
Response of Extremely Small Populations to Climate Change—A Case of Trachycarpus nanus in Yunnan, China

 and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
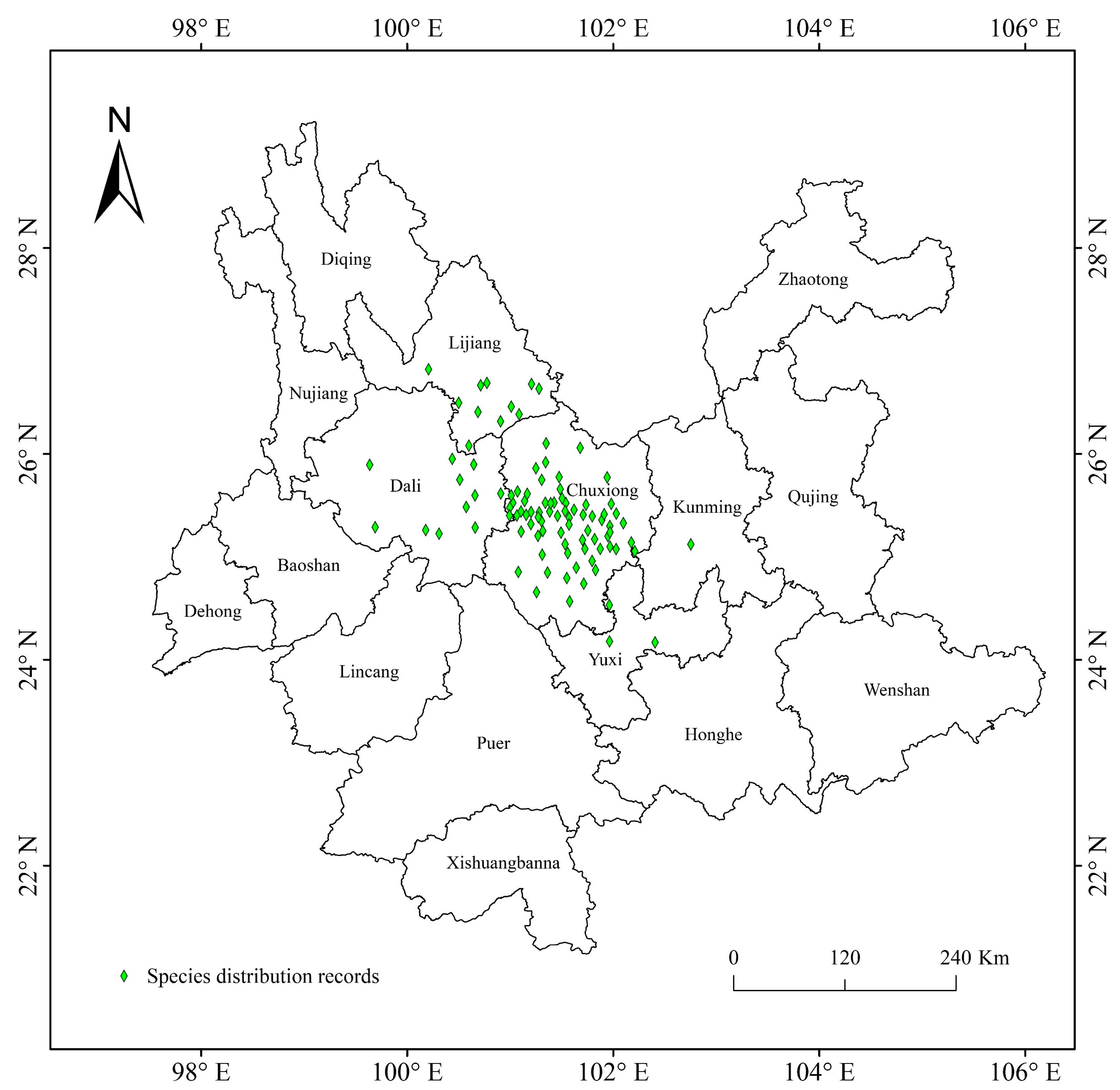
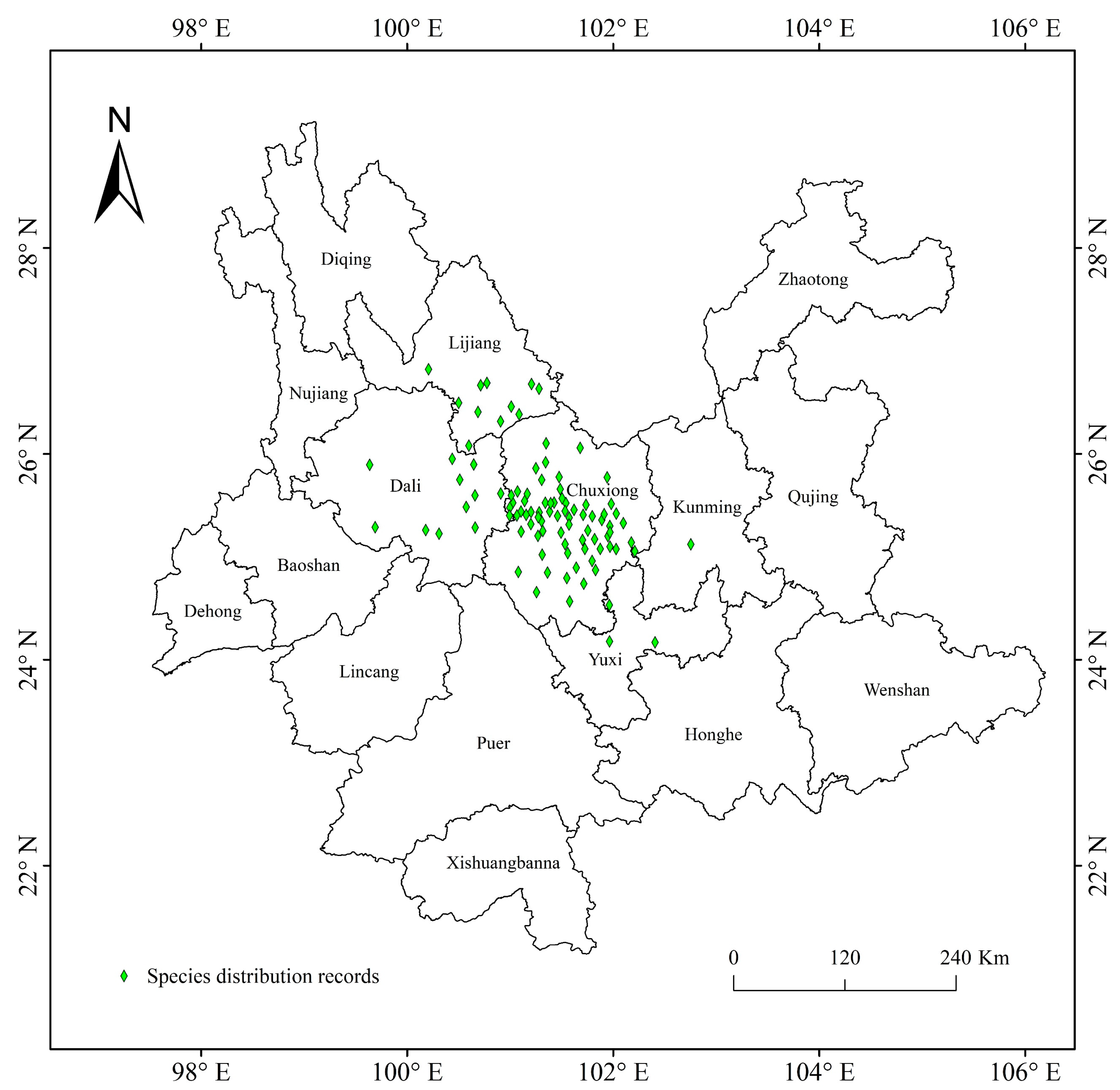
2.1. Species Distribution Data
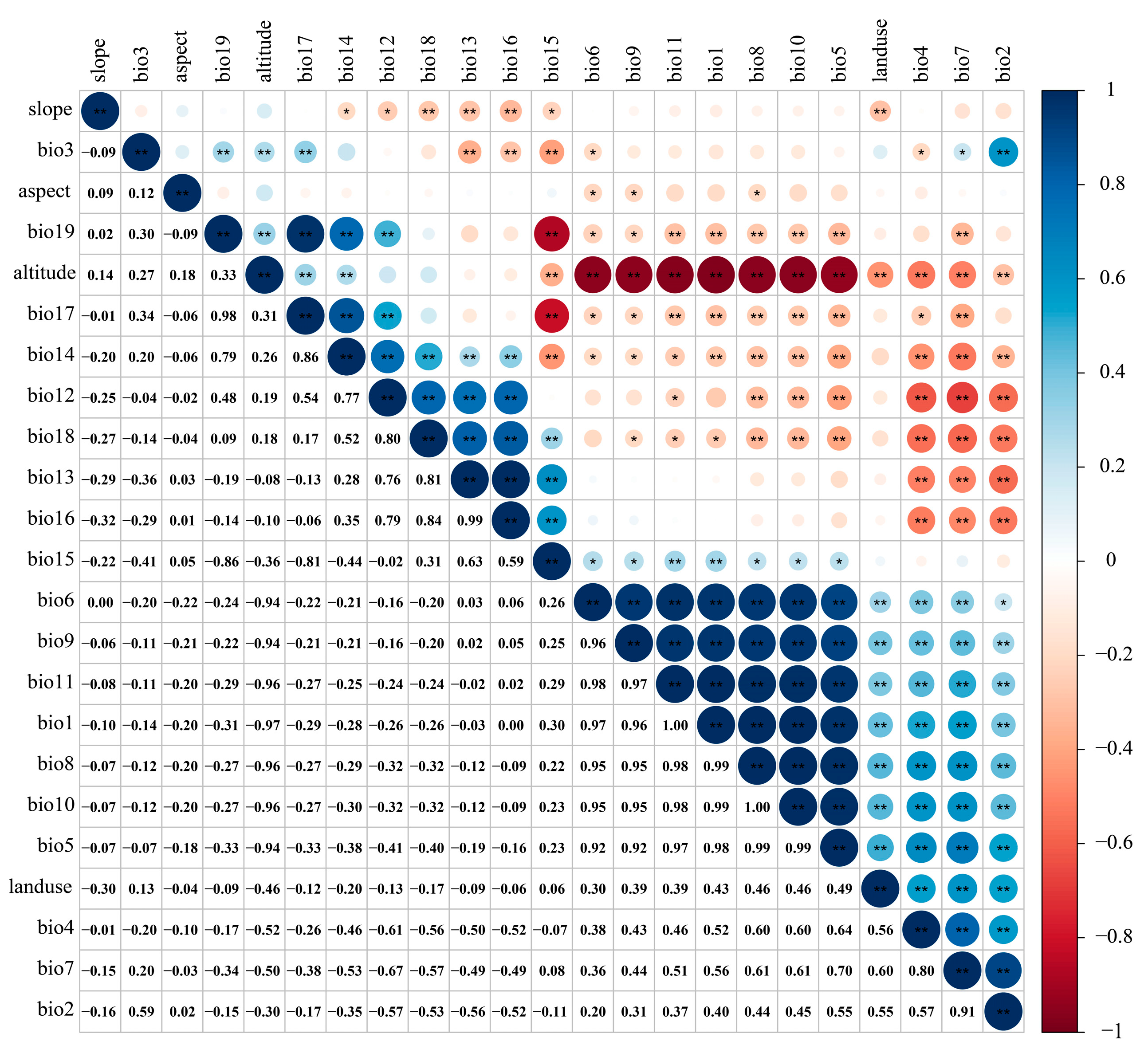
2.2. Environment Data
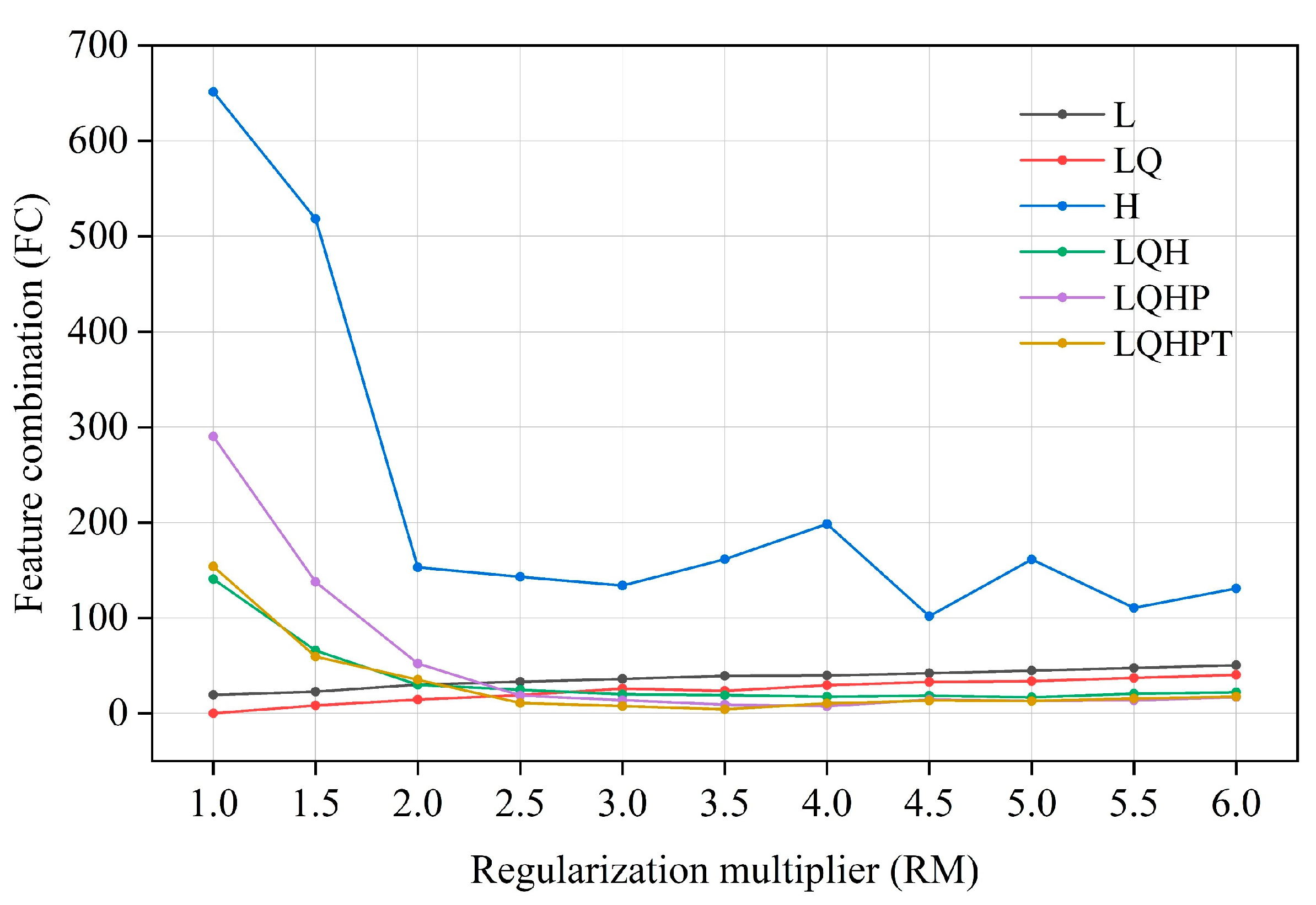
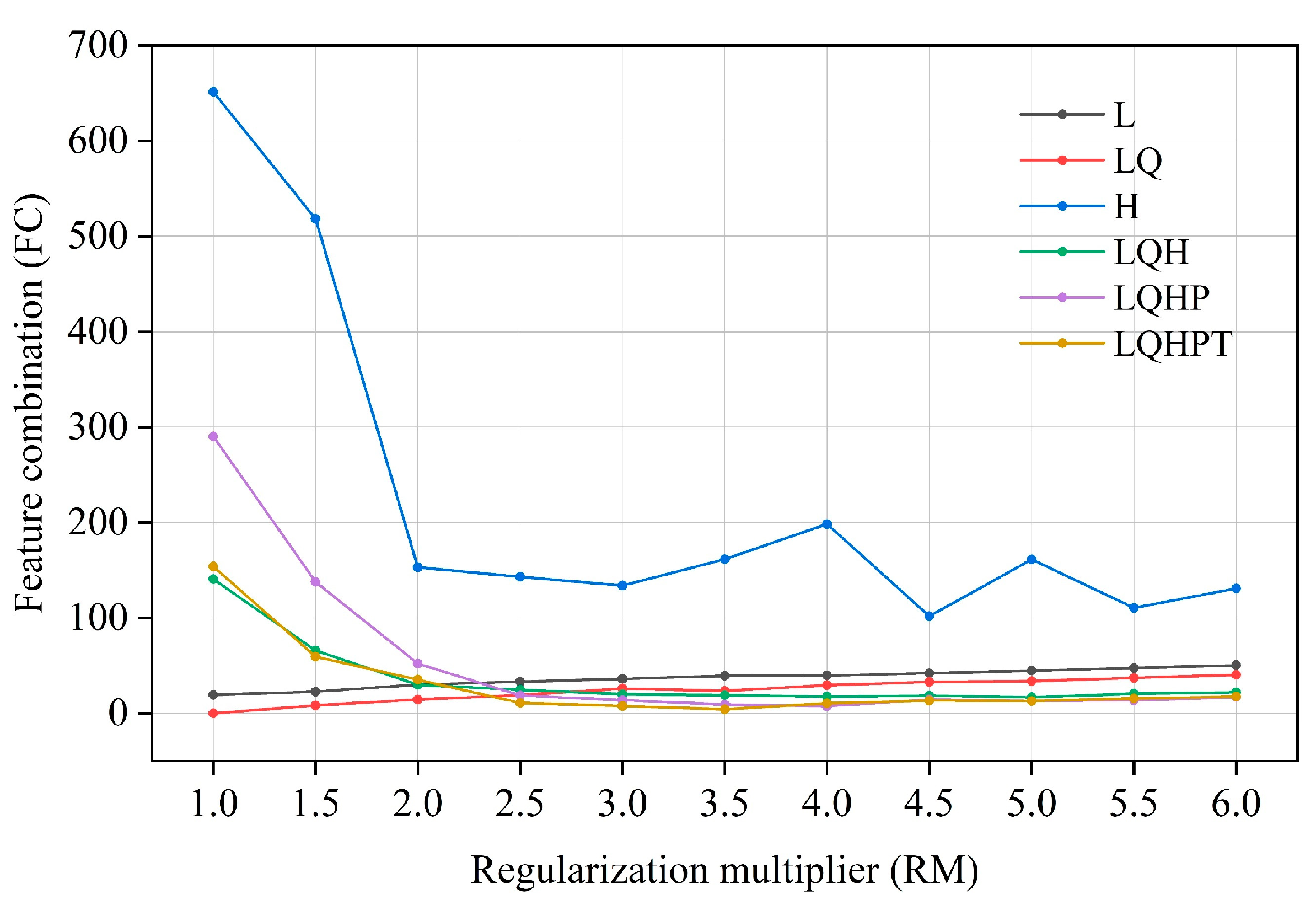
2.3. Model Setting and Evaluation
2.4. Classification of Potentially Suitable Area
2.5. Spatial Pattern Change and Centroid Analysis of Species’ Potentially Suitable Area
3. Results
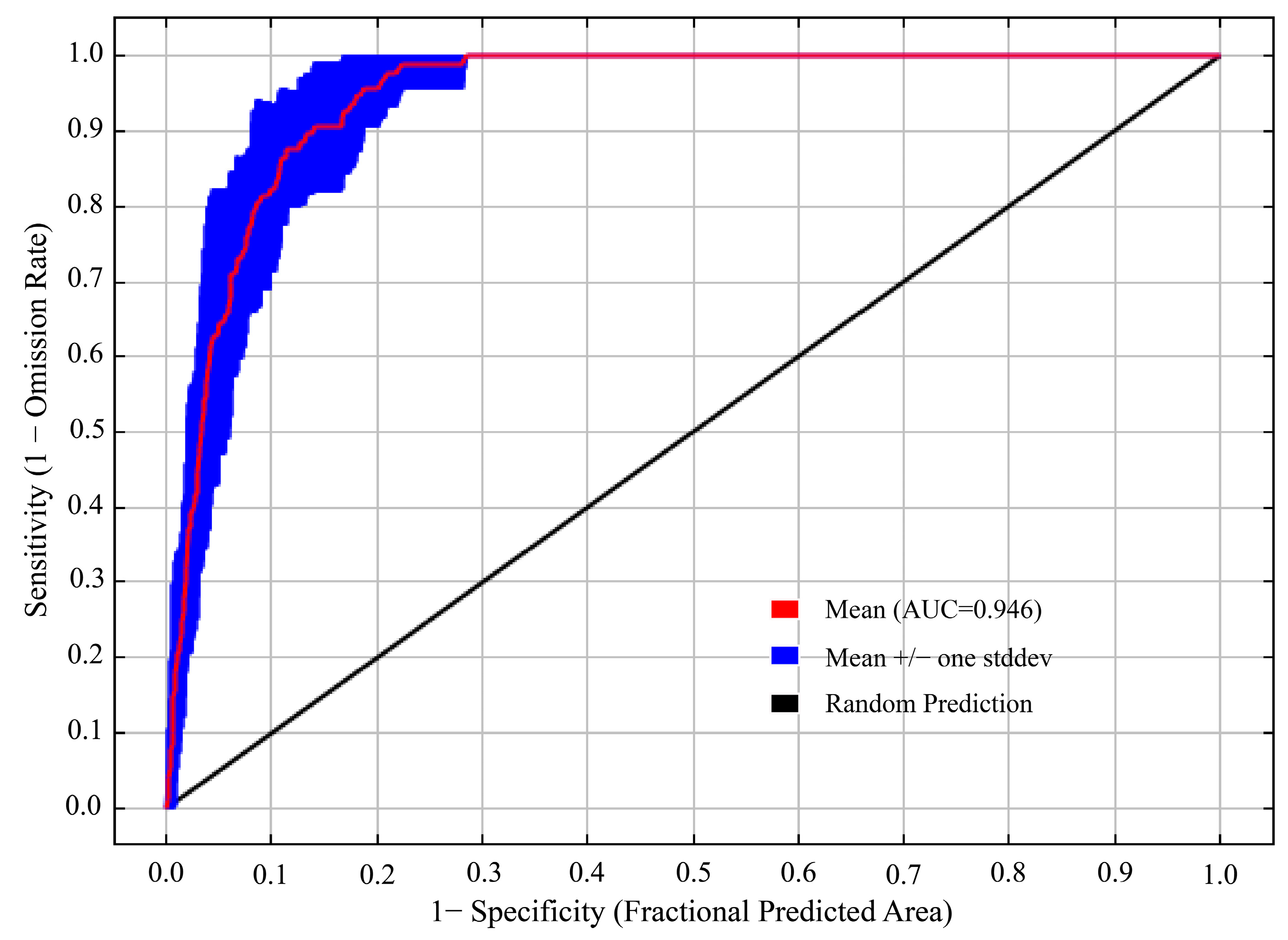
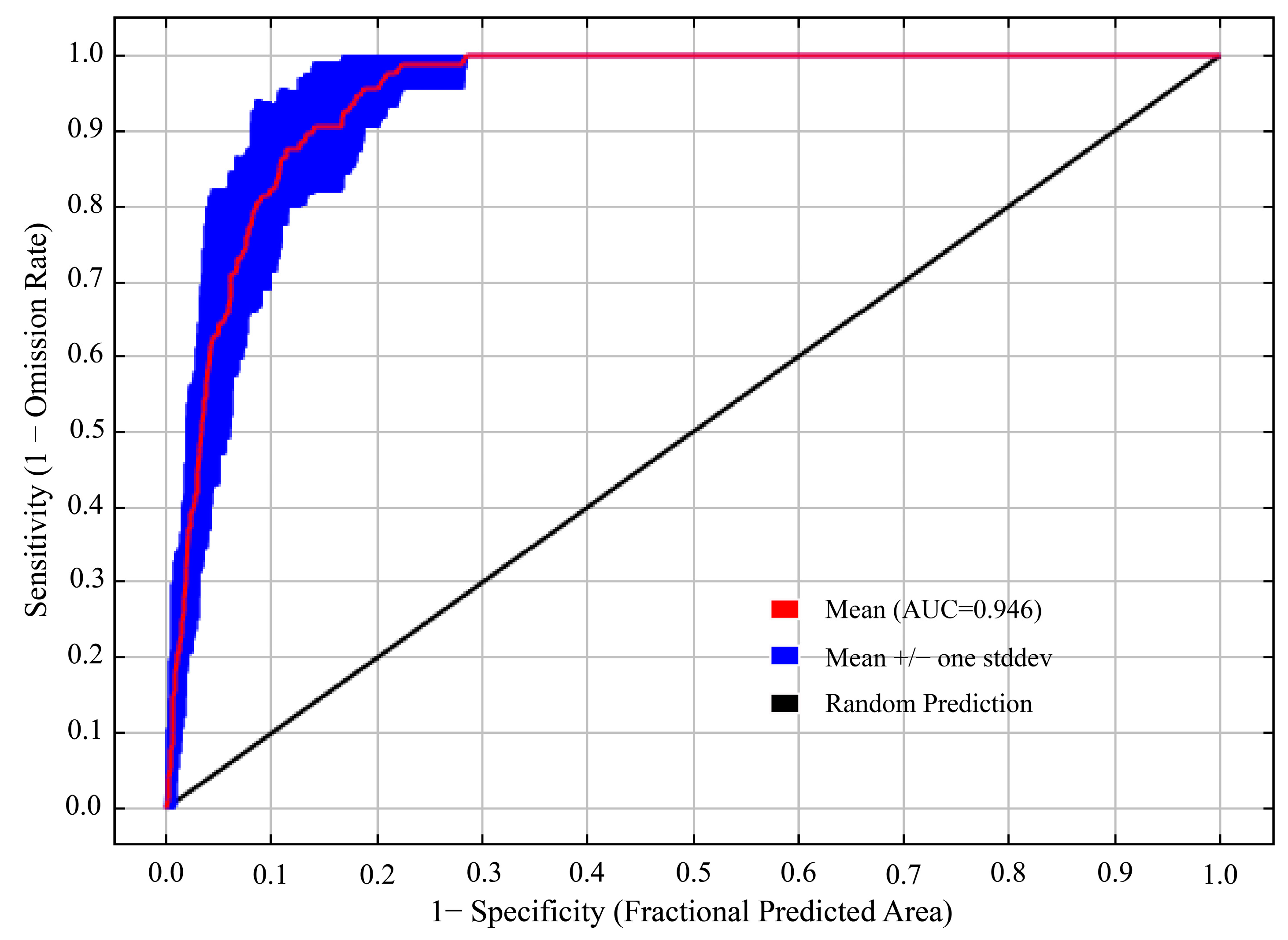
3.1. Accuracy Evaluation of the MaxEnt Model
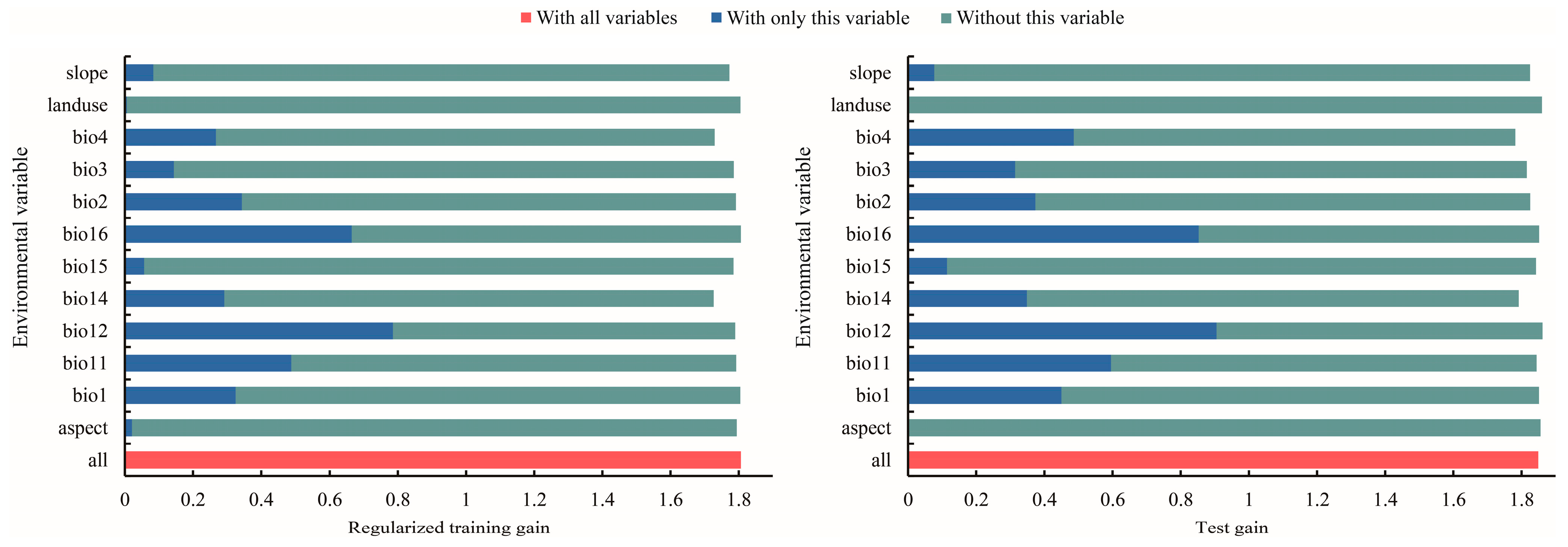
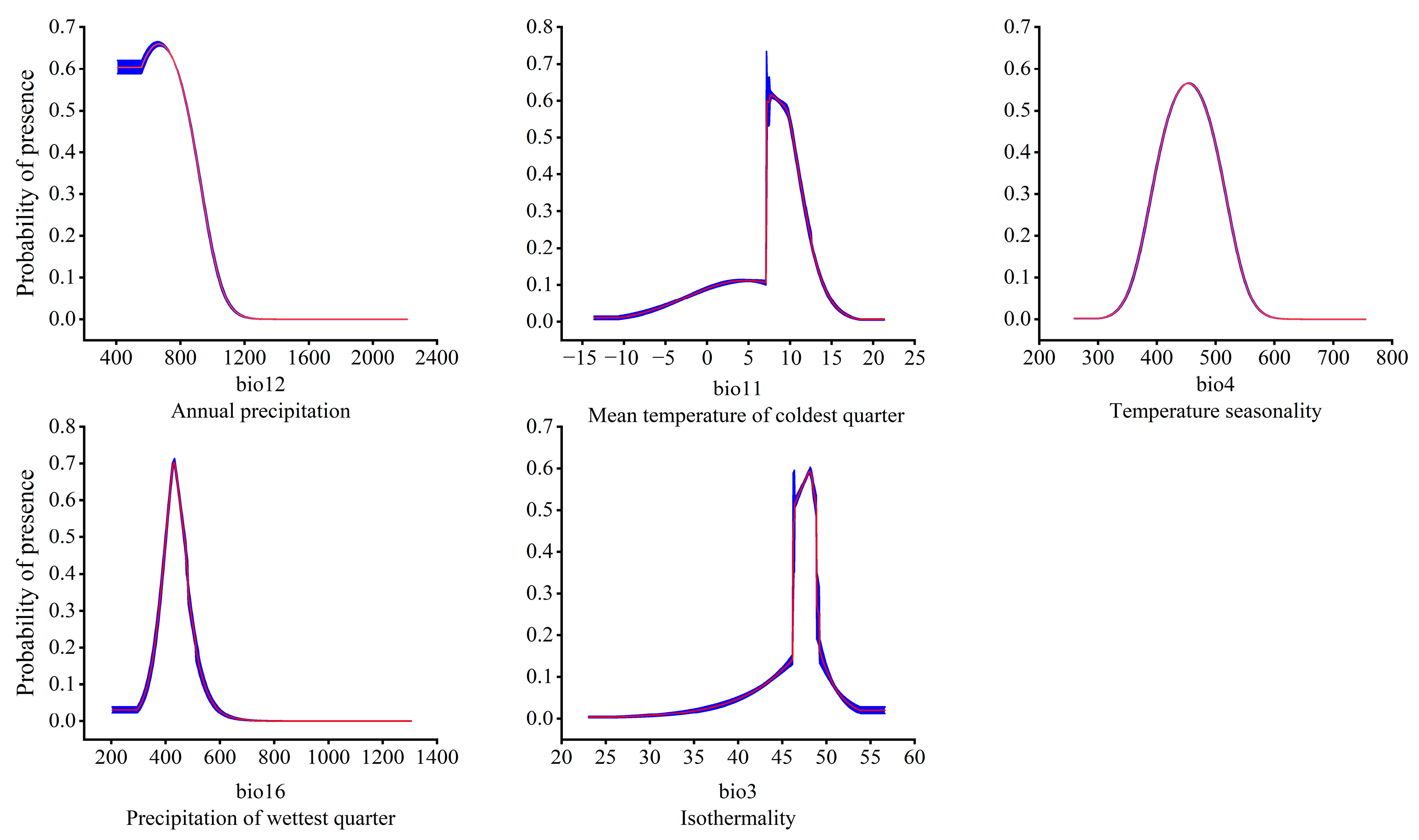
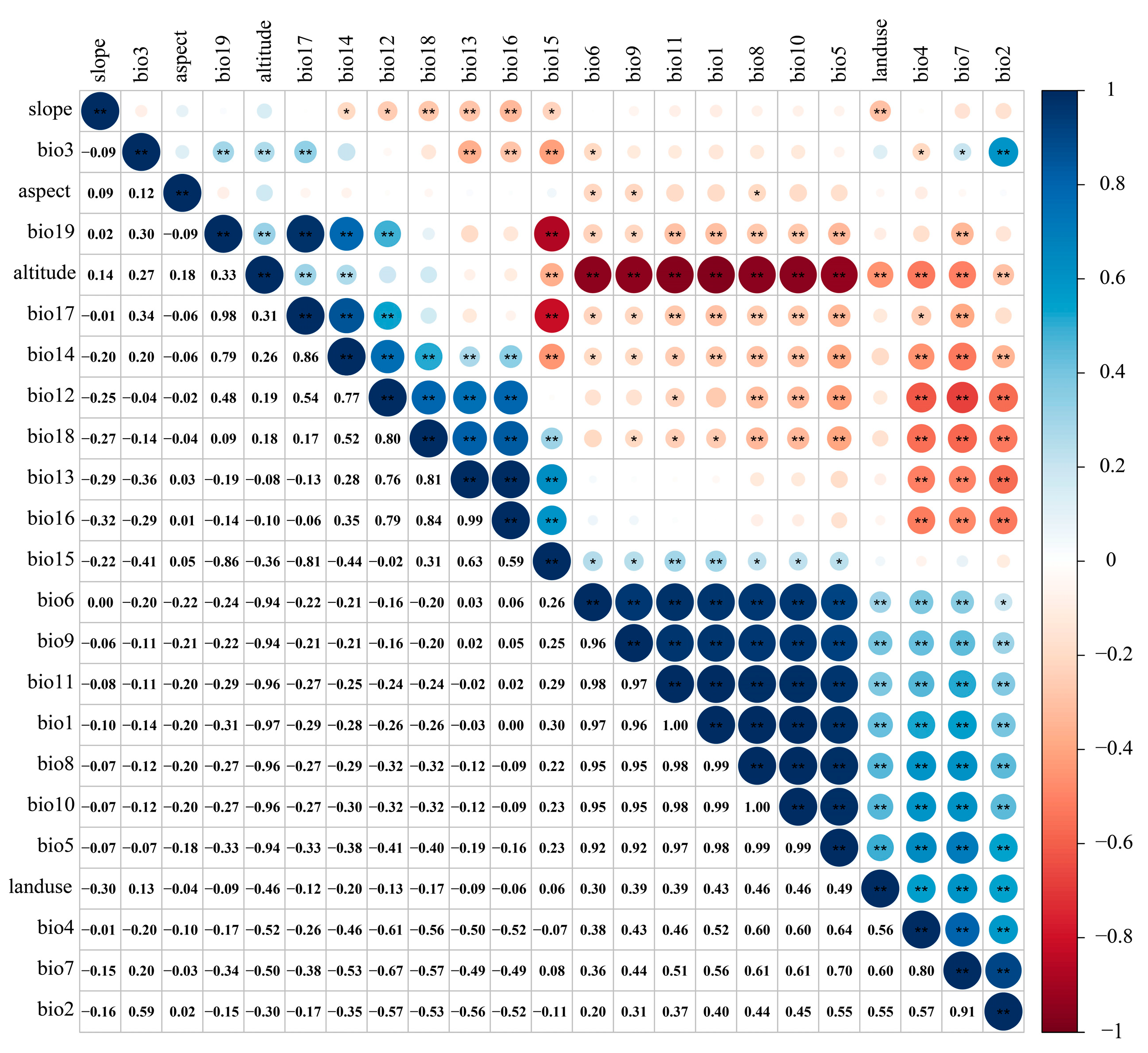
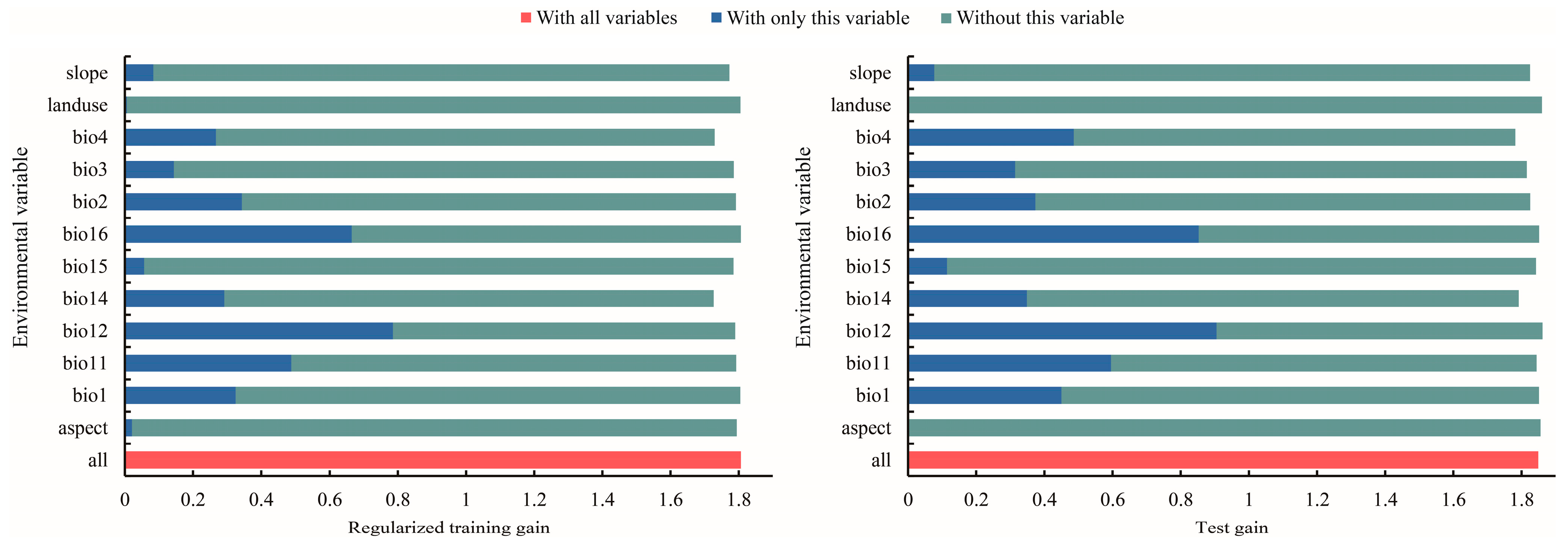
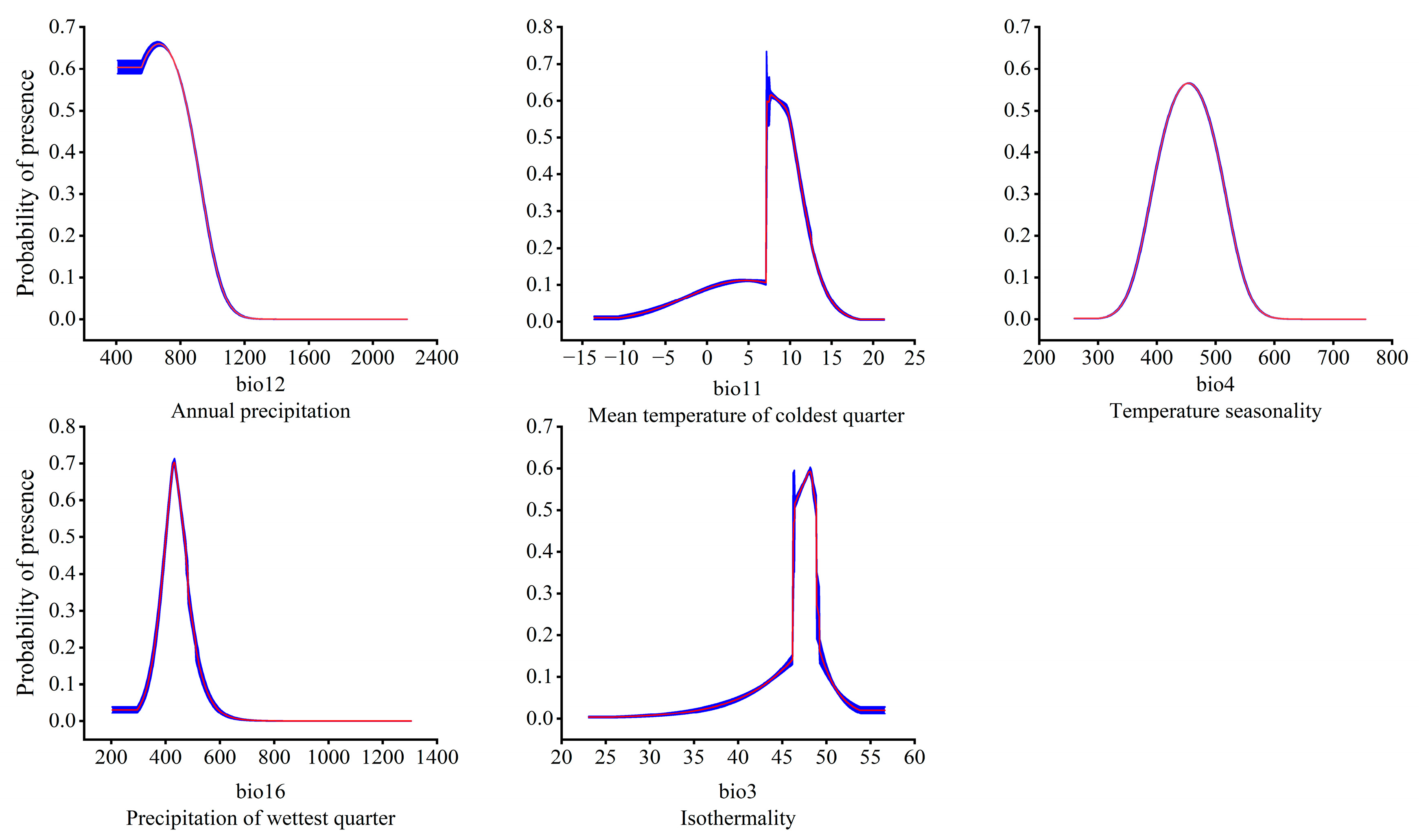
3.2. Dominant Environmental Factors Affecting the Distribution of T. nanus
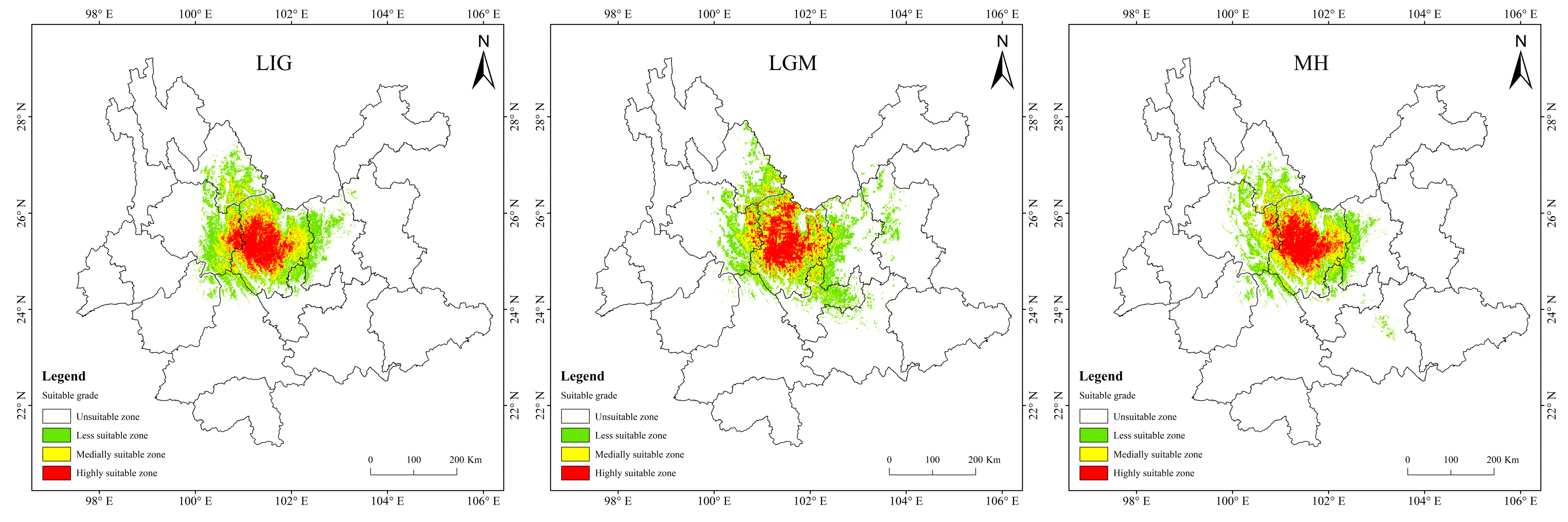
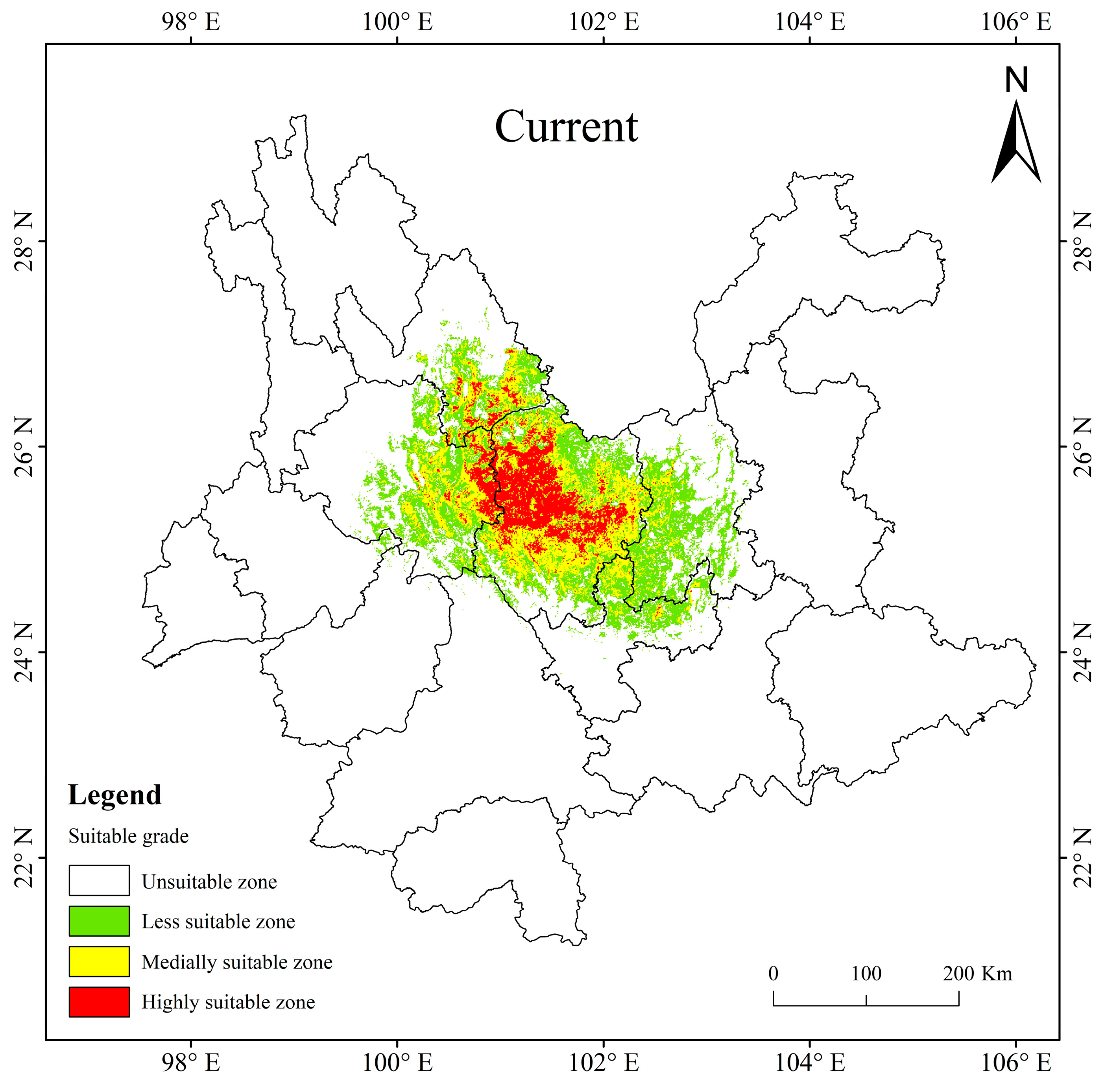
3.3. Present Potentially Suitable Area for T. nanus
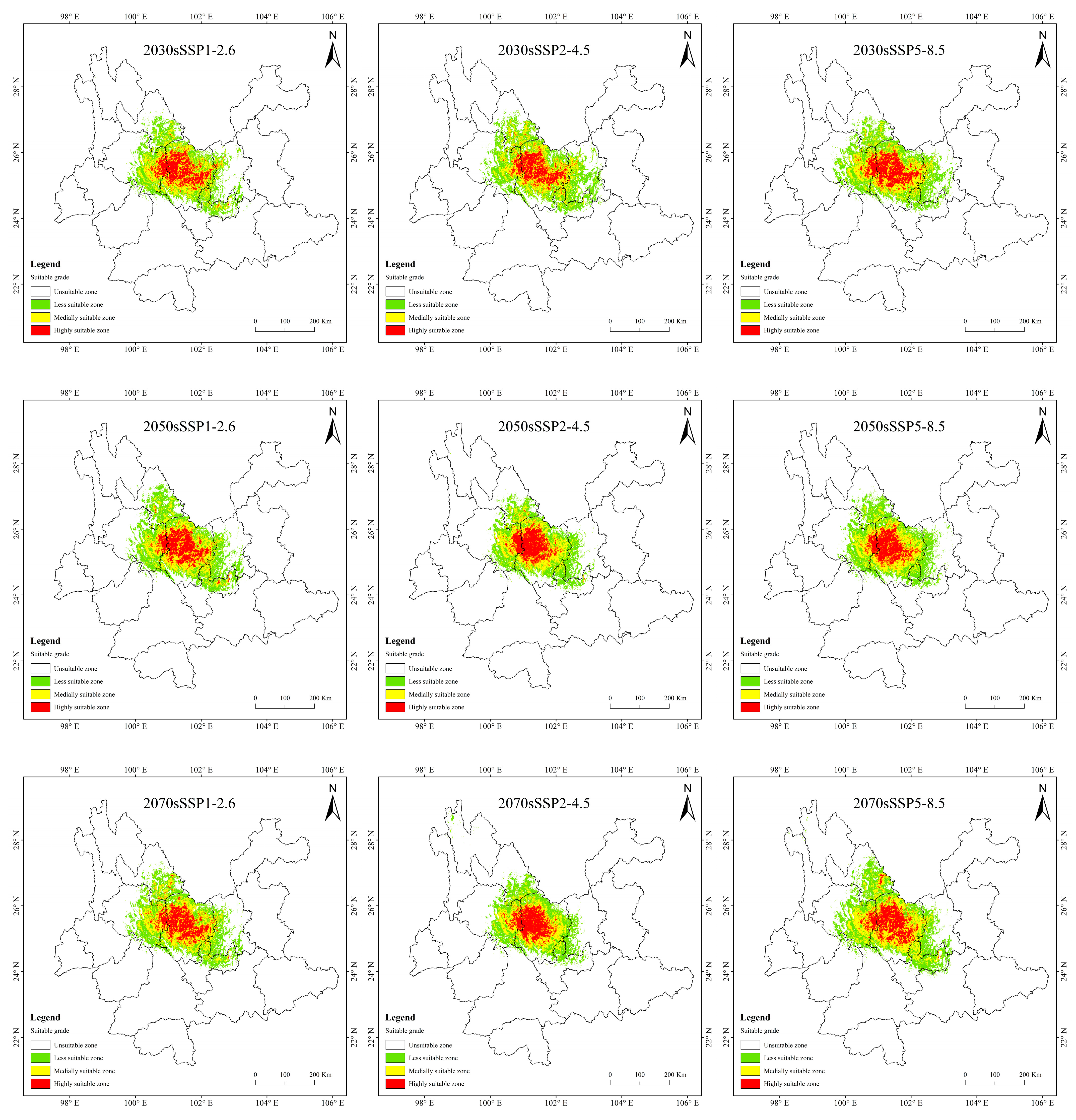
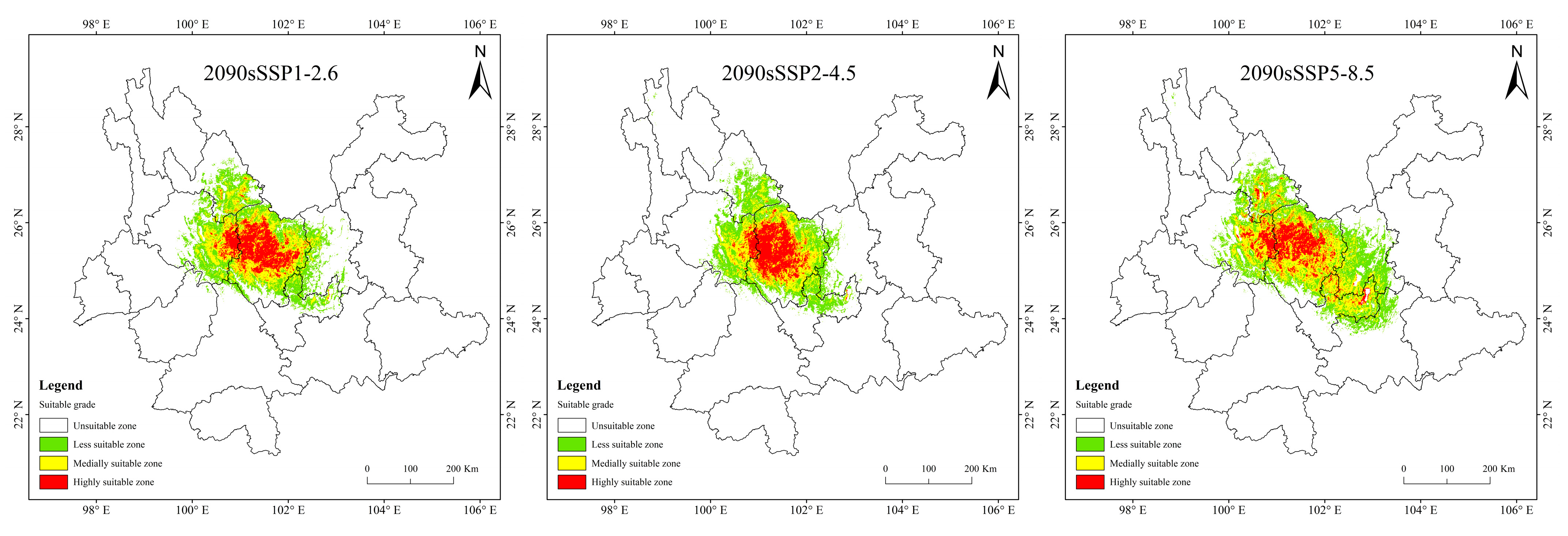
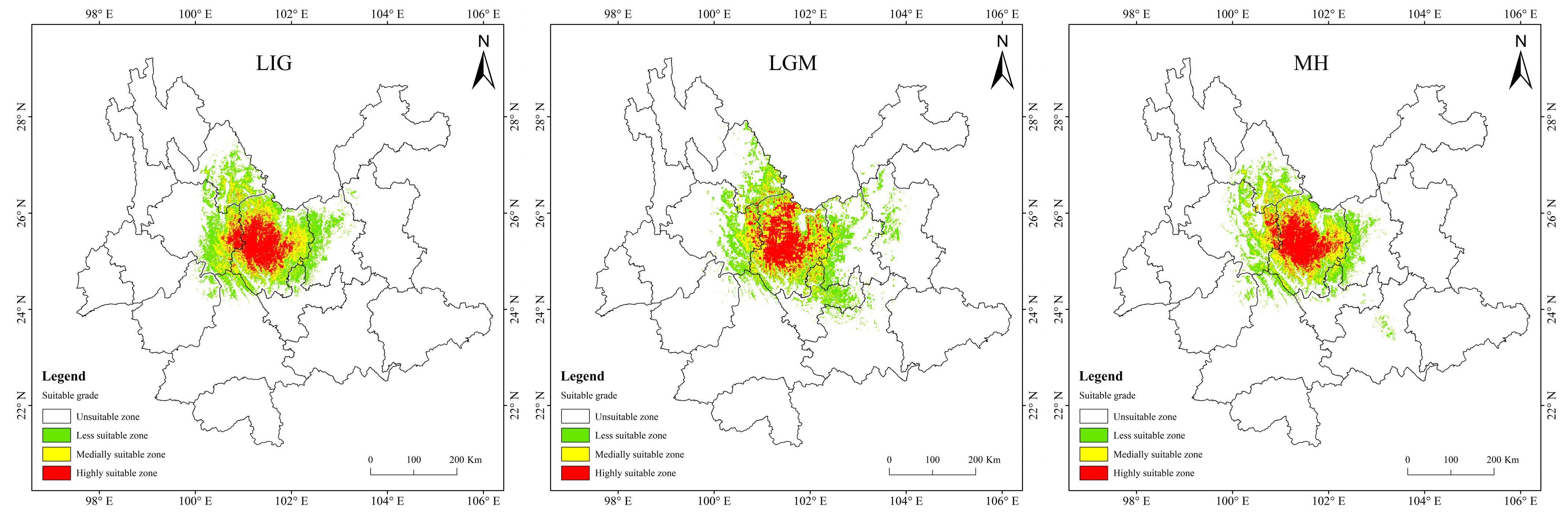
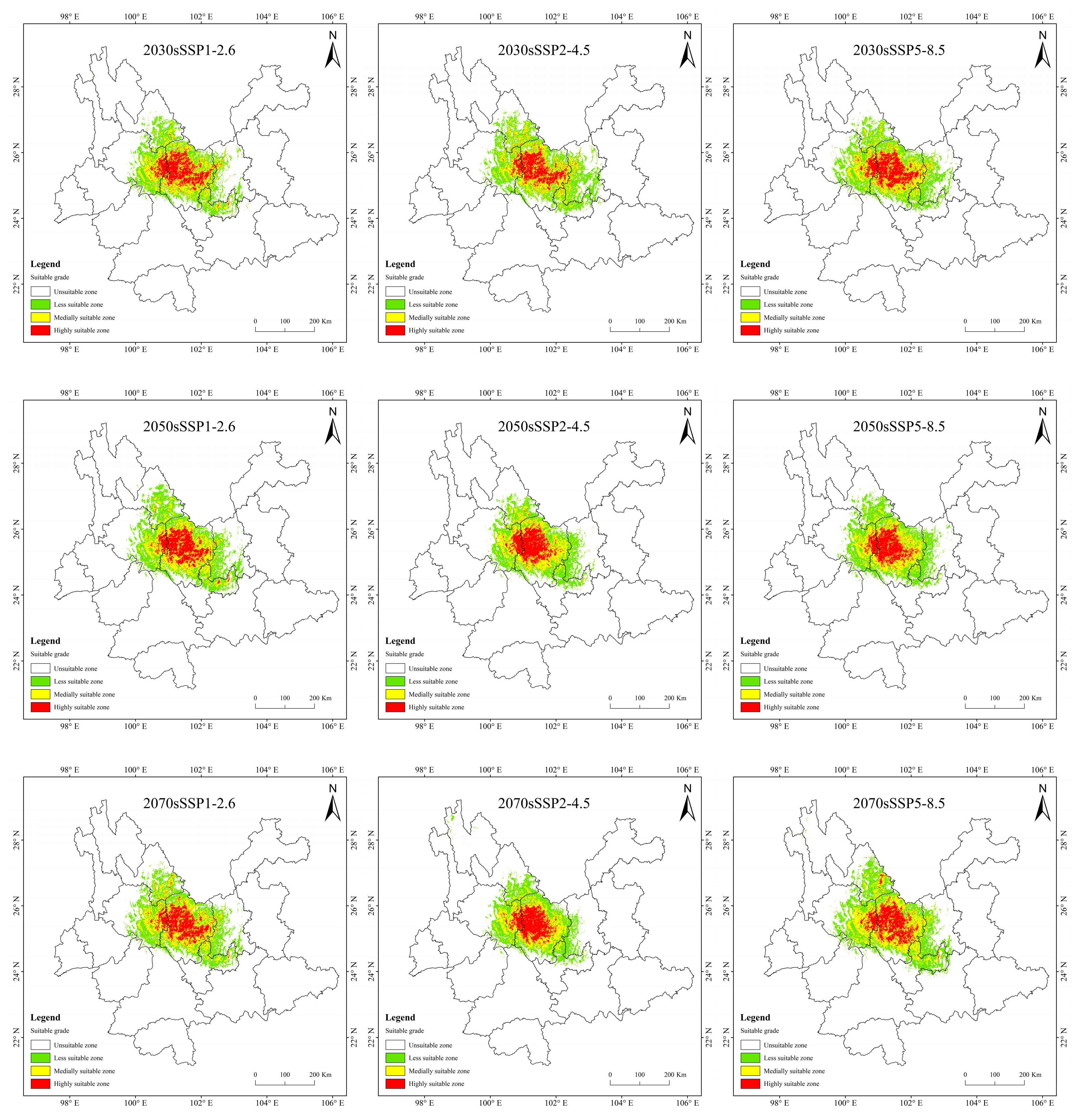
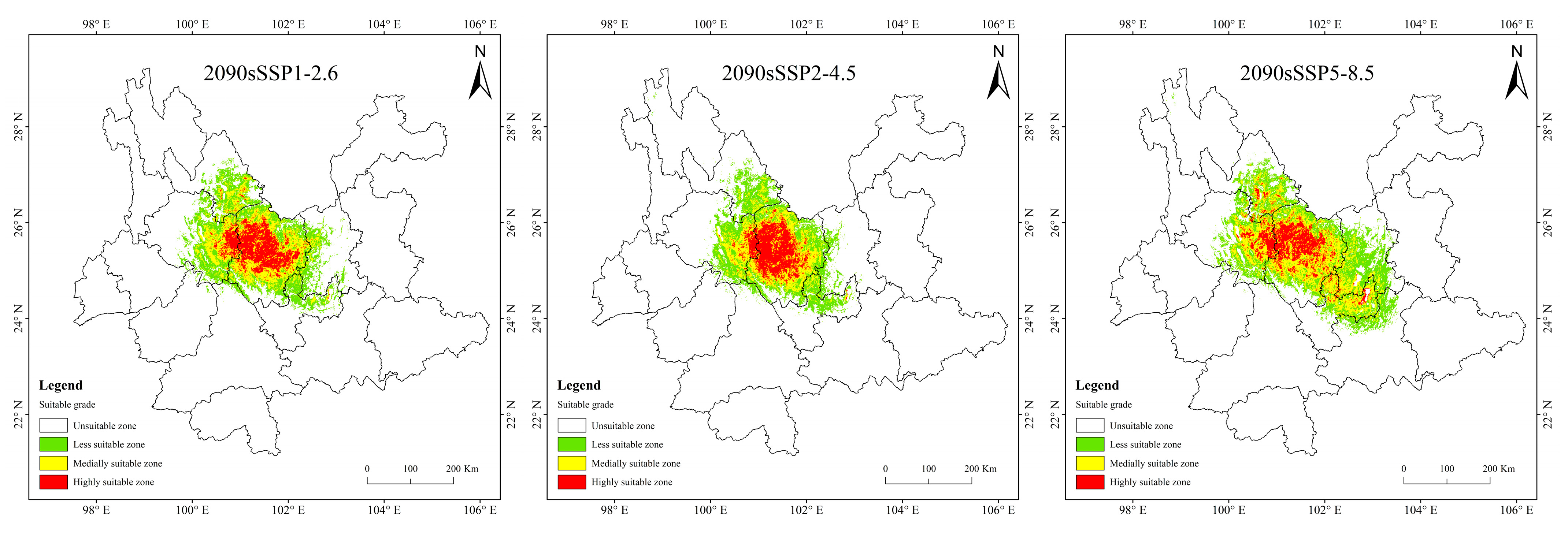
3.4. Potentially Suitable Area for T. nanus under Past and Future Climate Scenario
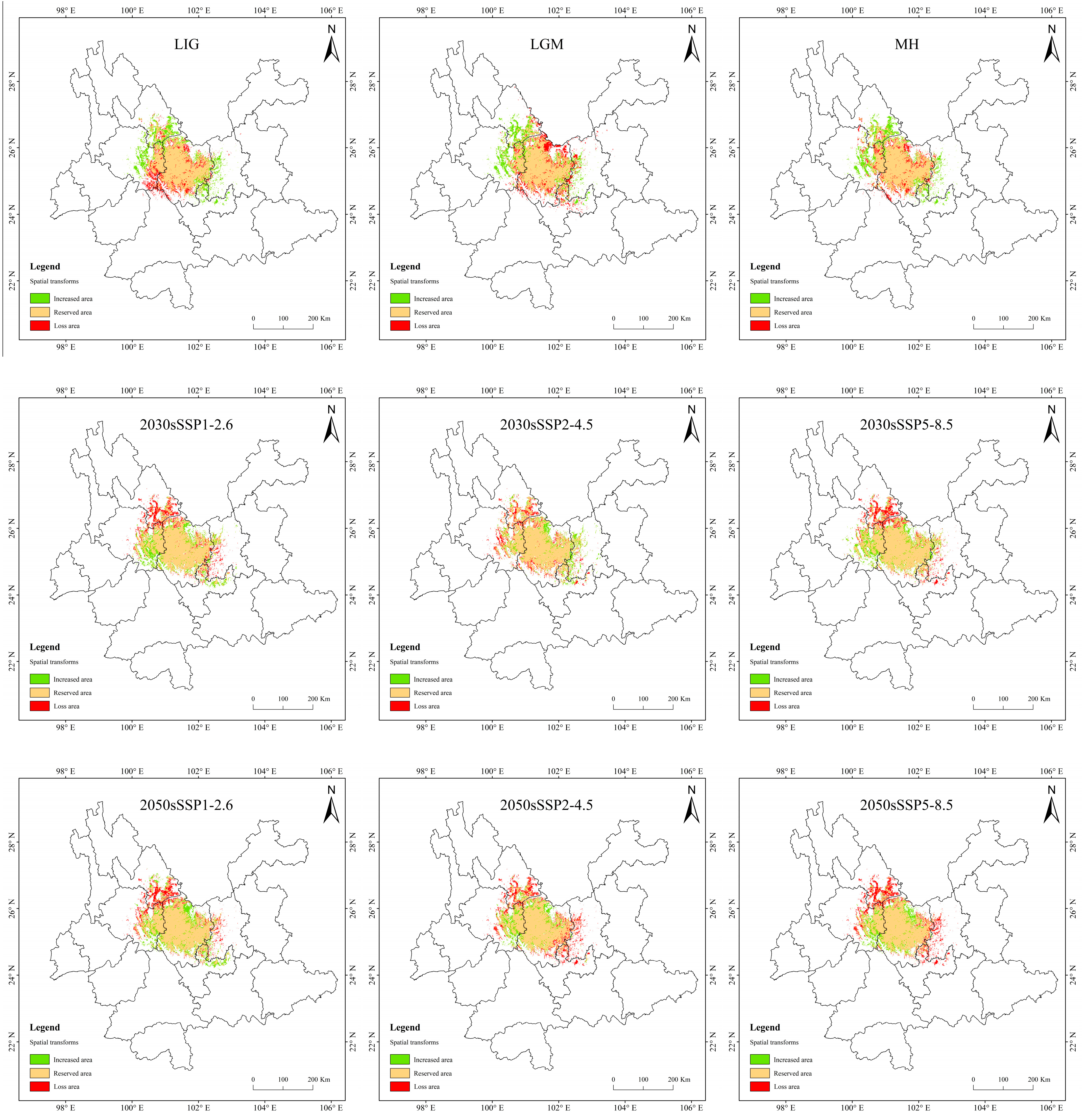
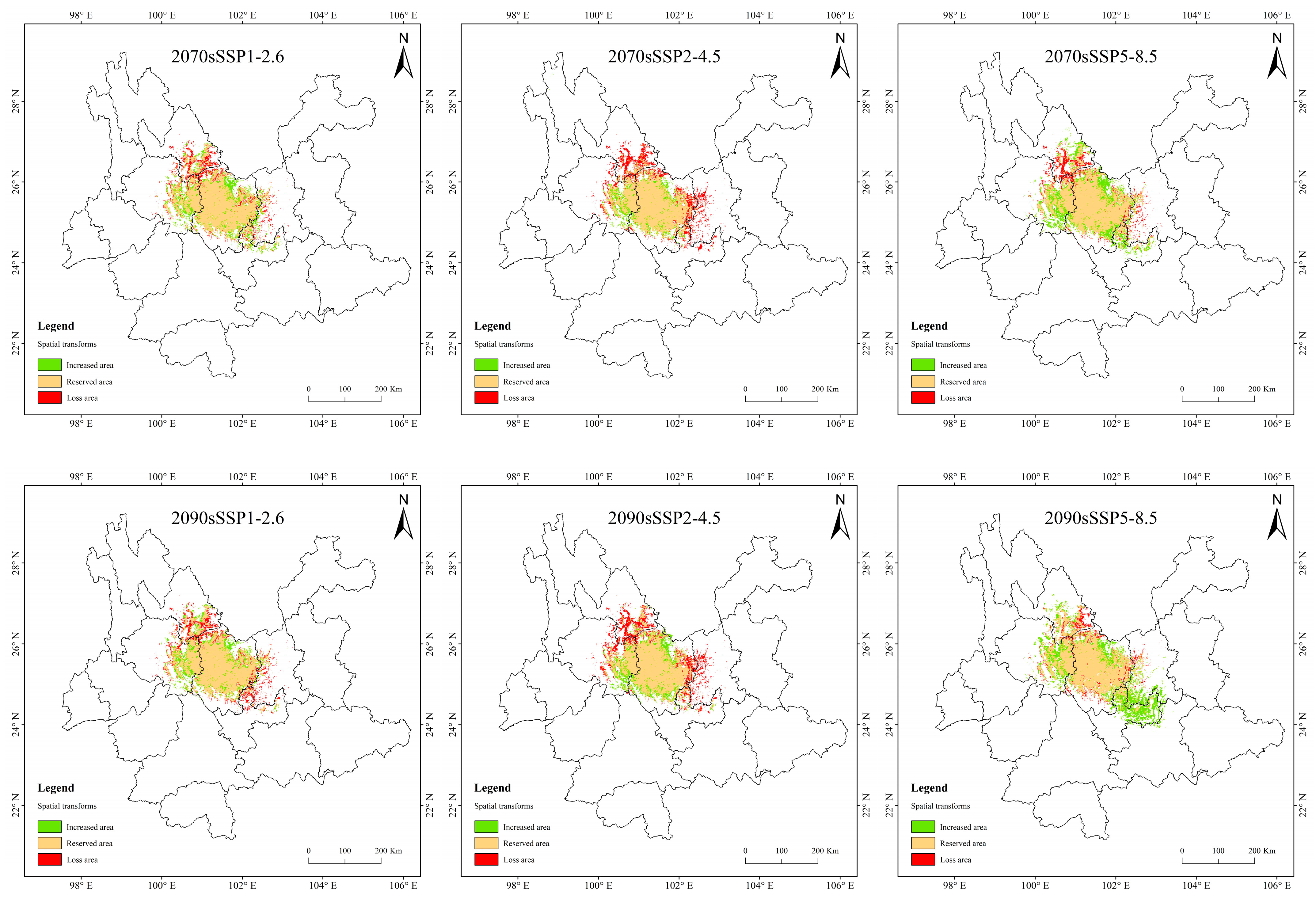
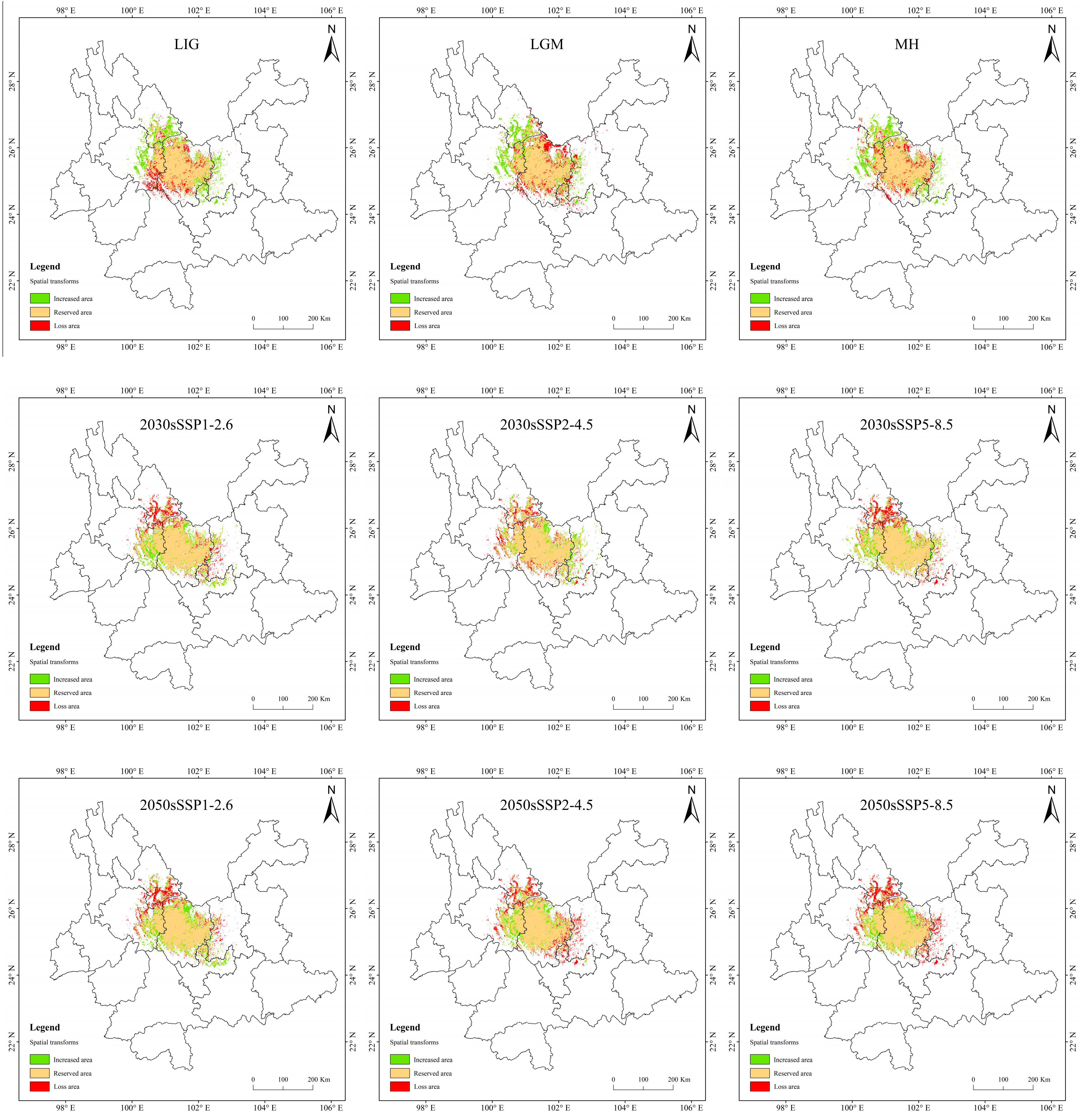
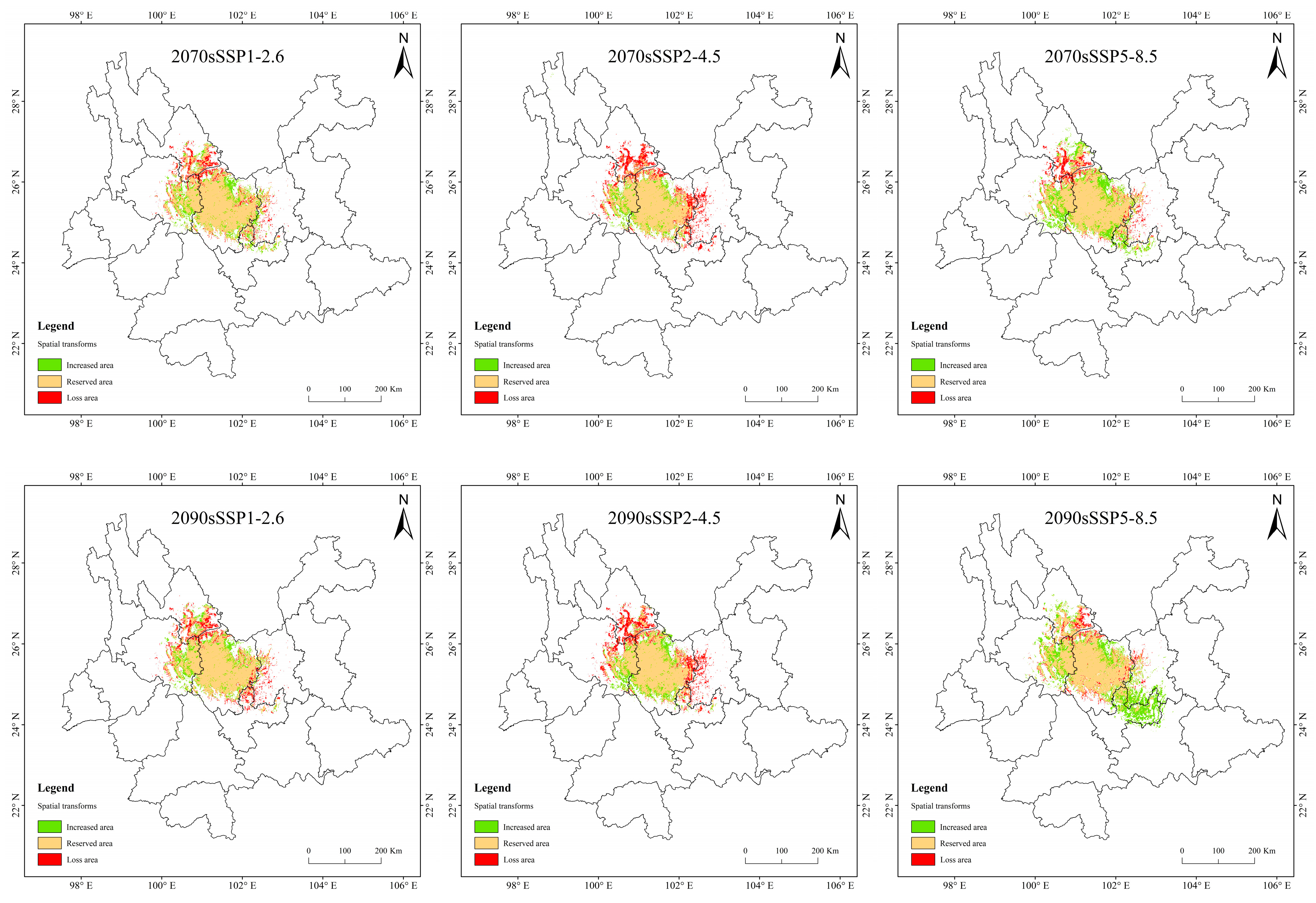
3.5. Spatial Variation Pattern of Potentially Suitable Areas for T. nanus
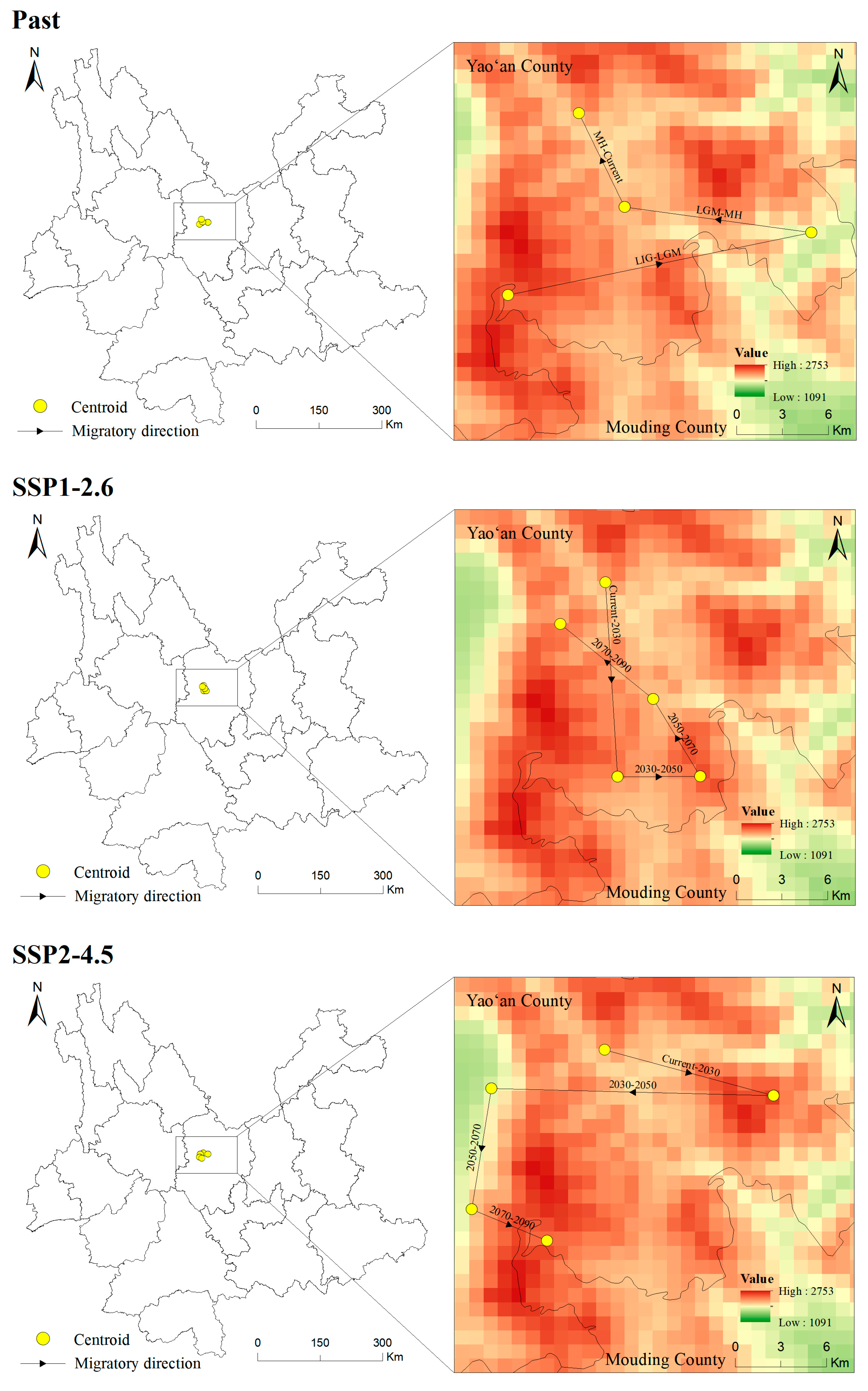
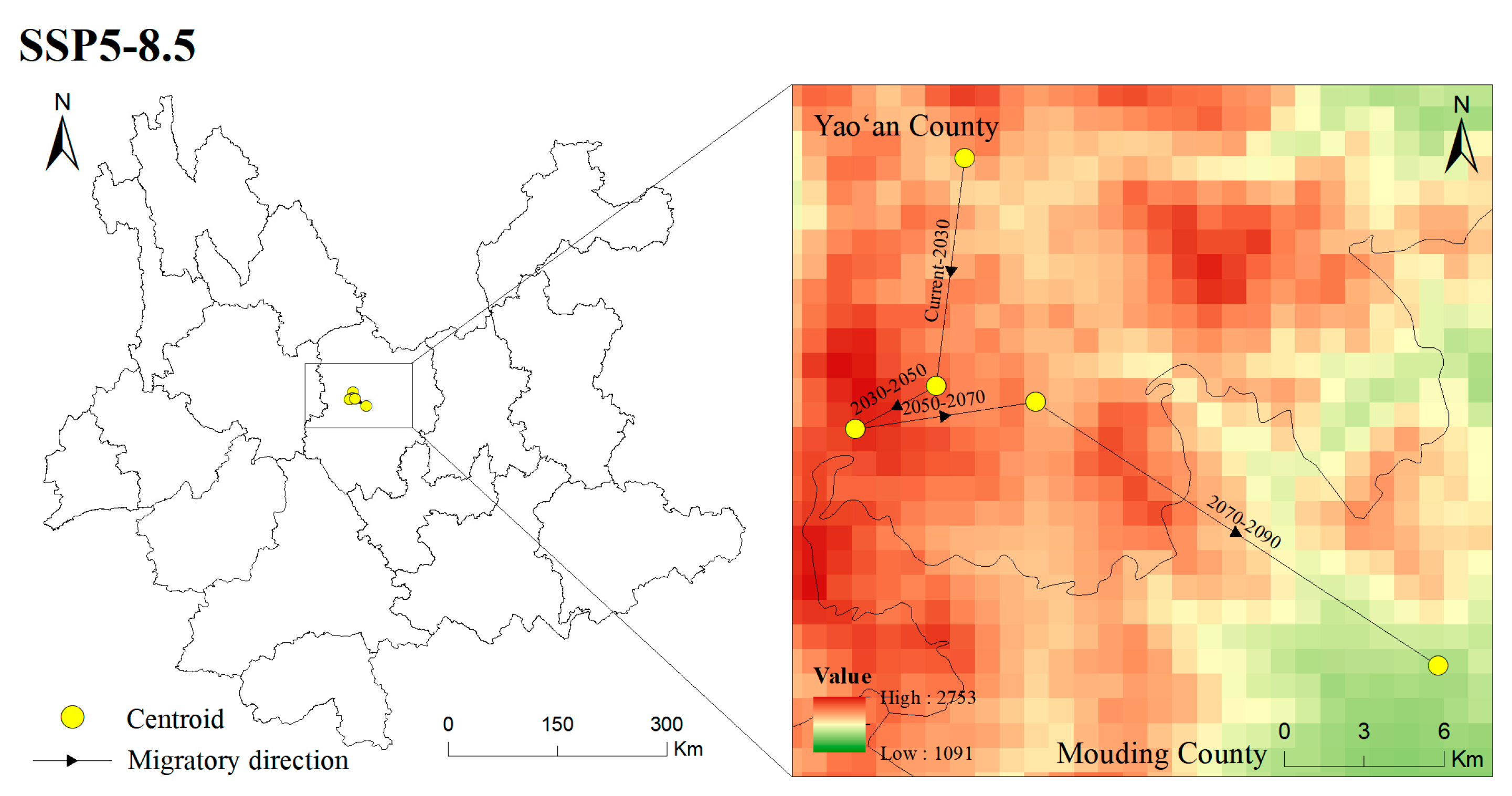
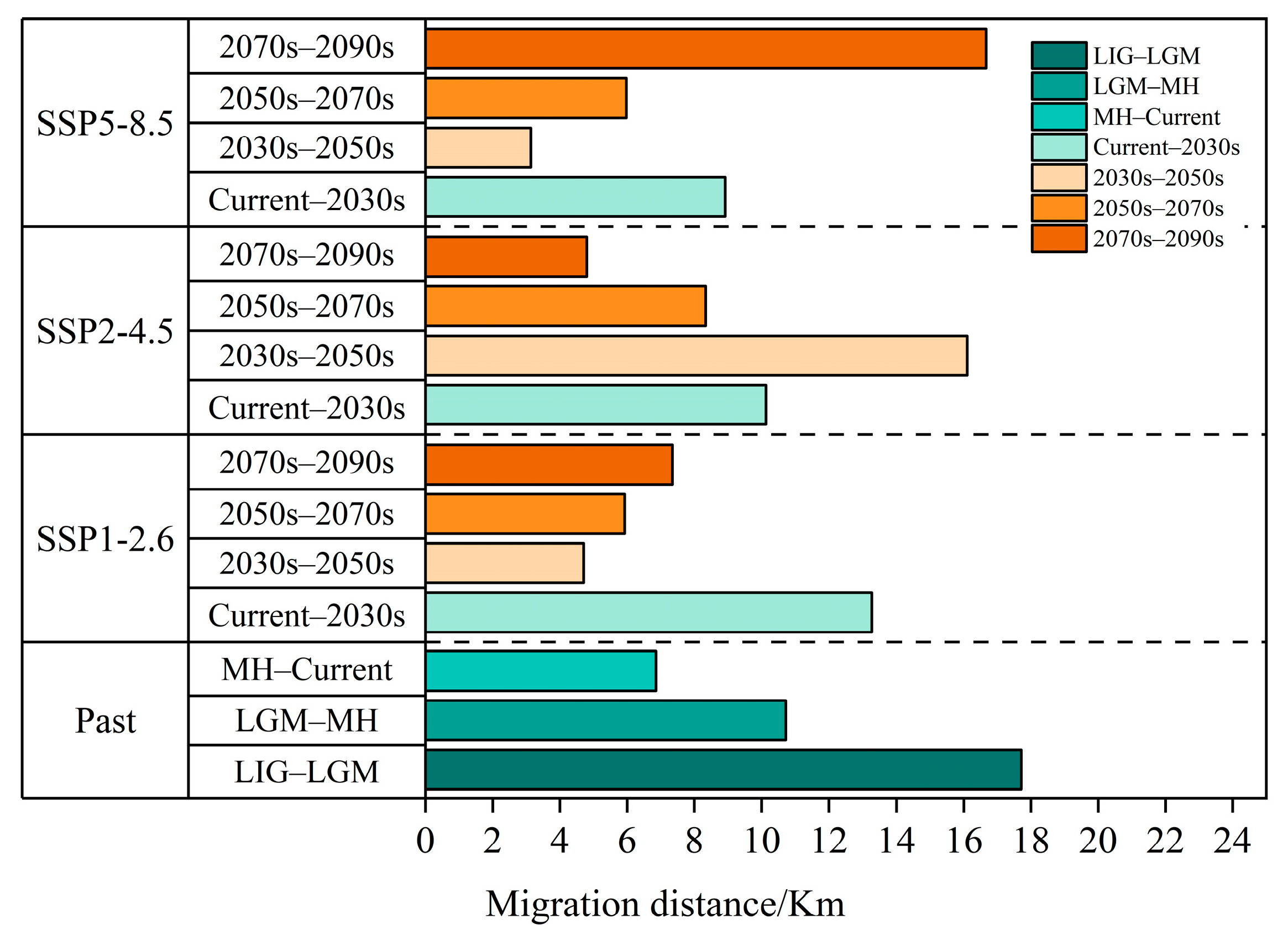
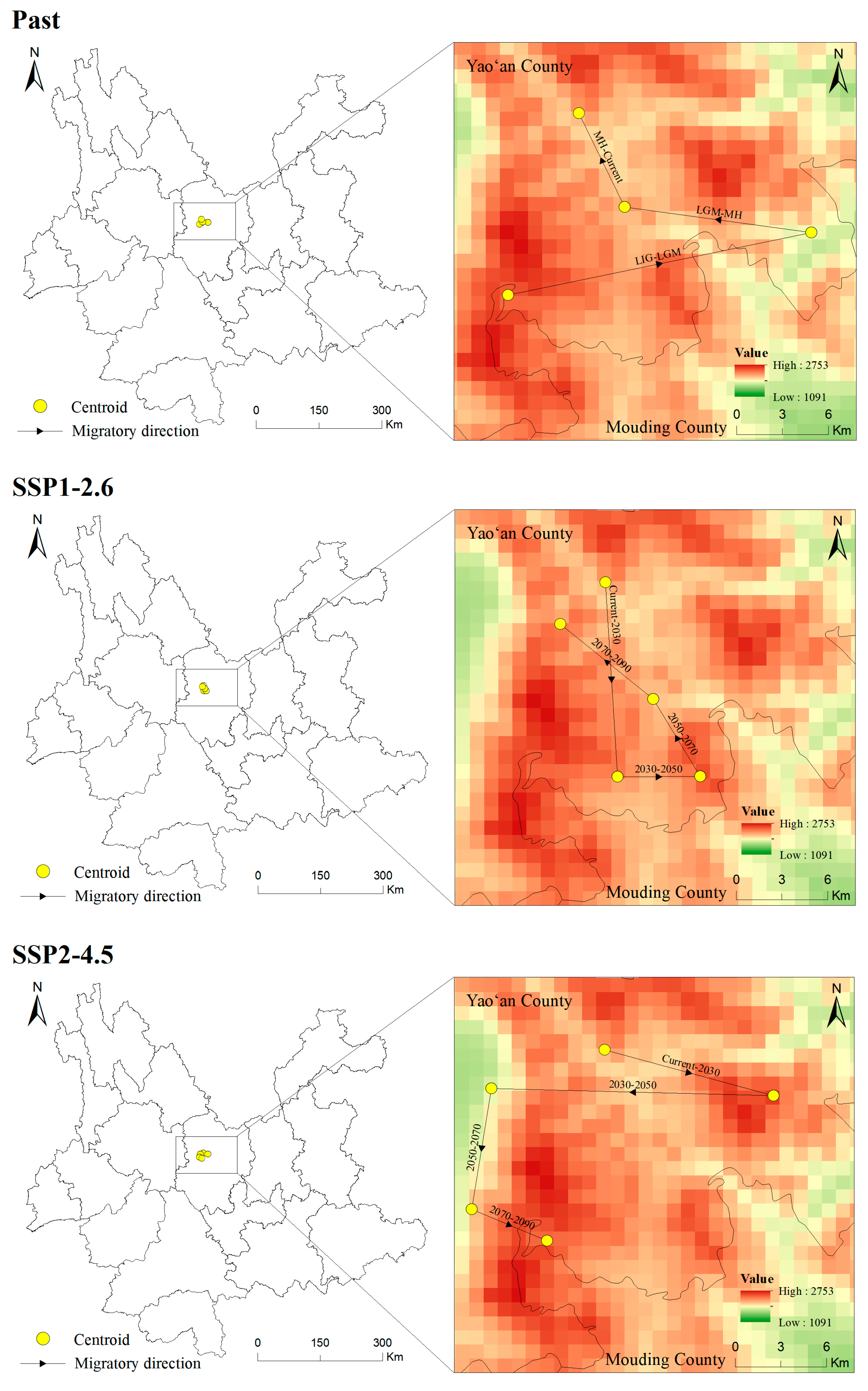
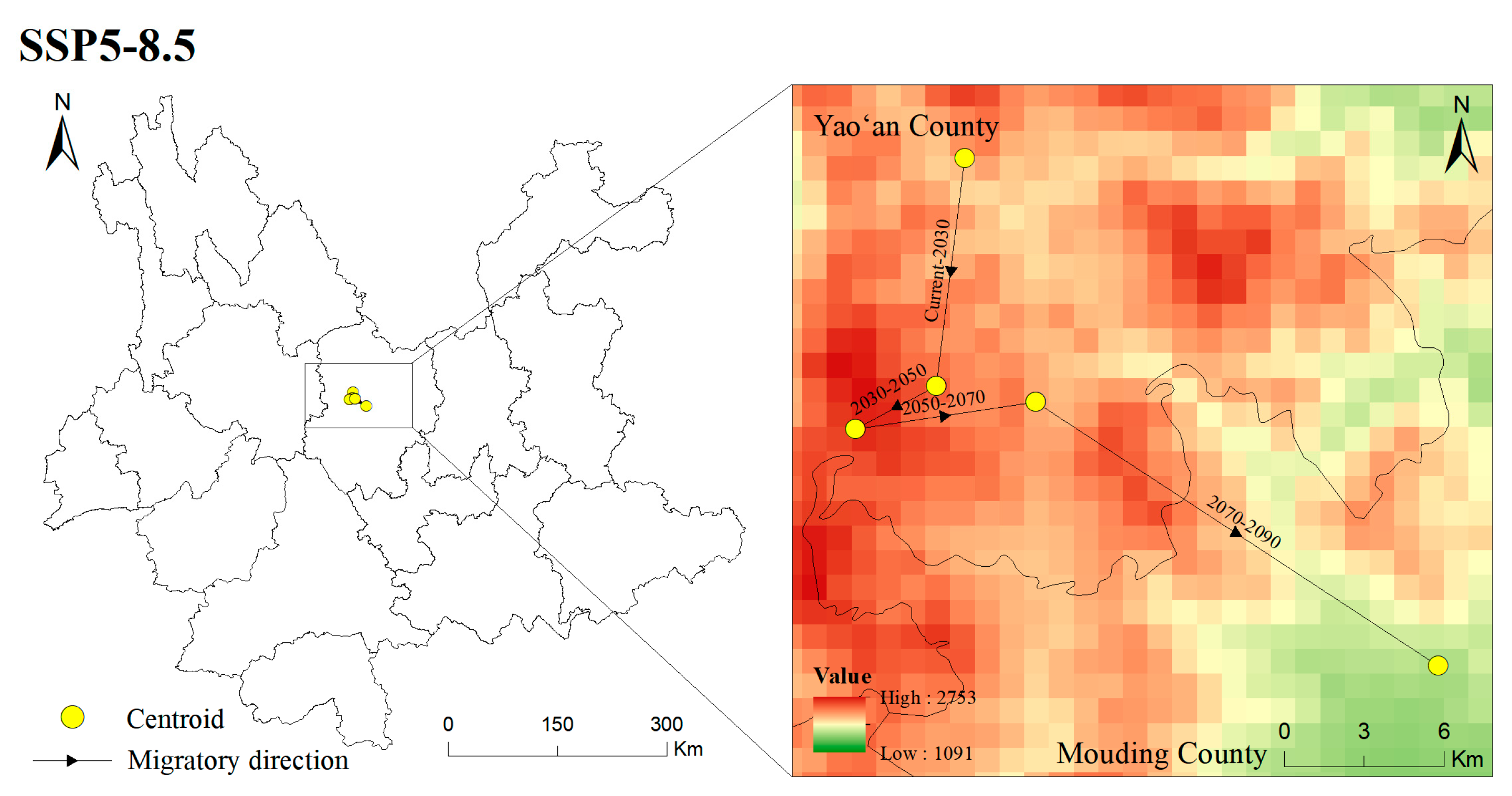
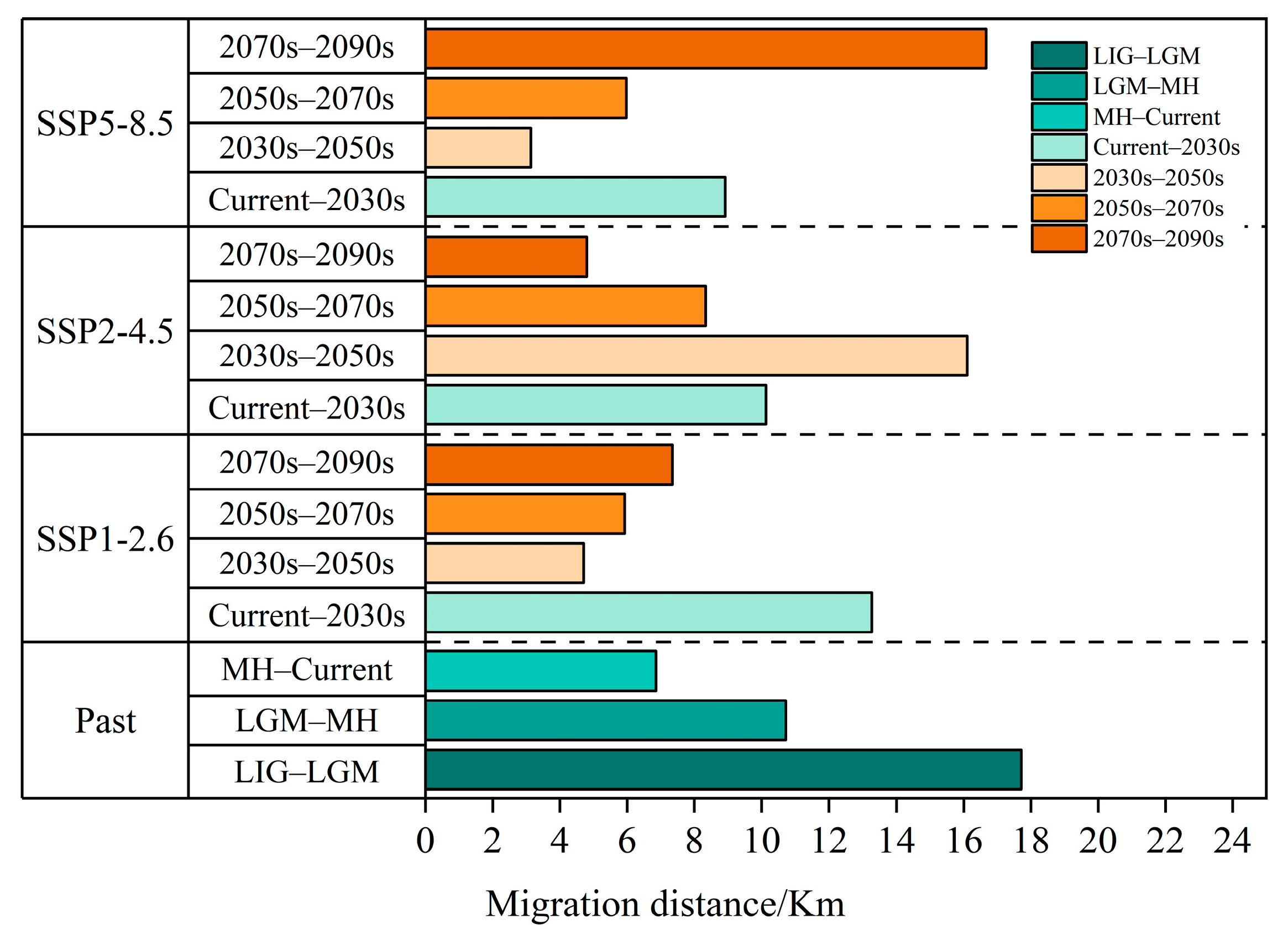
3.6. Centroid Shifts in the Potentially Suitable Areas for T. nanus during Different Periods
4. Discussion
4.1. Model Rationality and Prospects
4.2. Main Environmental Factors Affecting the Distribution of T. nanus
4.3. Spatial Distribution Change in Potentially Suitable Areas for T. nanus
4.4. Centroid Analysis for T. nanus
4.5. Protection Countermeasures and Suggestions for T. nanus
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soberon, J.; Peterson, A.T. Interpretation of models of fundamental ecological niches andspecies’ distributional areas. Biodivers. Inform. 2005, 2, 1–10. [Google Scholar] [CrossRef]
- Alberto, F.J.; Aitken, S.N.; Alía, R.; González-Martínez, S.C.; Hänninen, H.; Kremer, A.; Lefèvre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for evolutionary responses to climate change–evidence from tree populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2004, 359, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Thackeray, C.W.; Hall, A.; Norris, J.; Chen, D. Constraining the increased frequency of global precipitation extremes under warming. Nat. Clim. Chang. 2022, 12, 441–448. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Aleksi, L.; Raimo, V. North by north-west: Climate change and directions of density shifts in birds. Glob. Chang. Biol. 2016, 22, 1121–1129. [Google Scholar] [CrossRef]
