet analysis of the parrotfish (Scarus globiceps) in coral reefs of the Nansha islands. J. Trop. Oceanogr. 2024, 43, 100–108. [Google Scholar]
- Pan, W.; Qin, C.; Zuo, T.; Yu, G.; Zhu, W.; Ma, H.; Xi, S. Is Metagenomic Analysis an Effective Way to Analyze Fish Feeding Habits? A Case of the Yellowfin Sea Bream Acanthopagrus Latus (Houttuyn) in Daya Bay. Front. Mar. Sci. 2021, 8, 634651. [Google Scholar] [CrossRef]
- Chai, J.; Zhuang, Y.; Cui, K.; Bi, Y.; Zhang, N. Metagenomics Reveals the Temporal Dynamics of the Rumen Resistome and Microbiome in Goat Kids. Microbiome 2024, 12, 14. [Google Scholar] [CrossRef]
- Kim, J.; Guk, J.-H.; Mun, S.-H.; An, J.-U.; Song, H.; Kim, J.; Ryu, S.; Jeon, B.; Cho, S. Metagenomic Analysis of Isolation Methods of a Targeted Microbe, Campylobacter Jejuni, from Chicken Feces with High Microbial Contamination. Microbiome 2019, 7, 67. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.; Feng, H.; Wang, C.; Tang, Y. Analysis of feeding habits of cultured jellyfish (Rhopilema esculentum) in Tongzhou Bay based on fatty acid and stable carbon and nitrogen isotopic analysis. South China Fish. Sci. 2021, 17, 25–31. [Google Scholar]
- Härer, A.; Torres-Dowdall, J.; Rometsch, S.J.; Yohannes, E.; Machado-Schiaffino, G.; Meyer, A. Parallel and Non-Parallel Changes of the Gut Microbiota during Trophic Diversification in Repeated Young Adaptive Radiations of Sympatric Cichlid Fish. Microbiome 2020, 8, 149. [Google Scholar] [CrossRef] [PubMed]
- Tissue-Specific Turnover Rates of the Nitrogen Stable Isotope as Functions of Time and Growth in a Cyprinid Fish|Hydrobiologia. Available online: https://link.springer.com/article/10.1007/s10750-017-3276-2 (accessed on 2 May 2024).
- Yin, Y.; Liu, K.; Li, G. Protective Effect of Prim-O-Glucosylcimifugin on Ulcerative Colitis and Its Mechanism. Front. Pharmacol. 2022, 13, 882924. [Google Scholar] [CrossRef]
- Chen, S.; Han, P.; Zhang, Q.; Liu, P.; Liu, J.; Zhao, L.; Guo, L.; Li, J. Lactobacillus Brevis Alleviates the Progress of Hepatocellular Carcinoma and Type 2 Diabetes in Mice Model via Interplay of Gut Microflora, Bile Acid and NOTCH 1 Signaling. Front. Immunol. 2023, 14, 1179014. [Google Scholar] [CrossRef] [PubMed]
- Hicks, A.L.; Lee, K.J.; Couto-Rodriguez, M.; Patel, J.; Sinha, R.; Guo, C.; Olson, S.H.; Seimon, A.; Seimon, T.A.; Ondzie, A.U.; et al. Gut Microbiomes of Wild Great Apes Fluctuate Seasonally in Response to Diet. Nat. Commun. 2018, 9, 1786. [Google Scholar] [CrossRef] [PubMed]
- Muegge, B.D.; Kuczynski, J.; Knights, D.; Clemente, J.C.; Gonzalez, A.; Fontana, L.; Henrissat, B.; Knight, R.; Gordon, J.I. Diet Drives Convergence in Gut Microbiome Functions across Mammalian Phylogeny and within Humans. Science 2011, 332, 970–974. [Google Scholar] [CrossRef]
- Yuan, S.; Wang, K.-S.; Meng, H.; Hou, X.-T.; Xue, J.-C.; Liu, B.-H.; Cheng, W.-W.; Li, J.; Zhang, H.-M.; Nan, J.-X.; et al. The Gut Microbes in Inflammatory Bowel Disease: Future Novel Target Option for Pharmacotherapy. Biomed. Pharmacother. 2023, 165, 114893. [Google Scholar] [CrossRef]
