

Transcriptomic Analysis of Light-Induced Genes in Nasonia vitripennis: Possible Implications for Circadian Light Entrainment Pathways


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Lines and Maintenance
2.2. Entrainment and Sample Collection
2.3. RNA Isolation and RNA Sequencing
2.4. Differential Expression and Functional Annotation Analysis of RNA-Seq
3. Results
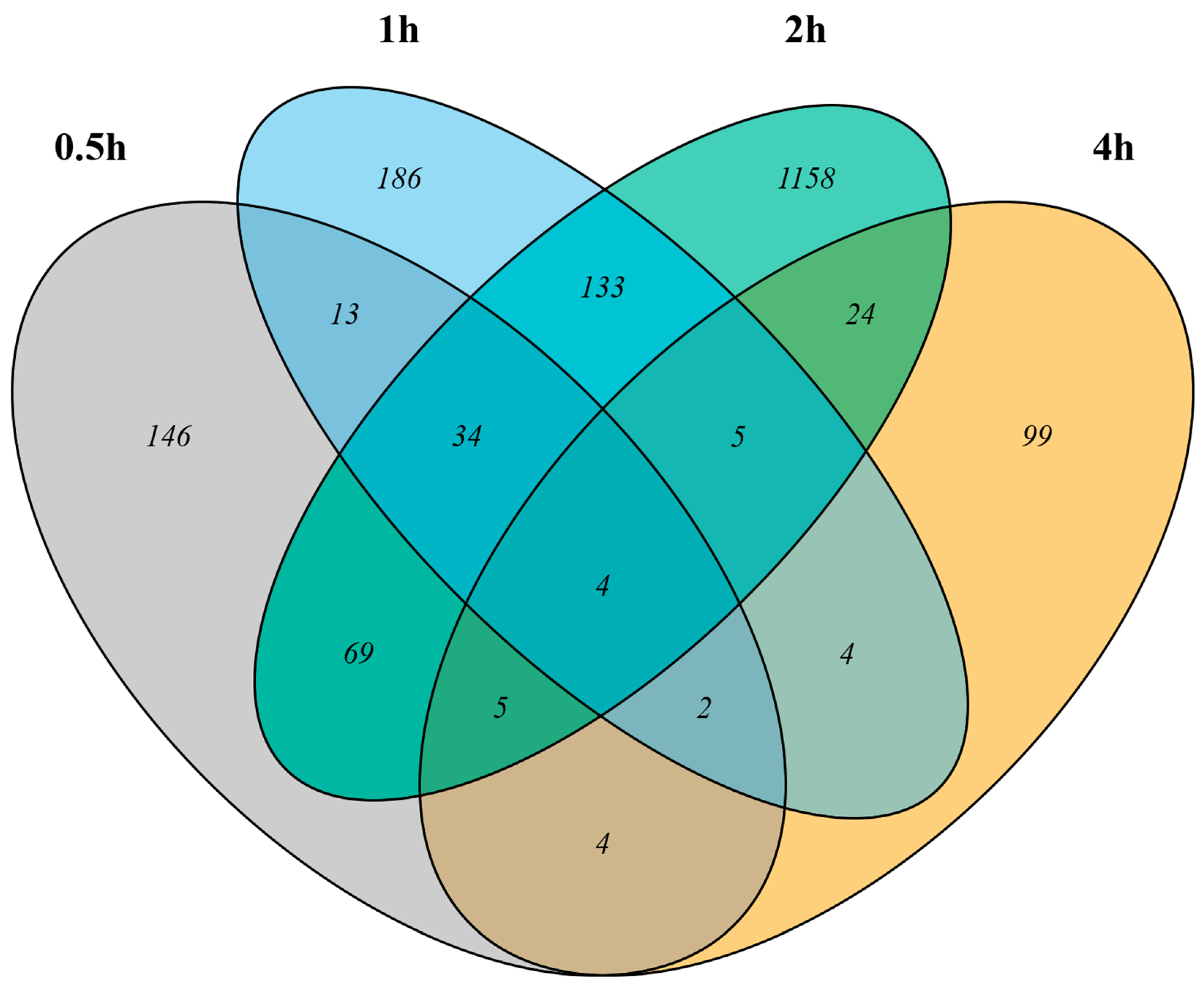
3.1. Differentially Expressed Genes Induced by Light
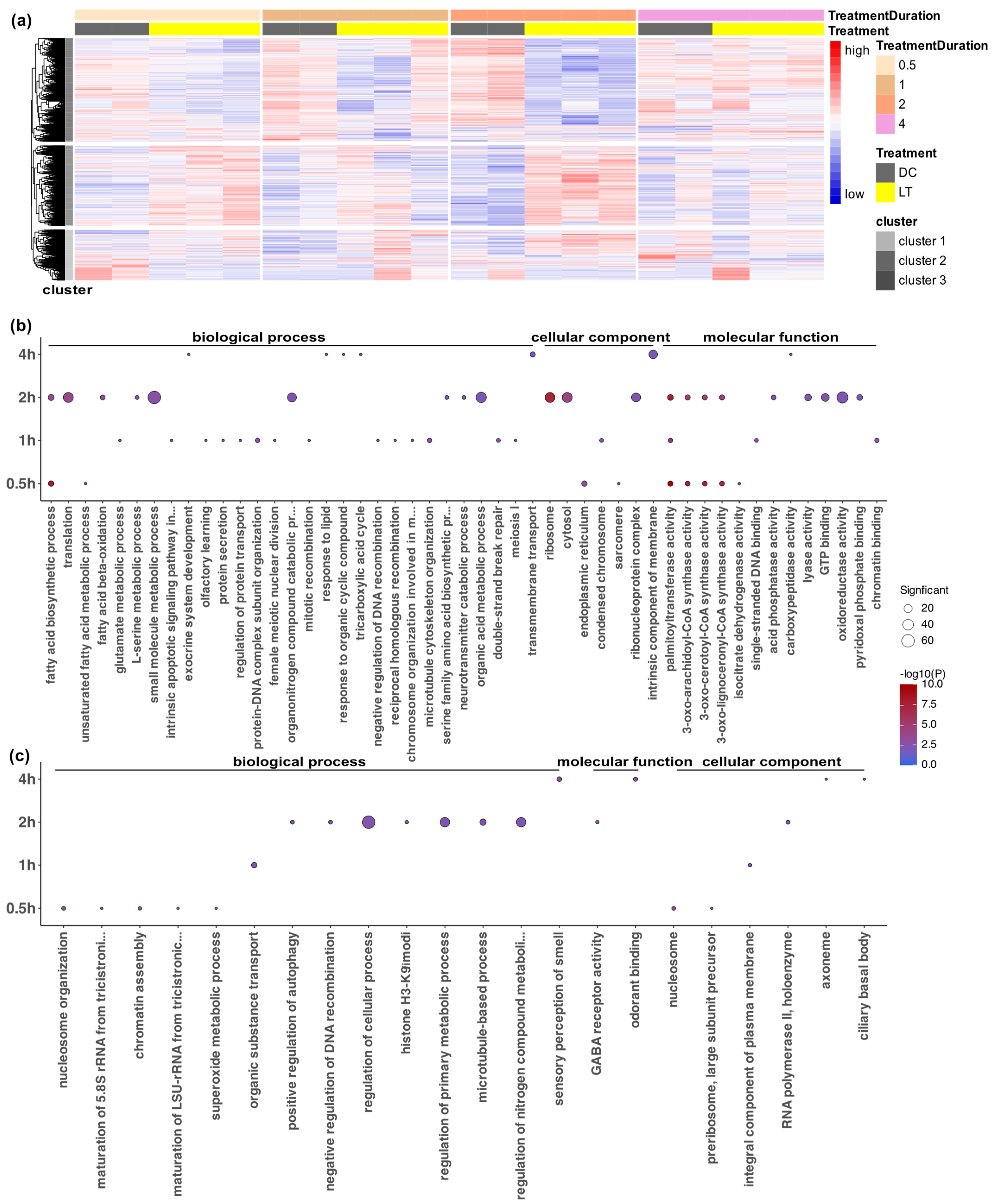
3.2. Functional Annotation Analysis of Differentially Expressed Genes
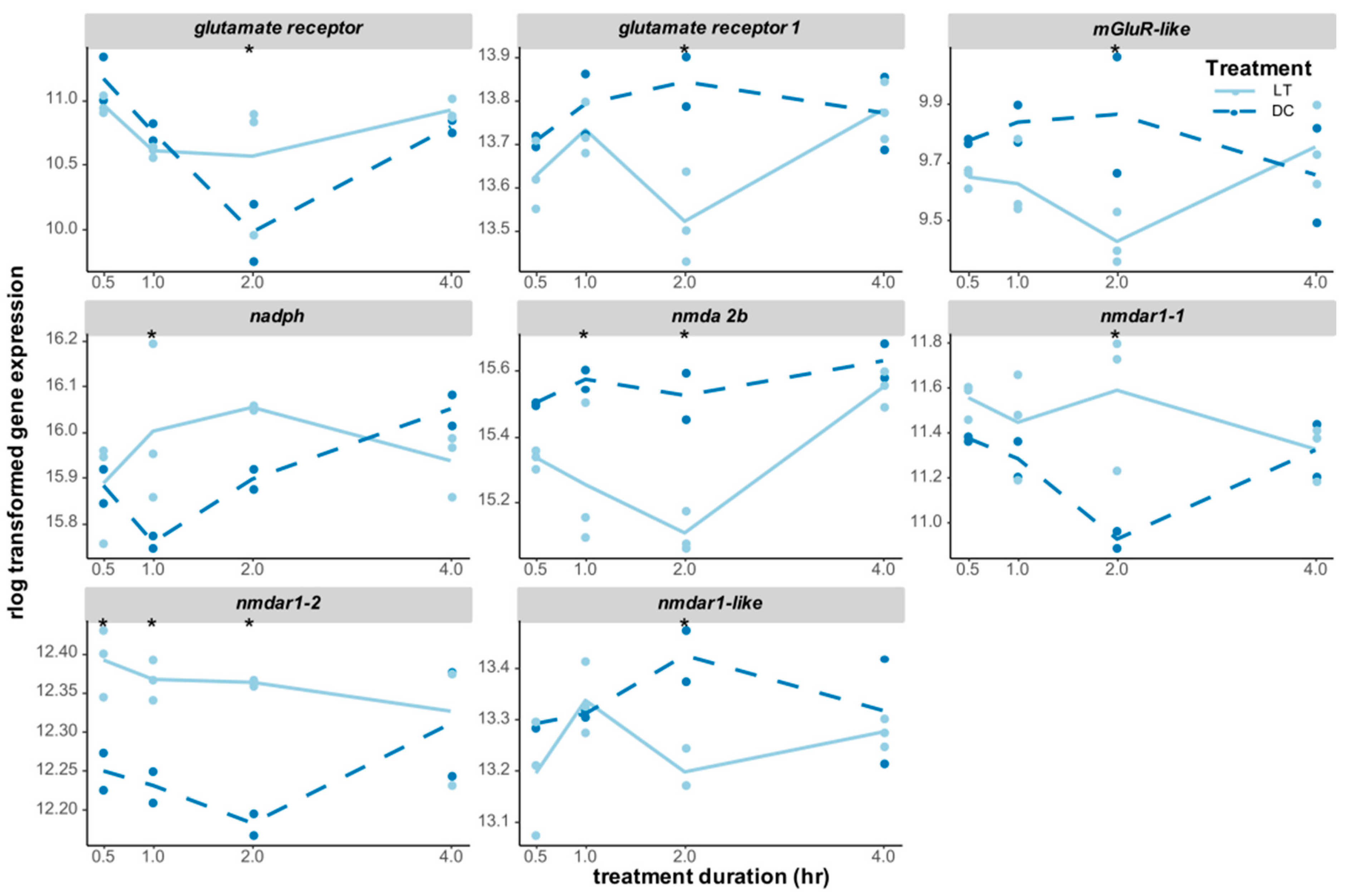
3.3. Candidate Genes for the Circadian Light Input Pathway
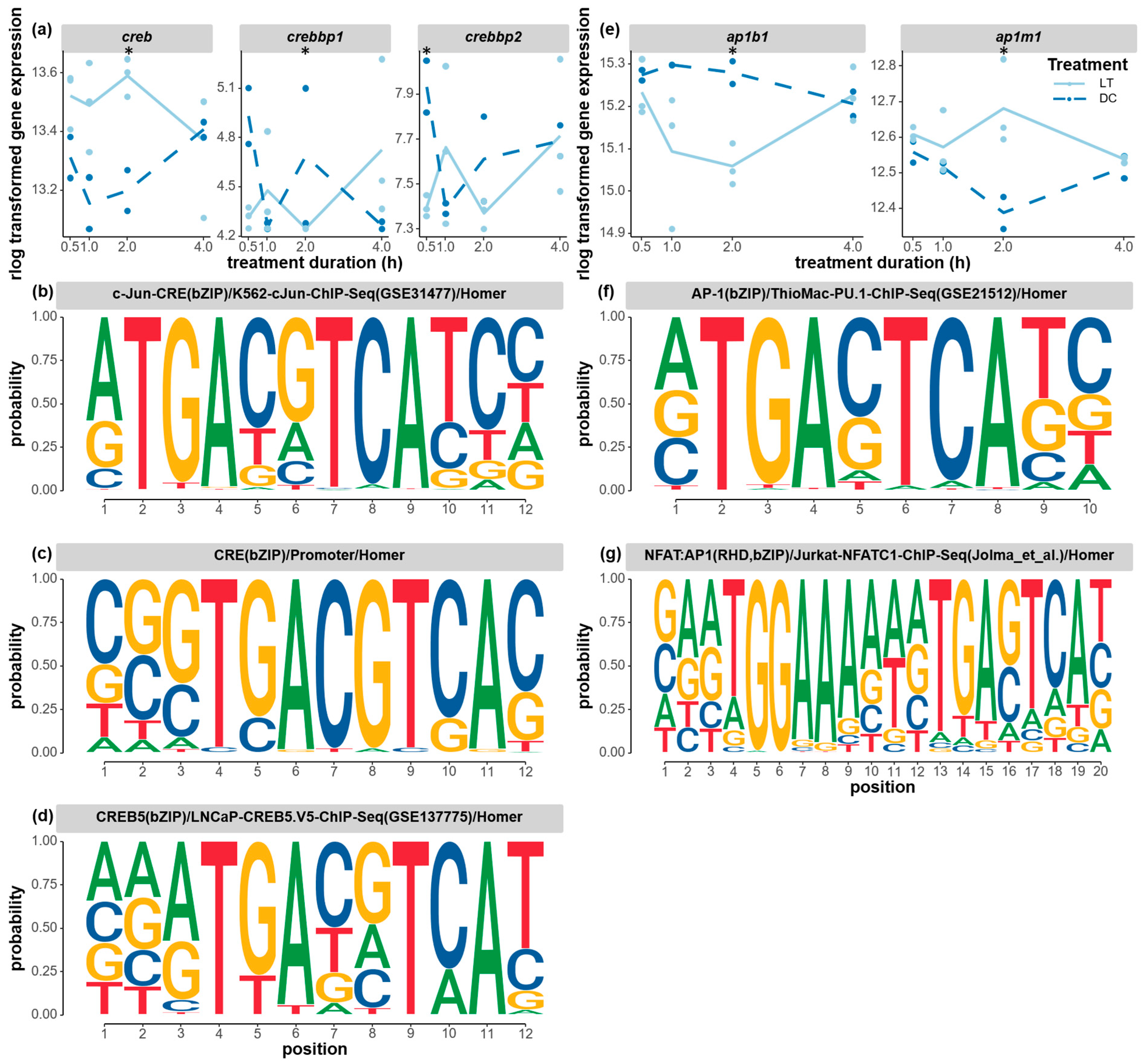
3.4. Candidate Transcription Factors Involved in Circadian Light Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allada, R.; Chung, B.Y. Circadian organization of behavior and physiology in Drosophila. Annu. Rev. Physiol. 2010, 72, 605–624. [Google Scholar] [CrossRef]
- Beer, K.; Helfrich-Förster, C. Model and non-model insects in chronobiology. Front. Behav. Neurosci. 2020, 14, 601676. [Google Scholar] [CrossRef]
- Duffy, J.F.; Czeisler, C.A. Effect of light on human circadian physiology. Sleep Med. Clin. 2009, 2, 165–177. [Google Scholar] [CrossRef]
- Sletten, T.L.; Cappuccio, F.P.; Davidson, A.J.; van Cauter, E.; Rajaratnam, S.M.W.; Scheer, F.A.J.L. Health consequences of circadian disruption. Sleep 2020, 43, zsz194. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.A.; Davidson, A.J. Health consequences of circadian disruption in humans and animal models. Prog. Mol. Biol. Transl. Sci. 2013, 119, 283–323. [Google Scholar] [PubMed]
