

Trunk Injection Delivery of Biocontrol Strains of Trichoderma spp. Effectively Suppresses Nut Rot by Gnomoniopsis castaneae in Chestnut (Castanea sativa Mill.)



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Soil Sampling and Trichoderma Isolation
2.3. Identification of G. castaneae and Trichoderma spp. Strains
2.4. In Vitro Antagonism Tests
2.5. Biocontrol Agent Mixture Preparation
2.6. In Vivo Tests
2.7. Assessments of G. castaneae Incidence
2.8. Statistical Analyses
3. Results
3.1. Gnomoniopsis castaneae and Trichoderma spp. Identification
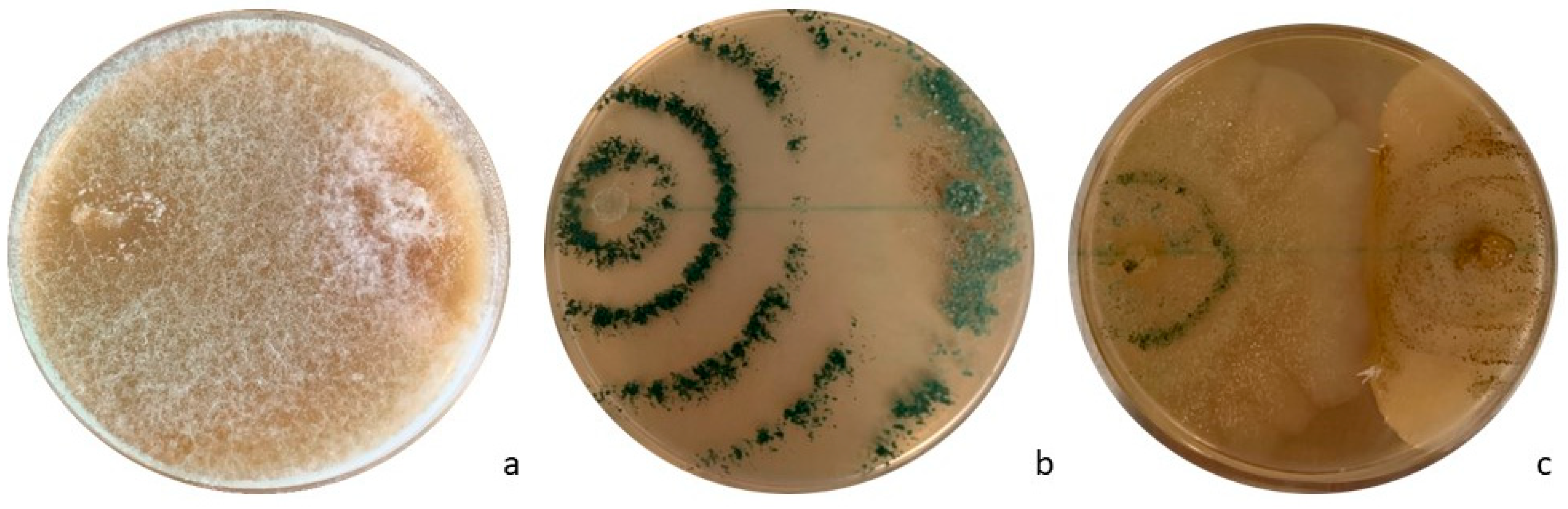
3.2. In Vitro Tests
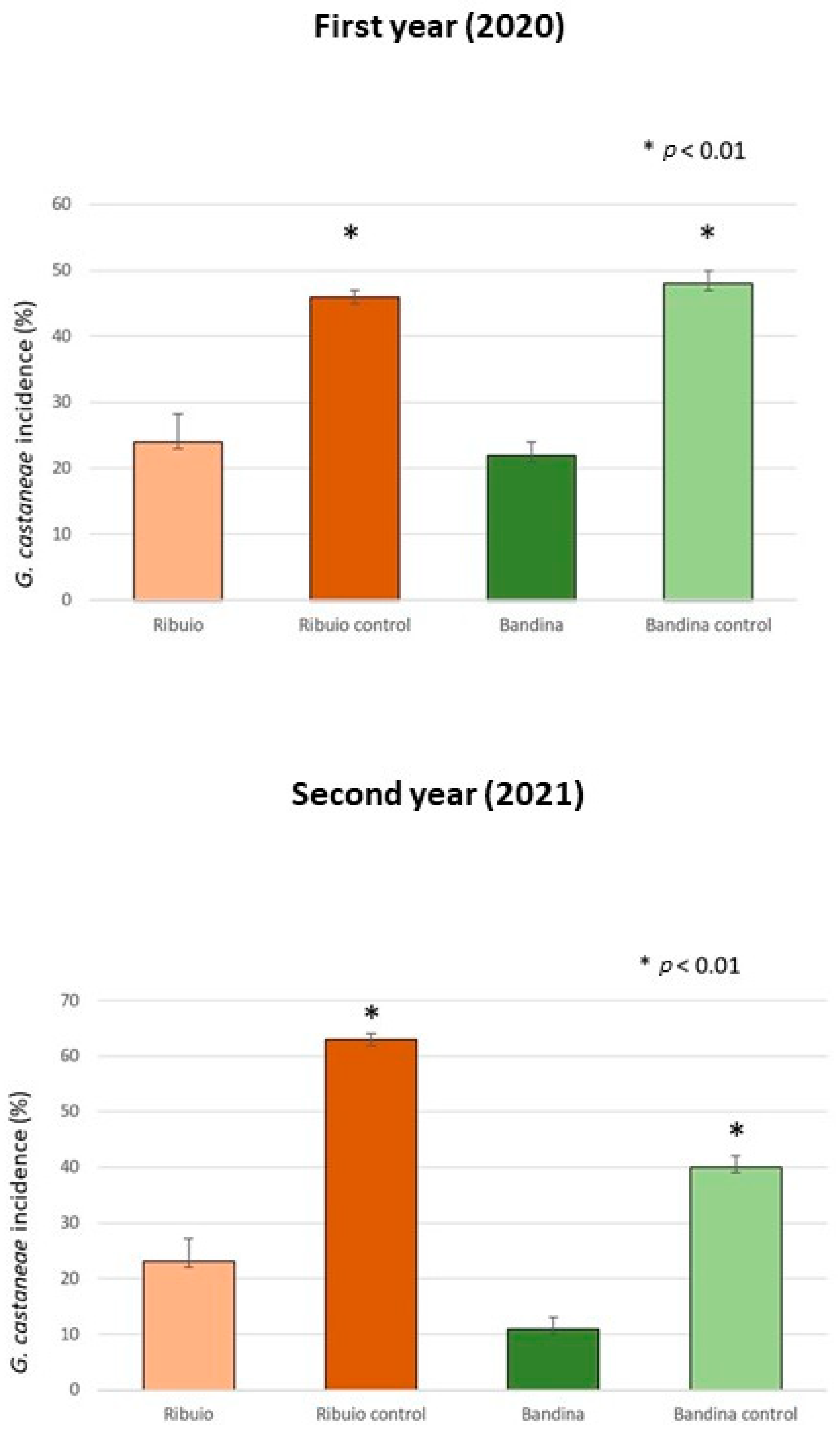
3.3. Endotherapic Treatments in the Field
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aglietti, C.; Cappelli, A.; Andreani, A. From Chestnut Tree (Castanea sativa) to Flour and Foods: A Systematic Review of the Main Criticalities and Control Strategies towards the Relaunch of Chestnut Production Chain. Sustainability 2022, 14, 12181. [Google Scholar] [CrossRef]
- Mattioni, C.; Martin, M.A.; Chiocchini, F.; Cherubini, M.; Gaudet, M.; Pollegioni, P.; Velichkov, I.; Jarman, R.; Chambers, F.M.; Damian, V.L.; et al. Landscape genetics structure of European sweet chestnut (Castanea sativa Mill): Indications for conservation priorities. Tree Genet. Genomes. 2017, 13, 39. [Google Scholar] [CrossRef]
- FAO. World Food and Agriculture Statistical Yearbook 2022; FAO: Rome, Italy, 2022. [Google Scholar]
- López-Sáez, J.A.; Glais, A.; Robles-López, S.; Alba-Sánchez, F.; Pérez-Díaz, S.; Abel-Schaad, D.; Luelmo-Lautenschlaeger, R. Unraveling the naturalness of sweet chestnut forests (Castanea sativa Mill.) in central Spain. Veg. Hist. Archaeobot. 2017, 26, 167–182. [Google Scholar] [CrossRef]
- Bussotti, F.; Papitto, G.; Di Martino, D.; Cocciufa, C.; Cindolo, C.; Cenni, E.; Bettini, D.; Iacopetti, G.; Pollastrini, M. Defoliation, recovery and increasing mortality in Italian forests: Levels, patterns and possible consequences for forest multifunctionality. Forests 2021, 12, 1476. [Google Scholar] [CrossRef]
- Fernandes, P.; Colavolpe, M.B.; Serrazina, S.; Costa, R.L. European and American Chestnuts: An Overview of the Main Threats and Control Efforts. Front. Plant Sci. 2022, 13, 951844. [Google Scholar] [CrossRef] [PubMed]