- Li, X.; Wu, P.; Zeng, X.; Lang, Q.; Lin, Y.; Huang, H.; Qian, P. Protocol for Correlation Analysis of the Murine Gut Microbiome and Meta-Metabolome Using 16S rDNA Sequencing and UPLC-MS. STAR Protoc. 2022, 3, 101494. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, J.; He, F. The Construction of Mycobacterium Tuberculosis 16S rDNA MSPQC Sensor Based on Exonuclease III-Assisted Cyclic Signal amplification. Biosens. Bioelectron. 2019, 138, 111322. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, E.; Nakamoto, J.A.; Cabrera-Sosa, L.; Huaihua, P.; Cruz, M.; Arévalo, J.; Milón, P.; Adaui, V. Novel CRISPR-Based Detection of Leishmania Species. Front. Microbiol. 2022, 13, 958693. [Google Scholar] [CrossRef] [PubMed]
- Hugerth, L.W.; Muller, E.E.L.; Hu, Y.O.O.; Lebrun, L.A.M.; Roume, H.; Lundin, D.; Wilmes, P.; Andersson, A.F. Systematic Design of 18S rRNA Gene Primers for Determining Eukaryotic Diversity in Microbial Consortia. PLoS ONE 2014, 9, e95567. [Google Scholar] [CrossRef]
- Cruz-Saavedra, L.; Ospina, C.; Patiño, L.H.; Villar, J.C.; Sáenz Pérez, L.D.; Cantillo-Barraza, O.; Jaimes-Dueñez, J.; Ballesteros, N.; Cáceres, T.; Vallejo, G.; et al. Enhancing Trypanosomatid Identification and Genotyping with Oxford Nanopore SequencingOxford Nanopore. J. Mol. Diagn. 2024, 26, 323–336. [Google Scholar] [CrossRef]
- Zhao, Y.-E.; Xu, J.-R.; Hu, L.; Wu, L.-P.; Wang, Z.-H. Complete Sequence Analysis of 18S rDNA Based on Genomic DNA Extraction from Individual Demodex Mites (Acari: Demodicidae). Exp. Parasitol. 2012, 131, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Makwanise, T.; Dube, S.; Sibula, M.S. Molecular Characterization of Raillietina Isolates from the Gastrointestinal Tract of Free Range Chickens (Gallus Gallus Domesticus) from the Southern Region of Zimbabwe Using the 18S rDNA gene. Vet. Parasitol. Reg. Stud. Rep. 2020, 20, 100389. [Google Scholar] [CrossRef]
- Lin, H.-J.; Kao, W.-Y.; Wang, Y.-T. Analyses of Stomach Contents and Stable Isotopes Reveal Food Sources of Estuarine Detritivorous Fish in Tropical/Subtropical Taiwan. Estuar. Coast. Shelf Sci. 2007, 73, 527–537. [Google Scholar] [CrossRef]
- Prado, P.; Carmichael, R.H.; Watts, S.A.; Cebrian, J.; Heck, K., Jr. Diet-Dependent δ13C and δ15N Fractionation among Sea Urchin Lytechinus Variegatus Tissues: Implications for Food Web Models. Mar. Ecol. Prog. Ser. 2012, 462, 175–190. [Google Scholar] [CrossRef]
- Yang, S.; Li, P.; Sun, K.; Wei, N.; Liu, J.; Feng, X. Mercury isotope compositions in seawater and marine fish revealed the sources and processes of mercury in the food web within differing marine compartments. Water Res. 2023, 241, 120150. [Google Scholar] [CrossRef]
- Prado, P.; Heck, K., Jr.; Watts, S.A.; Cebrian, J. δ13C and δ18O Signatures from Sea Urchin Skeleton: Importance of Diet Type in Metabolic Contributions. Mar. Ecol. Prog. Ser. 2013, 476, 153–166. [Google Scholar] [CrossRef]
- Macroalgae Contribution to the Diet of Two Sea Urchins in Sargassum Beds: Tripneustes Depressus (Camarodonta: Toxopneustidae) and Eucidaris Thouarsii (Cidaroide: Cidaridae). Reg. Stud. Mar. Sci. 2022, 53, 102456. [CrossRef]
