
Evidence for Selection on Mitochondrial OXPHOS Genes in the Mediterranean Killifish Aphanius fasciatus Valenciennes, 1821


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
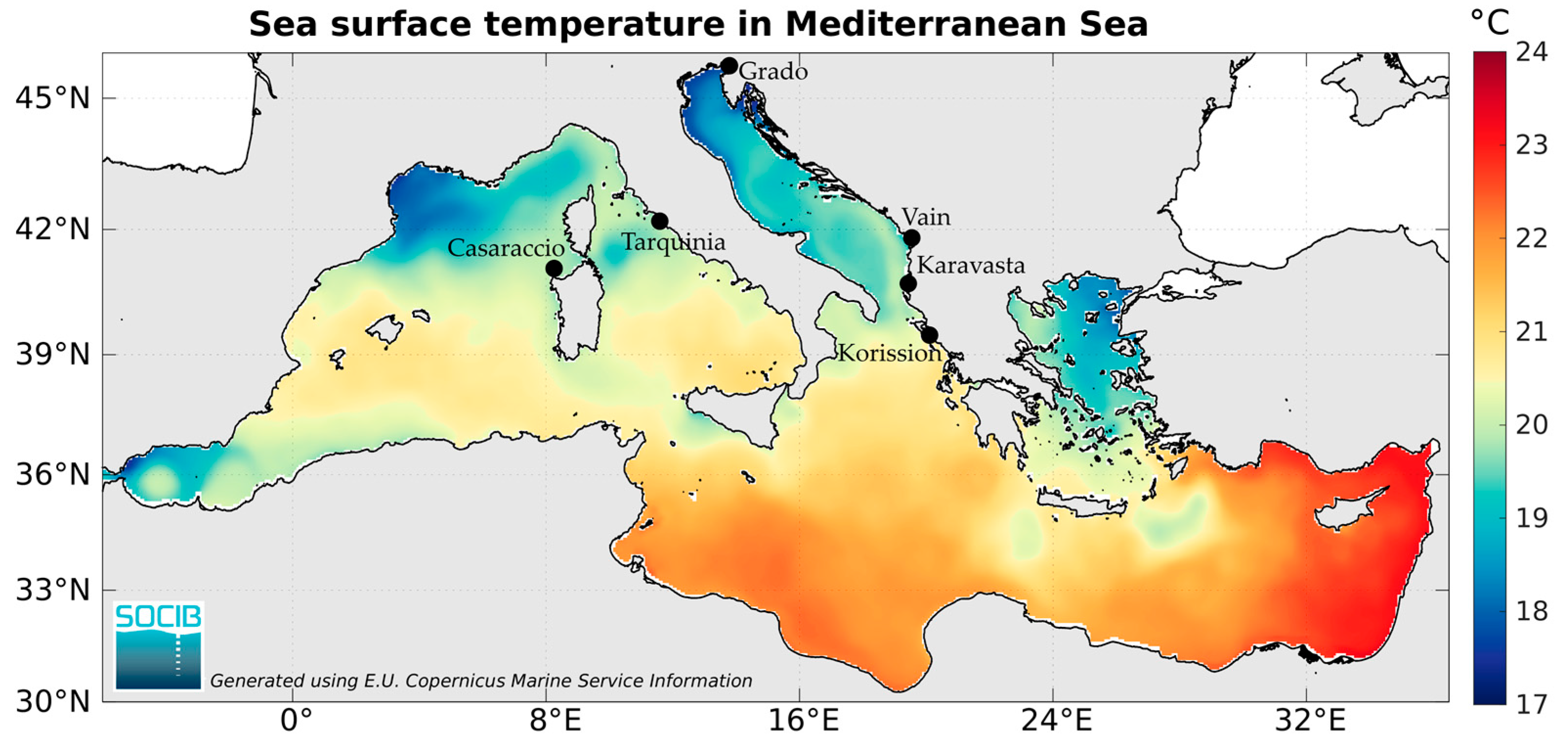
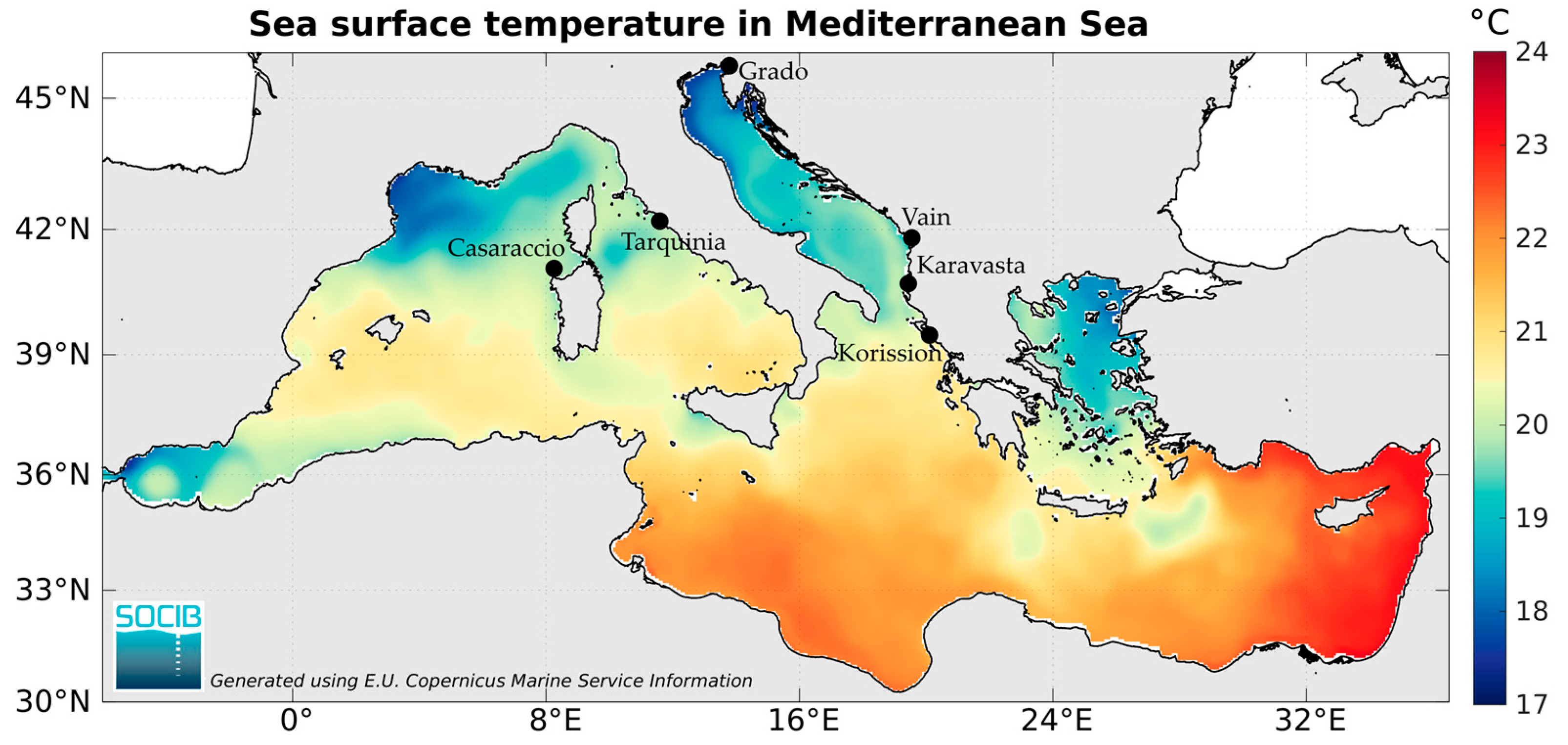
2.1. Sampling Sites
2.2. DNA Extraction
2.3. Amplification and Sequencing of the OXPHOS Target Genes
2.4. Data Analysis
2.5. Recombination and Selection Tests
2.6. Protein Structure Analysis
3. Results
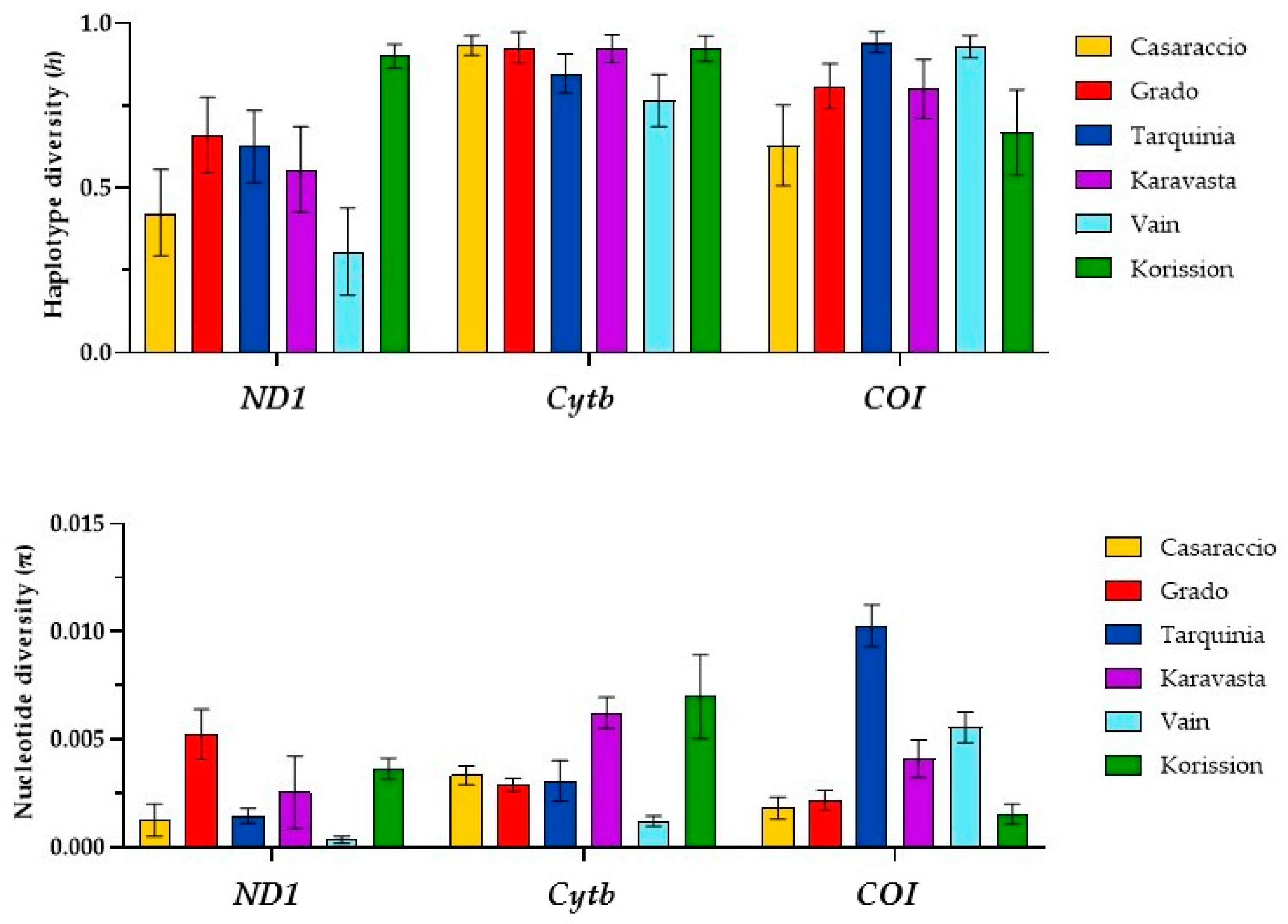
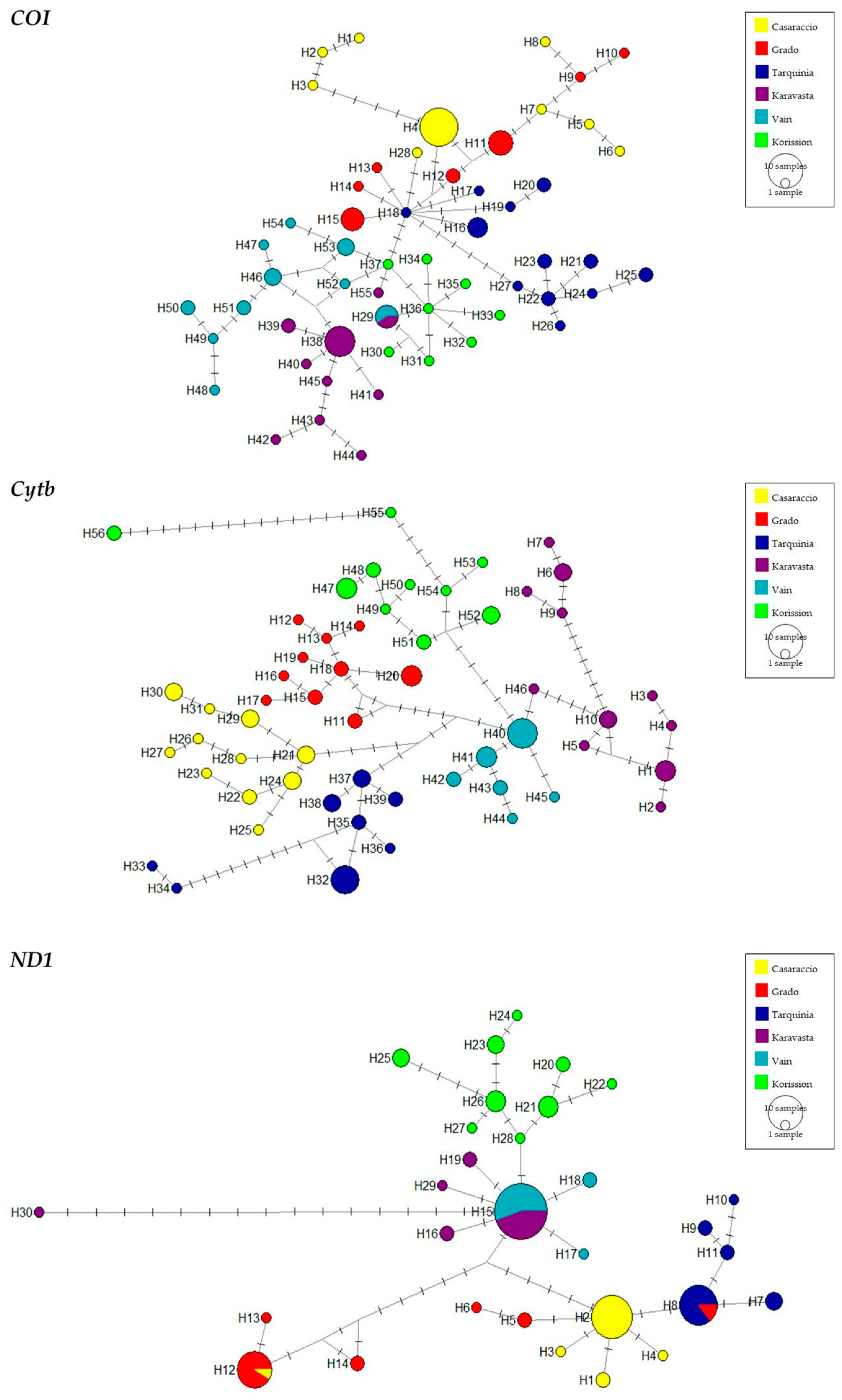
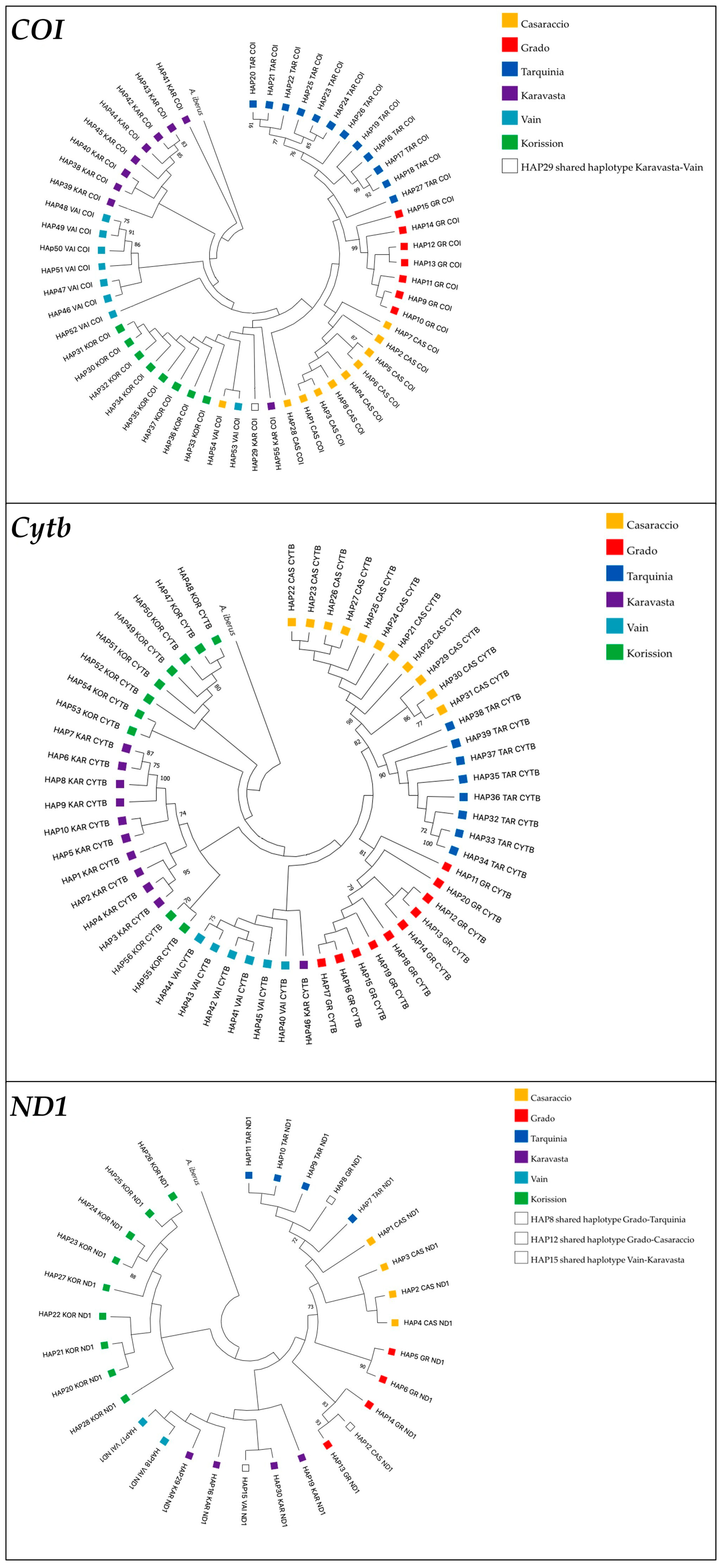
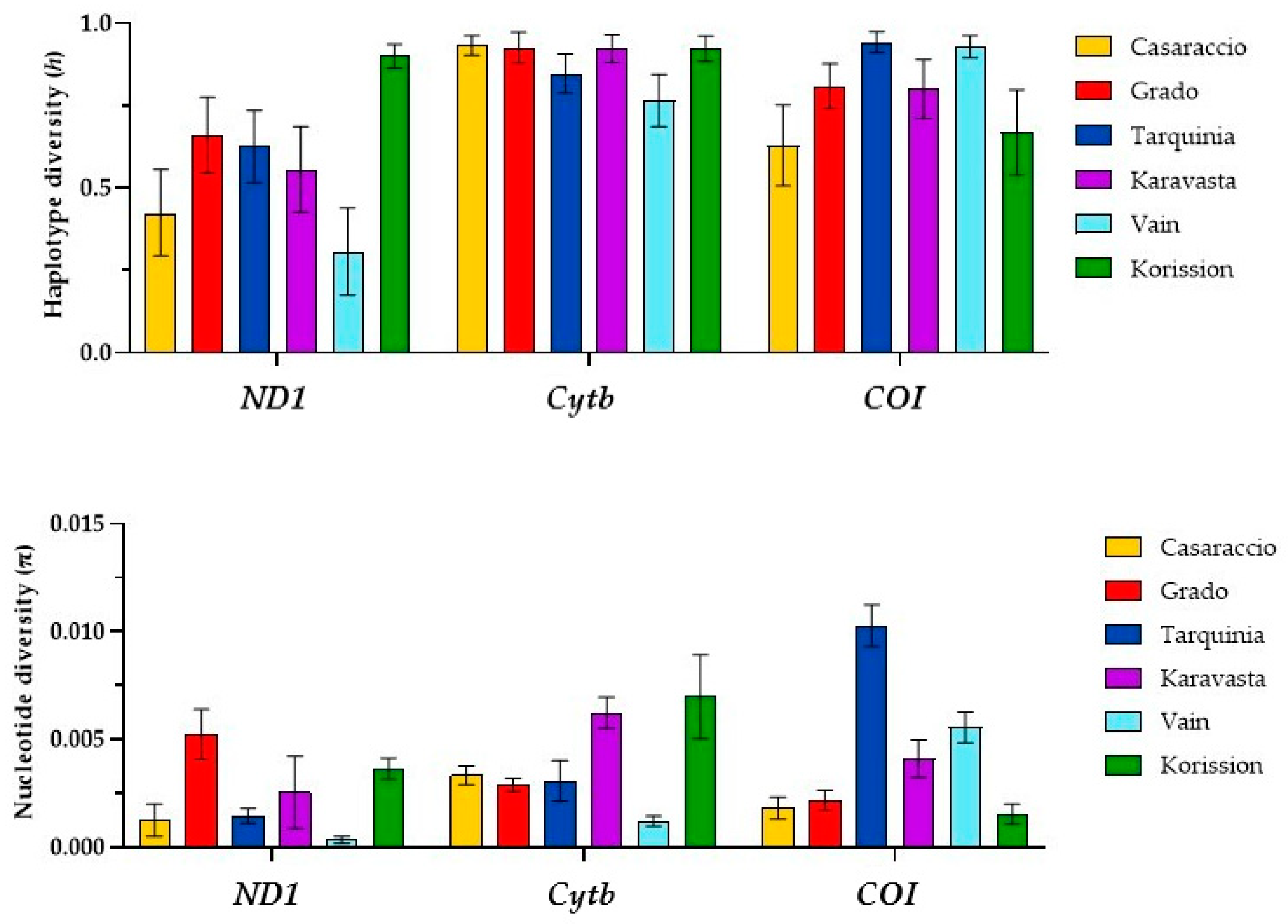
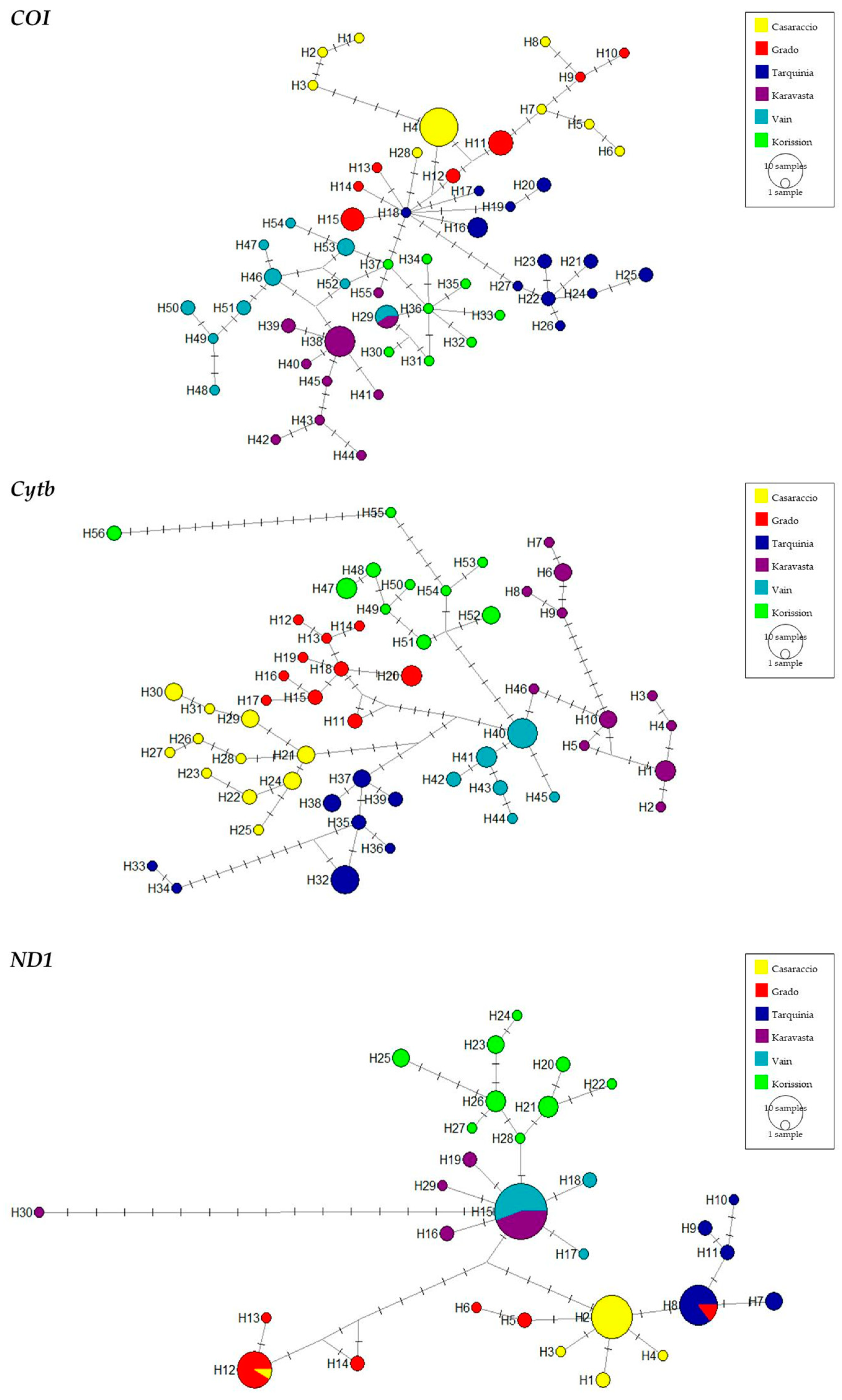
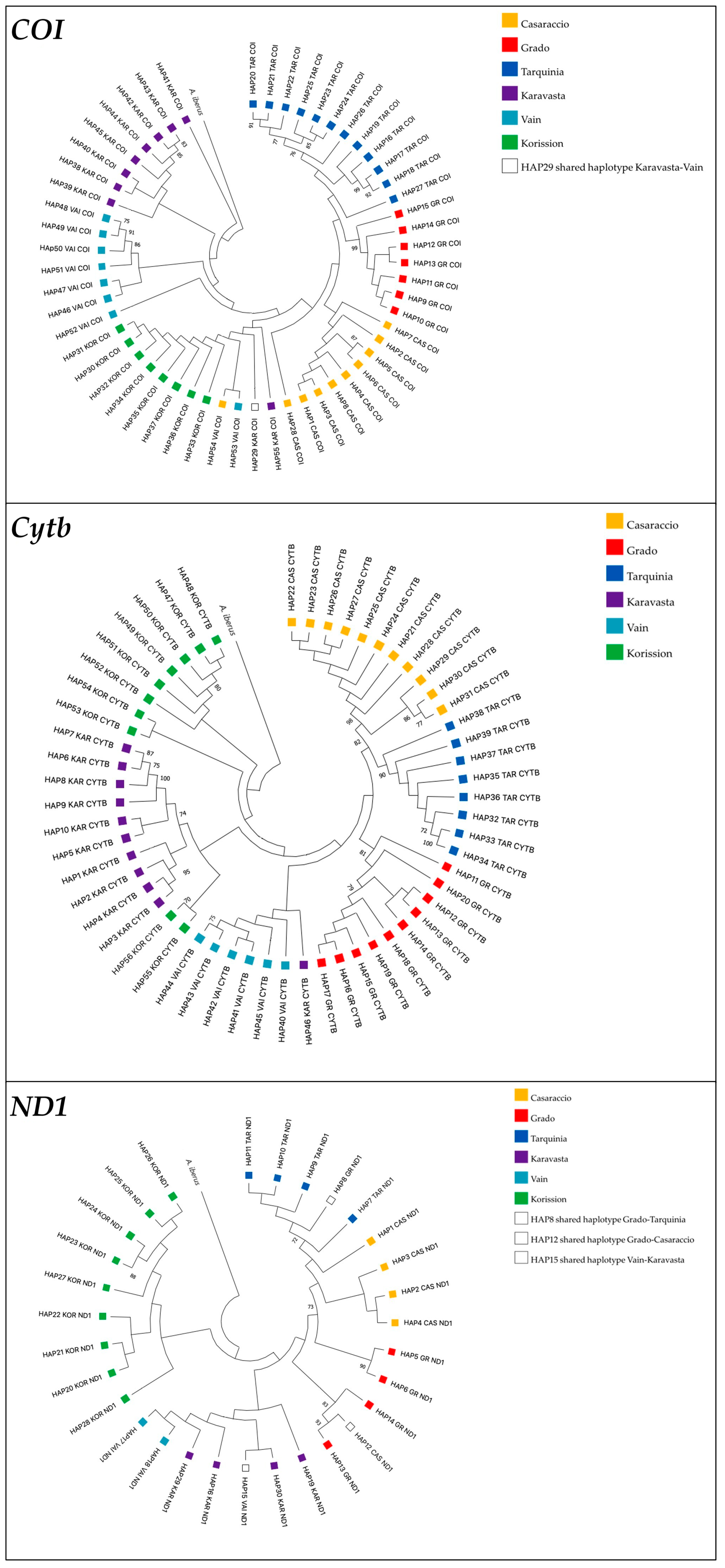
3.1. Gene Sequences Analysis and Distribution of Genetic Variation
3.2. Evidence of Selection Signals in Mitochondrial Genes
3.3. Non-Synonymous Changes Analysis
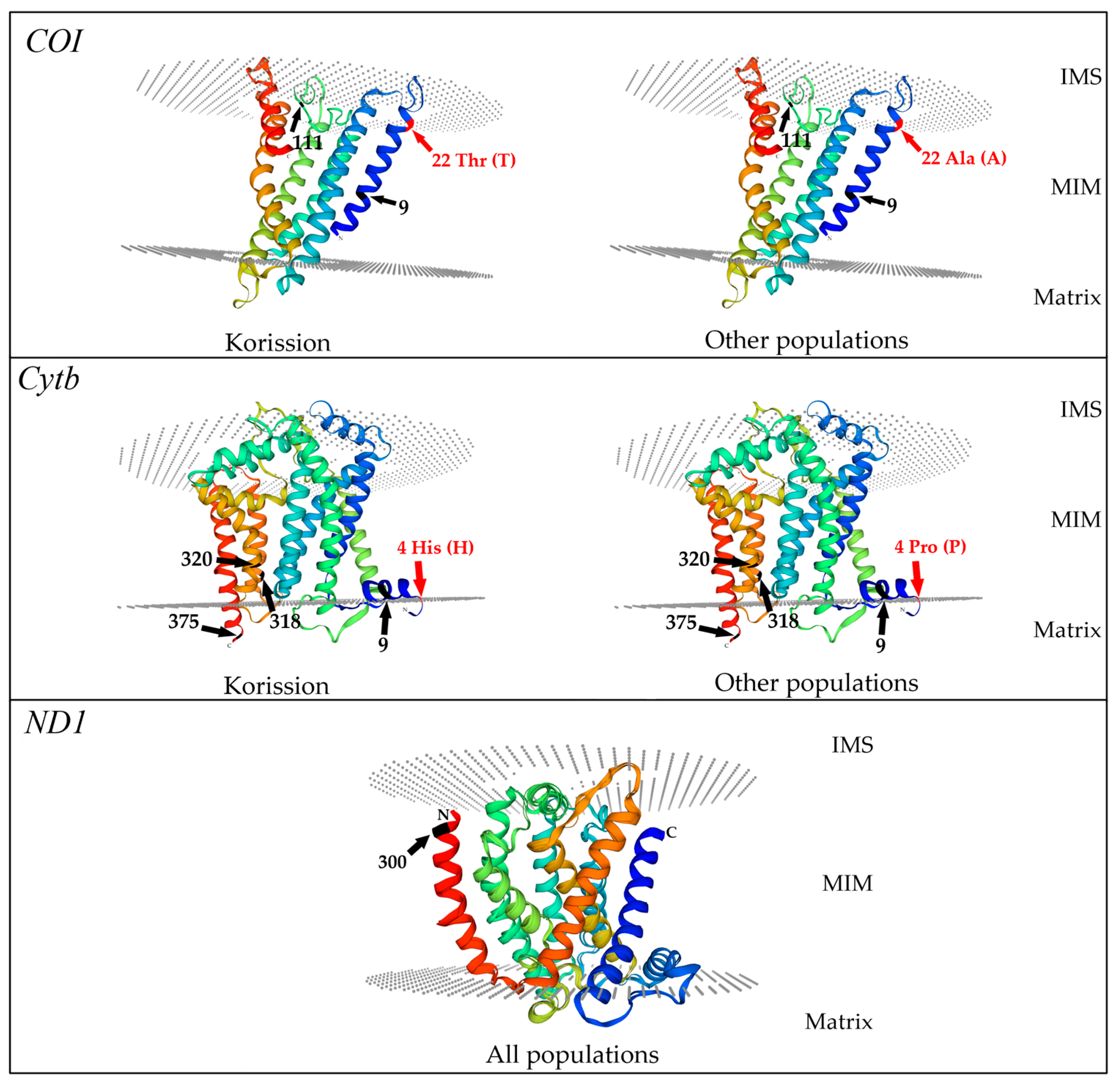
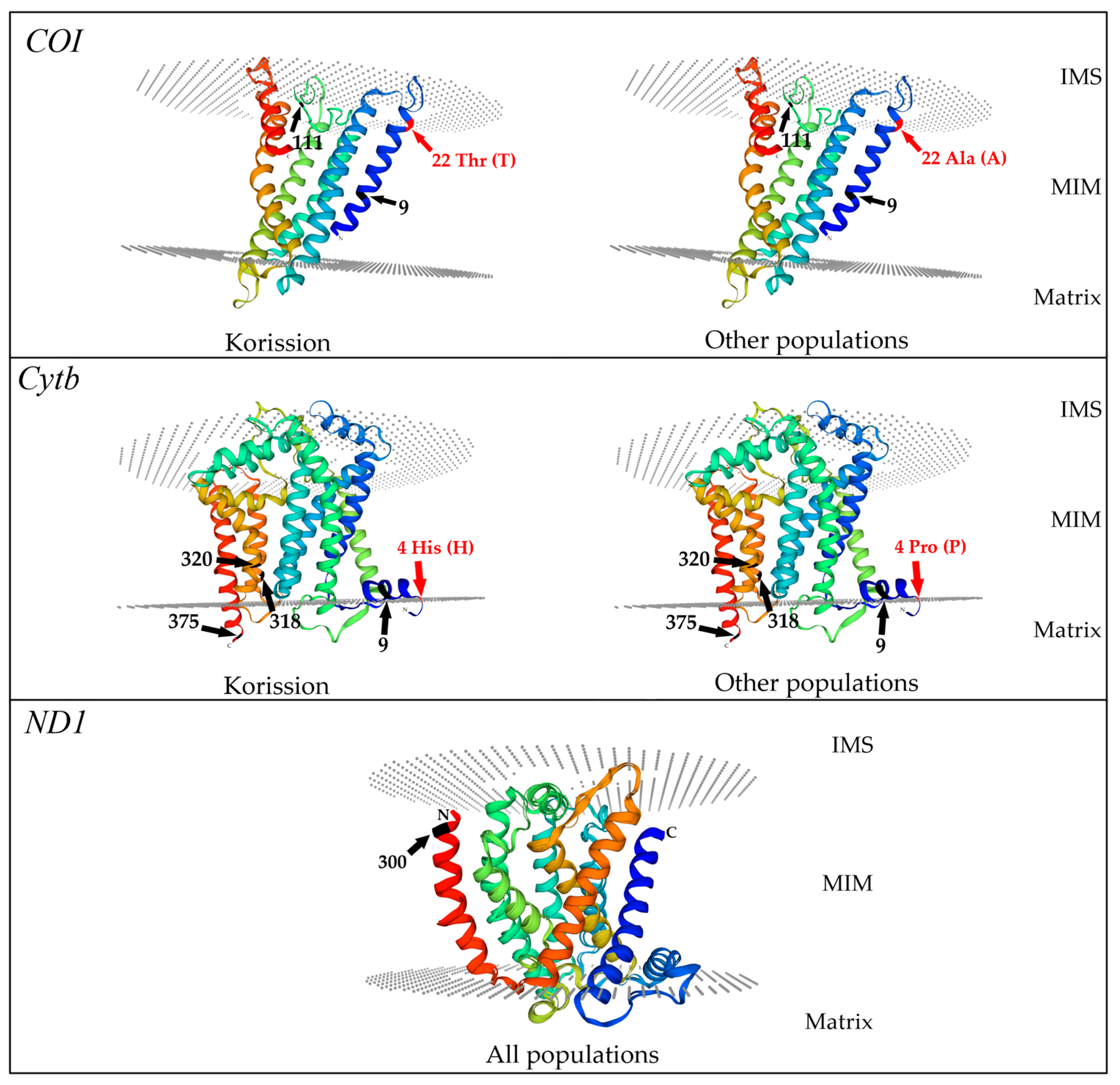
3.4. 3D Models of COI, Cytb, and ND1 Fragment Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lajbner, Z.; Pnini, R.; Camus, M.F.; Miller, J.; Dowling, D.K. Experimental Evidence That Thermal Selection Shapes Mitochondrial Genome Evolution. Sci. Rep. 2018, 8, 9500. [Google Scholar] [CrossRef]
- Lu, Z.; Liu, T.; Liu, Y.; Wang, Y.; Liu, J.; Liu, B.; Gong, L.; Liu, L. Climate Adaptation and Drift Shape the Genomes of Two Eel-Goby Sister Species Endemic to Contrasting Latitude. Animals 2023, 13, 3240. [Google Scholar] [CrossRef]
- Noll, D.; Leon, F.; Brandt, D.; Pistorius, P.; Le Bohec, C.; Bonadonna, F.; Trathan, P.N.; Barbosa, A.; Rey, A.R.; Dantas, G.P.M.; et al. Positive Selection over the Mitochondrial Genome and Its Role in the Diversification of Gentoo Penguins in Response to Adaptation in Isolation. Sci. Rep. 2022, 12, 3767. [Google Scholar] [CrossRef]
- Mori, S.; Matsunami, M. Signature of Positive Selection in Mitochondrial DNA in Cetartiodactyla. Genes Genet. Syst. 2018, 93, 65–73. [Google Scholar] [CrossRef]
