
Exploring the Immune Response against RSV and SARS-CoV-2 Infection in Children


Abstract
Simple Summary
Abstract
1. Introduction
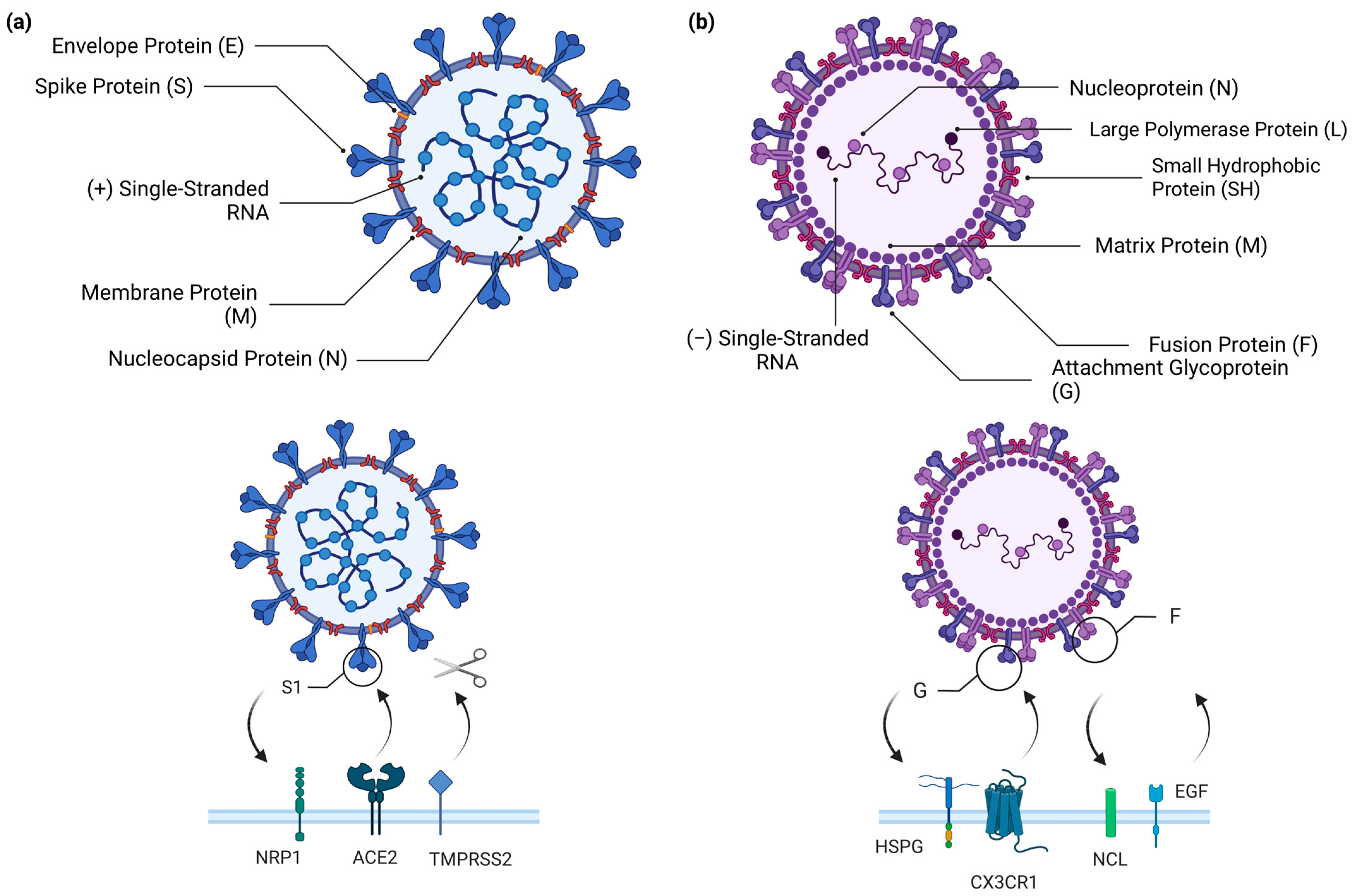
RSV, SARS-CoV-2, and Cellular Airway Infections
2. General Immunology of Viral Respiratory Infection in the Mucosal Airway
3. Comparing the Immune Responses to RSV and COVID-19 in Children
3.1. Innate Immunity
3.1.1. RIG-I and MDA5
3.1.2. Interferons
3.1.3. ISGs
4. Adaptive Immune Response
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, F.F.; Yu, S.J. Global morbidity and mortality of lower respiratory infections: A population-based study. Respir. Med. 2022, 205, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Alimohamadi, Y.; Tola, H.H. Case fatality rate of COVID-19: A systematic review and meta-analysis. J. Prev. Med. Hyg. 2021, 62, E311–E320. [Google Scholar]
- Chonmaitree, T.; Revai, K. Viral upper respiratory tract infection and otitis media complication in young children. Clin. Infect. Dis. 2008, 46, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Caballero, M.T.; Bianchi, A.M. Mortality Associated with Acute Respiratory Infections among Children at Home. J. Infect. Dis. 2019, 219, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Hemming, V.G. Viral respiratory diseases in children: Classification, etiology, epidemiology, and risk factors. J. Pediatr. 1994, 124, S13–S16. [Google Scholar] [CrossRef]
- Taylor, A.; Whittaker, E. The Changing Epidemiology of Respiratory Viruses in Children During the COVID-19 Pandemic: A Canary in a COVID Time. Pediatr. Infect. Dis. J. 2022, 41, e46–e48. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in children younger than 5 years in 2019: A systematic analysis. Lancet 2022, 399, 2047–2064. [Google Scholar] [CrossRef] [PubMed]
- Staadegaard, L.; Caini, S. The Global Epidemiology of RSV in Community and Hospitalized Care: Findings from 15 Countries. Open Forum Infect. Dis. 2021, 8, ofab159. [Google Scholar] [CrossRef]
- Shi, T.; McAllister, D.A. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: A systematic review and modelling study. Lancet 2017, 390, 946–958. [Google Scholar] [CrossRef]
- Meissner, H.C. Viral Bronchiolitis in Children. N. Engl. J. Med. 2016, 374, 62–72. [Google Scholar] [CrossRef]
- Child Mortality and COVID-19. Available online: https://data.unicef.org/topic/child-survival/covid-19/ (accessed on 8 May 2023).
- Ledford, H. Deaths from COVID ‘incredibly rare’ among children. Nature 2021, 595, 639. [Google Scholar] [CrossRef]
- Tagarro, A.; Garcia-Salido, A. Low COVID-19 mortality in Spanish children. Lancet Child. Adolesc. Health 2021, 5, e24–e25. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.L.; Harwood, R. Risk factors for PICU admission and death among children and young people hospitalized with COVID-19 and PIMS-TS in England during the first pandemic year. Nat. Med. 2022, 28, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Rinaldi, E. Coronavirus disease 2019 (COVID-19) in children and/or adolescents: A meta-analysis. Pediatr. Res. 2021, 89, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Karim, S.S.A.; Karim, Q.A. Omicron SARS-CoV-2 variant: A new chapter in the COVID-19 pandemic. Lancet 2021, 398, 2126–2128. [Google Scholar] [CrossRef] [PubMed]
