



Characterization of the Intracellular Acidity Regulation of Brain Tumor Cells and Consequences for Therapeutic Optimization of Temozolomide
 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Microscope
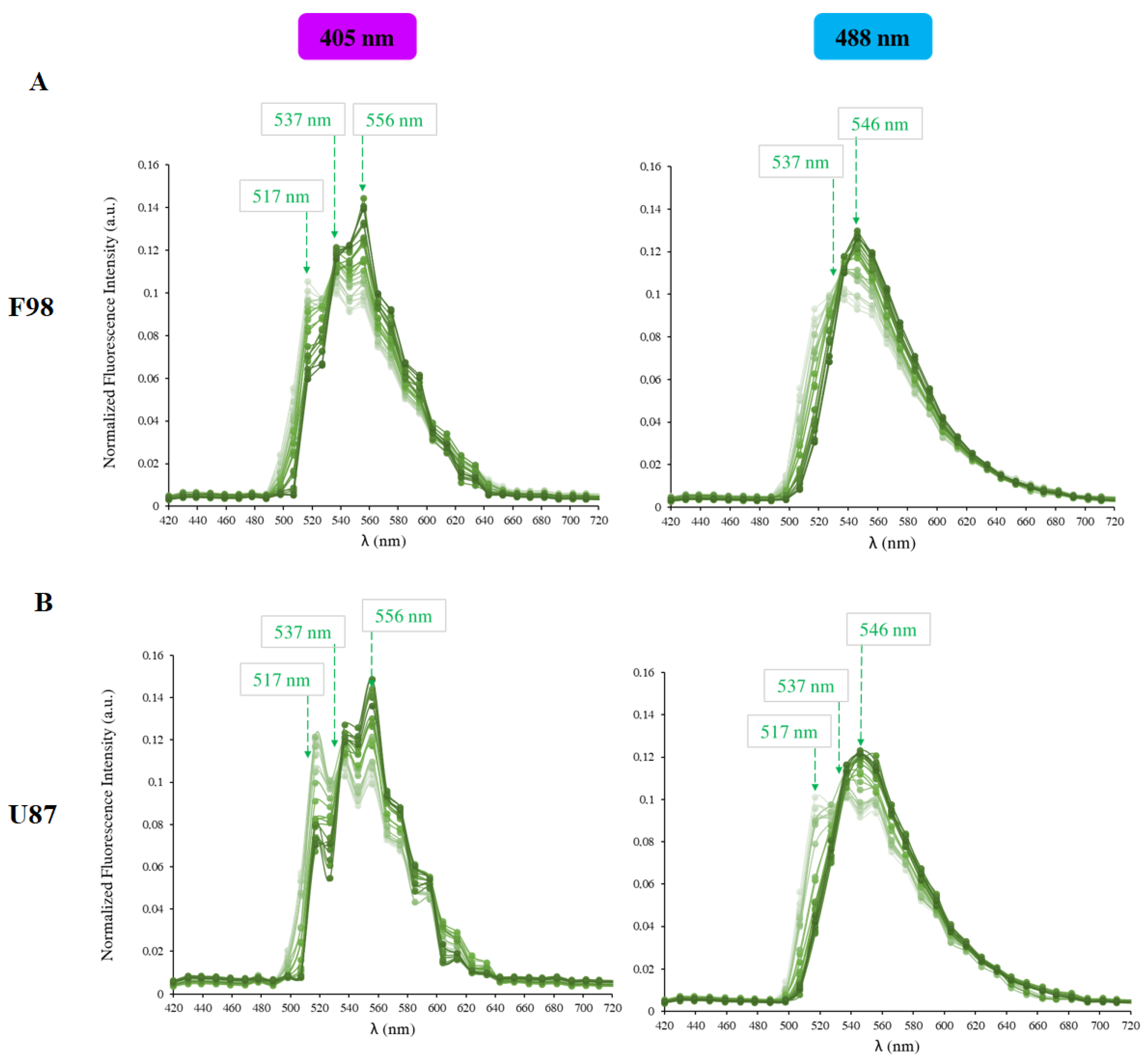
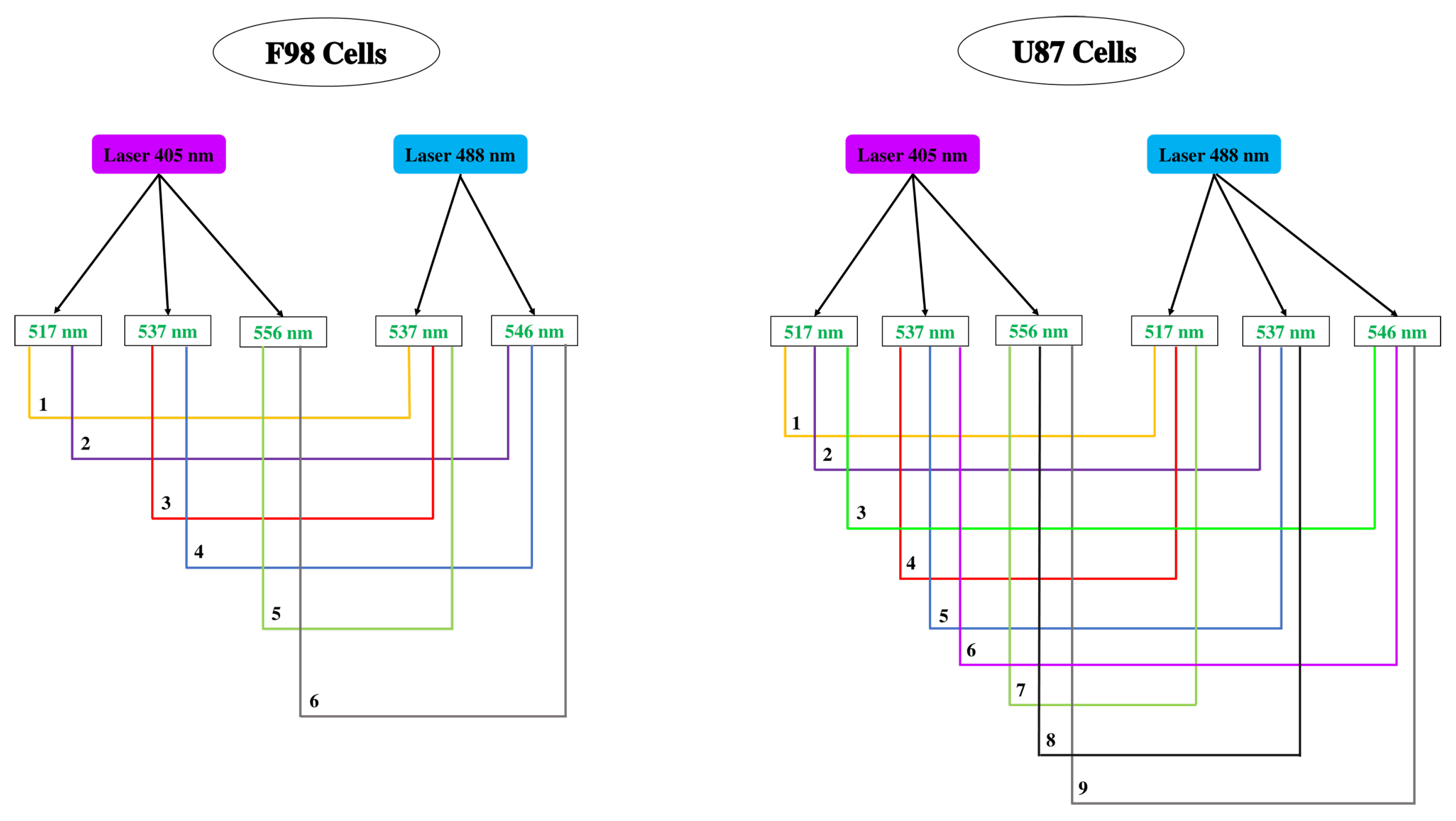
2.3. pH Measurements
2.4. Loading Cells with pH Probes
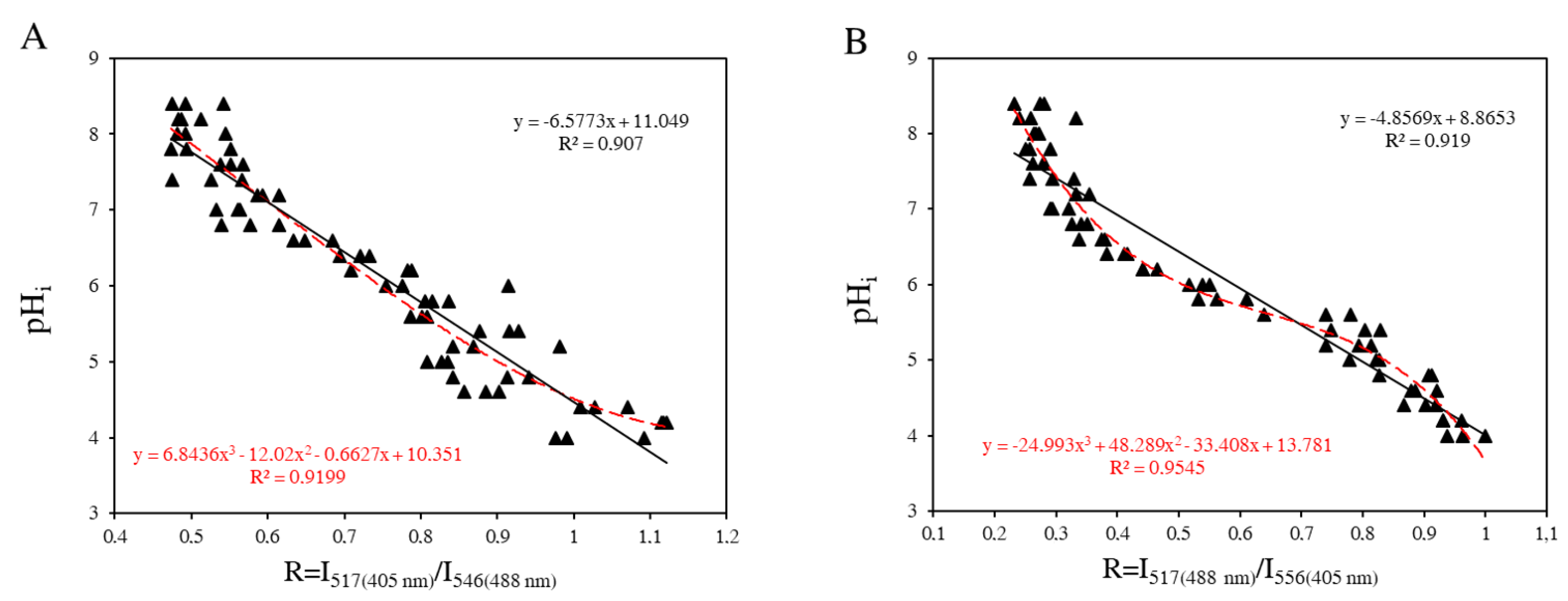
2.5. Intracellular pH Calibration
2.6. pH Measurements in Spheroids
2.7. Assessment of Cell Resistance to Acidity
2.8. Measure of Cell Viability after TMZ Treatment
2.9. Evaluation of IC of TMZ
2.10. TMZ Treatment
2.11. Statistical Analysis
3. Results
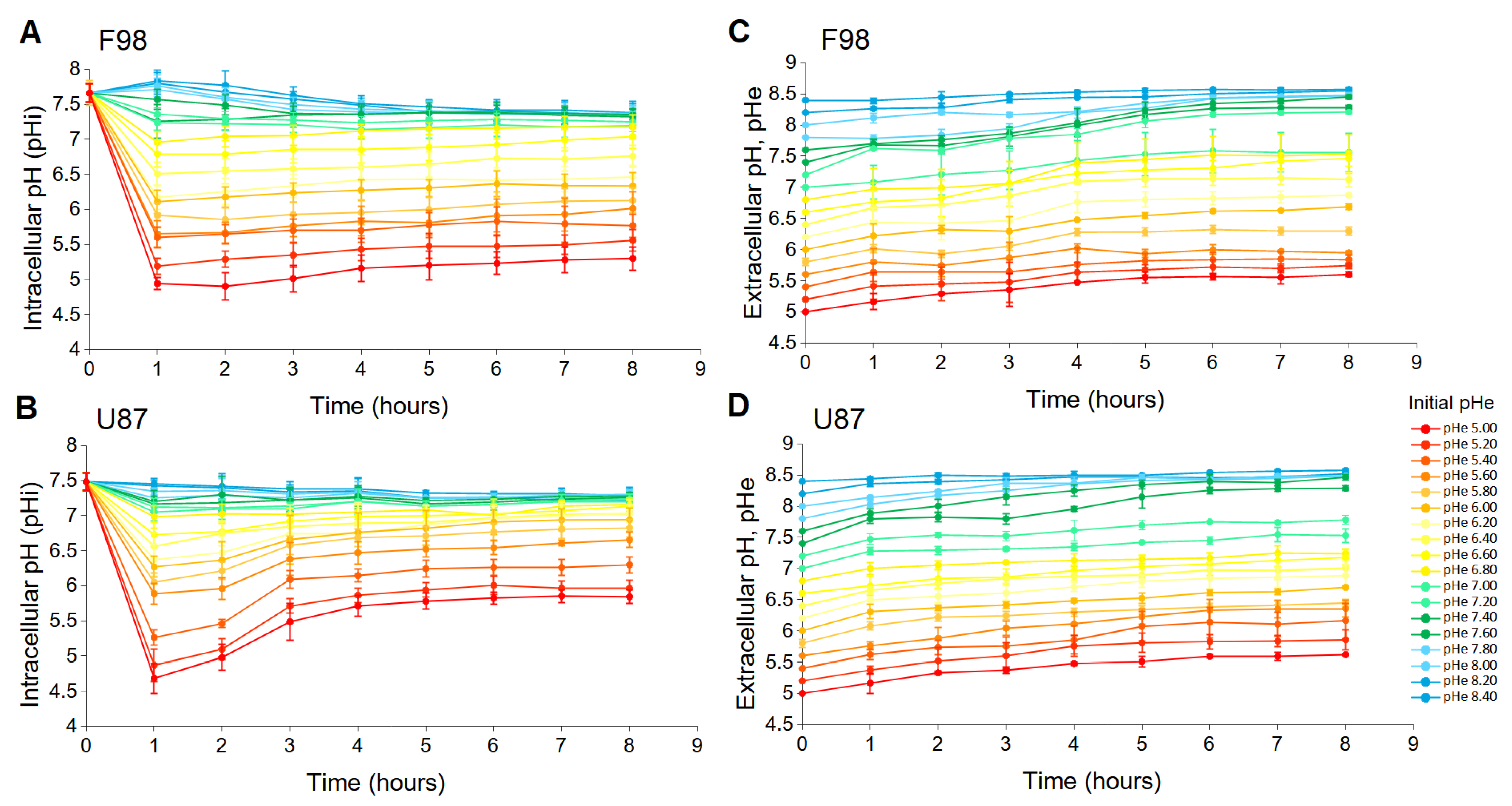
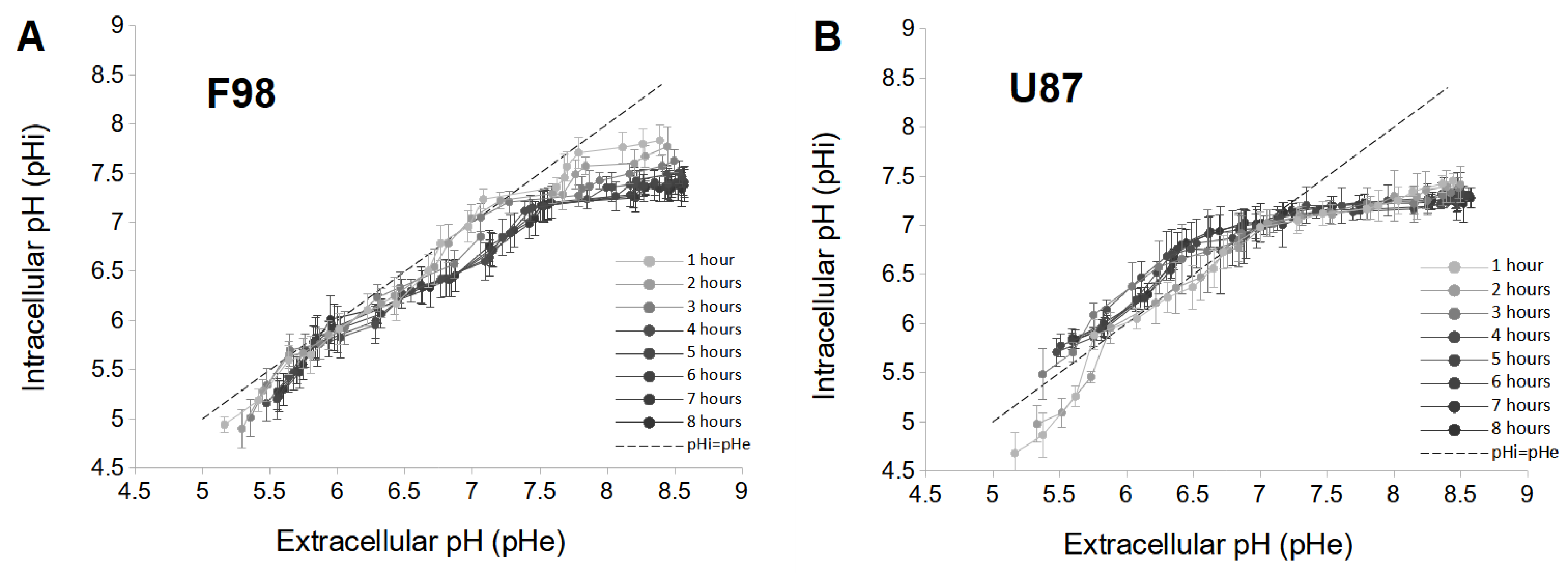
3.1. Intracellular pH Changes in Cell Cultures
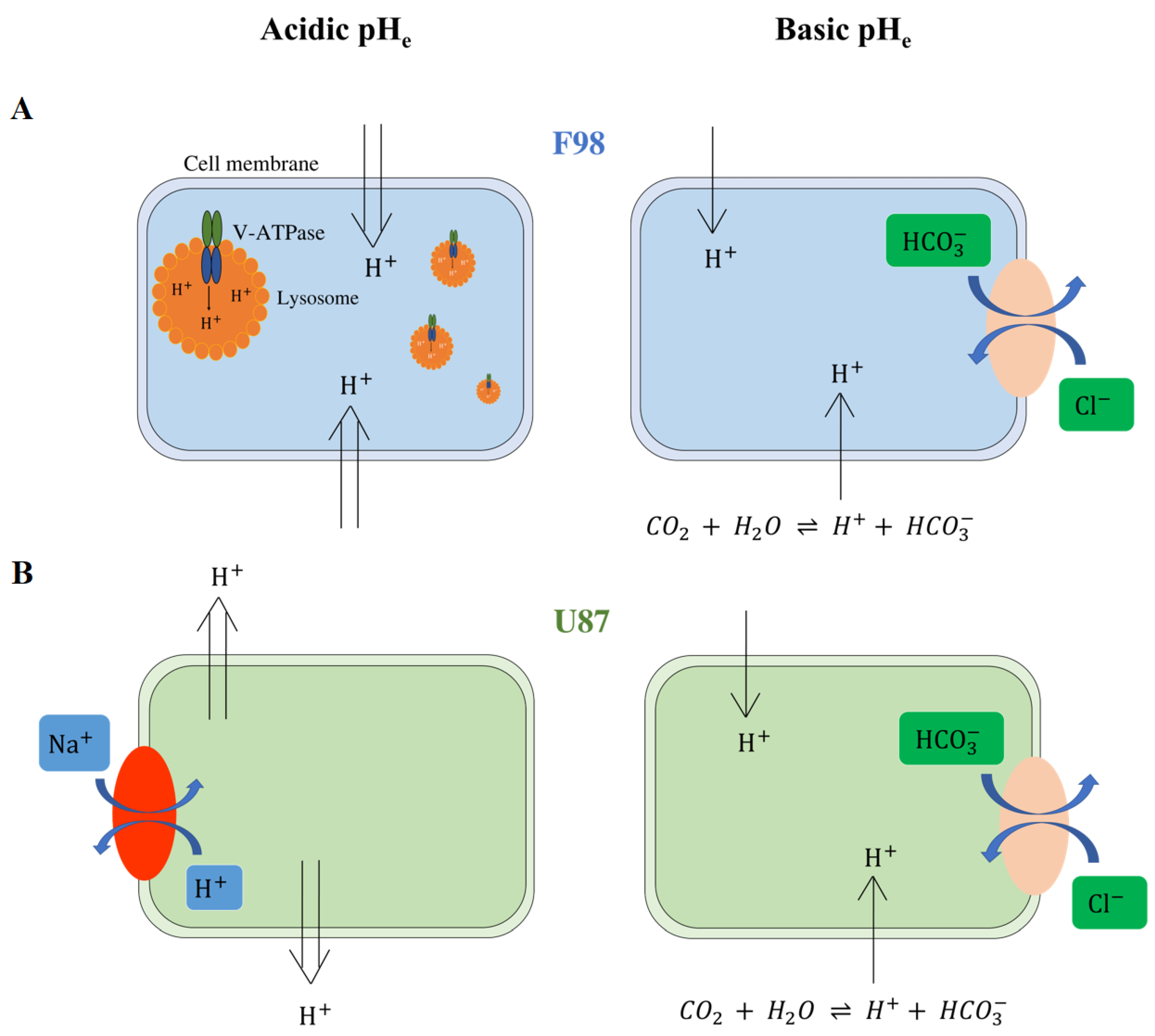
3.1.1. pH Changes in F98 Cells
3.1.2. pH Changes in U87 Cells
3.1.3. Interpretation
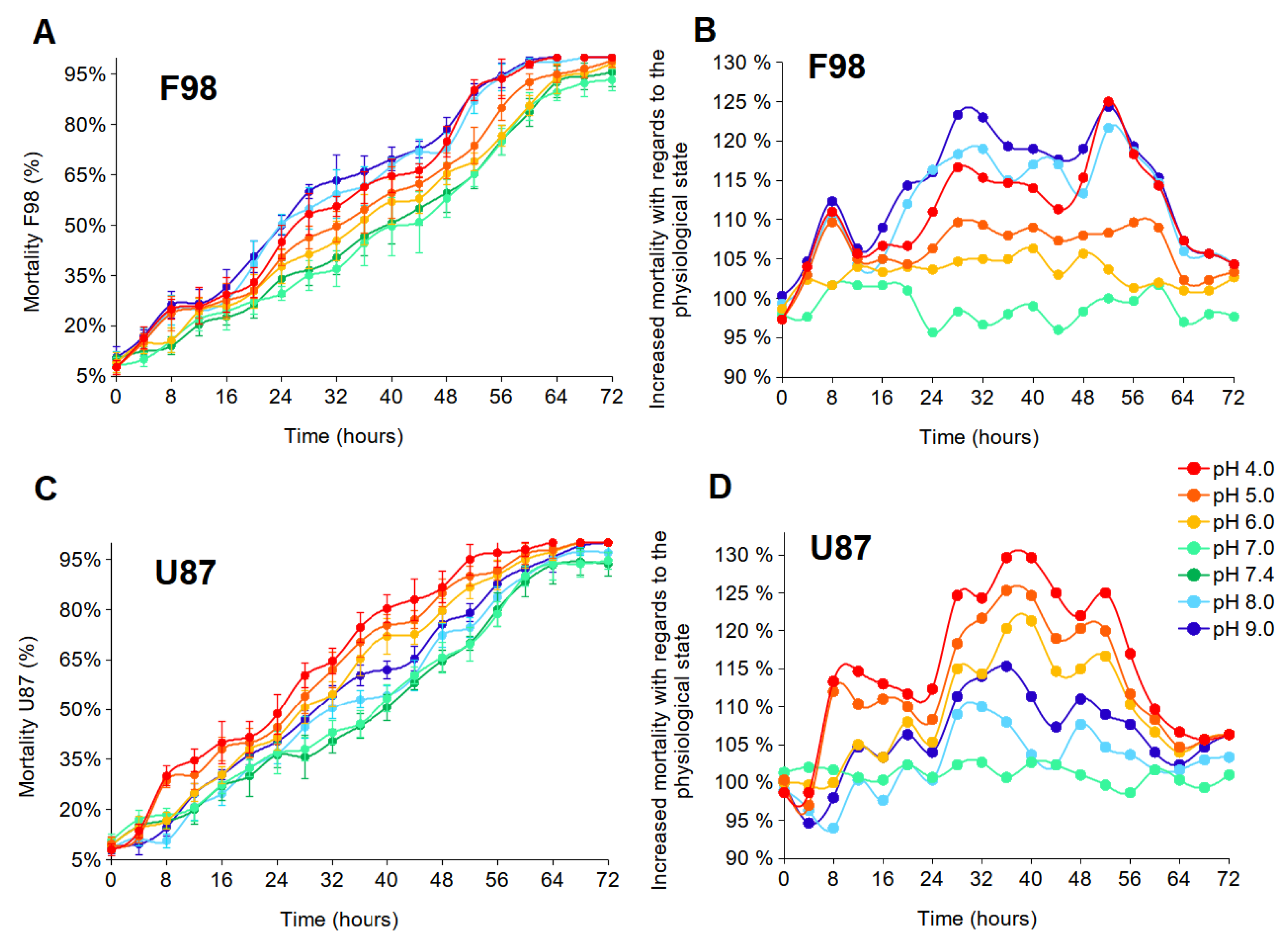
3.2. Resistance to Extracellular pH Changes
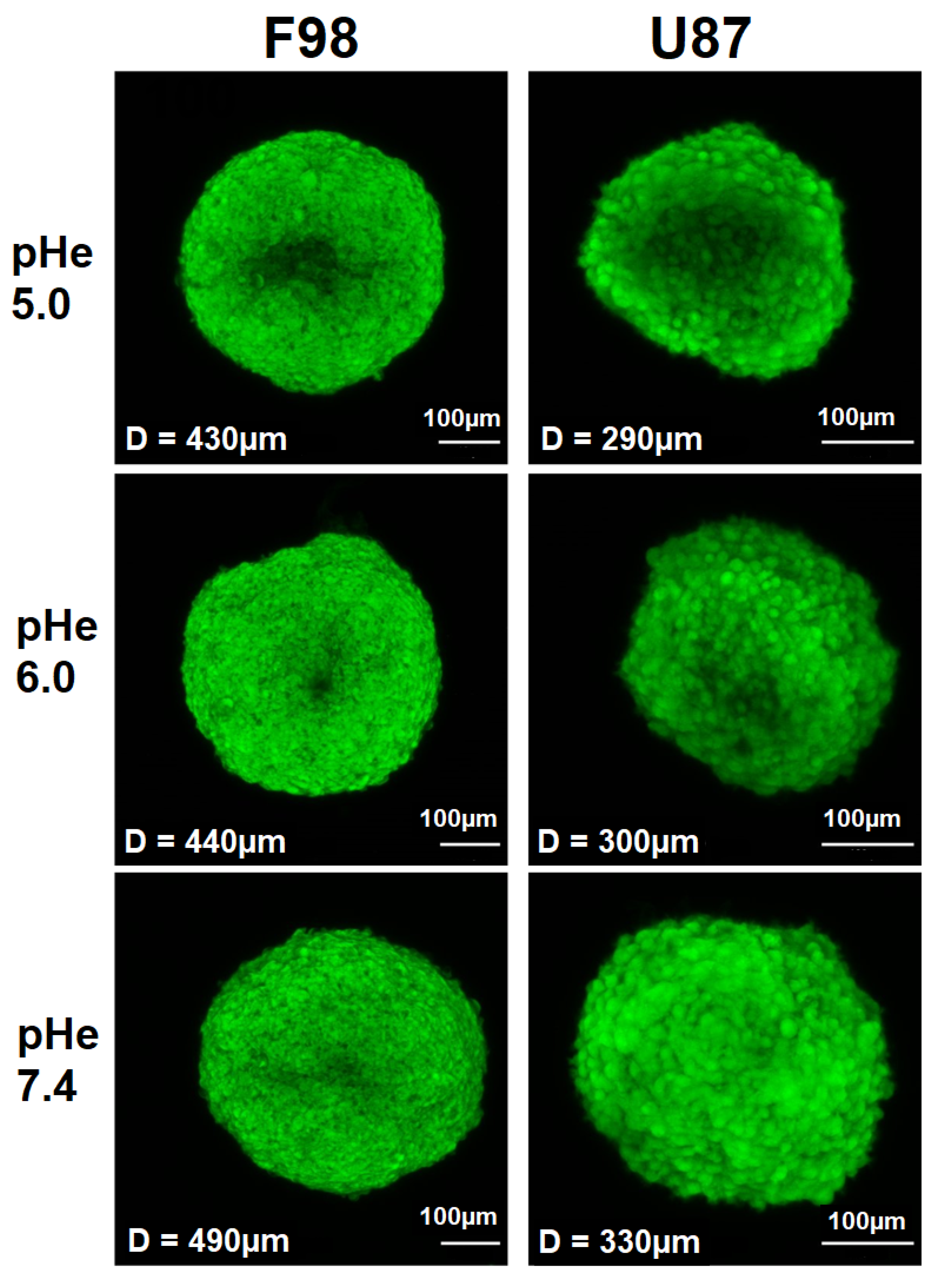
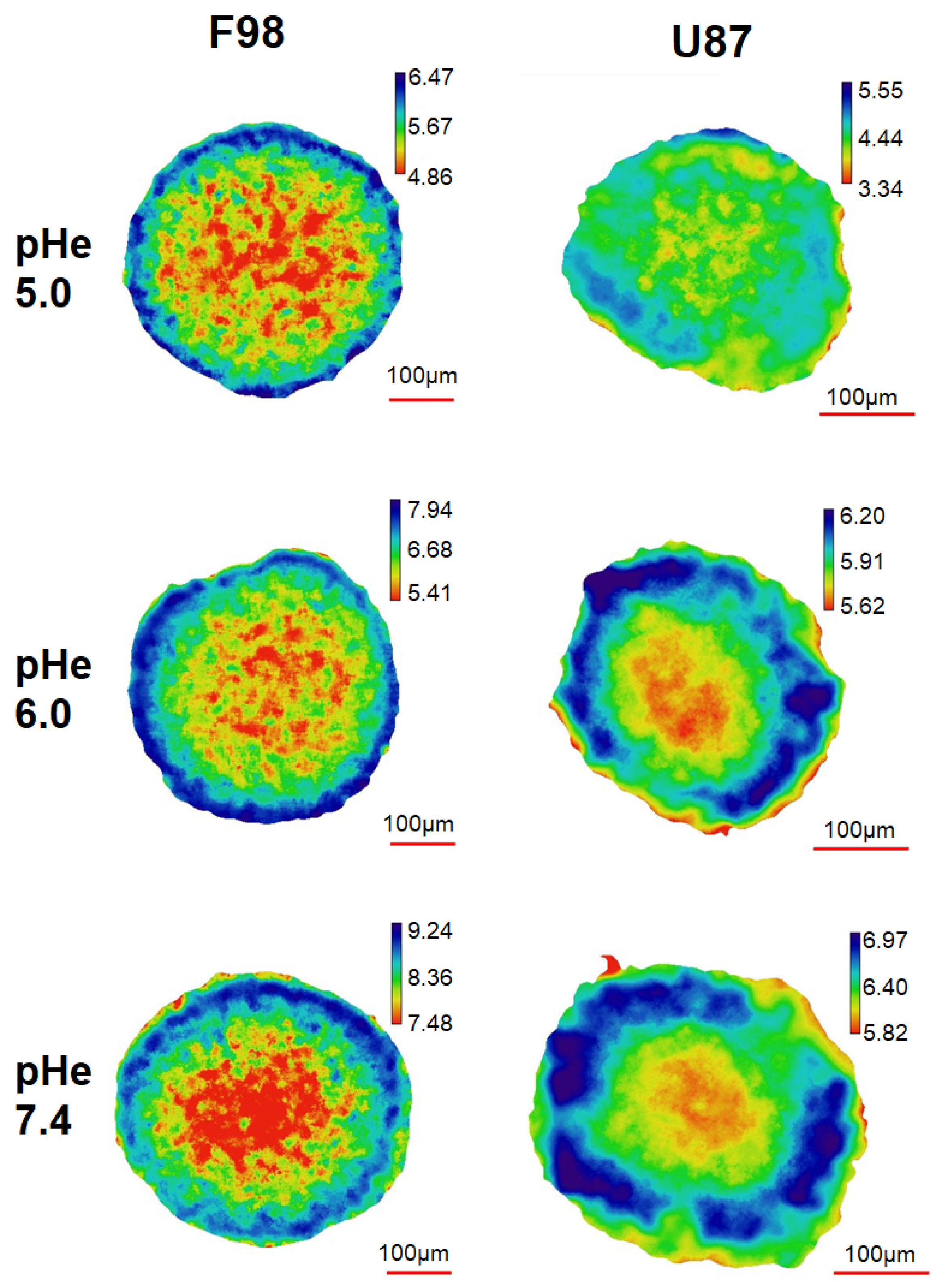
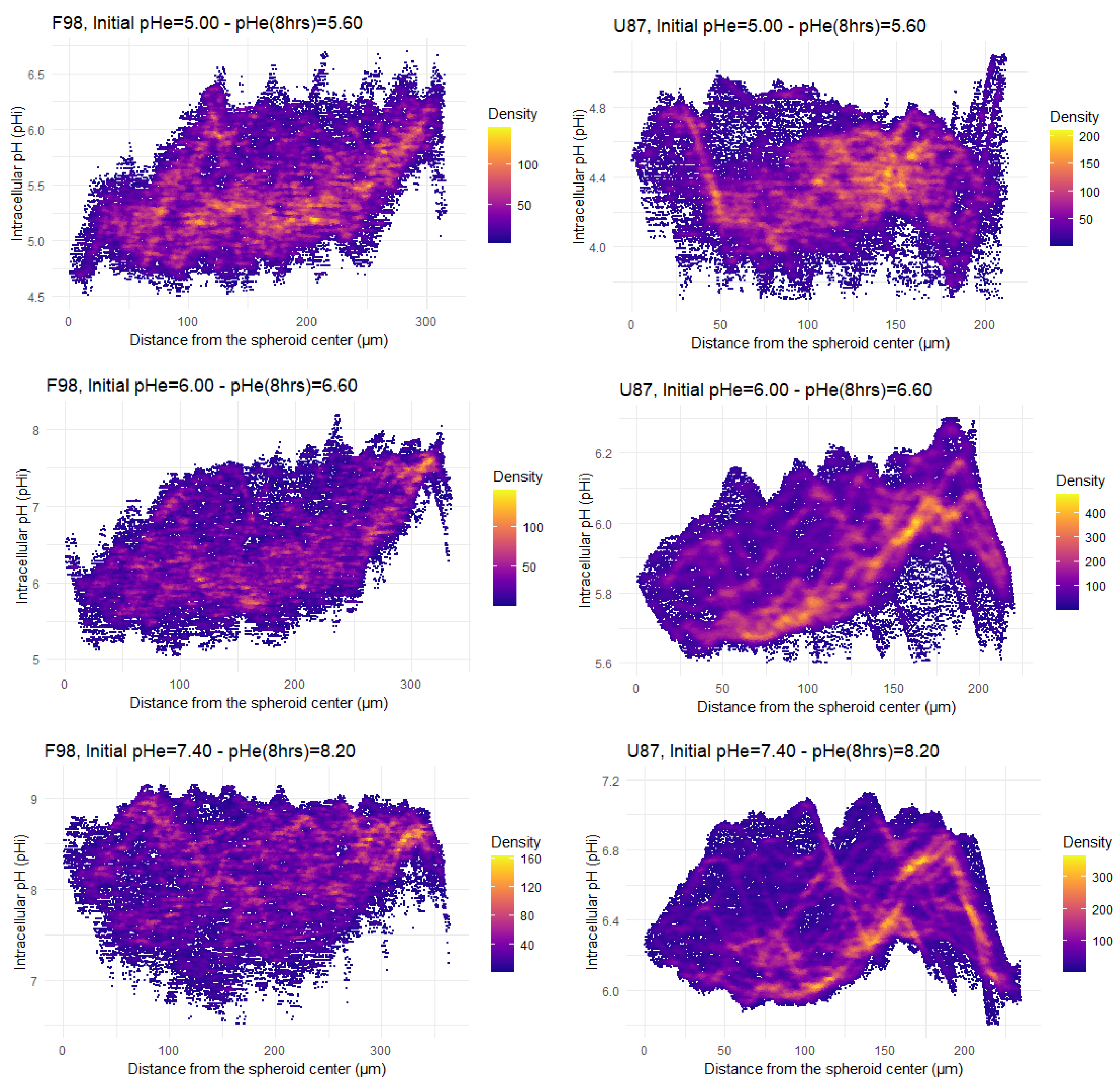
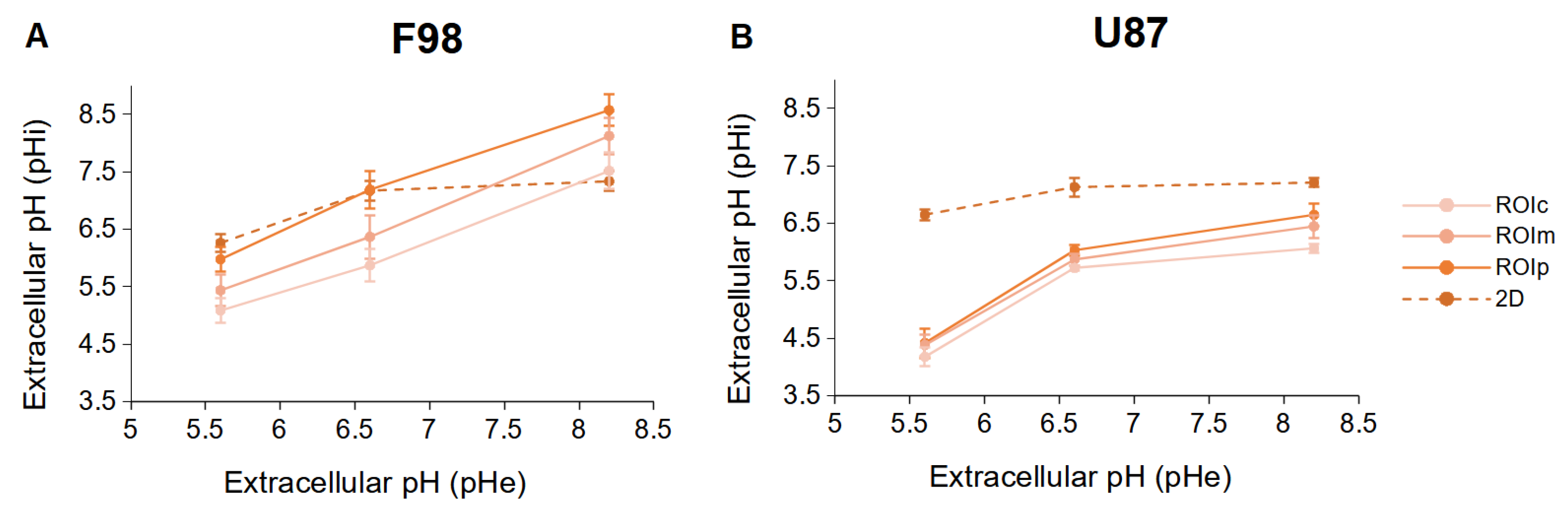
3.3. Intracellular pH in Spheroids
3.3.1. pH Distribution
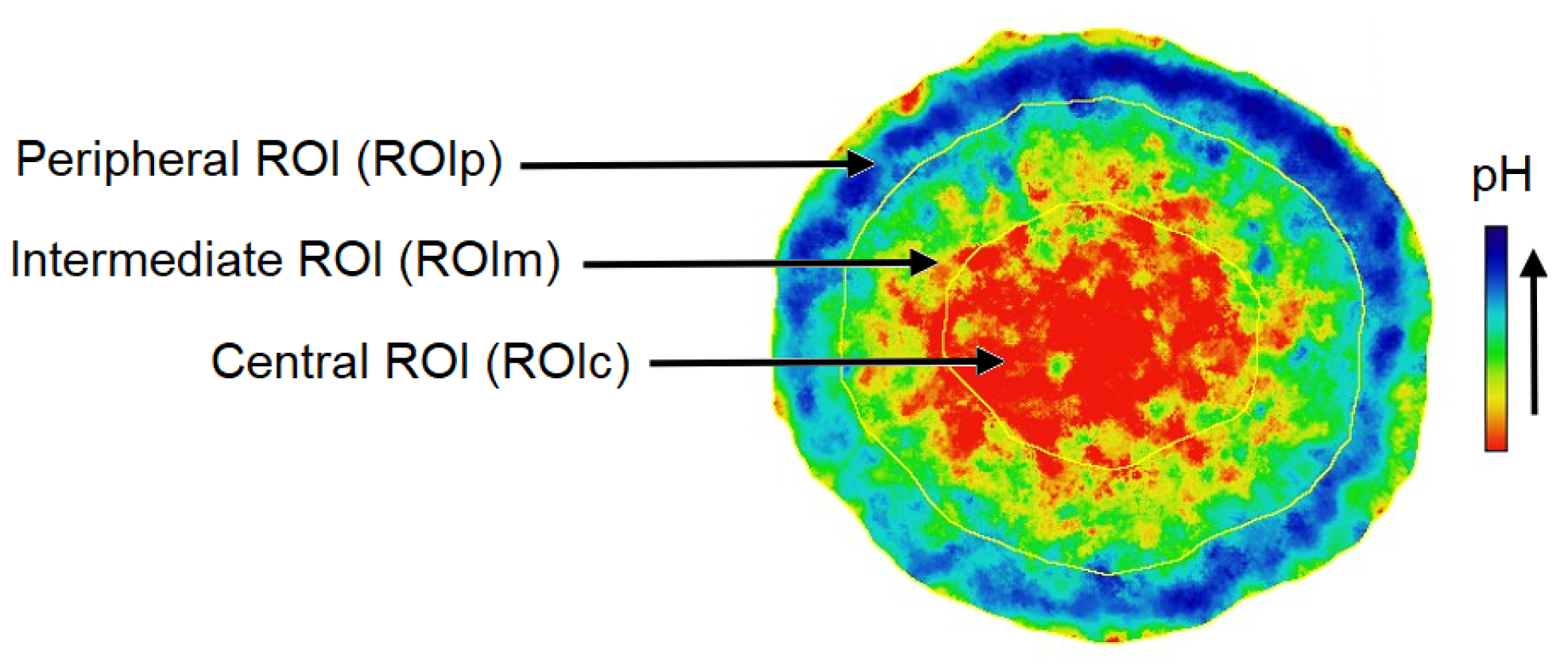
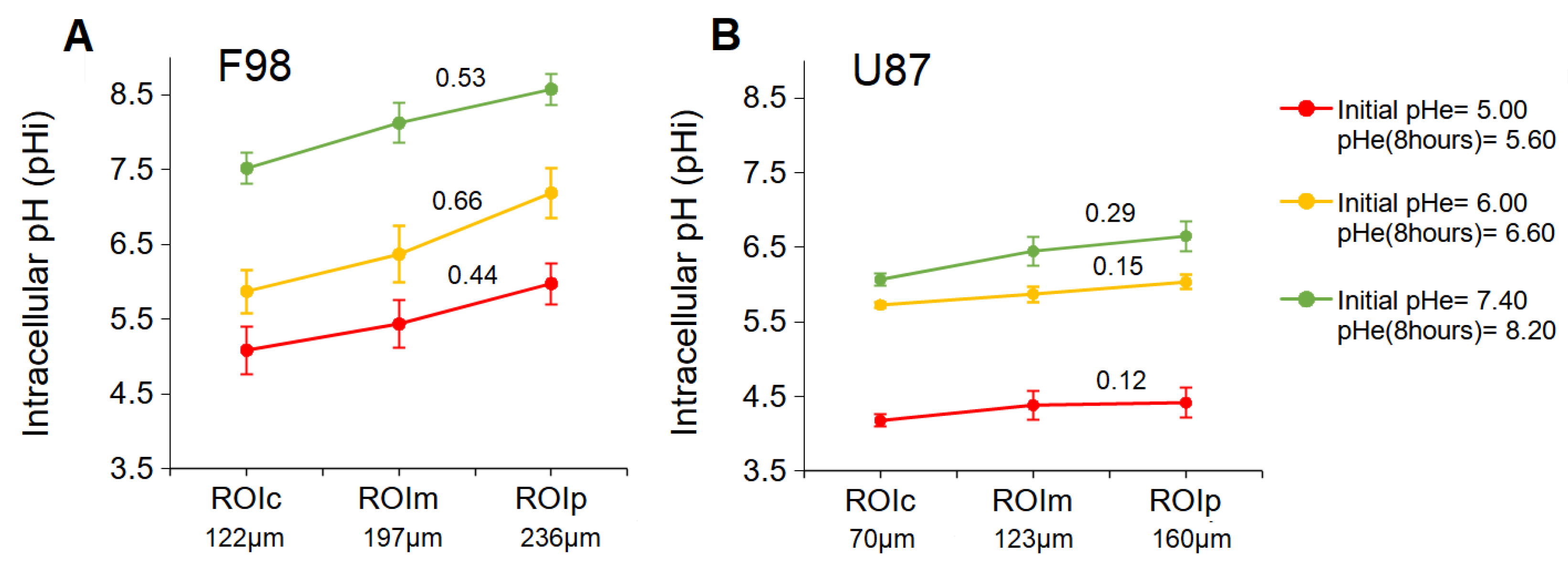
3.3.2. pH in ROI
3.3.3. 2D vs. 3D Cultures
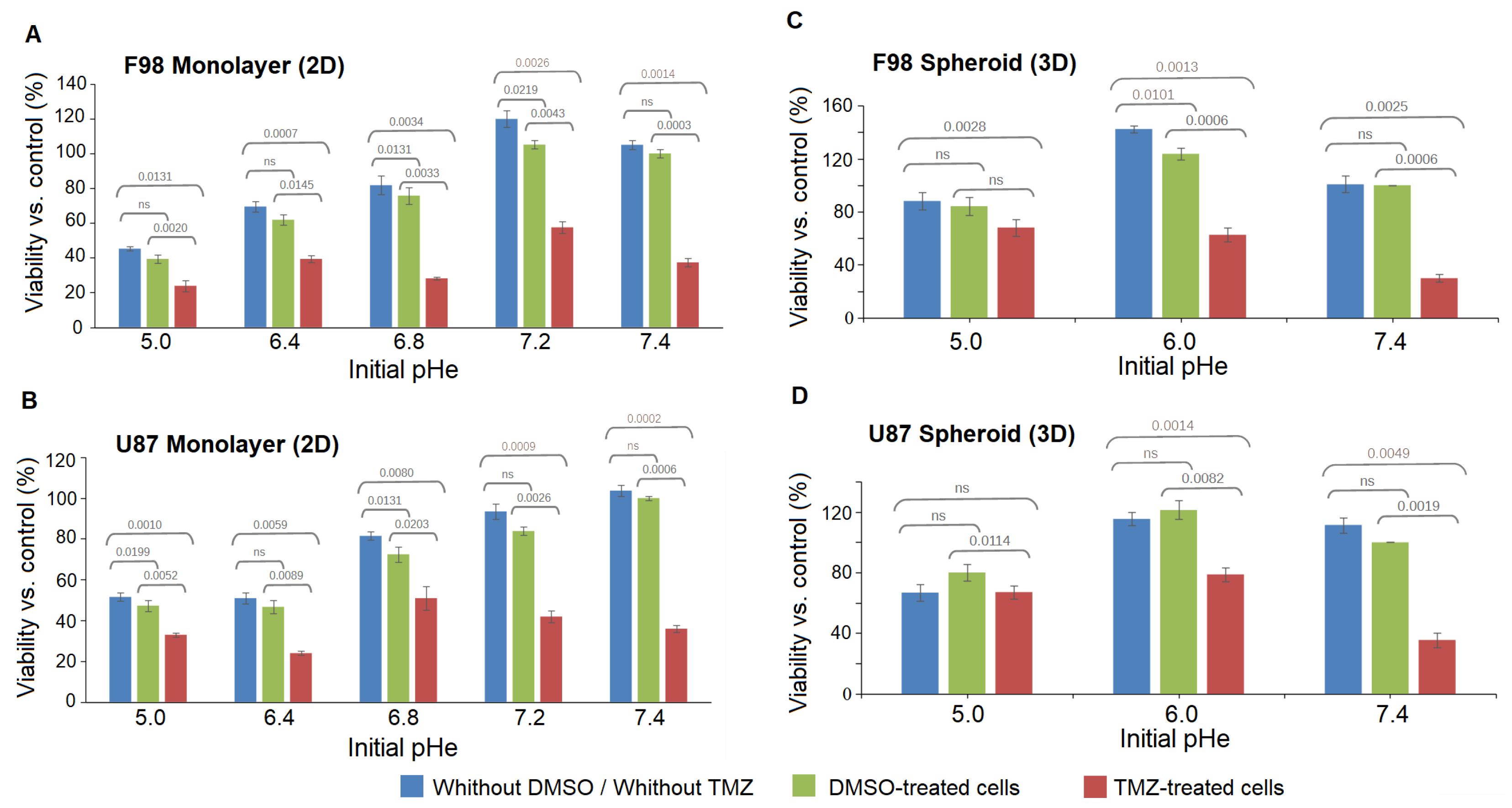
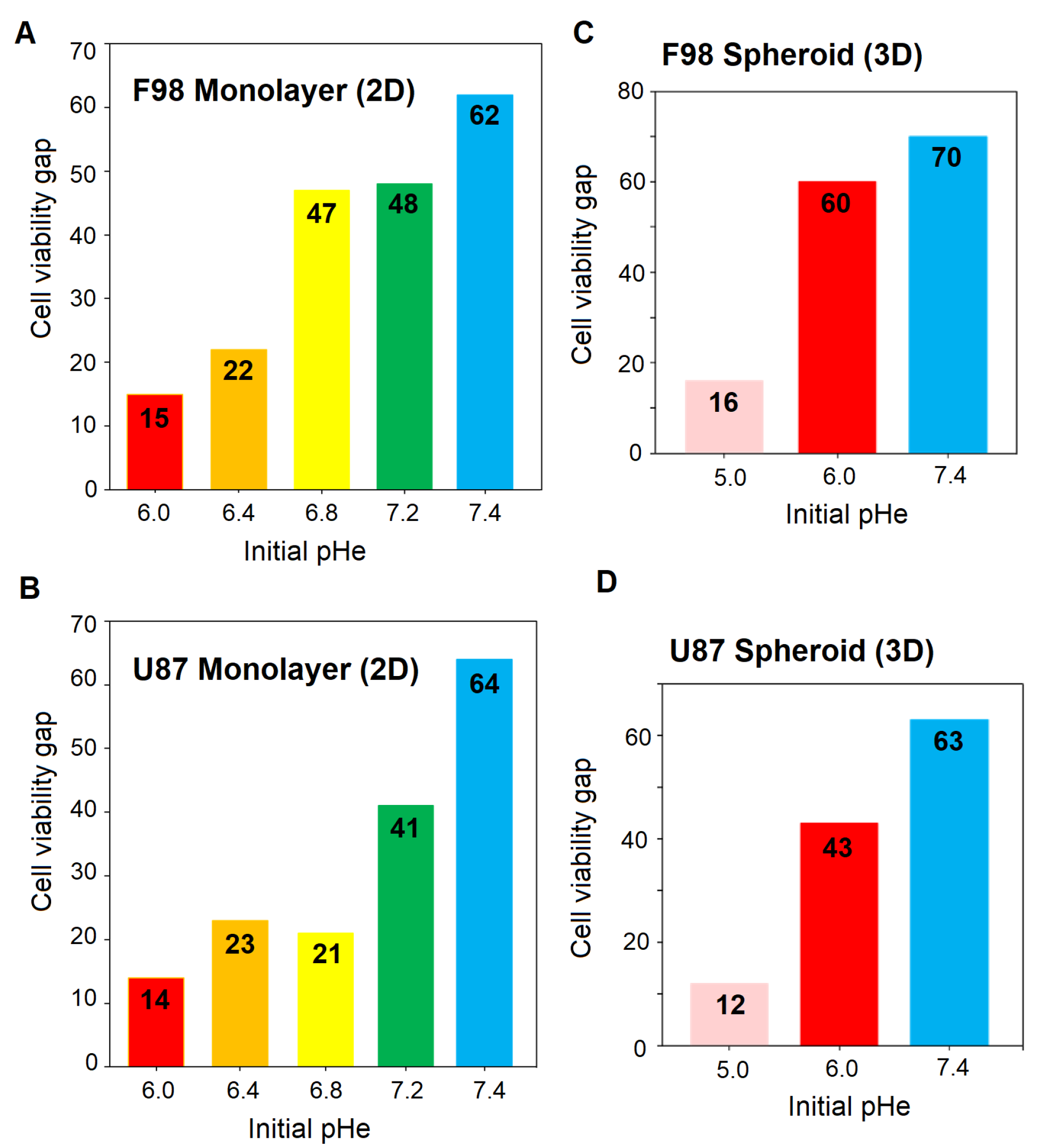
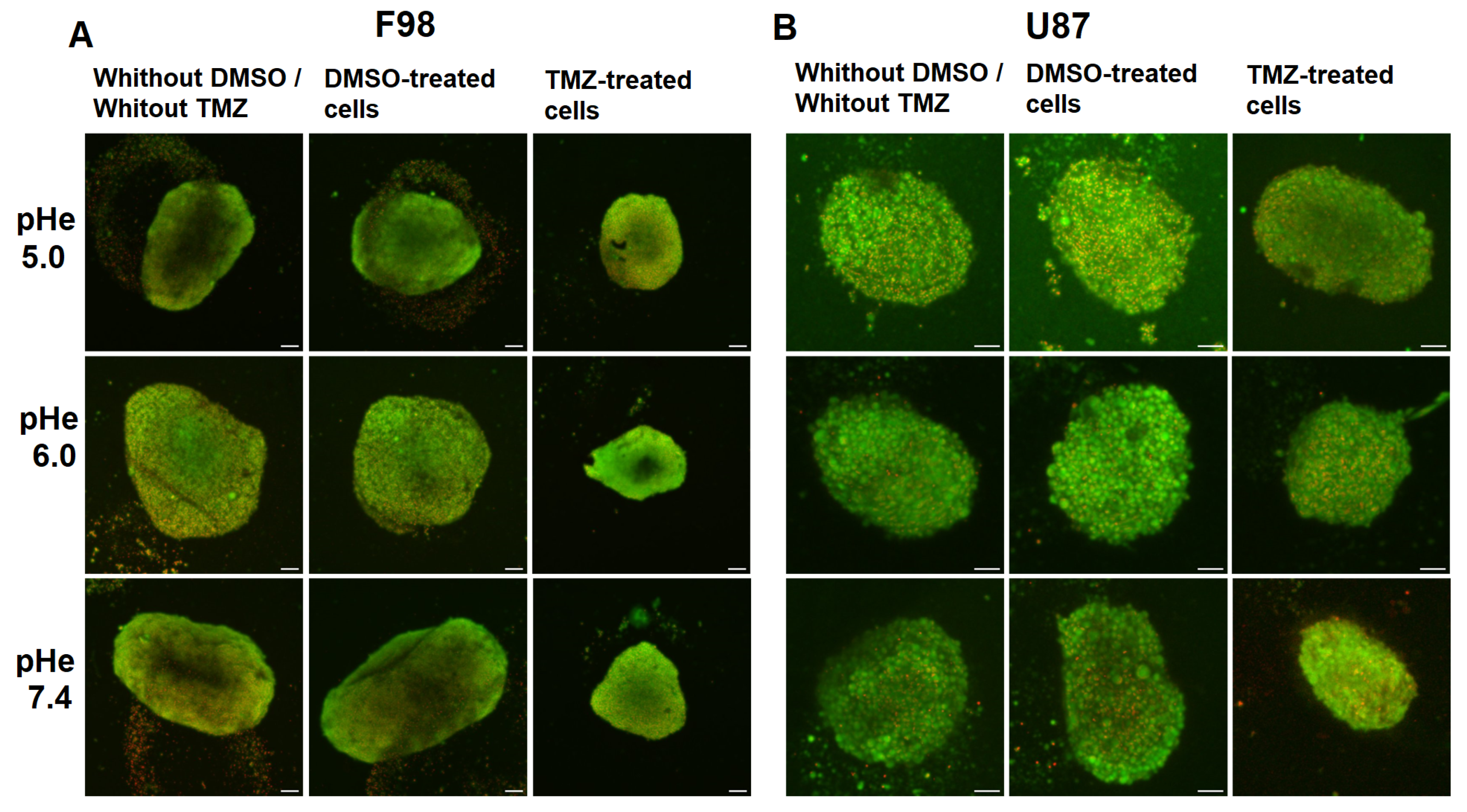
3.4. Effect of pH on Temozolomide Efficacy
Toxicity in Spheroids
4. Discussion
Conclusions and Future Work
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Vaupel, P.; Kallinowski, F.; Okunieff, P. Blood flow, oxygen and nutrient supply, and metabolic microenvironment of human tumors: A review. Cancer Res. 1989, 49, 6449–6465. [Google Scholar] [PubMed]
- Kallinowski, F.; Vaupel, P. pH distributions in spontaneous and isotransplanted rat tumours. Br. J. Cancer 1988, 58, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Chiche, J.; Brahimi-Horn, M.C.; Pouysségur, J. Tumour hypoxia induces a metabolic shift causing acidosis: A common feature in cancer. J. Cell. Mol. Med. 2010, 14, 771–794. [Google Scholar] [CrossRef] [PubMed]
- Smallbone, K.; Gavaghan, D.J.; Gatenby, R.A.; Maini, P.K. The role of acidity in solid tumour growth and invasion. J. Theor. Biol. 2005, 235, 476–484. [Google Scholar] [CrossRef]
- Justus, C.R.; Dong, L.; Yang, L.V. Acidic tumor microenvironment and pH-sensing G protein-coupled receptors. Front. Physiol. 2013, 4, 354. [Google Scholar] [CrossRef]
- Parks, S.K.; Cormerais, Y.; Pouysségur, J. Hypoxia and cellular metabolism in tumour pathophysiology. J. Physiol. 2017, 595, 2439–2450. [Google Scholar] [CrossRef]
- Druzhkova, I.; Shirmanova, M.; Lukina, M.; Dudenkova, V.; Sergeeva, T.; Belousov, V.; Lukyanov, S.; Zagaynova, E. Registration of intracellular pH in cancer cells with genetically encoded ratiometric sensor. In European Conference on Biomedical Optics; Optica Publishing Group: Washington, DC, USA, 2015. [Google Scholar] [CrossRef]
- Shirmanova, M.V.; Druzhkova, I.N.; Lukina, M.M.; Matlashov, M.E.; Belousov, V.V.; Snopova, L.B.; Prodanetz, N.N.; Dudenkova, V.V.; Lukyanov, S.A.; Zagaynova, E.V. Intracellular pH imaging in cancer cells in vitro and tumors in vivo using the new genetically encoded sensor SypHer2. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2015, 1850, 1905–1911. [Google Scholar] [CrossRef]