- Hughes, T.P.; Rodrigues, M.J.; Bellwood, D.R.; Ceccarelli, D.; Hoegh-Guldberg, O.; McCook, L.; Moltschaniwskyj, N.; Pratchett, M.S.; Steneck, R.S.; Willis, B. Phase Shifts, Herbivory, and the Resilience of Coral Reefs to Climate Change. Curr. Biol. 2007, 17, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.R.; Anderson, R.J. Biological and Physical Factors Controlling the Spatial Distribution of the Intertidal Alga Gelidium Pristoides in the Eastern Cape, South Africa. J. Mar. Biol. Assoc. UK 1991, 71, 555–568. [Google Scholar] [CrossRef]
- Bulleri, F.; Thiault, L.; Mills, S.C.; Nugues, M.M.; Eckert, E.M.; Corno, G.; Claudet, J. Erect Macroalgae Influence Epilithic Bacterial Assemblages and Reduce Coral Recruitment. Mar. Ecol. Prog. Ser. 2018, 597, 65–77. [Google Scholar] [CrossRef]
- Vieira, C.; Engelen, A.H.; Guentas, L.; Aires, T.; Houlbreque, F.; Gaubert, J.; Serrão, E.A.; De Clerck, O.; Payri, C.E. Species Specificity of Bacteria Associated to the Brown Seaweeds Lobophora (Dictyotales, Phaeophyceae) and Their Potential for Induction of Rapid Coral Bleaching in Acropora Muricata. Front. Microbiol. 2016, 7, 316. [Google Scholar] [CrossRef] [PubMed]
- Nugues, M.M.; Smith, G.W.; Hooidonk, R.J.; Seabra, M.I.; Bak, R.P.M. Algal Contact as a Trigger for Coral Disease. Ecol. Lett. 2004, 7, 919–923. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, Q.; Huang, L.; Wang, J.; Lin, R.; Huang, D.; Sun, X. Feeding Habits for Two Dominant Amphipod Species in the Yundang Lagoon Based on Stable Carbon and Nitrogen Isotope Analysis. Acta Ecol. Sin. 2015, 35, 7589–7597. [Google Scholar]
- Lyons, D.A.; Scheibling, R.E. Effect of Dietary History and Algal Traits on Feeding Rate and Food Preference in the Green Sea Urchin Strongylocentrotus Droebachiensis. J. Exp. Mar. Biol. Ecol. 2007, 349, 194–204. [Google Scholar] [CrossRef]
- Trenzado, C.E.; Hidalgo, F.; Villanueva, D.; Furné, M.; Díaz-Casado, M.E.; Merino, R.; Sanz, A. Study of the Enzymatic Digestive Profile in Three Species of Mediterranean Sea Urchins. Aquaculture 2012, 344–349, 174–180. [Google Scholar] [CrossRef]
- Hernández-Almaraz, P.; Méndez-Rodríguez, L.; Zenteno-Savín, T.; O’Hara, T.M.; Harley, J.R.; Serviere-Zaragoza, E. Concentrations of Trace Elements in Sea Urchins and Macroalgae Commonly Present in Sargassum Beds: Implications for Trophic Transfer. Ecol. Res. 2016, 31, 785–798. [Google Scholar] [CrossRef]
- Lawrence, J.M.; Lawrence, A.L.; Watts, S.A. Chapter 9—Feeding, Digestion and Digestibility of Sea Urchins. In Developments in Aquaculture and Fisheries Science; Lawrence, J.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 38, pp. 135–154. ISBN 0167-9309. [Google Scholar]
- Giraldes, B.W.; Al-Thani, J.A.K.H.; Dib, S.; Engmann, A.; Alsaadi, H.A.; Vethamony, P.; Alatalo, J.M.; Yigiterhan, O. Target Gastropods for Standardizing the Monitoring of Tar Mat Contamination in the Arabian Gulf. Reg. Stud. Mar. Sci. 2022, 53, 102328. [Google Scholar] [CrossRef]
- Rotolo, F.; Roncalli, V.; Cieslak, M.; Gallo, A.; Buttino, I.; Carotenuto, Y. Transcriptomic Analysis Reveals Responses to a Polluted Sediment in the Mediterranean Copepod Acartia Clausi. Environ. Pollut. 2023, 335, 122284. [Google Scholar] [CrossRef] [PubMed]
- Scheibling, R.; Hennigar, A.; Balch, T. Destructive Grazing, Epiphytism, and Disease: The Dynamics of Sea Urchin—Kelp Interactions in Nova Scotia. Can. J. Fish. Aquat. Sci. 1999, 56, 2300–2314. [Google Scholar] [CrossRef]
