



Unveiling Lovastatin’s Anti-Inflammatory Potential in Mouse’s Brain during Acute Trypanosoma cruzi Infection
 , ,
, ,  , ,
, ,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Parasite
2.3. Experimental Groups
2.4. Drug and Treatment Scheme
2.5. Parasitemia, Body Weight, and Survival
2.6. Intravital Video Microscopy
2.7. Assessment of Functional Capillary Density
2.8. Leukocyte Rolling and Adhesion Analysis
2.9. Analysis of Brain Blood Flow
2.10. Immunohistochemistry
2.11. Western Blot
2.12. Statistical Analysis
3. Results
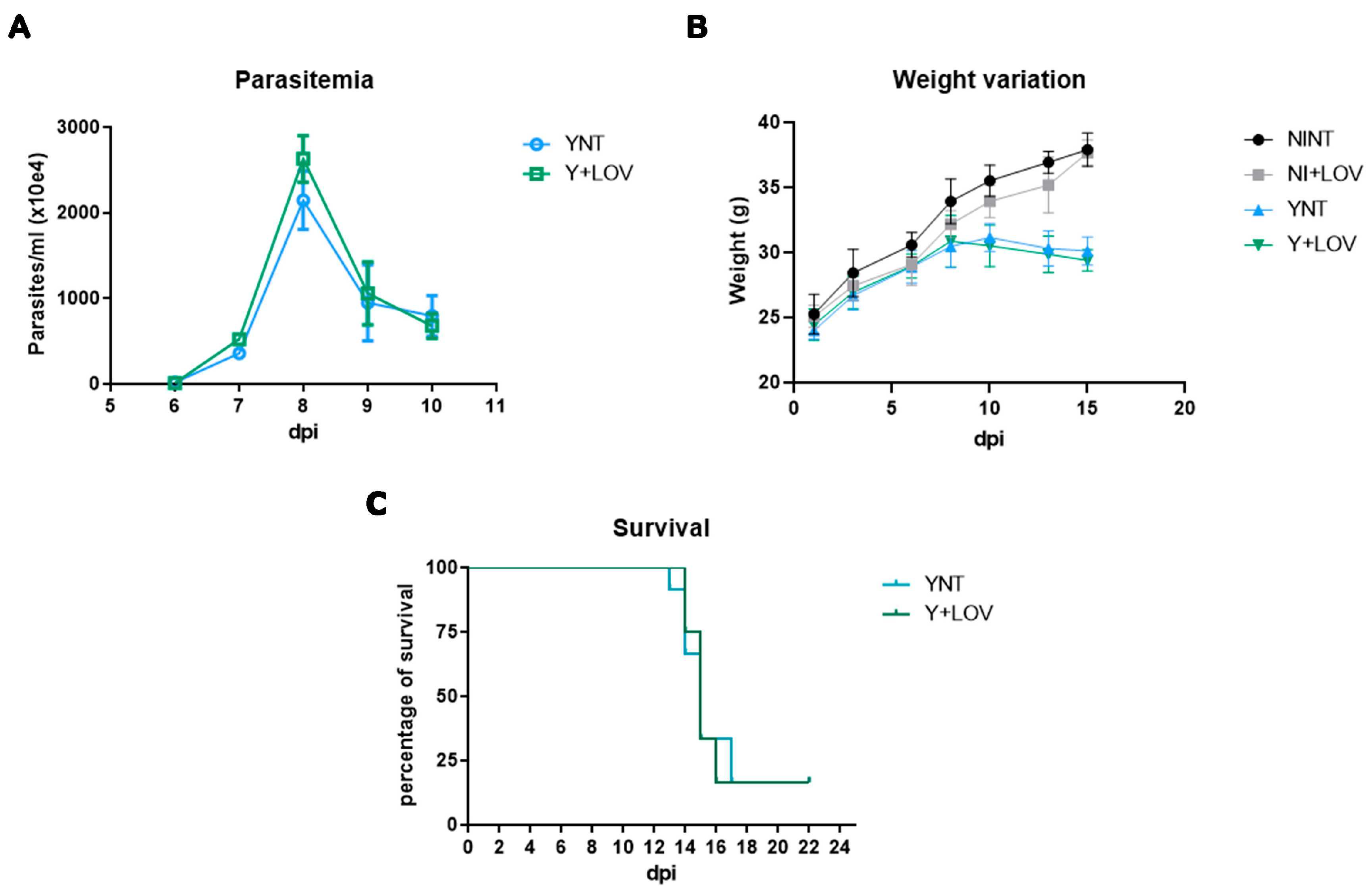
3.1. Effect of Lovastatin on Parasitemia, Body Weight, and Survival during T. cruzi Infection in Mice
3.2. Influence of Lovastatin on Cerebral Microvasculopathy Caused by T. cruzi Infection in Mice
3.3. Effect of Lovastatin Treatment on Cerebral Leukocyte–Endothelium Interactions during Acute T. cruzi Infection in Mice
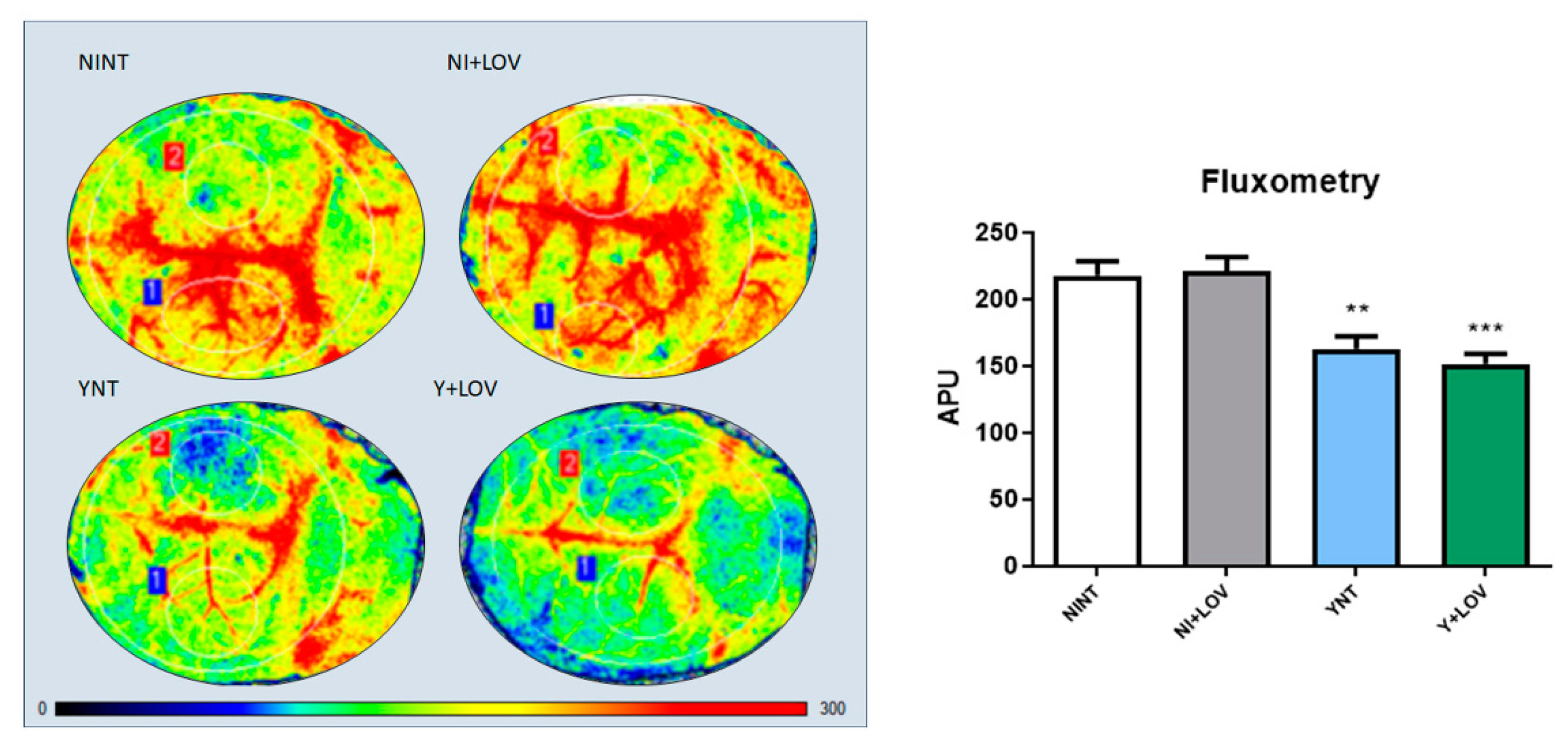
3.4. Influence of Lovastatin on the Cerebral Blood Flow Reduction Induced by Acute T. cruzi Infection in Mice
3.5. Influence of Lovastatin Treatment on the Increase in F4/80+ and CD3+ Cells in the Brain Caused by Acute T. cruzi Infection in Mice
3.6. Impact of Lovastatin on the Increased Inflammatory Mediators ICAM-1 and VCAM-1 in the Brain during Acute T. cruzi Infection in Mice
3.7. Influence of Lovastatin Treatment on MCP-1 Levels in the Brain during Acute T. cruzi Infection in Mice
3.8. Effect of Lovastatin on eNOS Levels in the Brain during Acute T. cruzi Infection in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Chagas’ Disease (American Trypanosomiasis)—Fact Sheet; WHO: Geneva, Switzerland, 2022. [Google Scholar]
- DNDi. Drugs for Neglected Diseases Initiative—Chagas Plataform; DNDi: Geneva, Switzerland, 2020. [Google Scholar]
- Rassi, A.; Rassi, A.; Marin-Neto, J.A. Chagas disease. Lancet 2020, 375, 1388–1402. [Google Scholar] [CrossRef]
- Prata, A. Clinical and epidemiological aspects of Chagas disease. Lancet Infect. Dis. 2001, 1, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Córdova, E.; Maiolo, E.; Corti, M.; Orduña, T. Neurological manifestations of Chagas’ disease. Neurol. Res. 2010, 32, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.C. Chagas disease: Still a challenge around the world. Rev. Soc. Bras. Med. Trop. 2015, 48, 367–369. [Google Scholar] [CrossRef]
- Vianna, G. Contribuição para o estudo da anatomia patolojica da “Molestia de Carlos Chagas” (Esquizotripanoze humana ou tireoidite parazitaria). Mem. Inst. Oswaldo Cruz 1911, 3, 276–294. [Google Scholar] [CrossRef]