- Gillies, R.J.; Raghunand, N.; Karczmar, G.S.; Bhujwalla, Z.M. MRI of the tumor microenvironment. J. Magn. Reson. Imaging 2002, 16, 430–450. [Google Scholar] [CrossRef]
- Roos, A.; Boron, W.F. Intracellular pH. Physiol. Rev. 1981, 61, 296–434. [Google Scholar] [CrossRef]
- Madshus, I.H. Regulation of intracellular pH in eukaryotic cells. Biochem. J. 1988, 250, 1–8. [Google Scholar] [CrossRef]
- Zhuang, Y.X.; Cragoe, E.J.; Shaikewitz, T.; Glaser, L.; Cassel, D. Characterization of potent sodium/proton exchange inhibitors from the amiloride series in A431 cells. Biochemistry 1984, 23, 4481–4488. [Google Scholar] [CrossRef] [PubMed]
- Barathova, M.; Takacova, M.; Holotnakova, T.; Gibadulinova, A.; Ohradanova, A.; Zatovicova, M.; Hulikova, A.; Kopacek, J.; Parkkila, S.; Supuran, C.T.; et al. Alternative splicing variant of the hypoxia marker carbonic anhydrase IX expressed independently of hypoxia and tumour phenotype. Br. J. Cancer 2007, 98, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Hinton, A.; Sennoune, S.R.; Bond, S.; Fang, M.; Reuveni, M.; Sahagian, G.G.; Jay, D.; Martinez-Zaguilan, R.; Forgac, M. Function of a Subunit Isoforms of the V-ATPase in pH Homeostasis and in Vitro Invasion of MDA-MB231 Human Breast Cancer Cells. J. Biol. Chem. 2009, 284, 16400–16408. [Google Scholar] [CrossRef] [PubMed]
- Enerson, B.E.; Drewes, L.R. Molecular Features, Regulation, and Function of Monocarboxylate Transporters: Implications for Drug Delivery. J. Pharm. Sci. 2003, 92, 1531–1544. [Google Scholar] [CrossRef]
- Supuran, C.T.; Fiore, A.D.; Alterio, V.; Montib, S.M.; Simone, G.D. Recent Advances in Structural Studies of the Carbonic Anhydrase Family: The Crystal Structure of Human CA IX and CA XIII. Curr. Pharm. Des. 2010, 16, 3246–3254. [Google Scholar] [CrossRef] [PubMed]
- Pouysségur, J.; Dayan, F.; Mazure, N.M. Hypoxia signalling in cancer and approaches to enforce tumour regression. Nature 2006, 441, 437–443. [Google Scholar] [CrossRef]
- Sennoune, S.R.; Bakunts, K.; Martínez, G.M.; Chua-Tuan, J.L.; Kebir, Y.; Attaya, M.N.; Martínez-Zaguilán, R. Vacuolar H+-ATPase in human breast cancer cells with distinct metastatic potential: Distribution and functional activity. Am. J.-Physiol.-Cell Physiol. 2004, 286, C1443–C1452. [Google Scholar] [CrossRef]
- Montcourrier, P.; Silver, I.; Farnoud, R.; Bird, I.; Rochefort, H. Breast cancer cells have a high capacity to acidify extracellular milieu by a dual mechanism. Clin. Exp. Metastasis 1997, 15, 382–392. [Google Scholar] [CrossRef]
- Grillon, E.; Farion, R.; Fablet, K.; Waard, M.D.; Tse, C.M.; Donowitz, M.; Rémy, C.; Coles, J.A. The Spatial Organization of Proton and Lactate Transport in a Rat Brain Tumor. PLoS ONE 2011, 6, e17416. [Google Scholar] [CrossRef]
- Kapus, A.; Grinstein, S.; Wasan, S.; Kandasamy, R.; Orlowski, J. Functional characterization of three isoforms of the Na+/H+ exchanger stably expressed in Chinese hamster ovary cells. ATP dependence, osmotic sensitivity, and role in cell proliferation. J. Biol. Chem. 1994, 269, 23544–23552. [Google Scholar] [CrossRef]