- Agnoletti, M. The degradation of traditional landscape in a mountain area of Tuscany during the 19th and 20th centuries: Implications for biodiversity and sustainable management. For. Ecol. Manag. 2007, 249, 5–17. [Google Scholar] [CrossRef]
- Frati, A.; Landi, D.; Marinelli, C.; Gianni, G.; Fontana, L.; Migliorini, M.; Pierucci, F.; Garcia-Gil, M.; Meacci, E. Nutraceutical Properties of Chestnut Flours: Beneficial Effects on Skeletal Muscle Atrophy. Food Funct. 2014, 5, 2870–2882. [Google Scholar] [CrossRef]
- Cevasco, R.; Moreno, D.; Balzaretti, R.; Watkins, C. Historical chestnut cultures, climate and rural landscapes in the Apennines. In The Future of Heritage as Climates Change: Loss, Adaptation and Creativity; Harvey, D., Perry, J., Eds.; Routledge: London, UK, 2015; pp. 130–147. [Google Scholar]
- Morales-Rodríguez, C.; Bastianelli, G.; Caccia, R.; Bedini, G.; Massantini, R.; Moscetti, R.; Thomidis, T.; Vannini, A. Impact of ‘brown rot’caused by Gnomoniopsis castanea on chestnut fruits during the post-harvest process: Critical phases and proposed solutions. J. Sci. Food Agric. 2022, 102, 680–687. [Google Scholar] [CrossRef]
- Shuttleworth, L.A.; Liew, E.C.Y.; Guest, D.I. Survey of the incidence of chestnut rot in south-eastern Australia. Australas. Plant Pathol. 2013, 42, 63–72. [Google Scholar] [CrossRef]
- Sieber, T.N.; Jermini, M.; Conedera, M. Effects of the harvest method on the infestation of chestnuts (Castanea sativa) by insects and moulds. J. Phytopathol. 2007, 155, 497–504. [Google Scholar] [CrossRef]
- Lione, G.; Danti, R.; Fernandez-Conradi, P.; Ferreira-Cardoso, J.V.; Lefort, F.; Marques, G.; Meyer, J.B.; Prospero, S.; Radócz, L.; Robin, C.; et al. The emerging pathogen of chestnut Gnomoniopsis castaneae: The challenge posed by a versatile fungus. Eur. J. Plant Pathol. 2019, 153, 671–685. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Luchi, N.; Rizzo, D.; Pepori, A.L.; Pecori, F.; Santini, A. Rapid diagnostics for Gnomoniopsis smithogilvyi (syn. Gnomoniopsis castaneae) in chestnut nuts: New challenges by using LAMP and real-time PCR methods. AMB Express 2021, 11, 105. [Google Scholar] [CrossRef] [PubMed]
- Lema, F.; Baptista, P.; Oliveira, C.; Ramalhosa, E. Brown Rot Caused by Gnomoniopsis smithogilvyi (syn. Gnomoniopsis castaneae) at the Level of the Chestnut Tree (Castanea sativa Mill.). Appl. Sci. 2023, 13, 3969. [Google Scholar] [CrossRef]