- Awadi, A.; Ben Slimen, H.; Schaschl, H.; Knauer, F.; Suchentrunk, F. Positive Selection on Two Mitochondrial Coding Genes and Adaptation Signals in Hares (Genus Lepus) from China. BMC Ecol. Evol. 2021, 21, 100. [Google Scholar] [CrossRef]
- Dhar, D.; Dey, D.; Basu, S.; Fortunato, H. Insight into the adaptive evolution of mitochondrial genomes in intertidal chitons. J. Molluscan Stud. 2021, 87, eyab018. [Google Scholar] [CrossRef]
- Li, X.D.; Jiang, G.F.; Yan, L.Y.; Li, R.; Mu, Y.; Deng, W.A. Positive Selection Drove the Adaptation of Mitochondrial Genes to the Demands of Flight and High-Altitude Environments in Grasshoppers. Front. Genet. 2018, 9, 605. [Google Scholar] [CrossRef]
- Dalziel, A.C.; Martin, N.; Laporte, M.; Guderley, H.; Bernatchez, L. Adaptation and acclimation of aerobic exercise physiology in Lake Withefish ecotypes (Coregonus clupeaformis). Evolution 2015, 69, 2167–2186. [Google Scholar] [CrossRef]
- Consuegra, S.; John, E.; Verspoor, E.; De Leaniz, C.G. Patterns of Natural Selection Acting on the Mitochondrial Genome of a Locally Adapted Fish Species. Genet. Sel. Evol. 2015, 47, 58. [Google Scholar] [CrossRef]
- Iftikar, F.I.; MacDonald, J.R.; Baker, D.W.; Renshaw, G.M.C.; Hickey, A.J.R. Could Thermal Sensitivity of Mitochondria Determine Species Distribution in a Changing Climate? J. Exp. Biol. 2014, 217, 2348–2357. [Google Scholar] [CrossRef]
- Sokolova, I.M. Ectotherm Mitochondrial Economy and Responses to Global Warming. Acta Physiol. 2023, 237, e13950. [Google Scholar] [CrossRef]
- Richards, J.G. Physiological, Behavioral and Biochemical Adaptations of Intertidal Fishes to Hypoxia. J. Exp. Biol. 2011, 214, 191–199. [Google Scholar] [CrossRef]
- Sokolova, I. Mitochondrial Adaptations to Variable Environments and Their Role in Animals’ Stress Tolerance. Integr. Comp. Biol. 2018, 58, 519–531. [Google Scholar] [CrossRef]