- Westbrook, C.E.; Ringang, R.R.; Cantero, S.M.A.; Toonen, R.J. Survivorship and Feeding Preferences among Size Classes of Outplanted Sea Urchins, Tripneustes Gratilla, and Possible Use as Biocontrol for Invasive Alien Algae. PeerJ 2015, 3, e1235. [Google Scholar] [CrossRef]
- Conklin, E.J.; Smith, J.E. Abundance and Spread of the Invasive Red Algae, Kappaphycus Spp., in Kane’ohe Bay, Hawai’i and an Experimental Assessment of Management Options. Biol. Invasions 2005, 7, 1029–1039. [Google Scholar] [CrossRef]
- Mancuso, F.P.; Milazzo, M.; Sarà, G.; Chemello, R. Bi- and Three-Dimensional Fractal Analysis of the Brown Seaweed Gongolaria Montagnei and Their Relationship with Gastropod Molluscs Assemblage. Mar. Pollut. Bull. 2023, 186, 114396. [Google Scholar] [CrossRef]
- Mahadik, G.A.; Castellani, C.; Mazzocchi, M.G. Effect of Diatom Morphology on the Small-Scale Behavior of the Copepod Temora Stylifera (Dana, 1849). J. Exp. Mar. Biol. Ecol. 2017, 493, 41–48. [Google Scholar] [CrossRef]
- Wessels, H.; Hagen, W.; Molis, M.; Wiencke, C.; Karsten, U. Intra- and Interspecific Differences in Palatability of Arctic Macroalgae from Kongsfjorden (Spitsbergen) for Two Benthic Sympatric Invertebrates. J. Exp. Mar. Biol. Ecol. 2006, 329, 20–33. [Google Scholar] [CrossRef]
- Pavia, H.; Toth, G.B. Inducible Chemical Resistance to Herbivory in the Brown Seaweed Ascophyllum Nodosum. Ecology 2000, 81, 3212–3225. [Google Scholar] [CrossRef]
- Neighbors, M.A.; Horn, M.H. Nutritional Quality of Macrophytes Eaten and Not Eaten by Two Temperatezone Herbivorous Fishes: A Multivariate Comparison. Mar. Biol. 1991, 108, 471–476. [Google Scholar] [CrossRef]
- Wright, J.T.; Dworjanyn, S.A.; Rogers, C.N.; Steinberg, P.D.; Williamson, J.E.; Poore, A.G.B. Density-Dependent Sea Urchin Grazing: Differential Removal of Species, Changes in Community Composition and Alternative Community States. Mar. Ecol. Prog. Ser. 2005, 298, 143–156. [Google Scholar] [CrossRef]
- Sala, E.; Boudouresque, C.F.; Harmelin-Vivien, M. Fishing, Trophic Cascades, and the Structure of Algal Assemblages: Evaluation of an Old but Untested Paradigm. Oikos 1998, 82, 425–439. [Google Scholar] [CrossRef]
- Pinnegar, J.; Polunin, N.; Francour, P.; Badalamenti, F.; Chemello, R.; Harmelin-vivien, M.; Pipitone, C. Trophic cascades in benthic marine ecosystems: Lessons for fisheries and protected-area management. Environ. Conserv. 2000, 27, 179–200. [Google Scholar] [CrossRef]
- Liao, Y.; Cai, C.; Yang, C.; Zheng, Z.; Wang, Q.; Du, X.; Deng, Y. Effect of Protein Sources in Formulated Diets on the Growth, Immune Response, and Intestinal Microflora of Pearl Oyster Pinctada Fucata Martensii. Aquac. Rep. 2020, 16, 100253. [Google Scholar] [CrossRef]
- Wang, C.; Li, P.; Guo, L.; Cao, H.; Mo, W.; Xin, Y.; Jv, R.; Zhao, Y.; Liu, X.; Ma, C.; et al. A New Potential Risk: The Impacts of Klebsiella Pneumoniae Infection on the Histopathology, Transcriptome and Metagenome of Chinese Mitten Crab (Eriocheir Sinensis). Fish Shellfish Immunol. 2022, 131, 918–928. [Google Scholar] [CrossRef] [PubMed]