- Blume, C.; Garbazza, C.; Spitschan, M. Effects of light on human circadian rhythms, sleep and mood. Somnologie 2019, 3, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Desouhant, E.; Gomes, E.; Mondy, N.; Amat, I. Mechanistic, ecological, and evolutionary consequences of artificial light at night for insects: Review and prospective. Entomol. Exp. Appl. 2019, 1, 37–58. [Google Scholar] [CrossRef]
- Oklejewicz, M.; Hut, R.A.; Daan, S.; Loudon, A.; Stirland, A.J. Metabolic rate changes proportionally to circadian frequency in tau mutant Syrian hamsters. J. Biol. Rhythm. 1997, 5, 413–422. [Google Scholar] [CrossRef]
- Spoelstra, K.; Wikelski, M.; Daan, S.; Loudon, A.S.I.; Hau, M. Natural selection against a circadian clock gene mutation in mice. Proc. Natl. Acad. Sci. USA 2016, 3, 686–691. [Google Scholar] [CrossRef]
- Ozturk, N.; Selby, C.P.; Annayev, Y.; Zhong, D.; Sancar, A. Reaction mechanism of Drosophila cryptochrome. Proc. Natl. Acad. Sci. USA 2011, 2, 516–521. [Google Scholar] [CrossRef]
- Provencio, I.; Jiang, G.S.; De Grip, W.J.; Hayes, W.P.; Rollag, M.D. Melanopsin: An opsin in melanophores, brain, and eye. Proc. Natl. Acad. Sci. USA 1998, 1, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by retinal ganglion cells that set the circadian clock. Science 2002, 5557, 1070–1073. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Sato, T.K.; Castrucci, A.M.; Rollag, M.D.; DeGrip, W.J.; Hogenesch, J.B.; Provencio, I.; Kay, S.A. Melanopsin (Opn4) requirement for normal light-induced circadian phase shifting. Science 2002, 5601, 2213–2216. [Google Scholar] [CrossRef] [PubMed]
- Hattar, S.; Liao, H.W.; Takao, M.; Berson, D.M.; Yau, K.W. Melanopsin-containing retinal ganglion cells: Architecture, projections, and intrinsic photosensitivity. Science 2002, 5557, 1065–1070. [Google Scholar] [CrossRef]
- Walmsley, L.; Hanna, L.; Mouland, J.; Martial, F.; West, A.; Smedley, A.R.; Bechtold, D.A.; Webb, A.R.; Lucas, R.J.; Brown, T.M. Colour as a signal for entraining the mammalian circadian clock. PLoS Biol. 2015, 4, e1002127. [Google Scholar] [CrossRef]
