



Prevalence and Diversity of Trypanosoma cruzi in Triatomine Vectors and Their Blood Meal Sources from South Central Texas, USA
 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
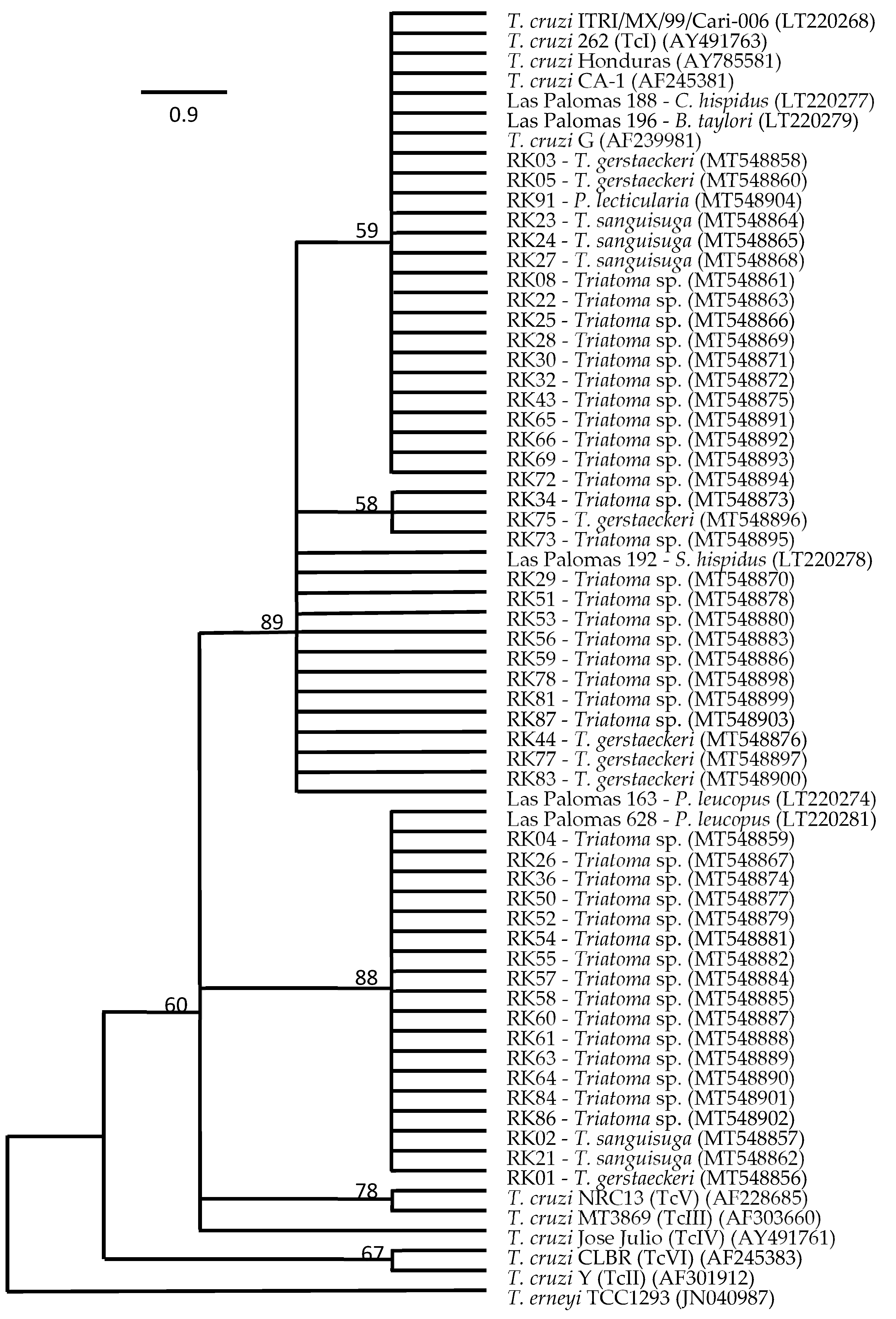
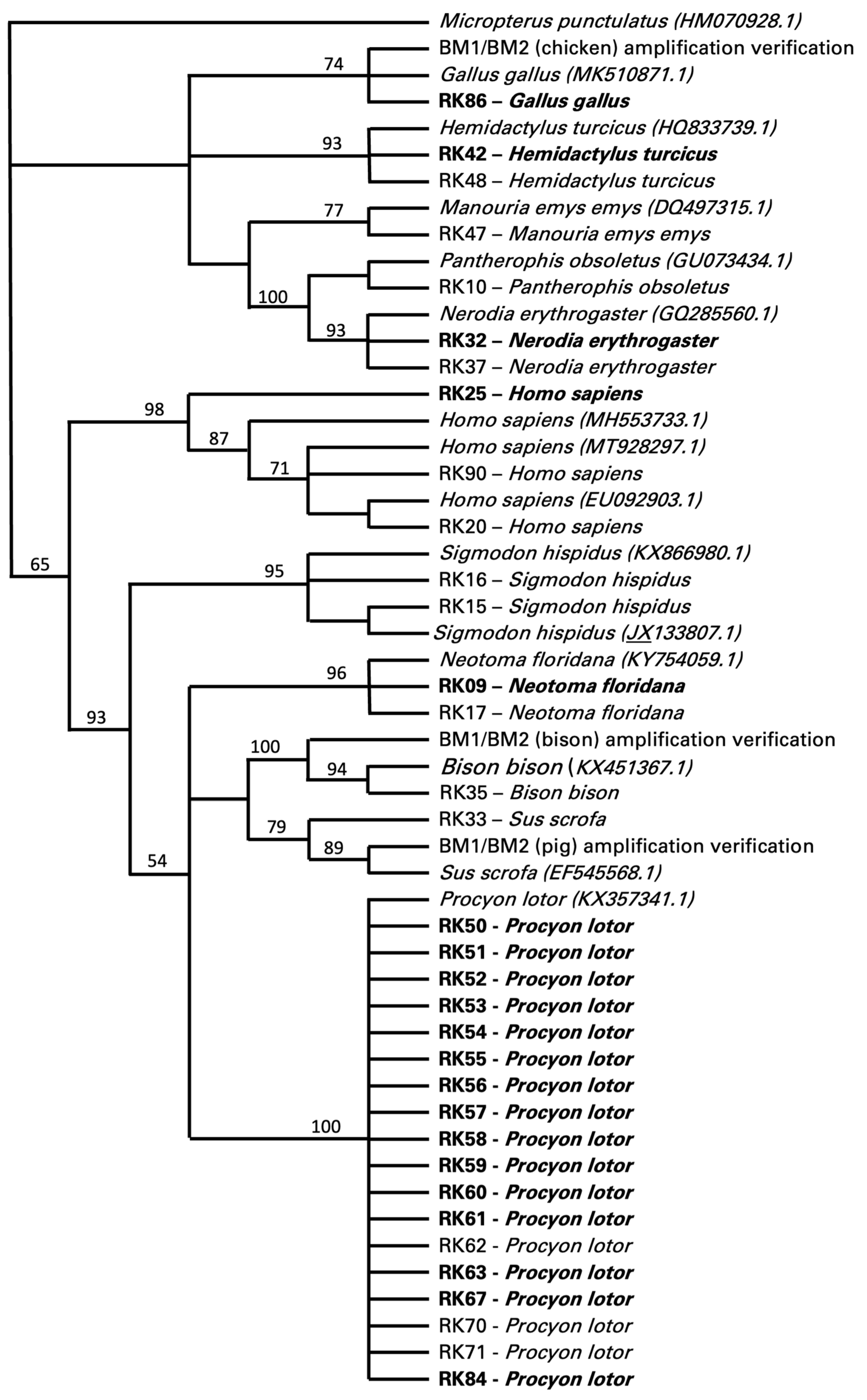
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bern, C.; Messenger, L.A.; Whitman, J.D.; Maguire, J.H. Chagas disease in the United States: A public health approach. Clin. Microbiol. Rev. 2019, 33, e00023-19. [Google Scholar] [CrossRef] [PubMed]
- Bern, C.; Kjos, S.; Yabsley, M.J.; Montgomery, S.P. Trypanosoma cruzi and Chagas’ disease in the United States. Clin. Microbiol. Rev. 2011, 24, 655–681. [Google Scholar] [CrossRef]
- Curtis-Robles, R.; Lewis, B.C.; Hamer, S.A. High Trypanosoma cruzi infection prevalence associated with minimal cardiac pathology among wild carnivores in central Texas. Int. J. Parasitol. Parasites Wildl. 2016, 5, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Munoz-San Martin, C.; Campo Verde Arbocco, F.; Saavedra, M.; Actis, E.A.; Rios, T.A.; Abba, A.M.; Morales, M.E.; Cattan, P.E.; Jahn, G.A.; Superina, M. High rates of Trypanosoma cruzi infection in goats from Mendoza province, Argentina: Parasite loads in blood and seasonal variation. Acta Trop. 2020, 208, 105493. [Google Scholar] [CrossRef] [PubMed]
- Herrera, L.; D’Andrea, P.S.; Xavier, S.C.; Mangia, R.H.; Fernandes, O.; Jansen, A.M. Trypanosoma cruzi infection in wild mammals of the National Park ‘Serra da Capivara’ and its surroundings (Piaui, Brazil), an area endemic for Chagas disease. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 379–388. [Google Scholar] [CrossRef]
- Rodriguez, F.; Luna, B.S.; Calderon, O.; Manriquez-Roman, C.; Amezcua-Winter, K.; Cedillo, J.; Garcia-Vazquez, R.; Tejeda, I.A.; Romero, A.; Waldrup, K.; et al. Surveillance of Trypanosoma cruzi infection in triatomine vectors, feral dogs and cats, and wild animals in and around El Paso County, Texas, and New Mexico. PLoS Negl. Trop. Dis. 2021, 15, e0009147. [Google Scholar] [CrossRef]