- Long, F.; Zhu, S. Update on the treatment of multisystem inflammatory syndrome in children associated with COVID-19. Future Virol. 2023; Epub ahead of print. [Google Scholar]
- Carl, A. Pierce et al. COVID-19 and children. Science 2022, 377, 1144–1149. [Google Scholar]
- Halabi, K.C.; Saiman, L. The epidemiology of respiratory syncytial virus in New York city during the coronavirus disease—2019 pandemic compared with previous years. J. Pediatr. 2022, 242, 242–244. [Google Scholar] [CrossRef]
- Mosscrop, L.G.; Williams, T.C. Respiratory syncytial virus after the SARS-CoV-2 pandemic—What next? Nat. Rev. Immunol. 2022, 22, 589–590. [Google Scholar] [CrossRef]
- Halabi, K.C.; Wang, H. Respiratory syncytial virus and SARS-CoV-2 coinfections in children. Pediatr. Pulmonol. 2022, 57, 3158–3160. [Google Scholar] [CrossRef]
- Swets, M.C.; Russell, C.D. SARS-CoV-2 co-infection with influenza viruses, respiratory syncytial virus, or adenoviruses. Lancet 2022, 399, 1463–1464. [Google Scholar] [CrossRef]
- Ke, Z.; Dillard, R.S. The Morphology and Assembly of Respiratory Syncytial Virus Revealed by Cryo-Electron Tomography. Viruses 2018, 10, 446. [Google Scholar] [CrossRef] [PubMed]
- Kung, Y.A.; Lee, K.M. Molecular Virology of SARS-CoV-2 and Related Coronaviruses. Microbiol. Mol. Biol. Ver. 2022, 86, e0002621. [Google Scholar] [CrossRef]
- Griffiths, C.; Drews, S.J. Respiratory Syncytial Virus: Infection, Detection, and New Options for Prevention and Treatment. Clin. Microbiol. Rev. 2017, 30, 277–319. [Google Scholar] [CrossRef]
- Masre, S.F.; Jufri, N.F. Classical and alternative receptors for SARS-CoV-2 therapeutic strategy. Rev. Med. Virol. 2021, 31, 1–9. [Google Scholar] [CrossRef]
- Yu, J.M.; Fu, Y.H.; Peng, X.L.; Zheng, Y.P.; He, J.S. Genetic diversity and molecular evolution of human respiratory syncytial virus A and B. Sci. Rep. 2021, 11, 12941. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zeng, F. Comparison of Clinical Features and Outcomes of Medically Attended COVID-19 and Influenza Patients in a Defined Population in the 2020 Respiratory Virus Season. Front. Public Health 2021, 9, 587425. [Google Scholar] [CrossRef] [PubMed]
- Bunyavanich, S.; Do, A. Nasal Gene Expression of Angiotensin-Converting Enzyme 2 in Children and Adults. JAMA 2020, 323, 2427–2429. [Google Scholar] [CrossRef] [PubMed]
- Koch, C.M.; Priggge, A.D. Age-related Differences in the Nasal Mucosal Immune Response to SARS-CoV-2. Am. J. Respir. Cell Mol. Biol. 2022, 66, 206–222. [Google Scholar] [CrossRef]
- Schuler, B.A.; Habermann, A.C. Age-determined expression of priming protease TMPRSS2 and localization of SARS-CoV-2 in lung epithelium. J. Clin. Investig. 2021, 131, e140766. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Chew, K.Y. Ancestral SARS-CoV-2, but not Omicron, replicates less efficiently in primary pediatric nasal epithelial cells. PLoS Biol. 2022, 20, e3001728. [Google Scholar] [CrossRef]
- Loske, J.; Röhmel, J. Pre-activated antiviral innate immunity in the upper airways controls early SARS-CoV-2 infection in children. Nat. Biotechnol. 2022, 40, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Travaglini, K.J.; Nabhan, A.N. A molecular cell atlas of the human lung from single-cell RNA sequencing. Nature 2020, 587, 619–625. [Google Scholar] [CrossRef]
- Maughan, E.F.; Hynds, R.E. Cell-intrinsic differences between human airway epithelial cells from children and adults. iScience 2022, 25, 105409. [Google Scholar] [CrossRef]