- Xu, L.L.; Yu, G.M.; Tu, Z.F.; Zhang, Y.C.; Tsendbazar, N.E. Monitoring vegetation change and their potential drivers in Yangtze River Basin of China from 1982 to 2015. Environ. Monit. Assess. 2020, 192, 642. [Google Scholar] [CrossRef]
- Zhang, Q.; Shen, X.B.; Jiang, X.L.; Fan, T.T.; Liang, X.C.; Yan, W.D. MaxEnt Modeling for Predicting Suitable Habitat for Endangered Tree Keteleeria davidiana (Pinaceae) in China. Forests 2023, 14, 394. [Google Scholar] [CrossRef]
- Samal, P.; Srivastava, J.; Charles, B.; Singarasubramanian, S.R. Species distribution models to predict the potential niche shift and priority conservation areas for mangroves (Rhizophora apiculata, R. mucronata) in response to climate and sea level fluctuations along coastal India. Ecol. Indic. 2023, 154, 110631. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Semwal, D.P.; Pradheep, K.; Ahlawat, S.P. Wild rice (Oryza spp.) germplasm collections from Gangetic Plains and eastern region of India: Diversity mapping and habitat prediction using ecocrop model. Vegetos 2016, 29, 96–100. [Google Scholar] [CrossRef]
- Souza, P.G.C.; Aidoo, O.F.; Farnezi, P.K.B.; Heve, W.K.; Júnior, P.A.S.; Picano, M.C.; Ninsin, K.D.; Ablormeti, F.K.; Shah, M.S.; Siddiqui, S.A.; et al. Tamarixia radiata global distribution to current and future climate using the climate change experiment (CLIMEX) model. Sci. Rep. 2023, 13, 3397. [Google Scholar] [CrossRef]
- Stockman, A.K.; Beamer, D.A.; Bond, J.E. An evaluation of a GARP model as an approach to predicting the spatial distribution of non-vagile invertebrate species. Divers. Distrib. 2006, 12, 81–89. [Google Scholar] [CrossRef]
- Ahmed, S.E.; McInerny, G.; O’Hara, K.; Harper, R.; Salido, L.; Emmott, S.; Joppa, L.N. Scientists and software–surveying the species distribution modelling community. Divers. Distrib. 2015, 21, 258–267. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Du, Z.Y.; He, Y.M.; Wang, H.T.; Wang, C.; Duan, Y.Z. Potential geographical distribution and habitat shift of the genus Ammopiptanthus in China under current and future climate change based on the MaxEnt model. J. Arid. Environ. 2021, 184, 104328. [Google Scholar] [CrossRef]
- Kumar, S.; Stohlgren, T.J. Maxent modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J. Ecol. Nat. Environ. 2009, 1, 94–98. [Google Scholar]
- Qin, A.L.; Liu, B.; Guo, Q.S.; Bussmann, R.W.; Ma, F.Q.; Jian, Z.J.; Xu, G.X.; Pei, S.X. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Zhang, K.L.; Sun, L.P.; Tao, J. Impact of climate change on the distribution of Euscaphis japonica (Staphyleaceae) trees. Forests 2020, 11, 525. [Google Scholar] [CrossRef]