- Coura, J.R.; Dias, J.C. Epidemiology, control and surveillance of Chagas disease: 100 years after its discovery. Mem. Inst. Oswaldo Cruz. 2009, 104 (Suppl. 1), 31–40. [Google Scholar] [CrossRef] [PubMed]
- Aleman, A.; Guerra, T.; Maikis, T.J.; Milholland, M.T.; Castro-Arellano, I.; Forstner, M.R.J.; Hahn, D. The prevalence of Trypanosoma cruzi, the causal agent of Chagas disease, in Texas rodent populations. Ecohealth 2017, 14, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Gorchakov, R.; Trosclair, L.P.; Wozniak, E.J.; Feria, P.T.; Garcia, M.N.; Gunter, S.M.; Murray, K.O. Trypanosoma cruzi infection prevalence and bloodmeal analysis in triatomine vectors of Chagas disease from rural peridomestic locations in Texas, 2013–2014. J. Med. Entomol. 2016, 53, 911–918. [Google Scholar] [CrossRef]
- Curtis-Robles, R.; Auckland, L.D.; Snowden, K.F.; Hamer, G.L.; Hamer, S.A. Analysis of over 1500 triatomine vectors from across the US, predominantly Texas, for Trypanosoma cruzi infection and discrete typing units. Infect. Genet. Evol. 2018, 58, 171–180. [Google Scholar] [CrossRef]
- Curtis-Robles, R.; Wozniak, E.J.; Auckland, L.D.; Hamer, G.L.; Hamer, S.A. Combining public health education and disease ecology research: Using citizen science to assess Chagas disease entomological risk in Texas. PLoS Neglect. Trop. D 2015, 9, e0004235. [Google Scholar] [CrossRef] [PubMed]
- Hudson, F.P.; Homer, N.; Epstein, A.; Mondy, K. Acute Chagas disease manifesting as orbital cellulitis, Texas, USA. Emerg Infect. Dis. 2021, 27, 2937–2939. [Google Scholar] [CrossRef] [PubMed]
- Comeaux, J.M.; Curtis-Robles, R.; Lewis, B.C.; Cummings, K.J.; Mesenbrink, B.T.; Leland, B.R.; Bodenchuk, M.J.; Hamer, S.A. Survey of feral swine (Sus scrofa) infection with the agent of Chagas disease (Trypanosoma cruzi) in Texas, 2013–2014. J. Wildl. Dis. 2016, 52, 627–630. [Google Scholar] [CrossRef] [PubMed]