- Chagas, C. Aspectos Clínicos y Anatomopatológicos de Latripanosomiasis Americana; La Prensa Médica Argentina: Buenos Aires, Argentina, 1916; Volume 3, pp. 125–127, 137–138, 153–158. [Google Scholar]
- Carod-Artal, F.J.; Ribeiro Lda, S.; Vargas, A.P. Awareness of stroke risk in chagasic stroke patients. J. Neurol. Sci. 2007, 263, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Coates, J.R.; Jeffery, N.D. Perspectives on meningoencephalomyelitis of unknown origin. Vet Clin. N. Am. Small Anim. Pract. 2014, 44, 1157–1185. [Google Scholar] [CrossRef] [PubMed]
- Py, M.O. Neurologic manifestations of Chagas disease. Curr. Neurol. Neurosci. Rep. 2011, 11, 536–542. [Google Scholar] [CrossRef]
- Pittella, J.E. Central nervous system involvement in Chagas disease: A hundred-year-old history. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 973–978. [Google Scholar] [CrossRef]
- Silva, A.A.; Roffe, E.; Marino, A.P.; dos Santos, P.V.; Quirico-Santos, T.; Paiva, C.N.; Lannes-Vieira, J. Chagas’ disease encephalitis: Intense CD8+ lymphocytic infiltrate is 67 restricted to the acute phase, but is not related to the presence of Trypanosoma cruzi antigens. Clin. Immunol. 1999, 92, 56–66. [Google Scholar] [CrossRef]
- Carod-Artal, F.J.; Vargas, A.P.; Falcao, T. Stroke in asymptomatic Trypanosoma cruzi-infected patients. Cerebrovasc. Dis. 2011, 31, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Carod-Artal, F.J.; Vargas, A.P.; Horan, T.A.; Nunes, L.G. Chagasic cardiomyopathy is independently associated with ischemic stroke in Chagas disease. Stroke 2005, 36, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Carod-Artal, F.J.; Gascon, J. Chagas disease and stroke. Lancet Neurol. 2010, 9, 533–542. [Google Scholar] [CrossRef]
- Nisimura, L.M.; Estato, V.; de Souza, E.M.; Reis, P.A.; Lessa, M.A.; Castro-Faria-Neto, H.C.; Pereira, M.C.; Tibiriçá, E.; Garzoni, L.R. Acute Chagas disease induces cerebral microvasculopathy in mice. PLoS Negl. Trop. Dis. 2014, 8, e2998. [Google Scholar] [CrossRef]
- Gonzaga, B.M.S.; Horita, S.I.M.; Beghini, D.G.; Gomes, F.; Nisimura, L.M.; Dos Santos, I.B.; Estato, V.; de Araújo-Jorge, T.C.; Garzoni, L.R. Effect of benznidazole on cerebral microcirculation during acute Trypanosoma cruzi infection in mice. Sci. Rep. 2022, 12, 21048. [Google Scholar] [CrossRef]
- Coura, J.R.; Borges-Pereira, J. Chagas disease. What is known and what should be improved: A systemic review. Rev. Soc. Bras. Med. Trop. 2012, 45, 286–296. [Google Scholar] [CrossRef]