- Ma, Y.P.; Chen, G.; Edward Grumbine, R.; Dao, Z.L.; Sun, W.B.; Guo, H.J. Conserving plant species with extremely small populations (PSESP) in China. Biodivers. Conserv. 2013, 22, 803–809. [Google Scholar] [CrossRef]
- Volis, S. How to conserve threatened Chinese plant species with extremely small populations? Plant Divers. 2016, 38, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Wade, E.M.; Nadarajan, J.; Yang, X.Y.; Ballesteros, D.; Sun, W.B.; Pritchard, H.W. Plant species with extremely small populations (PSESP) in China: A seed and spore biology perspective. Plant Divers. 2016, 38, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.D.; Li, J.H.; Yang, X.X. Surver of Yunnans Trachycarpus nanu and its biological features. Ecol. Sci. 2002, 21, 338–341. [Google Scholar] [CrossRef]
- Gibbons, M.; Spanner, T.W. In search of Trachycarpus nanus. Principes 1993, 37, 64–72. [Google Scholar]
- Pei, S.J.; Chen, S.Y.; Guo, L.X.; John, D.; Andrew, H. Flora of China, Arecaceae; Science Press: Beijing, China, 2013; pp. 145–146. [Google Scholar]
- Xu, C.D.; Dong, X.D. The present situation, the protection and the utilization of natural resources of Trachycarpus nanus in Chuxiong Prefecture. J. Chuxiong Norm. Univ. 1999, 14, 112–115. [Google Scholar]
- Dong, X.D.; Li, J.H.; Yang, X.X.; Su, H.Y. Study on Sexual Reproduction of Trchycarpus nana Becc. Spec. Wild Econ. Anim. Plant Res. 2003, 25, 13–15. [Google Scholar] [CrossRef]
- Xu, X.M.; Zhou, H.Q.; Huang, L.Y.; Liu, R.; Fan, H.K. Protection and utilization of rare and endangered species Trachycarpus nanus. J. Agric. 2010, 53–55. [Google Scholar] [CrossRef]
- Fu, J.B. Research on Yunnan Green Electric Power Development; Yunnan University: Kunming, China, 2013. [Google Scholar]
- Xie, Y.H. Effects of Power Station on Vegetation and Soil Properties; Inner Mongolia Normal University: Hohhot, China, 2015. [Google Scholar]
- Arroyo-Rodríguez, V.; Aguirre, A.; Benítez-Malvido, J.; Mandujano, S. Impact of rain forest fragmentation on the population size of a structurally important palm species: Astrocaryum mexicanum at Los Tuxtlas, Mexico. Biol. Conserv. 2007, 138, 198–206. [Google Scholar] [CrossRef]
- Wei, B.; Liu, L.S.; Gu, C.J.; Yu, H.B.; Zhang, Y.L.; Zhang, B.H.; Cui, B.H.; Gong, D.Q.; Tu, Y.L. The climate niche is stable and the distribution area of Ageratina adenophora is predicted to expand in China. Biodivers. Sci. 2022, 30, 88–99. [Google Scholar] [CrossRef]
- Wang, J.Y. The Wild Landscape and Ornamental Plant Resources of Palmae in China. Chin. Wild Pl. Resour. 2002, 21, 9–11. [Google Scholar]
- Li, P. Conservation Biology of Trachycarpus nanus: A Plant Species with Extremely Small Populations (PSESP) in Southwest Plateau Mountains of China. Master′s Thesis, Yunnan University, Kunming, China, 2020. [Google Scholar]
- Zhou, X.L.; Li, P.; Yang, L.; Long, B.; Wang, Y.H.; Shen, S.K. The complete chloroplast genome sequence of endangered plant Trachycarpus nanus (Arecaceae). Mitochondrial DNA Part B 2021, 6, 1772–1774. [Google Scholar] [CrossRef] [PubMed]