- Kjos, S.A.; Snowden, K.F.; Olson, J.K. Biogeography and Trypanosoma cruzi infection prevalence of Chagas disease vectors in Texas, USA. Vector-Borne Zoonot. 2009, 9, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Curtis-Robles, R.; Meyers, A.C.; Auckland, L.D.; Zecca, I.B.; Skiles, R.; Hamer, S.A. Parasitic interactions among Trypanosoma cruzi, triatomine vectors, domestic animals, and wildlife in Big Bend National Park along the Texas-Mexico border. Acta Trop. 2018, 188, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Piron, M.; Fisa, R.; Casamitjana, N.; Lopez-Chejade, P.; Puig, L.; Verges, M.; Gascon, J.; Prat, J.G.I.; Portus, M.; Sauleda, S. Development of a real-time PCR assay for Trypanosoma cruzi detection in blood samples. Acta Trop. 2007, 103, 195–200. [Google Scholar] [PubMed]
- Bosseno, M.F.; Barnabe, C.; Gastelum, E.M.; Kasten, F.L.; Ramsey, J.; Espinoza, B.; Breniere, S.F. Predominance of Trypanosoma cruzi lineage I in Mexico. J. Clin. Microbiol. 2002, 40, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Moreira, O.C.; Ramirez, J.D.; Velazquez, E.; Melo, M.F.A.D.; Lima-Ferreira, C.; Guhl, F.; Sosa-Estani, S.; Marin-Neto, J.A.; Morillo, C.A.; Britto, C. Towards the establishment of a consensus real-time qPCR to monitor Trypanosoma cruzi parasitemia in patients with chronic Chagas disease cardiomyopathy: A substudy from the BENEFIT trial. Acta Trop. 2013, 125, 23–31. [Google Scholar]
- Pinto, C.M.; Ocana-Mayorga, S.; Tapia, E.E.; Lobos, S.E.; Zurita, A.P.; Aguirre-Villacís, F.; MacDonald, A.; Villacís, A.G.; Lima, L.; Teixeira, M.M.G.; et al. Bats, trypanosomes, and triatomines in Ecuador: New insights into the diversity, transmission, and origins of Trypanosoma cruzi and Chagas disease. PLoS ONE 2015, 10, e0139999. [Google Scholar]
- Noyes, H.A.; Stevens, J.R.; Teixeira, M.; Phelan, J.; Holz, P. A nested PCR for the ssrRNA gene detects Trypanosoma binneyi in the platypus and Trypanosoma sp. in wombats and kangaroos in Australia. Int. J. Parasitol. 1999, 29, 331–339. [Google Scholar] [CrossRef]
- Murphy, W.J.; O’Brien, S.J. Designing and optimizing comparative anchor primers for comparative gene mapping and phylogenetic inference. Nat. Protoc. 2007, 2, 3022–3030. [Google Scholar] [CrossRef] [PubMed]