- Pouysségur, J.; Chambard, J.C.; Franchi, A.; Paris, S.; Obberghen-Schilling, E.V. Growth factor activation of an amiloride-sensitive Na+/H+ exchange system in quiescent fibroblasts: Coupling to ribosomal protein S6 phosphorylation. Proc. Natl. Acad. Sci. USA 1982, 79, 3935–3939. [Google Scholar] [CrossRef]
- Denker, S.P.; Huang, D.C.; Orlowski, J.; Furthmayr, H.; Barber, D.L. Direct Binding of the Na–H Exchanger NHE1 to ERM Proteins Regulates the Cortical Cytoskeleton and Cell Shape Independently of H+ Translocation. Mol. Cell 2000, 6, 1425–1436. [Google Scholar] [CrossRef] [PubMed]
- Turchi, L.; Loubat, A.; Rochet, N.; Rossi, B.; Ponzio, G. Evidence for a Direct Correlation between c-Jun NH2 Terminal Kinase 1 Activation, Cyclin D2 Expression, and G1/S Phase Transition in the Murine Hybridoma 7TD1 Cells. Exp. Cell Res. 2000, 261, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Putney, L.K.; Barber, D.L. Na-H Exchange-dependent Increase in Intracellular pH Times G2/M Entry and Transition. J. Biol. Chem. 2003, 278, 44645–44649. [Google Scholar] [CrossRef] [PubMed]
- Uzman, J.; Patil, S.; Uzgare, A.R.; Sater, A.K. The Role of Intracellular Alkalinization in the Establishment of Anterior Neural Fate inXenopus. Dev. Biol. 1998, 193, 10–20. [Google Scholar] [CrossRef][Green Version]
- Boussouf, A.; Gaillard, S. Intracellular pH changes during oligodendrocyte differentiation in primary culture. J. Neurosci. Res. 2000, 59, 731–739. [Google Scholar] [CrossRef]
- Sharma, M.; Astekar, M.; Soi, S.; Manjunatha, B.; Shetty, D.; Radhakrishnan, R. pH Gradient Reversal: An Emerging Hallmark of Cancers. Recent Patents-Anti-Cancer Drug Discov. 2015, 10, 244–258. [Google Scholar] [CrossRef]
- Brophy, G.T.; Sladek, N. Influence of pH on the cytotoxic activity of chlorambucil. Biochem. Pharmacol. 1983, 32, 79–84. [Google Scholar] [CrossRef]
- Skarsgard, L.D.; Skwarchuk, M.W.; Vinczan, A.; Kristl, J.; Chaplin, D.J. The cytotoxicity of melphalan and its relationship to pH, hypoxia and drug uptake. Anticancer Res. 1995, 15, 219–223. [Google Scholar]
- Born, R.; Eichholtz-Wirth, H. Effect of different physiological conditions on the action of adriamycin on Chinese hamster cells in vitro. Br. J. Cancer 1981, 44, 241–246. [Google Scholar] [CrossRef]
- Hindenburg, A.A.; Gervasoni, J.E.; Krishna, S.; Stewart, V.J.; Rosado, M.; Lutzky, J.; Bhalla, K.; Baker, M.A.; Taub, R.N. Intracellular distribution and pharmacokinetics of daunorubicin in anthracycline-sensitive and -resistant HL-60 cells. Cancer Res. 1989, 49, 4607–4614. [Google Scholar] [PubMed]
- Jähde, E.; Glüsenkamp, K.H.; Rajewsky, M.F. Protection of cultured malignant cells from mitoxantrone cytotoxicity by low extracellular pH: A possible mechanism for chemoresistance in vivo. Eur. J. Cancer Clin. Oncol. 1990, 26, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Vukovic, V.; Tannock, I. Influence of low pH on cytotoxicity of paclitaxel, mitoxantrone and topotecan. Br. J. Cancer 1997, 75, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- Stéphanou, A.; Ballesta, A. pH as a potential therapeutic target to improve temozolomide antitumor efficacy: A mechanistic modeling study. Pharmacol. Res. Perspect. 2019, 7, e00454. [Google Scholar] [CrossRef] [PubMed]
- Andersen, A.P.; Moreira, J.M.A.; Pedersen, S.F. Interactions of ion transporters and channels with cancer cell metabolism and the tumour microenvironment. Philos. Trans. R. Soc. B 2014, 369, 20130098. [Google Scholar] [CrossRef]