- Xia, K.; Fan, L.; Sun, W.B.; Chen, W.Y. Conservation and fruit biology of Sichou oak (Quercus sichourensis, Fagaceae)—A critically endangered species in China. Plant Divers. 2016, 38, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Ye, X.Z.; Zhang, M.Z.; Lai, W.F.; Yang, M.M.; Fan, H.H.; Zhang, G.F.; Chen, S.P.; Liu, B. Prediction of potential suitable distribution of Phoebe bournei based on MaxEnt optimization model. Acta Ecol. Sin. 2021, 41, 8135–8144. [Google Scholar] [CrossRef]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.; Stevens, B.; Stouffer, R.J.; Taylor, K. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organisation. Geosci. Model. Dev. Discuss. 2015, 8, 10539–10583. [Google Scholar] [CrossRef]
- Wu, T.W.; Lu, Y.X.; Fang, Y.J.; Xin, X.G.; Li, L.; Li, W.P.; Jie, W.H.; Zhang, J.; Liu, Y.M.; Zhang, L.; et al. The Beijing Climate Center Climate System Model (BCC-CSM): The main progress from CMIP5 to CMIP6. Geosci. Model. Dev. 2019, 12, 1573–1600. [Google Scholar] [CrossRef]
- Zhan, P.; Wang, F.Y.; Xia, P.G.; Zhao, G.H.; Wei, M.T.; Wei, F.G.; Han, R.L. Assessment of suitable cultivation region for Panax notoginseng under different climatic conditions using MaxEnt model and high-performance liquid chromatography in China. Ind. Crops Prod. 2022, 176, 114416. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dud’ık, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the 21st International Conference on Machine Learning, Banff, AB, Canada, 4–8 July 2004; ACM Press: New York, NY, USA, 2004; pp. 655–662. [Google Scholar]
- Sillero, N. What does ecological modelling model? A proposed classification of ecological niche models based on their underlying methods. Ecol. Model. 2011, 222, 1343–1346. [Google Scholar] [CrossRef]
- Ranjitkar, S.; Xu, J.C.; Shrestha, K.K.; Kindt, R. Ensemble forecast of climate suitability for the Trans-Himalayan Nyctaginaceae species. Ecol. Model. 2014, 282, 18–24. [Google Scholar] [CrossRef]
- Fotheringham, A.S.; Oshan, T.M. Geographically weighted regression and multicollinearity: Dispelling the myth. J. Geogr. Syst. 2016, 18, 303–329. [Google Scholar] [CrossRef]
- Zadrozny, B. Learning and evaluating classifiers under sample selection bias. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AB, Canada, 4–8 July 2004; pp. 114–121. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods. Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Yuan, Y.D.; Tang, X.G.; Liu, M.Y.; Liu, X.F.; Tao, J. Species distribution models of the Spartina alterniflora Loisel in its origin and invasive country reveal an ecological niche shift. Front. Plant Sci. 2021, 12, 738769. [Google Scholar] [CrossRef]