- Mettelman, R.C.; Allen, E.K. Mucosal immune responses to infection and vaccination in the respiratory tract. Imunnity 2022, 55, 749–780. [Google Scholar] [CrossRef] [PubMed]
- Pilette, C.; Ouadhiri, Y. Lung mucosal immunity: Immunoglobulin-A revisited. Eur. Respir. J. 2001, 18, 571–588. [Google Scholar] [CrossRef]
- Kurono, Y. The mucosal immune system of the upper respiratory tract and recent progress in mucosal vaccines. Auris Nasus Larynx 2022, 49, 1–10. [Google Scholar] [CrossRef]
- Zhang, N.; Van Crombruggen, K. Barrier function of the nasal mucosa in health and type-2 biased airway diseases. Allergy 2016, 71, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Persson, B.D.; Jaffe, A.B. Respiratory Syncytial Virus Can Infect Basal Cells and Alter Human Airway Epithelial Differentiation. PLoS ONE 2014, 9, e102368. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.E.; Gonzales, R.A. The histopathology of fatal untreated human respiratory syncytial virus infection. Mod. Pathol. 2007, 20, 108–119. [Google Scholar] [CrossRef] [PubMed]
- D’Agnillo, F.; Walters, K.A. Lung epithelial and endothelial damage, loss of tissue repair, inhibition of fibrinolysis, and cellular senescence in fatal COVID-19. Sci. Transl. Med. 2021, 13, eabj7790. [Google Scholar] [CrossRef] [PubMed]
- Calkovska, A.; Kolomaznik, M. Alveolar type II cells and pulmonary surfactant in COVID-19 era. Physiol. Res. 2021, 70, S195–S208. [Google Scholar] [CrossRef]
- Huang, B. Mucins produced by type II pneumocyte: Culprits in SARS-CoV-2 pathogenesis. Cell Mol. Immunol. 2021, 18, 1823–1825. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, J.J.; Avellaneda, A.M. Host Components Contributing to Respiratory Syncytial Virus Pathogenesis. Front. Immunol. 2019, 10, 2152. [Google Scholar] [CrossRef]
- Brookes, O.; Boland, S. Co-culture of type I and type II pneumocytes as a model of alveolar epithelium. PLoS ONE 2021, 16, e0248798. [Google Scholar] [CrossRef]
- Yamada, T.; Kawabata, Y. Pneumocyte injury and ubiquitin-positive pneumocytes in interstitial lung diseases. Histopathology 2015, 66, 161–172. [Google Scholar] [CrossRef][Green Version]
- Wang, L.; Cheng, W. Respiratory syncytial virus infection accelerates lung fibrosis through the unfolded protein response in a bleomycin-induced pulmonary fibrosis animal model. Mol. Med. Rep. 2017, 16, 310–316. [Google Scholar] [CrossRef]
- Rajan, A.; Weaver, A.M. The Human Nose Organoid Respiratory Virus Model: An Ex Vivo Human Challenge Model To Study Respiratory Syncytial Virus (RSV) and Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Pathogenesis and Evaluate Therapeutics. mBio 2022, 13, e0351121. [Google Scholar] [CrossRef]
- Ravindra, N.G.; Alfajaro, M.M. Single-cell longitudinal analysis of SARS-CoV-2 infection in human airway epithelium identifies target cells, alterations in gene expression, and cell state changes. PLoS Biol. 2021, 19, e3001143. [Google Scholar] [CrossRef] [PubMed]
- Lukassen, S.; Chua, R.L. SARS-CoV-2 receptor ACE2 and TMPRSS2 are primarily expressed in bronchial transient secretory cells. EMBO J. 2020, 39, e105114. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Curtis, N. Why Does the Severity of COVID-19 Differ With Age? Understanding the Mechanisms Underlying the Age Gradient in Outcome Following SARS-CoV-2 Infection. Pediatr. Infect. Dis. J. 2022, 41, 36–45. [Google Scholar] [CrossRef]