- Silva-Campos, M.; Nadiminti, P.; Cahill, D. Rapid and Accurate Detection of Gnomoniopsis smithogilvyi the Causal Agent of Chestnut Rot, through an Internally Controlled Multiplex PCR Assay. Pathogens 2022, 11, 907. [Google Scholar] [CrossRef]
- Robin, D.C.; Marchand, P.A. Evolution of the biocontrol active substances in the framework of the European Pesticide Regulation (EC) No. 1107/2009. Pest. Manag. Sci. 2019, 75, 950–958. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Ghazanfar, M.U.; Raza, M.; Raza, W.; Qamar, M.I. Trichoderma as Potential Biocontrol Agent, Its Exploitation in Agriculture: A Review. Plant Prot. 2018, 2, 109–135. [Google Scholar]
- Weindling, R. Trichoderma lignorum as a parasite of other soil fungi. Phytopathology 1932, 22, 837–845. [Google Scholar]
- Lorito, S.; Selva, J.; Basili, R.; Romano, F.; Tiberti, M.M.; Piatanesi, A. Probabilistic hazard for seismically induced tsunamis: Accuracy and feasibility of inundation maps. Geophys. J. Int. 2015, 200, 574–588. [Google Scholar] [CrossRef]
- Alfiky, A.; Weisskopf, L. Deciphering Trichoderma–Plant–Pathogen Interactions for Better Development of Biocontrol Applications. J. Fungi 2021, 7, 61. [Google Scholar] [CrossRef]
- Pasche, S.; Crovadore, J.; Pelleteret, P.; Jermini, M.; Mauch-Mani, B.; Oszako, T.; Lefort, F. Biological control of the latent pathogen Gnomoniopsis smithogylvyi in European chestnut grafting scions using Bacillus amyloliquefaciens and Trichoderma atroviride. Dendrobiology 2016, 75, 113–122. [Google Scholar] [CrossRef]
- Murolo, S.; Concas, J.; Romanazzi, G. Use of biocontrol agents as potential tools in the management of chestnut blight. Biol. Control 2019, 132, 102–109. [Google Scholar] [CrossRef]
- Schubert, M.; Fink, S.; Schwarze, F.W.M.R. Evaluation of Trichoderma spp. as a biocontrol agent against wood decay fungi in urban trees. Biol. Control 2008, 45, 111–123. [Google Scholar] [CrossRef]
- Prospero, S.; Botella, L.; Santini, A.; Robin, C. Biological control of emerging forest diseases: How can we move from dreams to reality? For. Ecol. Manag. 2021, 496, 119377. [Google Scholar] [CrossRef]
- Berger, G.; Czarnocka, K.; Cochard, B.; Oszako, T.; Lefort, F. Biocontrol Endotherapy with Trichoderma spp. and Bacillus amyloliquefaciens against Phytophthora spp.: A Comparative Study with Phosphite Treatment on Quercus robur and Fagus sylvatica. J. Agric. Sci. Technol. A 2015, 5, 428–439. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Moricca, S.; Ginetti, B.; Ragazzi, B. Species and organ-specificity in endophytes colonizing healthy and declining Mediterranean oaks. Phytopathol. Mediterr. 2012, 51, 587–598. [Google Scholar]