- Tin, A.; Marten, J.; Kuhns, V.L.H.; Li, Y.; Kttgen, A. Target genes, variants, tissues and transcriptional pathways influencing human serum urate levels. Nat. Genet. 2019, 51, 1459–1474. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, Z.Y.; Abdukeyum, N.; Ling, Y.B. Potential Geographical Distribution of Medicinal Plant Ephedra sinica Stapf under Climate Change. Forests 2022, 13, 2149. [Google Scholar] [CrossRef]
- Zhao, Y.; Deng, X.W.; Xiang, W.H.; Chen, L.; Ouyang, S. Predicting potential suitable habitats of Chinese fir under current and future climatic scenarios based on Maxent model. Ecol. Inf. 2021, 64, 101393. [Google Scholar] [CrossRef]
- Porfirio, L.L.; Harris, R.M.; Lefroy, E.C.; Hugh, S.; Gould, S.F.; Lee, G.; Bindoff, N.L.; Mackey, B. Improving the use of species distribution models in conservation planning and management under climate change. PLoS ONE 2014, 9, e113749. [Google Scholar] [CrossRef] [PubMed]
- Swets, J. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Alfonso-Corrado, C.; Naranjo-Luna, F.; Clark-Tapia, R.; Campos, J.E.; Rojas-Soto, O.R.; Luna-Krauletz, M.D.; Bodenhorn, B.; Gorgonio-Ramírez, M.; Pacheco-Cruz, N. Effects of environmental changes on the occurrence of Oreomunnea mexicana (Juglandaceae) in a biodiversity hotspot cloud forest. Forests 2017, 8, 261. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Wang, W.; Li, Z.J.; Zhang, Y.L.; Xu, X.Q. Current situation, global potential distribution and evolution of six almond species in China. Front. Plant Sci. 2021, 12, 619883. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.H.; Liu, L.; Zhao, L.J.; Cui, S.P. Potentially suitable geographical area for Monochamus alternatus under current and future climatic scenarios based on optimized MaxEnt model. Insects 2023, 14, 182. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.L.; Xiang, J.G.; Li, D.L.; Liu, X.H. Prediction of potential suitable distribution areas of Quasipaa spinosa in China based on MaxEnt optimization model. Biology 2023, 12, 366. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Liu, Y.L.; Qin, H.; Meng, Q.X. Prediction of spatial migration of suitable distribution of Elaeagnus mollis under climate change conditions in Shanxi Province, China. Chin. J. Appl. Ecol. 2019, 30, 496–502. [Google Scholar] [CrossRef]
- Zhao, H.X.; Zhang, H.; Xu, C.G. Study on Taiwania cryptomerioides under climate change: MaxEnt modeling for predicting the potential geographical distribution. Glob. Ecol. Conserv. 2020, 24, e01313. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Wang, H.C.; Chhin, S.; Zhang, J.G. Effects of competition, age and climate on tree slenderness of Chinese fir plantations in southern China. For. Ecol. Manag. 2020, 458, 117815. [Google Scholar] [CrossRef]
- Wang, X.F.; Duan, Y.X.; Jin, L.L.; Wang, C.Y.; Peng, M.C.; Li, Y.; Wang, X.H.; Ma, Y.F. Prediction of historical, present and future distribution of Quercus sect. Heterobalanus based on the optimized MaxEnt model in China. Acta Ecol. Sin. 2023, 43, 6590–6604. [Google Scholar] [CrossRef]