- Mueller, S.N.; Rouse, B.T. Immune responses to viruses. Clin. Immunol. 2008, 421–431. [Google Scholar] [CrossRef]
- Zhu, F.; Teng, Z. H1N1 Influenza Virus-Infected Nasal Mucosal Epithelial Progenitor Cells Promote Dendritic Cell Recruitment and Maturation. Front. Immunol. 2022, 13, 879575. [Google Scholar] [CrossRef] [PubMed]
- Kunisawa, J.; Nochi, T. Immunological commonalities and distinctions between airway and digestive immunity. Trends Immunol. 2008, 29, 505–513. [Google Scholar] [CrossRef]
- Ganesh, K.; Joshi, M.B. Neutrophil sub-types in maintaining immune homeostasis during steady state, infections and sterile inflammation. Inflamm. Res. 2023, 72, 1175–1192. [Google Scholar] [CrossRef] [PubMed]
- Zindel, J.; Kubes, P. DAMPs, PAMPs, and LAMPs in Immunity and Sterile Inflammation. Annu. Rev. Pathol. 2020, 15, 493–518. [Google Scholar] [CrossRef]
- Loo, Y.M.; Gale, M. Immune Signaling by RIG-I-Like Receptors. Immunity 2011, 34, 680–692. [Google Scholar] [CrossRef]
- Lazear, H.M.; Schoggins, J.W. Shared and Distinct Functions of Type I and Type III Interferons. Immunity 2019, 50, 907–923. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W. Interferon-stimulated genes: Roles in viral pathogenesis. Curr. Opin. Virol. 2014, 6, 40–46. [Google Scholar] [CrossRef]
- Schwartz, S.; Conn, G.L. RNA regulation of the antiviral protein 2′-5′-oligoadenylate synthetase. Wiley Interdiscip. Rev. RNA 2019, 10, e1534. [Google Scholar] [CrossRef]
- Liu, K. Dendritic cells. Encycl. Cell Biol. 2016, 3, 741–749. [Google Scholar]
- Netea, M.G.; Joosten, L.A.B. Trained immunity: A program of innate immune memory in health and disease. Science 2016, 352, aaf1098. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Domínguez-Andrés, J.; Barreiro, L.B.; Chavakis, T.; Divangahi, M.; Fuchs, E.; Joosten, L.A.B.; van der Meer, J.W.M.; Mhlanga, M.M.; Mulder, W.J.M.; et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 2020, 20, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Dagenais, A.; Villalba-Guerrero, C. Trained immunity: A “new” weapon in the fight against infectious diseases. Front. Immunol. 2023, 14, 1147476. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.Z.; Shuai, H. Bacillus Calmette-Guérin-induced trained immunity protects against SARS-CoV-2 challenge in K18-hACE2 mice. JCI Insight 2022, 7, e157393. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.K.; Hollander, G.A. Evolution of the immune system in humans from infancy to old age. Proc. Biol. Sci. 2015, 282, 20143085. [Google Scholar] [CrossRef] [PubMed]
- Tahamtan, A.; Besteman, S. Neutrophils in respiratory syncytial virus infection: From harmful effects to therapeutic opportunities. Br. J. Pharmacol. 2021, 178, 515–530. [Google Scholar] [CrossRef]
- Geerdink, R.J.; Pillay, J. Neutrophils in respiratory syncytial virus infection: A target for asthma prevention. J. Allergy Clin. Immunol. 2015, 136, 838–847. [Google Scholar] [CrossRef]
- George, P.M.; Reed, A. A persistent neutrophil-associated immune signature characterizes post–COVID-19 pulmonary sequelae. Sci. Transl. Med. 2022, 14, eabo5795. [Google Scholar] [CrossRef]
- McKenna, E.; Wubben, R. Neutrophils in COVID-19: Not Innocent Bystanders. Front. Immunol. 2022, 13, 864387. [Google Scholar] [CrossRef]