- Parks, S.K.; Chiche, J.; Pouysségur, J. Disrupting proton dynamics and energy metabolism for cancer therapy. Nat. Rev. Cancer 2013, 13, 611–623. [Google Scholar] [CrossRef]
- Neri, D.; Supuran, C.T. Interfering with pH regulation in tumours as a therapeutic strategy. Nat. Rev. Drug Discov. 2011, 10, 767–777. [Google Scholar] [CrossRef]
- Lee, E.S.; Gao, Z.; Bae, Y.H. Recent progress in tumor pH targeting nanotechnology. J. Control. Release Off. J. Control. Release Soc. 2008, 132, 164–170. [Google Scholar] [CrossRef]
- Song, S.; Chen, F.; Qi, H.; Li, F.; Xin, T.; Xu, J.; Ye, T.; Sheng, N.; Yang, X.; Pan, W. Multifunctional Tumor-Targeting Nanocarriers Based on Hyaluronic Acid-Mediated and pH-Sensitive Properties for Efficient Delivery of Docetaxel. Pharm. Res. 2014, 31, 1032–1045. [Google Scholar] [CrossRef]
- Tafech, A.; Beaujean, C.; Usson, Y.; Stéphanou, A. Generalization of the ratiometric method to extend pH range measurements of the BCECF probe. Biomolecules 2023, 13, 442. [Google Scholar] [CrossRef]
- Oraiopoulou, M.E.; Tzamali, E.; Tzedakis, G.; Vakis, A.; Papamatheakis, J.; Sakkalis, V. In Vitro/In Silico Study on the Role of Doubling Time Heterogeneity among Primary Glioblastoma Cell Lines. BioMed Res. Int. 2017, 2017, 8569328. [Google Scholar] [CrossRef] [PubMed]
- Mathews, M.S.; Blickenstaff, J.W.; Shih, E.C.; Zamora, G.; Vo, V.; Sun, C.H.; Hirschberg, H.; Madsen, S.J. Photochemical internalization of bleomycin for glioma treatment. J. Biomed. Opt. 2012, 17, 058001. [Google Scholar] [CrossRef] [PubMed]
- Tsien, R.Y. A non-disruptive technique for loading calcium buffers and indicators into cells. Nature 1981, 290, 527–528. [Google Scholar] [CrossRef] [PubMed]
- Graber, M.L.; DiLillo, D.C.; Friedman, B.L.; Pastoriza-Munoz, E. Characteristics of fluoroprobes for measuring intracellular pH. Anal. Biochem. 1986, 156, 202–212. [Google Scholar] [CrossRef]
- Damaghi, M.; Wojtkowiak, J.W.; Gillies, R.J. pH sensing and regulation in cancer. Front. Physiol. 2013, 4, 370. [Google Scholar] [CrossRef]
- Izutsu, K.T. Intracellular pH, H ion flux and H ion permeability coefficient in bullfrog toe muscle. J. Physiol. 1972, 221, 15–27. [Google Scholar] [CrossRef]
- Baltz, J.; Biggers, J.; Lechene, C. A novel H+ permeability dominating intracellular pH in the early mouse embryo. Development 1993, 118, 1353–1361. [Google Scholar] [CrossRef]
- Doyen, D.; Poët, M.; Jarretou, G.; Pisani, D.F.; Tauc, M.; Cougnon, M.; Argentina, M.; Bouret, Y.; Counillon, L. Intracellular pH Control by Membrane Transport in Mammalian Cells. Insights Into the Selective Advantages of Functional Redundancy. Front. Mol. Biosci. 2022, 9, 825028. [Google Scholar] [CrossRef]
- Carmelo, V.; Santos, H.; Sá-Correia, I. Effect of extracellular acidification on the activity of plasma membrane ATPase and on the cytosolic and vacuolar pH of Saccharomyces cerevisiae. Biochim. Biophys. Acta (BBA)-Biomembr. 1997, 1325, 63–70. [Google Scholar] [CrossRef]
- Glunde, K.; Dussmann, H.; Juretschke, H.P.; Leibfritz, D. Na+/H+ exchange subtype 1 inhibition during extracellular acidification and hypoxia in glioma cells. J. Neurochem. 2002, 80, 36–44. [Google Scholar] [CrossRef]