- Wang, Q.; Fan, B.G.; Zhao, G.H. Prediction of potential distribution area of Corylus mandshurica in China under climate change. Chin. J. Ecol. 2020, 39, 3774–3784. [Google Scholar] [CrossRef]
- Guo, Y.X.; Zhang, S.Y.; Tang, S.C.; Pan, J.Y.; Ren, L.H.; Tian, X.; Sun, Z.R.; Zhang, Z.L. Analysis of the prediction of the suitable distribution of Polygonatum kingianum under different climatic conditions based on the MaxEnt model. Front. Earth Sci. 2023, 11, 1111878. [Google Scholar] [CrossRef]
- Kyriazis, C.C.; Wayne, R.K.; Lohmueller, K.E. Strongly deleterious mutations are a primary determinant of extinction risk due to inbreeding depression. Evol. Lett. 2020, 5, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Byeon, D.H.; Jung, S.; Lee, W.H. Review of CLIMEX and MaxEnt for studying species distribution in South Korea. J. Asia-Pac. Biodivers. 2018, 11, 325–333. [Google Scholar] [CrossRef]
- Iverson, L.R.; McKenzie, D. Tree-species range shifts in a changing climate: Detecting, modeling, assisting. Landsc. Ecol. 2013, 28, 879–889. [Google Scholar] [CrossRef]
- Feng, L.; Sun, J.J.; El-Kassaby, Y.A.; Yang, X.Y.; Tian, X.N.; Wang, T.L. Predicting potential habitat of a plant species with small populations under climate change: Ostrya rehderiana. Forests 2022, 13, 129. [Google Scholar] [CrossRef]
- Gao, X.X.; Liu, J.; Huang, Z.H. The impact of climate change on the distribution of rare and endangered tree Firmiana kwangsiensis using the Maxent modeling. Ecol. Evol. 2022, 12, e9165. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Z.; Ye, X.Z.; Li, J.H.; Liu, Y.P.; Chen, S.P.; Liu, B. Prediction of potential suitable area of Ulmus elongata in China under climate change scenarios. Chin. J. Ecol. 2021, 40, 3822–3835. [Google Scholar] [CrossRef]
- Falk, W.; Mellert, K.H. Species distribution models as a tool for forest management planning under climate change: Risk evaluation of Abies alba in Bavaria. J. Veg. Sci. 2011, 22, 621–634. [Google Scholar] [CrossRef]
- Deb, J.C.; Phinn, S.; Butt, N.; McAlpine, C.A. The impact of climate change on the distribution of two threatened Dipterocarp trees. Ecol. Evol. 2017, 7, 2238–2248. [Google Scholar] [CrossRef]
- Root, T. Energy constraints on avian distributions and abundances. Ecology 1988, 69, 330–339. [Google Scholar] [CrossRef]
- Corlett, R.T.; Westcott, D.A. Will plant movements keep up with climate change? Trends Ecol. Evol. 2013, 28, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, J.A.; Thomas, C.D.; Wintle, B.A.; Moilanen, A. Climate change, connectivity and conservation decision making: Back to basics. J. Appl. Ecol. 2009, 46, 964–969. [Google Scholar] [CrossRef]
- Wei, Z.F. The geographic distribution of the Palmae. J. Trop. Subtrop. Bot. 1995, 3, 1–18. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, K.L.; Li, L.F.; Peng, C.; Wang, K. Floral resources and areal-types of palms in Dehong, Yunnan. J. West. China For. Sci. 2013, 42, 70–75. [Google Scholar] [CrossRef]