- Acquavita, A.; Aleffi, I.F.; Benci, C.; Bettoso, N.; Crevatin, E.; Milani, L.; Tamberlich, F.; Toniatti, L.; Barbieri, P.; Licen, S.; et al. Annual characterization of the nutrients and trophic state in a Mediterranean coastal lagoon: The Marano and Grado Lagoon (northern Adriatic Sea). Reg. Stud. Mar. Sci. 2015, 2, 132–144. [Google Scholar] [CrossRef]
- Çomo, E.; Hasimi, A.; Murtaj, B.; Hoxhaj, J.; Lushaj, B. Evaluation of Physic-Chemical Features of the Main Coastal Lagoons of Narta and Karavasta, in Albania. Online Int. Interdiscip. Res. J. 2018, 8, 14–26. [Google Scholar]
- Roselli, L.; Stanca, E.; Ludovisi, A.; Durante, G.; Souza, J.S.D.; Dural, M.; Alp, T.; Bulent, S.; Gjoni, V.; Ghinis, S.; et al. Multi-scale biodiverity patterns in phytoplankton from coastal lagoons: The Eastern Mediterranean. Transitional Waters Bull. 2013, 7, 202–219. [Google Scholar] [CrossRef]
- Bonifazi, A.; Galli, S.; Gravina, M.F.; Ventura, D. Macrozoobenthos Structure and Dynamics in a Mediterranean Hypersaline Ecosystem with Implications for Wetland Conservation. Water 2023, 15, 1411. [Google Scholar] [CrossRef]
- Ferrarin, C.; Umgiesser, G.; Bajo, M.; Bellafiore, D.; De Pascalis, F.; Ghezzo, M.; Mattassi, G.; Scroccaro, I. Hydraulic zonation of the lagoons of Marano and Grado, Italy. A modeling approach. Estuar. Coast. Shelf Sci. 2010, 87, 561–572. [Google Scholar] [CrossRef]
- Koto, R.; Bani, A.; Skuka, N. Physico-Chemical Characteristics and Heavy Metal Contents of Water from Karavasta lagoon, Albania. Albanian J. Agric. Sci. 2014, 13, 55–60. [Google Scholar]
- Gravina, M.F.; Cabiddu, S.; Como, S.; Floris, A.; Padedda, B.M.; Pusceddu, A.; Magni, P. Disentangling heterogeneity and commonalities in nanotidal Mediterranean lagoons through environmental features and macrozoobenthic assemblages. Estuar. Coast. Shelf Sci. 2020, 237, 106688. [Google Scholar] [CrossRef]
- Bacu, A.; Babani, F.; Malollari, I. A comparative study on the efficiency of use of different physical and biological parameters for the evaluation of the level of trophy in the lagoon system of Kune-Vain, Albania. J. Environ. Prot. Ecol. 2011, 12, 45–53. [Google Scholar]