- Pierce, C.A.; Sy, S. Natural mucosal barriers and COVID-19 in children. JCI Insight 2021, 6, e148694. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.Q.; Zhang, M. Characteristics of immune and inflammatory responses among different age groups of pediatric patients with COVID-19 in China. World J. Pediatr. 2021, 17, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Marr, N.; Wang, T.I. Attenuation of respiratory syncytial virus-induced and RIG-I-dependent type I IFN responses in human neonates and very young children. J. Immunol. 2014, 192, 948–957. [Google Scholar] [CrossRef]
- Lifland, A.W.; Jung, J. Human Respiratory Syncytial Virus Nucleoprotein and Inclusion Bodies Antagonize the Innate Immune Response Mediated by MDA5 and MAVS. ASM J. 2012, 86, 8245–8258. [Google Scholar] [CrossRef]
- Pei, J.; Wagner, N.D. Structural basis for IFN antagonism by human respiratory syncytial virus nonstructural protein 2. Proc. Natl. Acad. Sci. USA 2021, 118, e2020587118. [Google Scholar] [CrossRef] [PubMed]
- Ban, J.; Lee, N.R. Human Respiratory Syncytial Virus NS 1 Targets TRIM25 to Suppress RIG-I Ubiquitination and Subsequent RIG-I-Mediated Antiviral Signaling. Viruses 2018, 10, 716. [Google Scholar] [CrossRef]
- Lamborn, I.T.; Jing, H. Recurrent rhinovirus infections in a child with inherited MDA5 deficiency. J. Exp. Med. 2017, 214, 1949–1972. [Google Scholar] [CrossRef]
- Antunes, K.H.; Stein, R.T. Short-chain fatty acid acetate triggers antiviral response mediated by RIG-I in cells from infants with respiratory syncytial virus bronchiolitis. EBioMedicine 2022, 77, 103891. [Google Scholar] [CrossRef] [PubMed]
- Asgari, S.; Schlapbach, L.J. Severe viral respiratory infections in children with IFIH1 loss-of-function mutations. Proc. Natl. Acad. Sci. USA 2017, 114, 8342–8347. [Google Scholar] [CrossRef] [PubMed]
- Schwab, L.S.U.; Farrukee, R. Retinoic Acid-Inducible Gene I Activation Inhibits Human Respiratory Syncytial Virus Replication in Mammalian Cells and in Mouse and Ferret Models of Infection. J. Infect. Dis. 2022, 226, 2079–2088. [Google Scholar] [CrossRef]
- Goritzka, M.; Makris, S. Alveolar macrophage-derived type I interferons orchestrate innate immunity to RSV through recruitment of antiviral monocytes. J. Exp. Med. 2015, 212, 699–714. [Google Scholar] [CrossRef]
- Paulsen, M.; Varese, A. MAVS Deficiency Is Associated With a Reduced T Cell Response Upon Secondary RSV Infection in Mice. Front. Immunol. 2020, 11, 572747. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Sato, S. RIG-I triggers a signaling-abortive anti-SARS-CoV-2 defense in human lung cells. Nat. Immunol. 2021, 22, 820–828. [Google Scholar] [CrossRef]
- Deng, J.; Zheng, Y. SARS-CoV-2 NSP7 inhibits type I and III IFN production by targeting the RIG-I/MDA5, TRIF, and STING signaling pathways. J. Med. Virol. 2023, 95, e28561. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Ma, L. Main Protease of SARS-CoV-2 Serves as a Bifunctional Molecule in Restricting Type I Interferon Antiviral Signaling. Signal Transduct. Target. Ther. 2020, 5, 221. [Google Scholar] [CrossRef]
- Liu, Y.; Qin, C. SARS-CoV-2 Nsp5 Demonstrates Two Distinct Mechanisms Targeting RIG-I and MAVS To Evade the Innate Immune Response. mBio 2021, 12, e0233521. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Shi, Y. SARS-CoV-2 ORF9b inhibits RIG-I-MAVS antiviral signaling by interrupting K63-linked ubiquitination of NEMO. Cell Rep. 2021, 34, 108761. [Google Scholar] [CrossRef]