- Lacoste, I.; Harvey, B.; Ehrenfeld, J. Cl- permeability of the basolateral membrane of the Rana esculenta epithelium: Activation of Cl-/HCO3- exchange by alkaline intracellular pH. Biochim. Biophys. Acta (BBA)-Biomembr. 1991, 1063, 103–110. [Google Scholar] [CrossRef]
- Vilariño, N.; Vieytes, M.R.; Vieites, J.M.; Botana, L.M. Role of HCO-3Ions in Cytosolic pH Regulation in Rat Mast Cells: Evidence for a New Na+-Independent, HCO-3-Dependent Alkalinizing Mechanism. Biochem. Biophys. Res. Commun. 1998, 253, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Rivarola, V.; Ford, P.; Chara, O.; Parisi, M.; Capurro, C. Functional and Molecular Adaptation of Cl−/HCO3− Exchanger to Chronic Alkaline Media in RenalCells. Cell. Physiol. Biochem. 2005, 16, 271–280. [Google Scholar] [CrossRef]
- Swietach, P.; Vaughan-Jones, R.D.; Harris, A.L.; Hulikova, A. The chemistry, physiology and pathology of pH in cancer. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130099. [Google Scholar] [CrossRef] [PubMed]
- Estrella, V.; Chen, T.; Lloyd, M.; Wojtkowiak, J.; Cornnell, H.H.; Ibrahim-Hashim, A.; Bailey, K.; Balagurunathan, Y.; Rothberg, J.M.; Sloane, B.F.; et al. Acidity Generated by the Tumor Microenvironment Drives Local Invasion. Cancer Res. 2013, 73, 1524–1535. [Google Scholar] [CrossRef] [PubMed]
- Corbet, C.; Feron, O. Tumour acidosis: From the passenger to the driver’s seat. Nat. Rev. Cancer 2017, 17, 577–593. [Google Scholar] [CrossRef]
- Hackam, D.J.; Grinstein, S.; Rotstein, O.D. Intracellular pH regulation in leukocytes. Shock 1996, 5, 17–21. [Google Scholar] [CrossRef]
- Rao, M.; Streur, T.L.; Aldwell, F.E.; Cook, G.M. Intracellular pH regulation by Mycobacterium smegmatis and Mycobacterium bovis BCG. Microbiology 2001, 147, 1017–1024. [Google Scholar] [CrossRef]
- Zeng, W.P.; Yang, S.; Zhou, B. Intracellular pH-regulated Cell Intrinsic Control of Death and Proliferation of Lymphocytes in Immune Response and Tumor Cells. BioRxiv 2021. [Google Scholar] [CrossRef]
- Man, C.H.; Mercier, F.E.; Liu, N.; Dong, W.; Stephanopoulos, G.; Jiang, L.; Jung, Y.; Lin, C.P.; Leung, A.Y.H.; Scadden, D.T. Proton export alkalinizes intracellular pH and reprograms carbon metabolism to drive normal and malignant cell growth. Blood 2022, 139, 502–522. [Google Scholar] [CrossRef]
- Hirschhaeuser, F.; Menne, H.; Dittfeld, C.; West, J.; Mueller-Klieser, W.; Kunz-Schughart, L.A. Multicellular tumor spheroids: An underestimated tool is catching up again. J. Biotechnol. 2010, 148, 3–15. [Google Scholar] [CrossRef]
- Freyer, J.P.; Sutherland, R.M. A reduction in the in situ rates of oxygen and glucose consumption of cells in EMT6/Ro spheroids during growth. J. Cell. Physiol. 1985, 124, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, R.M. Cell and Environment Interactions in Tumor Microregions: The Multicell Spheroid Model. Science 1988, 240, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, J.; Seidel, C.; Ebner, R.; Kunz-Schughart, L.A. Spheroid-based drug screen: Considerations and practical approach. Nat. Protoc. 2009, 4, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.C.; Hung, B.P.; Browne-Bourne, S.; Zhou, D.; Yeung, J.; Genetos, D.C.; Leach, J.K. Measurement of oxygen tension within mesenchymal stem cell spheroids. J. R. Soc. Interface 2017, 14, 20160851. [Google Scholar] [CrossRef] [PubMed]