- Ferreira, A.M.; Damasceno, R.F.; Monteiro-Junior, R.S.; Oliveira, I.A.C.; de Prates, T.E.C.; Nunes, M.C.P.; Haikal, D.S. Reações adversas ao benzonidazol no tratamento da Doença de Chagas: Revisão sistemática de ensaios clínicos randomizados e controlados. Cad. Saúde Coletiva 2019, 27, 354–362. [Google Scholar] [CrossRef]
- Viotti, R.; Vigliano, C.; Lococo, B.; Bertocchi, G.; Petti, M.; Alvarez, M.G.; Postan, M.; Armenti, A. Long-Term Cardiac Outcomes of Treating Chronic Chagas Disease with Benznidazole versus No Treatment. Ann. Intern. Med. 2006, 144, 724. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Kang, H.J.; Jhon, M.; Kim, J.W.; Lee, J.Y.; Walker, A.J.; Agustini, B.; Kim, J.M.; Berk, M. Statins and Inflammation: New Therapeutic Opportunities in Psychiatry. Front. Psychiatry 2019, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Fernandez, C.H.; Cameron, J.C. Is statin-associated cognitive impairment clinically relevant? A narrative review and clinical recommendations. Ann. Pharmacother. 2012, 46, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Kessler, R.L.; Soares, M.J.; Probst, C.M.; Krieger, M.A. Trypanosoma cruzi response to sterol biosynthesis inhibitors: Morphophysiological alterations leading to cell death. PLoS ONE 2013, 8, e55497. [Google Scholar] [CrossRef]
- Urbina, J.A.; Lazardi, K.; Marchan, E.; Visbal, G.; Aguirre, T.; Piras, M.M.; Piras, R.; Maldonado, R.A.; Payares, G.; de Souza, W. Mevinolin (lovastatin) potentiates the antiproliferative effects of ketoconazole and terbinafine against Trypanosoma (Schizotrypanum) cruzi: In vitro and in vivo studies. Antimicrob. Agents Chemother. 1993, 37, 580–591. [Google Scholar] [CrossRef]
- Silva, R.R.; Shrestha-Bajracharya, D.; Almeida-Leite, C.M.; Leite, R.; Bahia, M.T.; Talvani, A. Short-term therapy with simvastatin reduces inflammatory mediators and heart inflammation during the acute phase of experimental Chagas disease. Mem. Inst. Oswaldo Cruz 2012, 107, 513–521. [Google Scholar] [CrossRef]
- Pena, D.A.; Eger, I.; Nogueira, L.; Heck, N.; Menin, A.; Báfica, A.; Steindel, M. Selection of TcII Trypanosoma cruzi Population Following Macrophage Infection. J. Infect. Dis. 2011, 204, 478–486. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Melo, L.; Caldas, I.S.; Azevedo, M.A.; Gonçalves, K.R.; do Nascimento, A.F.d.S.; Figueiredo, V.P.; de Figueiredo Diniz, L.; de Lima, W.G.; Torres, R.M.; Bahia, M.T.; et al. Low doses of simvastatin therapy ameliorate cardiac inflammatory remodeling in Trypanosoma cruzi-infected dogs. Am. J. Trop. Med. Hyg. 2011, 84, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Freitas, J.M.; Lages-Silva, E.; Crema, E.; Pena, S.D.J.; Macedo, A.M. Real time PCR strategy for the identification of major lineages of Trypanosoma cruzi directly in chronically infected humans tissues. Int. J. Parasitol. 2005, 35, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Reis, P.A.; Estato, V.; da Silva, T.I.; d’Avila, J.C.; Siqueira, L.D.; Assis, E.F.; Bozza, P.T.; Bozza, F.A.; Tibiriça, E.V.; Zimmerman, G.A.; et al. Statins Decrease Neuroinflammation and Prevent Cognitive Impairment after Cerebral Malaria. PLoS Pathog. 2012, 8, e1003099. [Google Scholar] [CrossRef] [PubMed]
- Da Mata, J.R.; Camargos, M.R.; Chiari, E.; Machado, C.R. Trypanosoma cruzi infection and the rat central nervous system: Proliferation of parasites in astrocytes and the brain reaction to parasitism. Brain Res. Bull. 2000, 53, 153–162. [Google Scholar] [PubMed]
- Roffê, E.; Silva, A.A.; Marino, A.P.; dos Santos, P.V.; Lannes-Vieira, J. Essential role of VLA-4/VCAM-1 pathway in the establishment of CD8+ T-cell-mediated Trypanosoma cruzi-elicited meningoencephalitis. J. Neuroimmunol. 2003, 142, 17–30. [Google Scholar] [CrossRef] [PubMed]
- González-Herrera, F.; Cramer, A.; Pimentel, P.; Castillo, C.; Liempi, A.; Kemmerling, U.; Machado, F.S.; Maya, J.D. Simvastatin Attenuates Endothelial Activation through 15-Epi-Lipoxin A4 Production in Murine Chronic Chagas Cardiomyopathy. Antimicrob. Agents Chemother. 2017, 61, e02137-16. [Google Scholar] [CrossRef]
- Rachid, M.A.; Teixeira, A.L.; Barcelos, L.S.; Machado, C.R.; Chiari, E.; Tanowitz, H.B.; Camargos, E.R.; Teixeira, M.M. Role of endothelin receptors in the control of central nervous system parasitism in Trypanosoma cruzi infection in rats. J. Neuroimmunol. 2010, 220, 64–68. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, D.; Yang, X.; Zhang, Y.; Li, P.; Su, X. Lysophosphatidic acid and lovastatin might protect kidney in renal I/R injury by downregulating MCP-1 in rat. Ren Fail. 2011, 33, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, B.; Melby, P.C.; Troyer, D.A.; Freeman, G.L. Differential regulation of nitric oxide synthase isoforms in experimental acute chagasic cardiomyopathy. Clin. Exp. Immunol. 