- Gau, D.; Lemberger, T.; von Gall, C.; Kretz, O.; Minh, N.L.; Gass, P.; Schmid, W.; Schibler, U.; Korf, H.W.; Schutz, G. Phosphorylation of CREB Ser142 regulates light-induced phase shifts of the circadian clock. Neuron 2002, 2, 245–252. [Google Scholar] [CrossRef]
- Lee, B.; Li, A.; Hansen, K.F.; Cao, R.; Yoon, J.H.; Obrietan, K. CREB Influences Timing and Entrainment of the SCN Circadian Clock. J. Biol. Rhythm. 2010, 6, 410–420. [Google Scholar] [CrossRef]
- Jagannath, A.; Butler, R.; Godinho, S.I.H.; Couch, Y.; Brown, L.A.; Vasudevan, S.R.; Flanagan, K.C.; Anthony, D.; Churchill, G.C.; Wood, M.J.A.; et al. The CRTC1-SIK1 pathway regulates entrainment of the circadian clock. Cell 2013, 5, 1100–1111. [Google Scholar] [CrossRef]
- Steindal, I.A.F.; Whitmore, D. Zebrafish circadian clock entrainment and the importance of broad spectral light sensitivity. Front. Physiol. 2020, 11, 1002. [Google Scholar] [CrossRef]
- Vatine, G.; Vallone, D.; Gothilf, Y.; Foulkes, N.S. It’s time to swim! Zebrafish and the circadian clock. FEBS Lett. 2011, 10, 1485–1494. [Google Scholar] [CrossRef]
- Pagano, C.; Siauciunaite, R.; Idda, M.L.; Ruggiero, G.; Ceinos, R.M.; Pagano, M.; Frigato, E.; Bertolucci, C.; Foulkes, N.S.; Vallone, D. Evolution shapes the responsiveness of the D-box enhancer element to light and reactive oxygen species in vertebrates. Sci. Rep. 2018, 1, 13180. [Google Scholar] [CrossRef] [PubMed]
- Ziv, L.; Levkovitz, S.; Toyama, R.; Falcon, J.; Gothilf, Y. Functional development of the zebrafish pineal gland: Light-induced expression of period2 is required for onset of the circadian clock. J. Neuroendocrinol. 2005, 5, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Tamai, T.K.; Young, L.C.; Whitmore, D. Light signaling to the zebrafish circadian clock by Cryptochrome 1a. Proc. Natl. Acad. Sci. USA 2007, 37, 14712–14717. [Google Scholar] [CrossRef] [PubMed]