- Galkina, S.; Sud’ina, G.; Margolis, L. Regulation of intracellular pH by phospholipase A2 and protein kinase C upon neutrophil adhesion to solid substrata. FEBS Lett. 1996, 393, 117–120. [Google Scholar] [CrossRef]
- Galkina, S.; Sud’ina, G.; Dergacheva, G.; Margolis, L. Regulation of intracellular pH by cell–cell adhesive interactions. FEBS Lett. 1995, 374, 17–20. [Google Scholar] [CrossRef]
- Zagaynova, E.V.; Druzhkova, I.N.; Mishina, N.M.; Ignatova, N.I.; Dudenkova, V.V.; Shirmanova, M.V. Imaging of Intracellular pH in Tumor Spheroids Using Genetically Encoded Sensor SypHer2. In Advances in Experimental Medicine and Biology; Springer International Publishing: New York, NY, USA, 2017; pp. 105–119. [Google Scholar] [CrossRef]
- Ballesta, A.; Zhou, Q.; Zhang, X.; Lv, H.; Gallo, J. Multiscale Design of Cell-Type-Specific Pharmacokinetic/Pharmacodynamic Models for Personalized Medicine: Application to Temozolomide in Brain Tumors. CPT Pharmacomet. Syst. Pharmacol. 2014, 3, 112. [Google Scholar] [CrossRef]
- Gnochi, D.; Sabbà, C.; Mazzocca, A. Lactic acid fermentation: A maladaptative mechanism and an evolutionary throwback boosting cancer drug resistance. Biochimie 2023, 208, 180–185. [Google Scholar] [CrossRef]
- Gnochi, D.; Nikolic, D.; Paparella, R.R.; Sabbà, C.; Mazzocca, A. Cellular adaptation take advantage of atavistic regression programs during carcinogenesis. Cancers 2023, 15, 3942. [Google Scholar] [CrossRef]
- Jacquet, P.; Stéphanou, A. Metabolic Reprogramming, Questioning, and Implications for Cancer. Biology 2021, 10, 129. [Google Scholar] [CrossRef] [PubMed]
- Jacquet, P.; Stéphanou, A. Searching for the metabolic signature of cancer: A review from Warburg’s time to now. Biomolecules 2022, 12, 1412. [Google Scholar] [CrossRef] [PubMed]
- Jacquet, P.; Stéphanou, A. A reduced model of cell metabolism to revisit the glycolysis-OXPHOS relationship in the deregulated tumor microenvironment. J. Theor. Biol. 2023, 562, 111434. [Google Scholar] [CrossRef] [PubMed]
- Tannock, I.F.; Rotin, D. Acid pH in tumors and its potential for therapeutic exploitation. Cancer Res. 1989, 49 16, 4373–4384. [Google Scholar]
- Wahl, M.L.; Pooler, P.M.; Briand, P.; Leeper, D.B.; Owen, C.S. Intracellular pH regulation in a nonmalignant and a derived malignant human breast cell line. J. Cell. Physiol. 2000, 183, 373–380. [Google Scholar] [CrossRef]
- Piao, S.; Amaravadi, R.K. Targeting the lysosome in cancer. Ann. N. Y. Acad. Sci. 2015, 1371, 45–54. [Google Scholar] [CrossRef]
- Tang, T.; Yang, Z.; Wang, D.; Yang, X.; Wang, J.; Li, L.; Wen, Q.; Gao, L.; Bian, X.; Yu, S. The role of lysosomes in cancer development and progression. Cell Biosci. 2020, 10, 131. [Google Scholar] [CrossRef]
- Chen, R.; Jäättelä, M.; Liu, B. Lysosome as a Central Hub for Rewiring PH Homeostasis in Tumors. Cancers 2020, 12, 2437. [Google Scholar] [CrossRef]
- Banerjee, S.; Kane, P.M. Regulation of V-ATPase Activity and Organelle pH by Phosphatidylinositol Phosphate Lipids. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef]
- Kinsella, J.L.; Aronson, P.S. Properties of the Na+-H+ exchanger in renal microvillus membrane vesicles. Am. J.-Physiol.-Ren. Physiol. 1980, 238, F461–F469. [Google Scholar] [CrossRef]