- Pappalardo, A.M.; Gonzalez, E.G.; Tigano, C.; Doadrio, I.; Ferrito, V. Comparative Pattern of Genetic Structure in Two Mediterranean Killifishes Aphanius fasciatus and Aphanius iberus Inferred from Both Mitochondrial and Nuclear Data. J. Fish. Biol. 2015, 87, 69–87. [Google Scholar] [CrossRef] [PubMed]
- Pappalardo, A.M.; Federico, C.; Sabella, G.; Saccone, S.; Ferrito, V. A COI Nonsynonymous Mutation as Diagnostic Tool for Intraspecific Discrimination in the European Anchovy Engraulis encrasicolus (Linnaeus). PLoS ONE 2015, 10, e0143297. [Google Scholar] [CrossRef]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D.N. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Resour 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Messing, J. New M13 vectors for cloning. Meth. Enzymol. 1983, 101, 20–78. [Google Scholar]
- Schmidt, T.R.; Gold, J.R. Complete Sequence of the Mitochondrial Cytochrome b Gene in the Cherryfin Shiner, Lythrurus roseipinnis (Teleostei: Cyprinidae). ASIH 1993, 3, 880–883. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kosakovsky Pond, S.L.; Frost, S.D.W. Datamonkey: Rapid Detection of Selective Pressure on Individual Sites of Codon Alignments. Bioinformatics 2005, 21, 2531–2533. [Google Scholar] [CrossRef]
- Murrell, B.; Moola, S.; Mabona, A.; Weighill, T.; Sheward, D.; Kosakovsky Pond, S.L.; Scheffler, K. FUBAR: A Fast, Unconstrained Bayesian AppRoximation for Inferring Selection. Mol. Biol. Evol. 2013, 30, 1196–1205. [Google Scholar] [CrossRef]
- Murrell, B.; Wertheim, J.O.; Moola, S.; Weighill, T.; Scheffler, K.; Kosakovsky Pond, S.L. Detecting Individual Sites Subject to Episodic Diversifying Selection. PLoS Genet. 2012, 8, e1002764. [Google Scholar] [CrossRef]
- Silva, G.; Lima, F.P.; Martel, P.; Castilho, R. Thermal Adaptation and Clinal Mitochondrial DNA Variation of European Anchovy. Proc. R. Soc. B 2014, 281, 20141093. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Buchan, D.W.A.; Jones, D.T. The PSIPRED Protein Analysis Workbench: 20 Years On. Nucleic Acids Res. 2019, 47, W402–W407. [Google Scholar] [CrossRef]