- Ashton, A.; Foster, R.G.; Jagannath, A. Photic entrainment of the circadian system. Int. J. Mol. Sci. 2022, 2, 729. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Metterville, D.; Briscoe, A.D.; Reppert, S.M. Insect cryptochromes: Gene duplication and loss define diverse ways to construct insect circadian clocks. Mol. Biol. Evol. 2007, 4, 948–955. [Google Scholar] [CrossRef]
- Kotwica-Rolinska, J.; Chodáková, L.; Smýkal, V.; Damulewicz, M.; Provazník, J.; Wu, B.C.; Hejníková, M.; Chvalová, D.; Doležel, D. Loss of timeless underlies an evolutionary transition within the circadian clock. Mol. Biol. Evol. 2021, 1, msab346. [Google Scholar] [CrossRef]
- Helfrich-Förster, C. Light input pathways to the circadian clock of insects with an emphasis on the fruit fly Drosophila melanogaster. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2020, 2, 259–272. [Google Scholar] [CrossRef]
- Lam, V.H.; Chiu, J.C. Evolution and design of invertebrate circadian clocks. In The Oxford Handbook of Invertebrate Neurobiology; Byrne, J.H., Ed.; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Tomioka, K.; Matsumoto, A. Circadian molecular clockworks in non-model insects. Curr. Opin. Insect Sci. 2015, 7, 58–64. [Google Scholar] [CrossRef]
- Rubin, E.B.; Shemesh, Y.; Cohen, M.; Elgavish, S.; Robertson, H.M.; Bloch, G. Molecular and phylogenetic analyses reveal mammalian-like clockwork in the honey bee (Apis mellifera) and shed new light on the molecular evolution of the circadian clock. Genome Res. 2006, 11, 1352–1365. [Google Scholar] [CrossRef]
- Davies, N.J.; Tauber, E. Deep sequencing analysis of the circadian transcriptome of the jewel wasp Nasonia vitripennis. bioRxiv 2016, preprint. [Google Scholar] [CrossRef]