- McIntosh, K. Interferon in nasal secretions from infants with viral respiratory tract infections. J. Pediatr. 1978, 93, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Mejias, A.; Dimo, B. Whole Blood Gene Expression Profiles to Assess Pathogenesis and Disease Severity in Infants with Respiratory Syncytial Virus Infection. PLoS Med. 2013, 10, e1001549. [Google Scholar] [CrossRef]
- Chirkova, T.; Boyoglu-Barnum, S. Respiratory syncytial virus G protein CX3C motif impairs human airway epithelial and immune cell responses. J. Virol. 2013, 87, 13466–13479. [Google Scholar] [CrossRef]
- Kalinowski, A.; Galen, B.T. Respiratory syncytial virus activates epidermal growth factor receptor to suppress interferon regulatory factor 1-dependent interferon-lambda and antiviral defense in airway epithelium. Mucosal Immunol. 2018, 11, 958–967. [Google Scholar] [CrossRef]
- Selvaggi, C.; Pierangeli, A. Interferon lambda 1e3 expression in infants hospitalized for RSV or HRV associated bronchiolitis. J. Infect. 2014, 68, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Hijano, D.R.; Vu, L.D. Role of Type I Interferon (IFN) in the Respiratory Syncytial Virus (RSV) Immune Response and Disease Severity. Front. Immunol. 2019, 10, 566. [Google Scholar] [CrossRef]
- He, L.; Yang, L. Efficacy and safety of interferon on neonates with respiratory syncytial virus pneumonia. Exp. Ther. Med. 2020, 20, 220. [Google Scholar] [CrossRef] [PubMed]
- Hadjadj, J.; Yatim, N. Impaired type I interferon activity and inflammatory responses in severe COVID-19 patients. Science 2020, 369, 718–724. [Google Scholar] [CrossRef] [PubMed]
- da Silva, R.P.; Gonçalves, J.I.B. Circulating Type I Interferon Levels and COVID-19 Severity: A Systematic Review and Meta-Analysis. Front. Immunol. 2021, 12, 657363. [Google Scholar] [CrossRef]
- Yoshida, M.; Worlock, K.B. Local and systemic responses to SARS-CoV-2 infection in children and adults. Nature 2022, 602, 321–327. [Google Scholar] [CrossRef]
- Sposito, B.; Broggi, A. The interferon landscape along the respiratory tract impacts the severity of COVID-19. Cell 2021, 184, 4953–4968. [Google Scholar] [CrossRef]
- Brzoska, J.; von Eick, H. Interferons in COVID-19: Missed opportunities to prove efficacy in clinical phase III trials? Front. Med. 2023, 10, 1198576. [Google Scholar] [CrossRef]
- Reis, G.; Moreira Silva, E.A.S. Early Treatment with Pegylated Interferon Lambda for Covid-19. N. Engl. J. Med. 2023, 388, 518–528. [Google Scholar] [CrossRef]
- González-Sanz, R.; Mata, M. ISG15 Is Upregulated in Respiratory Syncytial Virus Infection and Reduces Virus Growth through Protein ISGylation. J. Virol. 2016, 90, 3428–3438. [Google Scholar] [CrossRef]
- Wydorski, P.M.; Osipiuk, J. Dual domain recognition determines SARS-CoV-2 PLpro selectivity for human ISG15 and K48-linked di-ubiquitin. Nat. Commun. 2023, 14, 2366. [Google Scholar] [CrossRef]
- Martin-Sancho, L.; Lewinski, M.K. Functional landscape of SARS-CoV-2 cellular restriction. Mol. Cell 2021, 81, 2656–2668. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Le Pen, J. Inborn errors of OAS–RNase L in SARS-CoV-2–related multisystem inflammatory syndrome in children. Science 2023, 379, eabo3627. [Google Scholar] [CrossRef]