- Ferrito, V.; Maltagliati, F.; Mauceri, A.; Adorno, A.; Tigano, C. Morphological and Genetic Variation in Four Italian Populations of Lebias fasciata (Teleostei, Cyprinodontidae). Ital. J. Zool. 2003, 70, 115–121. [Google Scholar] [CrossRef]
- Triantafyllidis, A.; Leonardos, I.; Bista, I.; Kyriazis, I.D.; Stoumboudi, M.T.; Kappas, I.; Amat, F.; Abatzopoulos, T.J. Phylogeography and Genetic Structure of the Mediterranean Killifish Aphanius fasciatus (Cyprinodontidae). Mar. Biol. 2007, 152, 1159–1167. [Google Scholar] [CrossRef]
- Pappalardo, A.M.; Ferrito, V.; Messina, A.; Guarino, F.; Patarnello, T.; De Pinto, V.; Tigano, C. Genetic structure of the killifish Aphanius fasciatus Nardo 1827 (Teleostei, Cyprinodontidae), results of mitochondrial DNA analysis. J. Fish Biol. 2008, 72, 1154–1173. [Google Scholar] [CrossRef]
- Ferrito, V.; Pappalardo, A.M.; Canapa, A.; Barucca, M.; Doadrio, I.; Olmo, E.; Tigano, C. Mitochondrial Phylogeography of the Killifish Aphanius fasciatus (Teleostei, Cyprinodontidae) Reveals Highly Divergent Mediterranean Populations. Mar. Biol. 2013, 160, 3193–3208. [Google Scholar] [CrossRef]
- Buj, I.; Miočić-Stošić, J.; Marčić, Z.; Mustafić, P.; Zanella, D.; Mrakovčić, M.; Mihinjač, T.; Ćaleta, M. Population Genetic Structure and Demographic History of Aphanius fasciatus (Cyprinodontidae: Cyprinodontiformes) from Hypersaline Habitats in the Eastern Adriatic. Sci. Mar. 2015, 79, 399–408. [Google Scholar] [CrossRef]
- Cavraro, F.; Malavasi, S.; Torricelli, P.; Gkenas, C.; Liousia, V.; Leonardos, I.; Kappas, I.; Abatzopoulos, T.J.; Triantafyllidis, A. Genetic Structure of the South European Toothcarp Aphanius fasciatus (Actinopterygii: Cyprinodontidae) Populations in the Mediterranean Basin with a Focus on the Venice Lagoon. Eur. Zool. J. 2017, 84, 153–166. [Google Scholar] [CrossRef]