- Werren, J.H.; Loehlin, D.W. The parasitoid wasp Nasonia: An emerging model system with haploid male genetics. Cold Spring Harb. Protoc. 2009, 10, pdb.emo134. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Richards, S.; Desjardins, C.A.; Niehuis, O.; Gadau, J.; Colbourne, J.K.; Nasonia Genome Working Group; Werren, J.H.; Richards, S.; Desjardins, C.A.; et al. Functional and evolutionary insights from the genomes of three parasitoid Nasonia species. Science 2010, 5963, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Bertossa, R.C.; van Dijk, J.; Beersma, D.G.; Beukeboom, L.W. Circadian rhythms of adult emergence and activity but not eclosion in males of the parasitic wasp Nasonia vitripennis. J. Insect Physiol. 2010, 7, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Bertossa, R.C.; van Dijk, J.; Diao, W.; Saunders, D.; Beukeboom, L.W.; Beersma, D.G.M. Circadian rhythms differ between sexes and closely related species of Nasonia wasps. PLoS ONE 2013, 3, e60167. [Google Scholar] [CrossRef] [PubMed]
- Floessner, T.S.E.; Boekelman, F.E.; Druiven, S.J.M.; de Jong, M.; Rigter, P.M.F.; Beersma, D.G.M.; Hut, R.A. Lifespan is unaffected by size and direction of daily phase shifts in Nasonia, a hymenopteran insect with strong circadian light resetting. J. Insect Physiol. 2019, 117, 103896. [Google Scholar] [CrossRef] [PubMed]
- Paolucci, S.; van de Zande, L.; Beukeboom, L.W. Adaptive latitudinal cline of photoperiodic diapause induction in the parasitoid Nasonia vitripennis in Europe. J. Evol. Biol. 2013, 4, 705–718. [Google Scholar] [CrossRef]
- Dalla Benetta, E.; Beukeboom, L.W.; van de Zande, L. Adaptive differences in circadian clock gene expression patterns and photoperiodic diapause induction in Nasonia vitripennis. Am. Nat. 2019, 6, 881–896. [Google Scholar] [CrossRef]