- Sears, C.L. A Dynamic Partnership: Celebrating Our Gut Flora. Anaerobe 2005, 11, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Wu, Y.; Ye, Q.; Yao, Y.; Li, L.; Guo, Z.; Yang, L.; Tian, W.; Jiang, J. Individual and Combined Effects of Microplastics and Cadmium on Intestinal Histology and Microflora of Procypris Merus. Aquac. Rep. 2023, 31, 101659. [Google Scholar] [CrossRef]
- Klase, G.; Lee, S.; Liang, S.; Kim, J.; Zo, Y.-G.; Lee, J. The Microbiome and Antibiotic Resistance in Integrated Fishfarm Water: Implications of Environmental Public Health. Sci. Total Environ. 2019, 649, 1491–1501. [Google Scholar] [CrossRef]
- Gil, D.G.; Boraso, A.L.; Lopretto, E.C.; Zaixso, H.E. Depth-Related Plasticity in the Diet Composition of Pseudechinus Magellanicus (Echinoidea, Temnopleuridae) in Nearshore Environments off Central Patagonia, Argentina. Aquat. Ecol. 2021, 55, 589–606. [Google Scholar] [CrossRef]
- Wu, W.-J.; Liu, Q.-Q.; Chen, G.-J.; Du, Z.-J. Roseimarinus Sediminis Gen. Nov., Sp. Nov., a Facultatively Anaerobic Bacterium Isolated from Coastal Sediment. Int. J. Syst. Evol. Microbiol. 2015, 65, 2260–2264. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Jiang, C.; Shang, S.; Wang, S.; Zhu, K.; Dong, X.; Zhou, D.; Jiang, P. Insight into the Relationship between Metabolite Dynamic Changes and Microorganisms of Sea Urchin (S. intermedius) Gonads during Storage. Food Chem. X 2023, 18, 100727. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Liu, Y.; Zhuang, S.; Sun, X.; Li, Y.; Hong, H.; Lv, Y.; Luo, Y. Effect of ε-Polylysine and Ice Storage on Microbiota Composition and Quality of Pacific White Shrimp (Litopenaeus vannamei) Stored at 0 °C. Food Microbiol. 2019, 83, 27–35. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alpha Diversity Indices | Stone Region | Algal Region |
|---|---|---|
| ACE | 2.8 ± 0.53 | 7.1 ± 1.29 |
| Chao1 | 2.8 ± 0.53 | 7.1 ± 1.29 |
| Simpson | 0.23 ± 0.06 | 0.33 ± 0.06 |
| Shannon | 0.68 ± 0.17 | 0.74 ± 0.13 |
| Alpha Diversity Indices | Stone Region | Algal Region |
|---|---|---|
| ACE | 149.1 ± 15.1 | 84 ± 9.87 |
| Chao1 | 149.1 ± 15.1 | 84 ± 9.87 |
| Simpson | 0.93 ± 0.03 | 0.9 ± 0.05 |
| Shannon | 5.8 ± 0.36 | 4.87 ± 0.37 |
| Alpha Diversity Indices | Stone Region | Algal Region |
|---|---|---|
| ACE | 149.1 ± 15.1 | 84 ± 9.87 |
| Chao1 | 149.1 ± 15.1 | 84 ± 9.87 |
| Simpson | 0.93 ± 0.03 | 0.9 ± 0.05 |
| Shannon | 5.8 ± 0.36 | 4.87 ± 0.37 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Guo, Y.; Qin, C.; Mu, X.; Zhang, J. High-Throughput Sequencing Analysis Revealed a Preference for Animal-Based Food in Purple Sea Urchins. Biology 2024, 13, 623. https://doi.org/10.3390/biology13080623
Liu Z, Guo Y, Qin C, Mu X, Zhang J. High-Throughput Sequencing Analysis Revealed a Preference for Animal-Based Food in Purple Sea Urchins. Biology. 2024; 13(8):623. https://doi.org/10.3390/biology13080623
Chicago/Turabian StyleLiu, Zerui, Yu Guo, Chuanxin Qin, Xiaohui Mu, and Jia Zhang. 2024. "High-Throughput Sequencing Analysis Revealed a Preference for Animal-Based Food in Purple Sea Urchins" Biology 13, no. 8: 623. https://doi.org/10.3390/biology13080623
APA StyleLiu, Z., Guo, Y., Qin, C., Mu, X., & Zhang, J. (2024). High-Throughput Sequencing Analysis Revealed a Preference for Animal-Based Food in Purple Sea Urchins. Biology, 13(8), 623. https://doi.org/10.3390/biology13080623