- Badalyan, S.M.; Garibyan, N.G.; Innocenti, G. Antagonistic activity of xylotrophic mushrooms against pathogenic fungi of cereals in dual culture. Antagonistic activity of xylotrophic mushrooms against pathogenic fungi of cereals in dual culture. Phytopathol. Med. 2002, 41, 220–225. [Google Scholar]
- Idris, H.A.; Labuschagne, N.; Korsten, L. Screening rhizobacteria for biological control of Fusarium root and crown rot of sorghum in Ethiopia. Biol. Control 2007, 40, 97–106. [Google Scholar] [CrossRef]
- Montecchio, L. A Venturi Effect Can Help Cure Our Trees. J. Vis. Exp. 2013, 80, e51199. [Google Scholar]
- Visentin, I.; Gentile, S.; Valentino, D.; Gonthier, P.; Tamietti, G.; Cardinale, F. Gnomoniopsis castanea sp. nov. (Gnomoniaceae, Diaporthales) as the causal agent of nut rot in sweet chestnut. J. Plant. Pathol. 2012, 94, 411–419. [Google Scholar]
- Shuttleworth, L.A.; Guest, D.I.; Liew, E.C.Y. Fungal planet description sheet 108–Gnomoniopsis smithogilvyi L.A. Shuttleworth, E.C.Y. Liew & D.I. Guest, sp. nov. Persoonia 2012, 28, 142–143. [Google Scholar]
- Beccaro, G.L.; Alma, A.; Gonthier, P.; Mellano, M.G.; Ferracini, C.; Giordano, L.; Lione, G.; Donno, D.; Boni, I.; Ebone, A.; et al. Chestnut R&D Centre, Piemonte (Italy): 10 years of activity. Acta Hortic. 2018, 1220, 133–140. [Google Scholar]
- Beccaro, G.; Alma, A.; Bounous, G.; Gomes-Laranjo, J. The Chestnut Handbook: Crop & Forest Management; CRC Press: Boca Raton, FL, USA, 2019; 378p. [Google Scholar]
- Conti, V.; Salusti, P.; Romi, M.; Cantini, C. Effects of drying methods and temperatures on the quality of chestnut flours. Foods 2022, 11, 1364. [Google Scholar] [CrossRef] [PubMed]
- Nerva, L.; Costa, L.D.; Ciacciulli, A.; Sabbadini, S.; Pavese, V.; Dondini, L.; Vendramin, E.; Caboni, E.; Perrone, I.; Moglia, A.; et al. The role of Italy in the use of advanced plant genomic techniques on fruit trees: State of the art and future perspectives. Int. J. Mol. 2023, 24, 977. [Google Scholar] [CrossRef] [PubMed]
- Drais, M.I.; Gusella, G.; Mazzaglia, A.; Polizzi, G. A quantitative PCR assay for the detection and quantification of Septoria pistaciarum, the causal agent of pistachio leaf spot in Italy. PLoS ONE 2023, 18, e0286130. [Google Scholar] [CrossRef]