- Langeneck, J.; Englezou, C.; Di Maggio, M.; Castelli, A.; Maltagliati, F. Phylogeography of Aphanius fasciatus (Osteichthyes: Aphaniidae) in the Mediterranean Sea, with a Focus on Its Conservation in Cyprus. Hydrobiologia 2021, 848, 4093–4114. [Google Scholar] [CrossRef]
- Betts, M.J.; Russell, R.B. Amino Acid Properties and Consequences of Substitutions. In Bioinformatics for Geneticists; Barnes, M.R., Gray, I.C., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2003; pp. 291–314. [Google Scholar]
- Ward, R.D.; Holmes, B.H. An analysis of nucleotide and amino acid variability in the barcode region of cytochrome c oxidase I (cox1) in fishes. Mol. Ecol. Notes 2007, 7, 899–907. [Google Scholar] [CrossRef]
- Degli Esposti, M.; De Vries, S.; Criml, M.; Ghelh, A.; Patarnello, T.; Meyer, A. Mitochondrial Cytochrome b: Evolution and Structure of the Protein. Biochim. Biophys. Acta (BBA)-Bioenerg. 1993, 1143, 243–271. [Google Scholar] [CrossRef]
- McClellan, D.A.; Palfreyman, E.J.; Smith, M.J.; Moss, J.L.; Christensen, R.G.; Sailsbery, J.K. Physicochemical Evolution and Molecular Adaptation of the Cetacean and Artiodactyl Cytochrome b Proteins. Mol. Biol. Evol. 2004, 22, 437–455. [Google Scholar] [CrossRef]
- Beckstead, W.A.; Ebbert, M.T.W.; Rowe, M.J.; McClellan, D.A. Evolutionary Pressure on Mitochondrial Cytochrome b Is Consistent with a Role of CytbI7T Affecting Longevity during Caloric Restriction. PLoS ONE 2009, 4, e5836. [Google Scholar] [CrossRef]
- Dalziel, A.C.; Moyes, C.D.; Fredriksson, E.; Lougheed, S.C. Molecular Evolution of Cytochrome c Oxidase in High-Performance Fish (Teleostei: Scombroidei). J. Mol. Evol. 2006, 62, 319–331. [Google Scholar] [CrossRef]
- Baltazar-Soares, M.; de Araújo Lima, A.R.; Silva, G. Targeted Sequencing of Mitochondrial Genes Reveals Signatures of Molecular Adaptation in a Nearly Panmictic Small Pelagic Fish Species. Genes 2021, 12, 91. [Google Scholar] [CrossRef]
- Chung, D.J.; Bryant, H.J.; Schulte, P.M. Thermal acclimation and subspecies-specific effects on heart and brain mitochondrial performance in a eurythermal teleost (Fundulus heteroclitus). J. Exp. Biol. 2017, 220, 1459–1471. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Locality | Coordinates | COI | Cytb | ND1 |