- Kjos, S.A.; Marcet, P.L.; Yabsley, M.J.; Kitron, U.; Snowden, K.F.; Logan, K.S.; Barnes, J.C.; Dotson, E.M. Identification of bloodmeal sources and Trypanosoma cruzi infection in triatomine bugs (Hemiptera: Reduviidae) from residential settings in Texas, the United States. J. Med. Entomol. 2013, 50, 1126–1139. [Google Scholar] [CrossRef]
- Monteiro, F.A.; Barrett, T.V.; Fitzpatrick, S.; Cordon-Rosales, C.; Feliciangeli, D.; Beard, C.B. Molecular phylogeography of the Amazonian Chagas disease vectors Rhodnius prolixus and R. robustus. Mol. Ecol. 2003, 12, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Boakye, D.A.; Tang, J.; Truc, P.; Merriweather, A.; Unnasch, T.R. Identification of bloodmeals in haematophagous Diptera by Cytochrome B heteroduplex analysis. Med. Vet Entomol. 1999, 13, 282–287. [Google Scholar] [CrossRef]
- Molaei, G.; Oliver, J.; Andreadis, T.G.; Armstrong, P.M.; Howard, J.J. Molecular identification of blood-meal sources in Culiseta melanura and Culiseta morsitans from an endemic focus of eastern equine encephalitis virus in New York. Am. J. Trop. Med. Hyg. 2006, 75, 1140–1147. [Google Scholar] [CrossRef]
- Cicero, C.; Johnson, N.K. Higher-level phylogeny of new world vireos (Aves: Vireonidae) based on sequences of multiple mitochondrial DNA genes. Mol. Phylogenet. Evol. 2001, 20, 27–40. [Google Scholar] [CrossRef]
- Pfeiler, E.; Bitler, B.G.; Ramsey, J.M.; Palacios-Cardiel, C.; Markow, T.A. Genetic variation, population structure, and phylogenetic relationships of Triatoma rubida and T. recurva (Hemiptera: Reduviidae: Triatominae) from the Sonoran Desert, insect vectors of the Chagas’ disease parasite Trypanosoma cruzi. Mol. Phylogenet. Evol. 2006, 41, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Justi, S.A.; Russo, C.A.M.; Mallet, J.R.D.; Obara, M.T.; Galvao, C. Molecular phylogeny of Triatomini (Hemiptera: Reduviidae: Triatominae). Parasite Vector 2014, 7, 149. [Google Scholar] [CrossRef]