- Chout, J.; Thomas, P.G. Immunology of SARS-CoV-2 infection in children. Nat. Immunol. 2022, 23, 177–185. [Google Scholar]
- Shehata, L.; Wieland-Alter, W.F. Systematic comparison of respiratory syncytial virus-induced memory B cell responses in two anatomical compartments. Nat. Commun. 2019, 10, 1126. [Google Scholar] [CrossRef]
- Vissers, M.; Ahout, I.M. Mucosal IgG Levels Correlate Better with Respiratory Syncytial Virus Load and Inflammation than Plasma IgG Levels. Clin. Vaccine Immunol. 2015, 23, 243–245. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, E.; Gilman, M.S.A. Infants Infected with Respiratory Syncytial Virus Generate Potent Neutralizing Antibodies that Lack Somatic Hypermutation. Immunity 2018, 48, 339–349.e5. [Google Scholar] [CrossRef]
- Zhivaki, D.; Lemoine, S. Respiratory Syncytial Virus Infects Regulatory B Cells in Human Neonates via Chemokine Receptor CX3CR1 and Promotes Lung Disease Severity. Immunity 2017, 46, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Legg, J.P.; Hussain, I.R. Type 1 and type 2 cytokine imbalance in acute respiratory syncytial virus bronchiolitis. Am. J. Respir. Crit. Care Med. 2003, 168, 633–639. [Google Scholar] [CrossRef]
- Russell, C.D.; Unger, S.A. The Human Immune Response to Respiratory Syncytial Virus Infection. Clin. Microbiol. Rev. 2017, 30, 481–502. [Google Scholar] [CrossRef] [PubMed]
- Siefker, D.T.; Vu, L. Respiratory Syncytial Virus Disease Severity Is Associated with Distinct CD8+ T-Cell Profiles. Am. J. Respir. Crit. Care Med. 2020, 201, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Chirkova, T.; Rosas-Salazar, C. Effect of Infant RSV Infection on Memory T Cell Responses at Age 2-3 Years. Front. Immunol. 2022, 13, 826666. [Google Scholar] [CrossRef] [PubMed]
- Mella, C.; Suarez-Arrabal, C. Innate Immune Dysfunction is Associated with Enhanced Disease Severity In Infants with Severe Respiratory Syncytial Virus Bronchiolitis. J. Infect. Dis. 2013, 207, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Johansson, C. Respiratory syncytial virus infection: An innate perspective [version 1; referees: 4 approved]. F1000Research 2016, 5, 2898. [Google Scholar] [CrossRef] [PubMed]
- Toh, Z.Q.; Anderson, J. Comparison of Seroconversion in Children and Adults With Mild COVID-19. JAMA Netw. Open 2022, 5, e221313. [Google Scholar] [CrossRef]
- Corr, M.; Christie, S. Seroprevalence of SARS-CoV-2 antibodies in children of United Kingdom healthcare workers: A prospective multicentre cohort study protocol. BMJ Open 2020, 10, e041661. [Google Scholar] [CrossRef]
- Vella, L.A.; Giles, J.R. Deep immune profiling of MIS-C demonstrates marked but transient immune activation compared with adult and pediatric COVID-19. Sci. Immunol. 2021, 6, eabf7570. [Google Scholar] [CrossRef]
- Khoo, W.H.; Jackson, K. Tracking the clonal dynamics of SARS-CoV-2-specific T cells in children and adults with mild/asymptomatic COVID-19. Clin. Immunol. 2023, 246, 109209. [Google Scholar] [CrossRef]
- Lima, K.; Fontoura, J.C. SARS-CoV-2 infected children form early immune memory responses dominated by nucleocapsid-specific CD8+ T cells and antibodies. Front. Immunol. 2022, 13, 1033364. [Google Scholar] [CrossRef]