|---|---|---|---|---|---|
| Italy | Casaraccio Lagoon | 40°55′01″ N 8°13′04″ E | 21 (9) | 20 (11) | 21 (4) |
| Grado Lagoon | 45°40′44″ N 13°23′45″ E | 17 (7) | 10 (10) | 19 (6) | |
| Tarquinia Salt works | 42°14′47″ N 11°44′50″ E | 20 (12) | 20 (8) | 20 (4) | |
| Albania | Karavasta Lagoon | 40°56′02″ N 19°29′08″ E | 20 (10) | 18 (11) | 18 (4) |
| Vain Lagoon | 41°45′46″ N 19°34′54″ E | 18 (9) | 18 (6) | 18 (3) | |
| Greece | Korission Lagoon | 39°26′45″ N 19°54′14″ E | 18 (8) | 18 (10) | 20 (9) |
| Total | 114 (55 + 1) * | 110 (56) | 116 (27 + 3) ** |
| Gene | ID Primer | Sequence | Reference |
|---|---|---|---|
| COI | VF2_t1 | 5′-TGTAAAACGACGGCCAGTCAACCAACCACAAAGACATTGGCAC-3′ | [24] |
| FishR2_t1 | 5′-CAGGAAACAGCTATGACACTTCAGGGTGACCGAAGAATCAGAA-3′ | ||
| M13_FW | 5′-TGTAAAACGACGGCCAGT-3′ | [25] | |
| M13_REV | 5′-CAGGAAACAGCTATGAC-3′ | ||
| Cytb | L14724 | 5′-GTGACTTGAAAAACCACCGTTG-3′ | [26] |
| H15915 | 5′-CAACGATCTCCGGTTTACAAGAC-3′ | ||
| ND1 | ND1_Aph_FW | 5′-TGATTTTAGTACTTTATGCAATTATTA-3′ | This study |
| ND1_Aph_REV | 5′-GTGGGGGGGCAAGCCAGA-3′ |
| Codon | 6 | 7 | 8 | 9 | 20 | 34 | 72 | 111 | 152 | 222 | |
| Selection type | Purifying | Purifying | Purifying | Positive | Purifying | Purifying | Purifying | Positive | Purifying | Purifying | |
| COI | FEL | * | * | * | * | * | * | * | |||
| FUBAR | # | # | # | # | ## | ||||||
| MEME | *** | ||||||||||
| Codon | 9 | 31 | 144 | 159 | 243 | 311 | 318 | 320 | 375 | ||
| Selection type | Positive | Purifying | Purifying | Purifying | Purifying | Purifying | Positive | Positive | Positive | ||
| Cytb | FEL | * | ** | * | * | ||||||
| FUBAR | # | # | # | # | # | # | # | ||||
| MEME | * | ** | |||||||||
| Codon | 5 | 70 | 78 | 183 | 223 | 225 | 244 | 300 | 302 | ||
| Selection type | Purifying | Purifying | Purifying | Purifying | Purifying | Purifying | Purifying | Positive | Purifying | ||
| ND1 | FEL | * | * | * | * | ||||||