- Rodrigues, A.O.; De Mio, L.L.M.; Soccol, C.R. Trichoderma as a powerful fungal disease control agent for a more sustainable and healthy agriculture: Recent studies and molecular insights. Planta 2023, 257, 31. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.; Hermosa, R.; Lorito, M.; Monte, E. Trichoderma: A multipurpose, plant-beneficial microorganism for eco-sustainable agriculture. Nat. Rev. Microbiol. 2023, 21, 312–326. [Google Scholar] [CrossRef]
- Klein, D.; Eveleigh, D.E. Ecology of Trichoderma. In Trichoderma & Gliocladium, Basic Biology, Taxonomy and Genetics; Kubicek, C.P., Harmna, G.E., Eds.; Taylor and Francis: London, UK, 1998; Volume 1, pp. 57–69. [Google Scholar]
- Chandra, S.; Singh, B.K. Trichoderma spp.: As potential bio-control agents (BCAs) against fungal plant pathogens. Indian J. Life Sci. 2016, 5, 105. [Google Scholar]
- Hidayah, B.N. Biological Control Potential of Trichoderma Species and Bacterial Antagonists against Sclerotinia sclerotiorum on Canola in Western Australia. Int. J. Agric. Biol. 2022, 27, 215–227. [Google Scholar] [CrossRef]
- Asad, S.A. Mechanisms of Action and Biocontrol Potential of Trichoderma against Fungal Plant Diseases—A Review. Ecol. Complex. 2022, 49, 100978. [Google Scholar] [CrossRef]
- Shalaby, T.A.; Taha, N.; El-Beltagi, H.S.; El-Ramady, H. Combined Application of Trichoderma harzianum and Paclobutrazol to Control Root Rot Disease Caused by Rhizoctonia solani of Tomato Seedlings. Agronomy 2022, 12, 3186. [Google Scholar] [CrossRef]
- Natsiopoulos, D.; Tziolias, A.; Lagogiannis, I.; Mantzoukas, S.; Eliopoulos, P.A. Growth-Promoting and Protective Effect of Trichoderma atrobrunneum and T. simmonsii on Tomato against Soil-Borne. Fungal Pathogens. Crops 2022, 2, 202–217. [Google Scholar] [CrossRef]
- Sehim, A.E.; Hewedy, O.A.; Altammar, K.A.; Alhumaidi, M.S.; Abd Elghaffar, R.Y. Trichoderma asperellum empowers tomato plants and suppresses Fusarium oxysporum through priming responses. Front. Microbiol. 2023, 14, 1140378. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Millen, M.R.; Bailey, A.M. Analysis of Trichoderma as an effective biological control agent against the honey fungus (Armillaria spp.). Biol. Control 2023, 188, 105424. [Google Scholar] [CrossRef]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar]