- Xu, Q.; Milanez-Almeida, P.; Martins, A.J.; Radtke, A.J.; Hoehn, K.B.; Oguz, C.; Chen, J.; Liu, C.; Tang, J.; Grubbs, G.; et al. Adaptive immune responses to SARS-CoV-2 persist in the pharyngeal lymphoid tissue of children. Nat. Immunol. 2023, 24, 186–199. [Google Scholar] [CrossRef]
- Renk, H.; Dulovic, A.; Seidel, A.; Becker, M.; Fabricius, D.; Zernickel, M.; Junker, D.; Groß, R.; Müller, J.; Hilger, A.; et al. Robust and durable serological response following pediatric SARS-CoV-2 infection. Nat. Commun. 2022, 13, 128. [Google Scholar] [CrossRef] [PubMed]
- Reguera, J.; Santiago, C. Structural bases of coronavirus attachment to host aminopeptidase N and its inhibition by neutralizing antibodies. PLoS Pathog. 2012, 8, e1002859. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Zheng, Q. The SARS-CoV-2 spike glycoprotein biosynthesis, structure, function, and antigenicity: Implications for the design of spikebased vaccine immunogens. Front. Immunol. 2020, 11, 576622. [Google Scholar] [CrossRef]
- Dubey, A.; Choudhary, S.; Kumar, P. Emerging SARS-CoV-2 variants: Genetic variability and clinical implications. Curr. Microbiol. 2022, 79, 20. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, K. The profile of peripheral blood lymphocyte subsets and serum cytokines in children with 2019 novel coronavirus pneumonia. J. Infect. 2020, 81, 115–120. [Google Scholar] [CrossRef]
- Jia, R.; Wang, X. Mild Cytokine Elevation, Moderate CD4+ T Cell Response and Abundant Antibody Production in Children with COVID-19. Virol. Sin. 2020, 35, 734–743. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| RSV Infection | SARS-CoV-2 Infection | |
|---|---|---|
| IFN-λ1 and IFN-λ3 | IFN-λ1 and IFN-λ3 are associated with the severity of respiratory disease caused by RSV | IFN-λ1 and IFN-λ3 are associated with the induction of ISGs, efficiently containing SARS-CoV-2 |
| RIG-I and MDA5 | NS1 protein inhibits ubiquitination of RIG-I and MDA5 | NSP7, NSP5, and ORF9b interact with RIG-I and MDA5, preventing the interaction with MAVS |
| IFN-I | IFN-I response is impaired during RSV infection | IFN-I response is impaired during SARS-CoV-2 infection |
| IFN-III | IFN-III response is impaired during RSV infection | IFN-III response is impaired during SARS-CoV-2 infection |
| OAS1 | OAS1 is significantly expressed in individuals with symptomatic RSV | OAS1 is deficient in children who have presented severe cases of COVID-19 |
| ISG15 | ISG15 is upregulated during RSV infection | SARS-CoV-2 tries to interfere with ISG15 function |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, R.P.; Thomé, B.L.; da Souza, A.P.D. Exploring the Immune Response against RSV and SARS-CoV-2 Infection in Children. Biology 2023, 12, 1223. https://doi.org/10.3390/biology12091223
da Silva RP, Thomé BL, da Souza APD. Exploring the Immune Response against RSV and SARS-CoV-2 Infection in Children. Biology. 2023; 12(9):1223. https://doi.org/10.3390/biology12091223
Chicago/Turabian Styleda Silva, Rafaela Pires, Bibiana Liberman Thomé, and Ana Paula Duarte da Souza. 2023. "Exploring the Immune Response against RSV and SARS-CoV-2 Infection in Children" Biology 12, no. 9: 1223. https://doi.org/10.3390/biology12091223
APA Styleda Silva, R. P., Thomé, B. L., & da Souza, A. P. D. (2023). Exploring the Immune Response against RSV and SARS-CoV-2 Infection in Children. Biology, 12(9), 1223. https://doi.org/10.3390/biology12091223