| FUBAR | ## | ## | # | # | ## | # | # | # | # | ||
| MEME |
| Gene | Population | Codon 22 | ||||
| COI | Grado | A (N-polar) | ||||
| Tarquinia | A (N-polar) | |||||
| Casaraccio | A (N-polar) | |||||
| Vain | A (N-polar) | |||||
| Karavasta | A (N-polar) | |||||
| Korission | T (polar) | |||||
| Gene | Population | Codon 3 | Codon 4 | Codon 6 | Codon 7 | Codon 111 |
| Cytb | Grado | N (polar) | P (N-polar) | P (N-polar) | L (N-polar) | V (N-polar) |
| Tarquinia | N (polar) | P (N-polar) | P (N-polar) | F (N-polar) | V (N-polar) | |
| Casaraccio | N (polar) | P (N-polar) | P (N-polar) | F (N-polar) | I (N-polar) | |
| Vain | N (polar) | P (N-polar) | P (N-polar) | F (N-polar) | V (N-polar) | |
| Karavasta | N (polar) | P (N-polar) | P (N-polar) | L (N-polar) | V (N-polar) | |
| Korission | T (polar) | H (polar, basic) | L (N-polar) | L (N-polar) | V (N-polar) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pappalardo, A.M.; Calogero, G.S.; Šanda, R.; Giuga, M.; Ferrito, V. Evidence for Selection on Mitochondrial OXPHOS Genes in the Mediterranean Killifish Aphanius fasciatus Valenciennes, 1821. Biology 2024, 13, 212. https://doi.org/10.3390/biology13040212
Pappalardo AM, Calogero GS, Šanda R, Giuga M, Ferrito V. Evidence for Selection on Mitochondrial OXPHOS Genes in the Mediterranean Killifish Aphanius fasciatus Valenciennes, 1821. Biology. 2024; 13(4):212. https://doi.org/10.3390/biology13040212
Chicago/Turabian StylePappalardo, Anna Maria, Giada Santa Calogero, Radek Šanda, Marta Giuga, and Venera Ferrito. 2024. "Evidence for Selection on Mitochondrial OXPHOS Genes in the Mediterranean Killifish Aphanius fasciatus Valenciennes, 1821" Biology 13, no. 4: 212. https://doi.org/10.3390/biology13040212
APA StylePappalardo, A. M., Calogero, G. S., Šanda, R., Giuga, M., & Ferrito, V. (2024). Evidence for Selection on Mitochondrial OXPHOS Genes in the Mediterranean Killifish Aphanius fasciatus Valenciennes, 1821. Biology, 13(4), 212. https://doi.org/10.3390/biology13040212