- Bunbury-Blanchette, A.; Walker, A. Trichoderma species show biocontrol potential in dual culture and greenhouse bioassays against Fusarium basal rot of onion. Biol. Control 2018, 130, 127–135. [Google Scholar] [CrossRef]
- Gajera, H.; Domadiya, R.; Patel, S.; Kapopara, M.; Golakiya, B. Molecular mechanism of Trichoderma as bio-control agents against phytopathogen system—A review. Curr. Res. Microbiol. Biotechnol. 2013, 1, 133–142. [Google Scholar]
- Nawrocka, J.; Małolepsza, U. Diversity in Plant Systemic Resistance Induced by Trichoderma. Biol. Control 2013, 67, 149–156. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Mendoza-Mendoza, A.; Zeilinger, S.; Horwitz, B.A. Mycoparasitism as a Mechanism of Trichoderma-Mediated Suppression of Plant Diseases. Fungal Biol. Rev. 2022, 39, 15–33. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ribuio Chestnut Stand | |||
|---|---|---|---|
| Number | Age (Y = Young, <30 years; M = Mature, 80–150 years; O = Overripe, >150 years) | Diameter at Breast Height DBH (cm) | Height (m) |
| 1 | O | 79.58 | 11.8 |
| 2 | O | 42.97 | 15.9 |
| 3 | O | 86.26 | 14 |
| 4 | O | 95.49 | 12.3 |
| 5 | O | 44.56 | 10 |
| 6 | M | 38.2 | 14.7 |
| 7 | M | 34.06 | 13.8 |
| 8 | O | 113.32 | 12.8 |
| 9 | M | 53.16 | 16.3 |
| 10 | M | 33.42 | 14.1 |
| 11 | O | 65.57 | 10.9 |
| 12 | O | 108.23 | 10.9 |
| 13 | O | 113 | 11.1 |
| 14 | O | 66.85 | 9.4 |
| 15 | O | 82.76 | 7.6 |
| 16 | O | 64.3 | 7.8 |
| 17 | O | 43.93 | 12.8 |
| 18 | O | 69.39 | 7.7 |
| 19 | M | 36.61 | 6.1 |
| 20 | NA | 28.87 | NA |
| 21 | O | 62.71 | 6.9 |
| 22 | O | 70.66 | 6.8 |
| 23 | O | 60.48 | NA |
| 24 | NA | 60.48 | NA |
| 25 | O | 63.66 | NA |
| 26 | NA | 49.34 | NA |
| 27 | O | 92.31 | NA |
| 28 | O | 98.04 | NA |
| 29 | O | 127.32 | NA |
| 30 | M | 49.34 | NA |
| 31 | O | 119.37 | NA |
| 32 | O | 93.9 | NA |
| Bandina chestnut stand | |||
| 1 | M | 70.03 | 10 |
| 2 | M | 38.83 | 11.9 |
| 3 | M | 55.7 | 10.02 |
| 4 | M | 53.16 | 11.06 |
| 5 | M | 59.21 | 13.6 |
| 6 | M | 41.38 | 13.4 |
| 7 | M | 61.75 | 15.1 |
| 8 | M | 51.57 | 14.1 |
| 9 | M | 45.2 | 14 |
| 10 | M | 48.38 | 16.1 |
| 11 | M | 74.8 | 9.5 |
| 12 | M | 44.56 | 12.9 |
| 13 | M | 67.16 | 10.5 |
| 14 | M | 73.21 | 11.1 |
| 15 | M | 49.34 | 16.8 |
| 16 | M | 53.48 | 16 |
| 17 | M | 46.47 | 11.4 |
| 18 | M | 48.38 | 12.2 |
| 19 | M | 59.84 | 14 |