- Cassel, D.; Katz, M.; Rotman, M. Depletion of cellular ATP inhibits Na+/H+ antiport in cultured human cells. Modulation of the regulatory effect of intracellular protons on the antiporter activity. J. Biol. Chem. 1986, 261, 5460–5466. [Google Scholar] [CrossRef]
- Brown, S.E.; Heming, T.A.; Benedict, C.R.; Bidani, A. ATP-sensitive Na(+)-H+ antiport in type II alveolar epithelial cells. Am. J.-Physiol.-Cell Physiol. 1991, 261, C954–C963. [Google Scholar] [CrossRef]
- Levine, S.A.; Montrose, M.H.; Tse, C.M.; Donowitz, M. Kinetics and regulation of three cloned mammalian Na+/H+ exchangers stably expressed in a fibroblast cell line. J. Biol. Chem. 1993, 268, 25527–25535. [Google Scholar] [CrossRef]
- Brown, S.V.; Hosking, P.; Li, J.; Williams, N. ATP Synthase Is Responsible for Maintaining Mitochondrial Membrane Potential in Bloodstream Form Trypanosoma brucei. Eukaryot. Cell 2006, 5, 45–53. [Google Scholar] [CrossRef]
- Vasan, K.; Clutter, M.; Dunne, S.F.; George, M.D.; Luan, C.H.; Chandel, N.S.; Martínez-Reyes, I. Genes Involved in Maintaining Mitochondrial Membrane Potential Upon Electron Transport Chain Disruption. Front. Cell Dev. Biol. 2022, 10, 781558. [Google Scholar] [CrossRef] [PubMed]
- Langer, G.A. The effect of pH on cellular and membrane calcium binding and contraction of myocardium. A possible role for sarcolemmal phospholipid in EC coupling. Circ. Res. 1985, 57, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Wanandi, S.; Yustisia, I.; Neolaka, G.; Jusman, S. Impact of extracellular alkalinization on the survival of human CD24−/CD44+ breast cancer stem cells associated with cellular metabolic shifts. Braz. J. Med. Biol. Res. 2017, 50. [Google Scholar] [CrossRef] [PubMed]
- Khajah, M.A.; Almohri, I.; Mathew, P.M.; Luqmani, Y.A. Extracellular Alkaline pH Leads to Increased Metastatic Potential of Estrogen Receptor Silenced Endocrine Resistant Breast Cancer Cells. PLoS ONE 2013, 8, e76327. [Google Scholar] [CrossRef] [PubMed]
- Avnet, S.; Lemma, S.; Cortini, M.; Pellegrini, P.; Perut, F.; Zini, N.; Kusuzaki, K.; Chano, T.; Grisendi, G.; Dominici, M.; et al. Altered pH gradient at the plasma membrane of osteosarcoma cells is a key mechanism of drug resistance. Oncotarget 2016, 7, 63408–63423. [Google Scholar] [CrossRef] [PubMed]
- Güvenalp, N.; Güvenç, D. An evaluation of the effects of medium pH on the viability of the HepG2 cell line. J. Etlik Vet. Microbiol. 2020, 31, 107–114. [Google Scholar] [CrossRef]
- Ichas, F.; Mazat, J.P. From calcium signaling to cell death: Two conformations for the mitochondrial permeability transition pore. Switching from low- to high-conductance state. Biochim. Biophys. Acta (BBA)-Bioenerg. 1998, 1366, 33–50. [Google Scholar] [CrossRef]
- Mazzio, E.A.; Boukli, N.; Rivera, N.; Soliman, K.F.A. Pericellular pH homeostasis is a primary function of the Warburg effect: Inversion of metabolic systems to control lactate steady state in tumor cells. Cancer Sci. 2012, 103, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, J.; Hassan, A.; Lee, C.H.; Xie, X.S.; Li, X. Molecular basis of V-ATPase inhibition by bafilomycin A1. Nat. Commun. 2021, 12, 1782. [Google Scholar] [CrossRef] [PubMed]