- Floessner, T. Latitudinal Differences in the Circadian System of Nasonia vitripennis. Ph.D. Thesis, University of Groningen, Groningen, The Netherlands, 2019. [Google Scholar]
- Wang, Y.F.; Belušič, G.; Pen, I.; Beukeboom, L.W.; Wertheim, B.; Stavenga, D.G.; Hut, R.A. Circadian rhythm entrainment of the jewel wasp, Nasonia vitripennis, by antagonistic interactions of multiple spectral inputs. Proc. Biol. Sci. 2023, 1992, 20222319. [Google Scholar] [CrossRef]
- Page, T.L.; Barrett, R.K. Effects of light on circadian pacemaker development. II. Responses to light. J. Comp. Physiol. A 1989, 1, 51–59. [Google Scholar] [CrossRef]
- Page, T.L.; Caldarola, P.C.; Pittendrigh, C.S. Mutual entrainment of bilaterally distributed circadian pacemakers. Proc. Natl. Acad. Sci. USA 1977, 3, 1277–1281. [Google Scholar] [CrossRef]
- Komada, S.; Kamae, Y.; Koyanagi, M.; Tatewaki, K.; Hassaneen, E.; Saifullah, A.S.M.; Yoshii, T.; Terakita, A.; Tomioka, K. Green-sensitive opsin is the photoreceptor for photic entrainment of an insect circadian clock. Zool. Lett. 2015, 1, 11. [Google Scholar] [CrossRef] [PubMed]
- van den Assem, J.; Jachmann, F. Changes in male perseverance in courtship and female readiness to mate in a strain of the parasitic wasp Nasonia Vitripennis over a period of 20+ years. Neth. J. Zool. 1999, 3, 125–137. [Google Scholar] [CrossRef]
- Perrot-Minnot, M.J.; Werren, J.H. Wolbachia infection and incompatibility dynamics in experimental selection lines. J. Evol. Biol. 1999, 12, 272–282. [Google Scholar] [CrossRef]
- Paolucci, S.; Dalla Benetta, E.; Salis, L.; Dolezel, D.; van de Zande, L.; Beukeboom, L.W. Latitudinal variation in circadian rhythmicity in Nasonia vitripennis. Behav. Sci. 2019, 11, 115. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 9, 1650–1667. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 11 March 2021).