| 20 | M | 60.8 | 11.9 |
| 21 | M | 50.93 | 11.5 |
| 22 | M | 66.85 | 11.5 |
| 23 | M | 58.89 | 10.3 |
| 24 | O | 194.17 | 10.9 |
| 25 | Y | 30.24 | 13.6 |
| 26 | Y | 35.01 | 13.4 |
| 27 | M | 48.7 | 13.5 |
| 28 | M | 49.02 | 12.4 |
| 29 | Y | 29.92 | 16.3 |
| 30 | M | 59.21 | 14.2 |
| 31 | M | 37.24 | 14.5 |
| 32 | M | 42.34 | 14.7 |
| 33 | M | 40.11 | 14.3 |
| 34 | M | 61.75 | 15.6 |
| Type of Interaction | Interaction | Value |
|---|---|---|
| A | Stop of colony growth by contact with mutual inhibition | 1 |
| B | Remote stop without mycelial contact | 2 |
| C | Growth of one colony over another without initial stop | 3 |
| CA1 | Partial growth of one colony over another after contact arrest | 3.5 |
| CA2 | Complete growth of one colony upon another after contact arrest | 4.5 |
| CB1 | Partial growth of one colony upon another after remote arrest | 4 |
| CB2 | Complete growth of one colony upon another after remote arrest | 5 |
| Isolates | A (cm2) * | I (%) * | CI ** |
|---|---|---|---|
| Trichoderma viride | 63.58 | 78.12 b | CA2 |
| Trichoderma harzianum | 63.58 | 99.14 c | CA2 |
| Trichoderma atroviride | 41.03 | 51.82 a | CA1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benigno, A.; Aglietti, C.; Cacciola, S.O.; Moricca, S. Trunk Injection Delivery of Biocontrol Strains of Trichoderma spp. Effectively Suppresses Nut Rot by Gnomoniopsis castaneae in Chestnut (Castanea sativa Mill.). Biology 2024, 13, 143. https://doi.org/10.3390/biology13030143
Benigno A, Aglietti C, Cacciola SO, Moricca S. Trunk Injection Delivery of Biocontrol Strains of Trichoderma spp. Effectively Suppresses Nut Rot by Gnomoniopsis castaneae in Chestnut (Castanea sativa Mill.). Biology. 2024; 13(3):143. https://doi.org/10.3390/biology13030143
Chicago/Turabian StyleBenigno, Alessandra, Chiara Aglietti, Santa Olga Cacciola, and Salvatore Moricca. 2024. "Trunk Injection Delivery of Biocontrol Strains of Trichoderma spp. Effectively Suppresses Nut Rot by Gnomoniopsis castaneae in Chestnut (Castanea sativa Mill.)" Biology 13, no. 3: 143. https://doi.org/10.3390/biology13030143
APA StyleBenigno, A., Aglietti, C., Cacciola, S. O., & Moricca, S. (2024). Trunk Injection Delivery of Biocontrol Strains of Trichoderma spp. Effectively Suppresses Nut Rot by Gnomoniopsis castaneae in Chestnut (Castanea sativa Mill.). Biology, 13(3), 143. https://doi.org/10.3390/biology13030143