2000, 121, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.F.; Capettini, L.S.; da Silva, J.F.; Sales-Junior, P.; Cruz, J.S.; Cortes, S.F.; Lemos, V.S. Mechanisms of vascular dysfunction in acute phase of Trypanosoma cruzi infection in mice. Vasc. Pharmacol. 2016, 82, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Sumeray, M.S.; Rees, D.D.; Yellon, D.M. Infarct size and nitric oxide synthase in murine myocardium. J. Mol. Cell. Cardiol. 2000, 32, 35–42. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies Used for Western Blot | Manufacturer | Catalog Number | Dilution | Protein Concentration |
|---|---|---|---|---|
| Primary antibody: Anti-ICAM-1 | Abcam | ab124760 | 1:1000 | 20 μg |
| Primary antibody: Anti-VCAM1 | Abcam | ab115135 | 1:1000 | 20 μg |
| Primary antibody: Anti-MCP-1 | Abcam | ab702 | 1:1000 | 20 μg |
| Primary antibody: Anti-eNOS | Abcam | ab5589 | 1:1000 | 100 μg |
| Primary antibody: Anti-GAPDH | Fitzgerald | 10R-G109a | 1:40,000 | - |
| Secondary antibody: Anti-rabbit with peroxidase | Thermo Scientific | 31,430 | 1:10,000 | - |
| Anticorpo secundário de cabra | Thermo Scientific | 31,460 | 1:10,000 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzaga, B.M.d.S.; Nisimura, L.M.; Coelho, L.L.; Ferreira, R.R.; Horita, S.I.M.; Beghini, D.G.; Estato, V.; Araújo-Jorge, T.C.d.; Garzoni, L.R. Unveiling Lovastatin’s Anti-Inflammatory Potential in Mouse’s Brain during Acute Trypanosoma cruzi Infection. Biology 2024, 13, 301. https://doi.org/10.3390/biology13050301
Gonzaga BMdS, Nisimura LM, Coelho LL, Ferreira RR, Horita SIM, Beghini DG, Estato V, Araújo-Jorge TCd, Garzoni LR. Unveiling Lovastatin’s Anti-Inflammatory Potential in Mouse’s Brain during Acute Trypanosoma cruzi Infection. Biology. 2024; 13(5):301. https://doi.org/10.3390/biology13050301
Chicago/Turabian StyleGonzaga, Beatriz Matheus de Souza, Líndice Mitie Nisimura, Laura Lacerda Coelho, Roberto Rodrigues Ferreira, Samuel Iwao Maia Horita, Daniela Gois Beghini, Vanessa Estato, Tania Cremonini de Araújo-Jorge, and Luciana Ribeiro Garzoni. 2024. "Unveiling Lovastatin’s Anti-Inflammatory Potential in Mouse’s Brain during Acute Trypanosoma cruzi Infection" Biology 13, no. 5: 301. https://doi.org/10.3390/biology13050301
APA StyleGonzaga, B. M. d. S., Nisimura, L. M., Coelho, L. L., Ferreira, R. R., Horita, S. I. M., Beghini, D. G., Estato, V., Araújo-Jorge, T. C. d., & Garzoni, L. R. (2024). Unveiling Lovastatin’s Anti-Inflammatory Potential in Mouse’s Brain during Acute Trypanosoma cruzi Infection. Biology, 13(5), 301. https://doi.org/10.3390/biology13050301