- Pearson, W.R.; Lipman, D.J. Improved tools for biological sequence comparison. Proc. Nat. Acad. Sci. USA 1988, 85, 2444–2448. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-joining Method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits of phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.E.; Lineberry, M.W. Detection of Trypansoma cruzi in kissing bugs (Hemiptera: Reduviidae: Triatominae) collected across Oklahoma. J. Med. Entomol. 2022, 59, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, D.H.; Koch, K.; Roachell, W.; Delgado, B.; Bast, J. First record of an established population of Triatoma sanguisuga (Hemiptera: Reduviidae) in Richardson County, Nebraska. J. Med. Entomol. 2021, 58, 2519–2523. [Google Scholar] [CrossRef]
- Behrens-Bradley, N.; Smith, S.; Beatty, N.L.; Love, M.; Ahmad, N.; Dorn, P.L.; Schmidt, J.O.; Klotz, S.A. Kissing bugs harboring Trypanosoma cruzi, frequently bite residents of the US Southwest but do not cause Chagas disease. Am. J. Med. 2020, 133, 108–114.e13. [Google Scholar] [CrossRef]
- Zingales, B.; Andrade, S.G.; Briones, M.R.S.; Campbell, D.A.; Chiari, E.; Fernandes, O.; Guhl, F.; Lages-Silva, E.; Macedo, A.M.; Machado, C.R.; et al. A new consensus for Trypanosoma cruzi intraspecific nomenclature: Second revision meeting recommends TcI to TcVI. Mem. I Oswaldo Cruz. 2009, 104, 1051–1054. [Google Scholar] [CrossRef]
- Wozniak, E.J.; Lawrence, G.; Gorchakov, R.; Alamgir, H.; Dotson, E.; Sissel, B.; Sarkar, S.; Murray, K.O. The biology of the triatomine bugs native to South Central Texas and assessment of the risk they pose for autochthonous Chagas disease exposure. J. Parasitol. 2015, 101, 520–528. [Google Scholar] [CrossRef]
- Majeau, A.; Pronovost, H.; Sanford, A.; Cloherty, E.; Anderson, A.N.; Balsamo, G.; Gee, L.; Straif-Bourgeois, S.C.; Herrera, C. Raccoons as an important reservoir for Trypanosoma cruzi: A prevalence study from two metropolitan areas in Louisiana. Vector-Borne Zoonot. 2020, 20, 535–540. [Google Scholar] [CrossRef]
- Pietrzak, S.M.; Pung, O.J. Trypanosomiasis in raccoons from Georgia. J. Wildl. Dis. 1998, 34, 132–136. [Google Scholar] [CrossRef]