- Ruan, Z.P.; Tang, Y.J.; Zeng, M.J. Influence of drought stress on photosynthetic characteristics and activity of antioxidant enzymes of four species of Palm seedlings. Chin. J. Trop. Crop. 2016, 37, 1914–1919. [Google Scholar]
- Xu, W.J.; Yuan, T.; Yang, Y.G.; Yu, S.Q. The physiological response to low temperature stress of six palme plants in Nanchang. Acta Agric. Univ. Jiangxiensis 2013, 35, 1212–1216. [Google Scholar] [CrossRef]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Funct. Plant Biol. 2000, 27, 1191. [Google Scholar] [CrossRef]
- Zhang, W.C.; Zheng, J.M.; Ma, T. Temporal and spatial distribution and variation of extreme temperature in Yunnan Province from 1961–2010. Resour. Sci. 2015, 37, 710–722. [Google Scholar]
- Yu, L. Spatio-Temporal Variation of Vegetation EVI in Central Yunnan Under the Background of Climate Aridity. Master′s Thesis, Yunnan University, Kunming, China, 2021. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.J.; Gómez-Aparicio, L.; Lortie, C.J.; Verdú, M.; Cavieres, L.A.; Huang, Z.Y.; Gao, R.R.; Liu, R.; Zhao, Y.L.; Cornelissen, J.H.C. Net plant interactions are highly variable and weakly dependent on climate at the global scale. Ecol. Lett. 2022, 25, 1580–1593. [Google Scholar] [CrossRef] [PubMed]
- Institute of Botany, Chinese Academy of Sciences. Rare and Endangered Plants in China; Shanghai Education Publishing House: Shanghai, China, 1987. [Google Scholar]
- Reich, P.B.; Bermudez, R.; Montgomery, R.A.; Rich, R.L.; Rice, K.E.; Hobbie, S.E.; Stefanski, A. Even modest climate change may lead to major transitions in boreal forests. Nature 2022, 608, 540–545. [Google Scholar] [CrossRef]
- Fu, Y.S.H.; Zhao, H.F.; Piao, S.L.; Peaucelle, M.; Peng, S.S.; Zhou, G.Y.; Ciais, P.; Huang, M.T.; Menzel, A.; Peñuelas, J.; et al. Declining global warming effects on the phenology of spring leaf unfolding. Nature 2015, 526, 104–107. [Google Scholar] [CrossRef]
- Qi, J.Q.; Yu, W.J.; Xie, T.; Ren, M.L. Spatial and temporal variation characteristics of drought disasters in Yun-nan province. Jiangsu J. Agr. Sci. 2019, 35, 631–638. [Google Scholar] [CrossRef]
- Bartlein, P.J.; Harrison, S.; Brewer, S.; Connor, S.; Davis, B.; Gajewski, K.; Guiot, J.; Harrison-Prentice, T.I.; Henderson, A.; Peyron, O. Pollen-based continental climate reconstructions at 6 and 21 ka: A global synthesis. Clim. Dynam. 2011, 37, 775–802. [Google Scholar] [CrossRef]
- Chen, K.; Khine, P.K.; Schneider, H. Diversity and geographical distribution of Pteridophytes in Southern Yunnan. J. Green Sci. Technol. 2021, 23, 120–125. [Google Scholar] [CrossRef]
- Karl, T.R.; Trenberth, K.E. Modern global climate change. Science 2003, 302, 1719–1723. [Google Scholar] [CrossRef]
- Zeke, H. How ‘Shared Socioeconomic Pathways’ explore future climate change. Carbon Brief Clear. Clim. 2018. Available online: https://www.carbonbrief.org/explainer-how-shared-socioeconomic-pathways-explore-future-climate-change/ (accessed on 26 March 2024).