- Ewels, P.; Magnusson, M.; Lundin, S.; Kaller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 19, 3047–3048. [Google Scholar] [CrossRef]
- Wingett, S.W.; Andrews, S. FastQ Screen: A tool for multi-genome mapping and quality control. F1000Research 2018, 7, 1338. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 15, 2114–2120. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 3, 290–295. [Google Scholar] [CrossRef]
- Pertea, G.; Pertea, M. GFF Utilities: GffRead and GffCompare. F1000Research 2020, 9, 304. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 1, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. InterProScan: Protein domains identifier. Nucleic Acids Res. 2005, 33, W116–W120. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.; Pethica, R.; Zhou, Y.; Talbot, C.; Vogel, C.; Madera, M.; Chothia, C.; Gough, J. SUPERFAMILY-sophisticated comparative genomics, data mining, visualization and phylogeny. Nucleic Acids Res. 2009, 37, D380–D386. [Google Scholar] [CrossRef]
- Finn, R.D.; Tate, J.; Mistry, J.; Coggill, P.C.; Sammut, S.J.; Hotz, H.; Ceric, G.; Forslund, K.; Eddy, S.R.; Sonnhammer, E.L.L.; et al. The Pfam protein families database. Nucleic Acids Res 2008, 36, D281–D288. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 12, 550. [Google Scholar] [CrossRef]
- Kolde, R. pheatmap: Pretty Heatmaps. 2019. Available online: https//CRAN.R-project.org/package=pheatmap (accessed on 16 June 2021).
- Brionne, A.; Juanchich, A.; Hennequet-Antier, C. ViSEAGO: A Bioconductor package for clustering biological functions using Gene Ontology and semantic similarity. BioData Min. 2019, 1, 16. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Wu, M.J.; Gu, L. TCseq: Time Course Sequencing Data Analysis. 2021. Available online: https://rdrr.io/bioc/TCseq/f/inst/doc/TCseq.pdf (accessed on 17 August 2021).
- Shen, W.; Le, S.; Li, Y.; Hu, F. SeqKit: A cross-platform and ultrafast toolkit for FASTA/Q file manipulation. PLoS ONE 2016, 10, e0163962. [Google Scholar] [CrossRef]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol. Cell 2010, 4, 576–589. [Google Scholar] [CrossRef]
- Nitta, K.R.; Jolma, A.; Yin, Y.; Morgunova, E.; Kivioja, T.; Akhtar, J.; Hens, K.; Toivonen, J.; Deplancke, B.; Furlong, E.E.M.; et al. Conservation of transcription factor binding specificities across 600 million years of bilateria evolution. eLife 2015, 4, e04837. [Google Scholar] [CrossRef] [PubMed]
- Wilson, V.; Conlon, F.L. The T-box family. Genome Biol. 2002, 6, reviews3008.1. [Google Scholar]
- Mira, H.; Morante, J. Neurogenesis from embryo to adult—Lessons from flies and mice. Front. Cell Dev. Biol. 2020, 8, 533. [Google Scholar] [CrossRef] [PubMed]
- Gluscevic, M.; Paradise, C.R.; Dudakovic, A.; Karperien, M.; Dietz, A.B.; van Wijnen, A.J.; Deyle, D.R. Functional expression of ZNF467 and PCBP2 supports adipogenic lineage commitment in adipose-derived mesenchymal stem cells. Gene 2020, 737, 144437. [Google Scholar] [CrossRef] [PubMed]
- Kuhlman, S.J.; Silver, R.; Le Sauter, J.; Bult-Ito, A.; McMahon, D.G. Phase resetting light pulses induce Per1 and persistent spike activity in a subpopulation of biological clock neurons. J. Neurosci. 2003, 4, 1441–1450. [Google Scholar] [CrossRef]
- Adewoye, A.B.; Kyriacou, C.P.; Tauber, E. Identification and functional analysis of early gene expression induced by circadian light-resetting in Drosophila. BMC Genom. 2015, 16, 570. [Google Scholar] [CrossRef]