- Masereel, B. An overview of inhibitors of Na+/H+ exchanger. Eur. J. Med. Chem. 2003, 38, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Tidwell, T.R.; Røsland, G.V.; Tronstad, K.J.; Søreide, K.; Hagland, H.R. Metabolic flux analysis of 3D spheroids reveals significant differences in glucose metabolism from matched 2D cultures of colorectal cancer and pancreatic ductal adenocarcinoma cell lines. Cancer Metab. 2022, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Denny, B.J.; Wheelhouse, R.T.; Stevens, M.F.G.; Tsang, L.L.H.; Slack, J.A. NMR and Molecular Modeling Investigation of the Mechanism of Activation of the Antitumor Drug Temozolomide and Its Interaction with DNA. Biochemistry 1994, 33, 9045–9051. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial pH | pH (t = 8 h) | pH (t = 48 h) |
|---|---|---|
| 5.00 | 5.60 | 6.07 |
| 6.00 | 6.60 | 6.91 |
| 6.40 | 7.12 | 7.33 |
| 6.80 | 7.53 | 7.67 |
| 7.20 | 7.88 | 8.15 |
| 7.40 | 8.20 | 8.52 |
| F98 Monolayer (2D) | Extracellular pH | % Cell viability with | % Cell viability after | Cell viability |
| (pH) | DMSO (control) | TMZ treatment | gap | |
| 6.91 | 39.28 ± 2.38 | 23.80 ± 3.31 | 15 | |
| 7.33 | 61.90 ± 3.14 | 39.28 ± 2.09 | 22 | |
| 7.67 | 75.79 ± 4.81 | 28.17 ± 0.68 | 47 | |
| 8.15 | 105.15 ± 2.47 | 57.53 ± 3.43 | 48 | |
| 8.52 | 100 ± 2.99 | 37 ± 2.47 | 62 | |
| U87 Monolayer (2D) | Extracellular pH | % Cell viability with | % Cell viability after | Cell viability |
| (pH) | DMSO (control) | TMZ treatment | gap | |
| 6.91 | 47.30 ± 2.74 | 32.93 ± 2.74 | 14 | |
| 7.33 | 48.30 ± 3.11 | 23.95 ± 3.11 | 23 | |
| 7.67 | 72.45 ± 3.73 | 50.89 ± 3.73 | 21 | |
| 8.15 | 83.83 ± 2.07 | 41.91 ± 2.07 | 41 | |
| 8.52 | 100 ± 1.03 | 35.92 ± 1.03 | 64 | |
| F98 Spheroid (3D) | Extracellular pH | % Cell viability with | % Cell viability after | Cell viability |
| (pH) | DMSO (control) | TMZ treatment | gap | |
| 6.07 | 84.44 ± 7.21 | 68.27 ± 3.58 | 16 | |
| 6.91 | 123.13 ± 7.89 | 62.27 ± 4.72 | 60 | |
| 8.52 | 100 ± 8.64 | 29.83 ± 6.81 | 70 | |
| U87 Spheroid (3D) | Extracellular pH | % Cell viability with | % Cell viability after | Cell viability |
| (pH) | DMSO (control) | TMZ treatment | gap | |
| 6.07 | 80.49 ± 6.89 | 67.99 ± 5.36 | 12 | |
| 6.91 | 121.73 ± 6.21 | 78.11 ± 5.29 | 43 | |
| 8.52 | 100 ± 3.31 | 35.62 ± 4.84 | 64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tafech, A.; Jacquet, P.; Beaujean, C.; Fertin, A.; Usson, Y.; Stéphanou, A. Characterization of the Intracellular Acidity Regulation of Brain Tumor Cells and Consequences for Therapeutic Optimization of Temozolomide. Biology 2023, 12, 1221. https://doi.org/10.3390/biology12091221
Tafech A, Jacquet P, Beaujean C, Fertin A, Usson Y, Stéphanou A. Characterization of the Intracellular Acidity Regulation of Brain Tumor Cells and Consequences for Therapeutic Optimization of Temozolomide. Biology. 2023; 12(9):1221. https://doi.org/10.3390/biology12091221
Chicago/Turabian StyleTafech, Alaa, Pierre Jacquet, Céline Beaujean, Arnold Fertin, Yves Usson, and Angélique Stéphanou. 2023. "Characterization of the Intracellular Acidity Regulation of Brain Tumor Cells and Consequences for Therapeutic Optimization of Temozolomide" Biology 12, no. 9: 1221. https://doi.org/10.3390/biology12091221
APA StyleTafech, A., Jacquet, P., Beaujean, C., Fertin, A., Usson, Y., & Stéphanou, A. (2023). Characterization of the Intracellular Acidity Regulation of Brain Tumor Cells and Consequences for Therapeutic Optimization of Temozolomide. Biology, 12(9), 1221. https://doi.org/10.3390/biology12091221