- Hodo, C.L.; Banuelos, R.M.; Edwards, E.E.; Wozniak, E.J.; Hamer, S.A. Pathology and discrete typing unit associations of Trypanosoma cruzi infection in coyotes (Canis latrans) and raccoons (Procyon lotor) of Texas, USA. J. Wildl. Dis. 2020, 56, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Ordonez-Krasnowski, P.C.; Lanati, L.A.; Gaspe, M.S.; Cardinal, M.V.; Ceballos, L.A.; Gurtler, R.E. Domestic host availability modifies human-triatomine contact and host shifts of the Chagas disease vector Triatoma infestans in the humid Argentine Chaco. Med. Vet Entomol. 2020, 34, 459–469. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequence (5′–3′) | Amplicon Size | PCR Cycles | Cited |
|---|---|---|---|---|
| Detection of T. cruzi (qPCR) | ||||
| Cruzi 1 Cruzi 2 | AST CGG CTG ATC GTT TTC GA AAT TCC TCC AAG CAG CGG ATA | 166 bp | 40 cycles 95 °C, 15 s 58 °C, 60 s | [16,18] |
| Identification of T. cruzi (comparative 18S rRNA gene amplicon sequence analysis) | ||||
| SSU4_F 18Sq1R | GTG CCA GCA CCC GCG GTA AT CCA CCG ACC AAA AGC GGC CA | 900 bp | 2 cycles each: 95 °C, 30 s 60–52 °C, 30 s 72 °C, 60 s | [19,20,21] |
| followed by | ||||
| SSU561F SSU561R | TGG GAT AAC AAA GGA GCA CTG AGA CTG TAA CCT CAA AGC | 650 bp | 30 cycles 95 °C, 30 s 50 °C, 30 s 72 °C, 60 s | |
| Identification of triatomines (comparative CytB amplicon sequence analysis) | ||||
| CytB7432F CytB7433R | GGACGW GGW ATT TAT TAT GGA TC GCW CCA ATT CAR GTT AR T AA | 663 bp | 35 cycles 94 °C, 30 s 47 °C, 30 s 72 °C, 60 s | [22,23] |
| Identification of triatomine food sources (comparative CytB amplicon sequence analysis) | ||||
| BM1 BM2 | CCC CTC AGA ATG ATA TTT GTC CTC A CCA TCC AAC ATC TCA GCA TGA TGA AA | 358 bp | 36 cycles 95 °C, 30 s 57 °C, 50 s 72 °C, 40 s | [22,24] |
| Mammalian a F Mammalian a R | CGA AGC TTG ATA TGA AAA ACC ATC GTT TGT AGT ART CWG GGT CHC CTA | 772 bp | 36 cycles 95 °C, 30 s 57 °C, 50 s 72 °C, 40 s | [22,25] |
| L15557 H16065 | GAC TGT GAC AAA ATC CCN TTC CA GGT CTT CAT CTY HGG YTT ACA AGA C | 508 bp | 36 cycles 95 °C, 30 s 57 °C, 50 s 72 °C, 40 s | [22,26] |
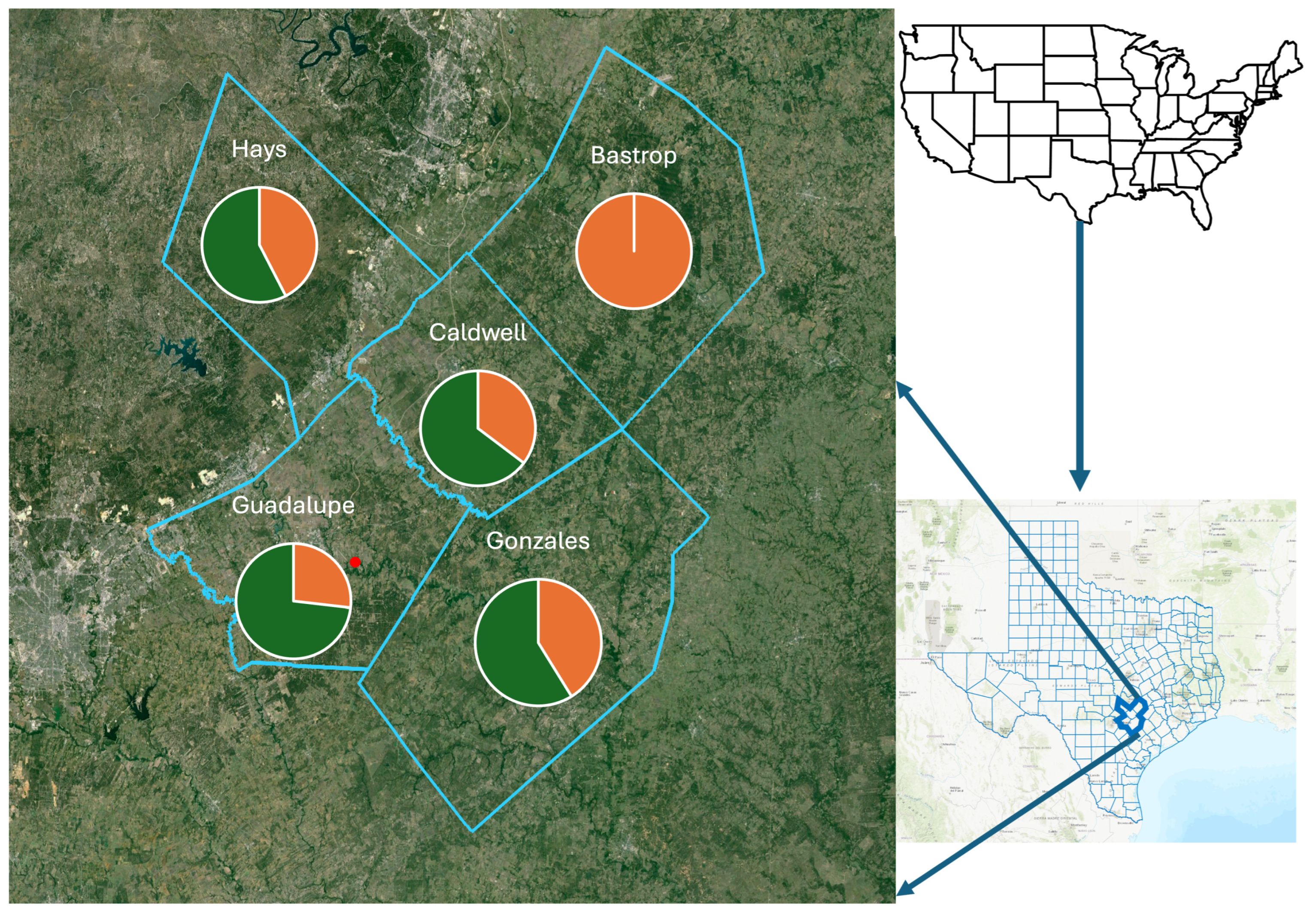
| County | Total Number of Triatomines | Triatomines Negative for T. cruzi | Triatomines Positive for T. cruzi | ||