- Deng, L.; Zhu, H.H.; Jiang, Z.H. Projection of climate change in China under carbon neutral scenarios. Trans. Atmos. Sci. 2022, 45, 364–375. [Google Scholar] [CrossRef]
- Yang, X.L.; Zhou, B.T.; Xu, Y.; Han, Z.Y. CMIP6 evaluation and projection of temperature and precipitation over China. Adv. Atmos. Sci. 2021, 38, 817–830. [Google Scholar] [CrossRef]
- Mo, X.G.; Hu, S.; Lu, H.J.; Lin, Z.H.; Liu, S.X. Drought trends over the terrestrial China in the 21st century in climate change scenarios with ensemble GCM projections. J. Nat. Resour. 2018, 33, 1244–1256. [Google Scholar] [CrossRef]
- Su, Z.K.; Li, W.X.; Zhang, J.Y.; Jin, J.L.; Xue, Q.; Wang, Y.T.; Wang, G.Q. Historical changes and future trends of extreme precipitation and high temperature in China. Strateg. Study CAE 2022, 24, 116–125. [Google Scholar] [CrossRef]
- Osman, M.B.; Tierney, J.E.; Zhu, J.; Tardif, R.; Hakim, G.J.; King, J.; Poulsen, C.J. Globally resolved surface temperatures since the Last Glacial Maximum. Nature 2021, 599, 239–244. [Google Scholar] [CrossRef]
- Turney, C.S.M.; Jones, R.T. Does the Agulhas Current amplify global temperatures during super-interglacials? J. Quat. Sci. 2010, 25, 839–843. [Google Scholar] [CrossRef]
- Hertzberg, J. A seasonal solution to a palaeoclimate puzzle. Nature 2021, 589, 521–522. [Google Scholar] [CrossRef]
- Park, H.S.; Kim, S.J.; Stewart, A.L.; Son, S.W.; Seo, K.H. Mid-holocene Northern Hemisphere warming driven by Arctic amplification. Sci. Adv. 2019, 5, eaax8203. [Google Scholar] [CrossRef]
- Fang, J.Y.; Zhu, J.L.; Shi, Y. The responses of ecosystems to global warming. Chin. Sci. Bull. 2018, 63, 136–140. [Google Scholar] [CrossRef]
- Loarie, S.R.; Carter, B.E.; Hayhoe, K.; McMahon, S.; Moe, R.; Knight, C.A.; Ackerly, D.D. Climate change and the future of California’s endemic flora. PLoS ONE 2008, 3, e2502. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.R.; Zhang, L.; Zhang, J.B.; He, L.R.; Sun, R.X. Maxent modeling for predicting the potential geographical distribution of Castanopsis carlesii under various climate change scenarios in China. Forests 2023, 14, 1397. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; De Siqueira, M.F.; Grainger, A.; Hannah, L. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, M.C.; Gove, A.D.; Sanders, N.J.; Dunn, R.R. Climate change, plant migration, and range collapse in a global biodiversity hotspot: The Banksia (Proteaceae) of Western Australia. Glob. Chang. Biol. 2008, 14, 1337–1352. [Google Scholar] [CrossRef]
- Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to Quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Chave, J. Neutral theory and community ecology. Ecol. Lett. 2004, 7, 241–253. [Google Scholar] [CrossRef]
- Hubbell, S.P. Neutral theory in community ecology and the hypothesis of functional equivalence. Funct. Ecol. 2005, 19, 166–172. [Google Scholar] [CrossRef]
- Malcolm, J.R.; Markham, A.; Neilson, R.P.; Garaci, M. Estimated migration rates under scenarios of global climate change. J. Biogeogr. 2002, 29, 835–849. [Google Scholar] [CrossRef]
- Iverson, L.R.; Schwartz, M.W.; Prasad, A.M. How fast and far might tree species migrate in the eastern United States due to climate change? Glob. Ecol. Biogeogr. 2004, 13, 209–219. [Google Scholar] [CrossRef]
- Etterson, J.R.; Shaw, R.G. Constraint to adaptive evolution in response to global warming. Science 2001, 294, 151–154. [Google Scholar] [CrossRef]
- Pitelka, L.F.; Plant Migration Workshop Group. Plant migration and climate change: A more realistic portrait of plant migration is essential to predicting biological responses to global warming in a world drastically altered by human activity. Am. Sci. 1997, 85, 464–473. [Google Scholar]
- Tucker, M.A.; Böhning-Gaese, K.; Fagan, W.F.; Fryxell, J.M.; Van Moorter, B.; Alberts, S.C.; Ali, A.H.; Allen, A.M.; Attias, N.; Avgar, T. Moving in the Anthropocene: Global reductions in terrestrial mammalian movements. Science 2018, 359, 466–469. [Google Scholar] [CrossRef]
- Smit, B.; Wandel, J. Adaptation, adaptive capacity and vulnerability. Glob. Environ. Chang. 2006, 16, 282–292. [Google Scholar] [CrossRef]
- Lv, J.J. The Impacts of Climate Change on the Distribution of Rare or Endangered Species in China and Adaptation Strategies; Chinese Research Academy of Environmental Sciences: Beijing, China, 2009. [Google Scholar]
- Xiao, H.W.; Huang, Y.B.; Wei, Y.K. Successful ex situ conservation of Salvia daiguii. Oryx 2022, 56, 650–651. [Google Scholar] [CrossRef]
- Patwardhan, A.; Pimputkar, M.; Mhaskar, M.; Agarwal, P.; Barve, N.; Gunaga, R.; Mirgal, A.; Salunkhe, C.; Vasudeva, R. Distribution and population status of threatened medicinal tree Saraca asoca (Roxb.) De Wilde from Sahyadri–Konkan ecological corridor. Curr. Sci. 2016, 111, 1500–1506. [Google Scholar] [CrossRef]
- Wang, M.Q.; Wang, Z.J. Ecological integrity conservation and management of group of national parks of Yellowstone to Yukon in North America. Landsc. Archit. 2021, 28, 113–118. [Google Scholar] [CrossRef]
- Fricke, E.C.; Ordonez, A.; Rogers, H.S.; Svenning, J.C. The effects of defaunation on plants’ capacity to track climate change. Science 2022, 375, 210–214. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description | Unit | Percent Contribution/% | Permutation Importance/% |
|---|---|---|---|---|
| bio12 | Annual precipitation | mm | 55.1 | 34.4 |
| bio4 | Temperature seasonality | - | 11.7 | 15.0 |
| bio11 | Mean temperature of coldest quarter | °C | 7.6 | 0.2 |
| bio15 | Precipitation seasonality | - | 5.2 | 0.5 |
| bio14 | Precipitation of driest month | mm | 3.6 | 3.8 |
| Alt | Elevation | m | 2.7 | 13.1 |
| asp | Aspect | ° | 2.5 | 1.1 |
| Slo | Slope | ° | 2.3 | 0.9 |
| Landuse | Land use type | - | 1.6 | 0.6 |
| bio3 | Isothermality | - | 1.2 | 7.4 |
| bio1 | Annual mean temperature | °C | 1.1 | 12.9 |
| bio16 | Precipitation of wettest quarter | mm | 1.1 | 1.3 |
| bio9 | Mean temperature of driest quarter | °C | 0.8 | 2.8 |
| bio7 | Temperature annual range | °C | 0.7 | 2.8 |
| bio6 | Min temperature of coldest month | °C | 0.7 | 2.5 |
| bio13 | Precipitation of wettest month | mm | 0.6 | 0.1 |
| bio18 | Precipitation of warmest quarter | mm | 0.3 | 0.3 |
| bio17 | Precipitation of driest quarter | mm | 0.3 | 0.0 |
| bio2 | Mean diurnal range | °C | 0.2 | 0.0 |
| bio8 | Mean temperature of wettest quarter | °C | 0.2 | 0.1 |
| bio19 | Precipitation of coldest quarter | mm | 0.1 | 0.1 |
| bio10 | Mean temperature of warmest quarter | °C | 0.1 | 0.0 |
| bio5 | Max temperature of warmest month | °C | 0.1 | 0.5 |
| Variable | Description | Unit | Percent Contribution/% | Permutation Importance/% |
|---|---|---|---|---|
| bio12 | Annual precipitation | mm | 48.9 | 57.2 |
| bio11 | Mean temperature of coldest quarter | °C | 11.3 | 1.3 |
| bio4 | Temperature seasonality | - | 9.6 | 24.8 |
| bio16 | Precipitation of wettest quarter | mm | 8.0 | 0.5 |
| bio3 | Isothermality | - | 7.1 | 5.4 |
| bio2 | Mean diurnal range | °C | 5.3 | 2.5 |
| bio15 | Precipitation seasonality | - | 4.9 | 1.9 |
| bio1 | Annual mean temperature | °C | 1.1 | 0.4 |
| asp | Aspect | ° | 1.1 | 0.5 |
| bio14 | Precipitation of driest month | mm | 0.9 | 4.5 |
| slope | Slope | ° | 0.9 | 0.6 |
| Land use | Land use type | - | 0.8 | 0.3 |
| Type | RM | FC | Delta. AICc | Avg. Diff. AUC | Mean. OR10 |
|---|---|---|---|---|---|
| Default | 1 | LQHPT | 153.875 | 0.038 | 0.292 |
| Optimized | 1 | LQ | 0 | 0.031 | 0.115 |
| Period | Less Suitable Area | Medially Suitable Area | Highly Suitable Area | Total Suitable Area |
|---|---|---|---|---|
| LIG | 2.43 | 1.43 | 1.09 | 4.95 |
| LGM | 2.87 | 1.59 | 1.15 | 5.61 |
| MH | 2.16 | 1.37 | 1.11 | 4.64 |
| Current | 2.86 | 1.79 | 1.00 | 5.65 |
| 2030s-SSP1-2.6 | 2.96 | 1.83 | 1.12 | 5.91 |
| 2030s-SSP2-4.5 | 2.46 | 1.57 | 1.07 | 5.10 |
| 2030s-SSP5-8.5 | 2.67 | 1.58 | 1.04 | 5.29 |
| 2050s-SSP1-2.6 | 2.56 | 1.76 | 1.23 | 5.45 |
| 2050s-SSP2-4.5 | 2.96 | 1.82 | 1.01 | 5.79 |
| 2050s-SSP5-8.5 | 2.75 | 1.71 | 1.15 | 5.61 |
| 2070s-SSP1-2.6 | 2.77 | 1.92 | 1.12 | 5.81 |
| 2070s-SSP2-4.5 | 2.38 | 1.43 | 1.06 | 4.87 |
| 2070s-SSP5-8.5 | 3.12 | 2.16 | 1.21 | 6.49 |
| 2090s-SSP1-2.6 | 2.61 | 1.74 | 1.13 | 5.48 |
| 2090s-SSP2-4.5 | 2.48 | 1.54 | 1.13 | 5.15 |
| 2090s-SSP5-8.5 | 3.29 | 2.63 | 1.16 | 7.08 |
| Period | Area/×104 km2 | Change Rate/% | ||||||
|---|---|---|---|---|---|---|---|---|
| Increase | Reserved | Lost | Change | Increase | Reserved | Lost | Change | |
| LIG | 0.81 | 1.99 | 0.54 | 0.27 | 14.34 | 35.22 | 9.56 | 4.78 |
| LGM | 0.79 | 2.03 | 0.72 | 0.07 | 13.98 | 35.93 | 12.74 | 1.24 |
| MH | 0.77 | 2.03 | 0.45 | 0.32 | 13.63 | 35.93 | 7.96 | 5.66 |
| 2030s-SSP1-2.6 | 0.63 | 2.26 | 0.54 | 0.09 | 11.15 | 40.00 | 9.56 | 1.59 |
| 2030s-SSP2-4.5 | 0.50 | 2.34 | 0.46 | 0.04 | 8.85 | 41.42 | 8.14 | 0.71 |
| 2030s-SSP5-8.5 | 0.58 | 2.28 | 0.51 | 0.07 | 10.26 | 40.35 | 9.03 | 1.24 |
| 2050s-SSP1-2.6 | 0.68 | 2.27 | 0.53 | 0.15 | 12.04 | 40.18 | 9.38 | 2.66 |
| 2050s-SSP2-4.5 | 0.47 | 2.18 | 0.61 | −0.14 | 8.32 | 38.58 | 10.80 | −2.48 |
| 2050s-SSP5-8.5 | 0.49 | 2.12 | 0.67 | −0.18 | 8.67 | 37.52 | 11.86 | −3.19 |
| 2070s-SSP1-2.6 | 0.70 | 2.34 | 0.45 | 0.25 | 12.39 | 41.42 | 7.96 | 4.42 |
| 2070s-SSP2-4.5 | 0.42 | 2.06 | 0.73 | −0.31 | 7.43 | 36.46 | 12.92 | −5.49 |
| 2070s-SSP5-8.5 | 1.02 | 2.36 | 0.44 | 0.58 | 18.05 | 41.77 | 7.79 | 10.27 |
| 2090s-SSP1-2.6 | 0.61 | 2.27 | 0.53 | 0.08 | 10.80 | 40.18 | 9.38 | 1.42 |
| 2090s-SSP2-4.5 | 0.59 | 2.07 | 0.73 | −0.14 | 10.44 | 36.64 | 12.92 | −2.48 |
| 2090s-SSP5-8.5 | 1.32 | 2.47 | 0.32 | 1.00 | 23.36 | 43.72 | 5.66 | 17.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wang, X.; Li, Y.; Wu, C.; Zhao, B.; Peng, M.; Chen, W.; Wang, C. Response of Extremely Small Populations to Climate Change—A Case of Trachycarpus nanus in Yunnan, China. Biology 2024, 13, 240. https://doi.org/10.3390/biology13040240
Wang X, Wang X, Li Y, Wu C, Zhao B, Peng M, Chen W, Wang C. Response of Extremely Small Populations to Climate Change—A Case of Trachycarpus nanus in Yunnan, China. Biology. 2024; 13(4):240. https://doi.org/10.3390/biology13040240
Chicago/Turabian StyleWang, Xiaofan, Xuhong Wang, Yun Li, Changhao Wu, Biao Zhao, Mingchun Peng, Wen Chen, and Chongyun Wang. 2024. "Response of Extremely Small Populations to Climate Change—A Case of Trachycarpus nanus in Yunnan, China" Biology 13, no. 4: 240. https://doi.org/10.3390/biology13040240
APA StyleWang, X., Wang, X., Li, Y., Wu, C., Zhao, B., Peng, M., Chen, W., & Wang, C. (2024). Response of Extremely Small Populations to Climate Change—A Case of Trachycarpus nanus in Yunnan, China. Biology, 13(4), 240. https://doi.org/10.3390/biology13040240