|---|---|---|---|---|---|
| Adults | Nymphs | Adults | Nymphs | ||
| Hays | 33 | 3 | 11 | 18 | 1 |
| Guadalupe | 41 | 5 | 6 | 23 | 7 |
| Caldwell | 17 | 4 | 2 | 11 | 0 |
| Gonzales | 17 | 7 | 0 | 10 | 0 |
| Bastrop | 9 | 9 | 0 | 0 | 0 |
| Total | 117 | 28 | 19 | 62 | 8 |
| Species | Texas County of Capture | Negative for T. cruzi | Positive for T. cruzi | ||
|---|---|---|---|---|---|
| Adults | Nymphs | Adults | Nymphs | ||
| Triatoma gerstaeckeri | Bastrop | 9 | 0 | 0 | 0 |
| Caldwell | 3 | * | 3 | 0 | |
| Gonzales | 7 | 0 | 10 | * | |
| Guadalupe | * | * | 2 | * | |
| Hays | * | 6 | 5 | 1 | |
| Triatoma sanguisuga | Caldwell | 1 | * | 1 | 0 |
| Guadalupe | 1 | * | 4 | * | |
| Hays | 3 | * | 3 | * | |
| Paratriatoma lecticularia | Guadalupe | * | * | 2 | * |
| Triatomine | Caldwell | 0 | 2 | 7 | 0 |
| Guadalupe | 4 | 6 | 16 | 6 | |
| Hays | * | 5 | 9 | 1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kilgore, R.J.; Guerra, T.; Beck, H.; Villamizar Gomez, A.; Forstner, M.R.J.; Hahn, D. Prevalence and Diversity of Trypanosoma cruzi in Triatomine Vectors and Their Blood Meal Sources from South Central Texas, USA. Biology 2024, 13, 489. https://doi.org/10.3390/biology13070489
Kilgore RJ, Guerra T, Beck H, Villamizar Gomez A, Forstner MRJ, Hahn D. Prevalence and Diversity of Trypanosoma cruzi in Triatomine Vectors and Their Blood Meal Sources from South Central Texas, USA. Biology. 2024; 13(7):489. https://doi.org/10.3390/biology13070489
Chicago/Turabian StyleKilgore, Rebecca J., Trina Guerra, Heather Beck, Andrea Villamizar Gomez, Michael R. J. Forstner, and Dittmar Hahn. 2024. "Prevalence and Diversity of Trypanosoma cruzi in Triatomine Vectors and Their Blood Meal Sources from South Central Texas, USA" Biology 13, no. 7: 489. https://doi.org/10.3390/biology13070489
APA StyleKilgore, R. J., Guerra, T., Beck, H., Villamizar Gomez, A., Forstner, M. R. J., & Hahn, D. (2024). Prevalence and Diversity of Trypanosoma cruzi in Triatomine Vectors and Their Blood Meal Sources from South Central Texas, USA. Biology, 13(7), 489. https://doi.org/10.3390/biology13070489