- Crosio, C.; Cermakian, N.; Allis, C.D.; Sassone-Corsi, P. Light induces chromatin modification in cells of the mammalian circadian clock. Nat. Neurosci. 2000, 12, 1241–1247. [Google Scholar] [CrossRef]
- Etchegaray, J.P.; Lee, C.; Wade, P.A.; Reppert, S.M. Rhythmic histone acetylation underlies transcription in the mammalian circadian clock. Nature 2003, 6919, 177–182. [Google Scholar] [CrossRef]
- Naruse, Y.; Oh-hashi, K.; Iijima, N.; Naruse, M.; Yoshioka, H.; Tanaka, M. Circadian and light-induced transcription of clock gene Per1 depends on histone acetylation and deacetylation. Mol. Cell. Biol. 2004, 14, 6278–6287. [Google Scholar] [CrossRef]
- Alejevski, F.; Saint-Charles, A.; Michard-Vanhee, C.; Martin, B.; Galant, S.; Vasiliauskas, D.; Rouyer, F. The HisCl1 histamine receptor acts in photoreceptors to synchronize Drosophila behavioral rhythms with light-dark cycles. Nat. Commun. 2019, 10, 252. [Google Scholar] [CrossRef]
- Colwell, C.S.; Foster, R.G.; Menaker, M. Nmda receptor antagonists block the effects of light on circadian behavior in the mouse. Brain Res. 1991, 554, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.M.; Faiman, L.E.; Hurst, W.J.; Kuriashkina, L.R.; Gillette, M.U. Resetting the biological clock: Mediation of nocturnal CREB phosphorylation via light, glutamate, and nitric oxide. J. Neurosci. 1997, 2, 667–675. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Golombek, D.A. The rhythmic GABAergic system. Neurochem. Res. 1998, 5, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Ono, D.; Honma, K.; Yanagawa, Y.; Yamanaka, A.; Honma, S. Role of GABA in the regulation of the central circadian clock of the suprachiasmatic nucleus. J. Physiol. Sci. 2018, 4, 333–343. [Google Scholar] [CrossRef]
- Tischkau, S.A.; Mitchell, J.W.; Tyan, S.H.; Buchanan, G.F.; Gillette, M.U. Ca2+/cAMP response element-binding protein (CREB)-dependent activation of Per1 is required for light-induced signaling in the suprachiasmatic nucleus circadian clock. J. Biol. Chem. 2003, 2, 718–723. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J.S.; Reddy, A.B. The essential role of cAMP/Ca2+ signalling in mammalian circadian timekeeping. Biochem. Soc. Trans. 2012, 40, 44–50. [Google Scholar] [CrossRef]
- DeBruyne, J.P.; Weaver, D.R.; Reppert, S.M. CLOCK and NPAS2 have overlapping roles in the suprachiasmatic circadian clock. Nat. Neurosci. 2007, 10, 543–545. [Google Scholar] [CrossRef]
- Debruyne, J.P. Oscillating perceptions: The ups and downs of the CLOCK protein in the mouse circadian system. J. Genet. 2008, 5, 437–446. [Google Scholar] [CrossRef]
- Hut, R.A.; Oklejewicz, M.; Rieux, C.; Cooper, H.M. Photic sensitivity ranges of hamster pupillary and circadian phase responses do not overlap. J. Biol. Rhythm. 2008, 23, 37. [Google Scholar] [CrossRef]
- Aranda-Martínez, P.; Fernández-Martínez, J.; Ramírez-Casas, Y.; Guerra-Librero, A.; Rodríguez-Santana, C.; Escames, G.; Acuña-Castroviejo, D. The zebrafish, an outstanding model for biomedical research in the field of melatonin and human diseases. Int. J. Mol. Sci. 2022, 23, 7438. [Google Scholar] [CrossRef]
- Hirayama, J.; Cardone, L.; Doi, M.; Sassone-Corsi, P. Common pathways in circadian and cell cycle clocks: Light-dependent activation of Fos/AP-1 in zebrafish controls CRY-1a and WEE-1. Proc. Natl. Acad. Sci. USA 2005, 102, 10194–10199. [Google Scholar] [CrossRef] [PubMed]
- Kutaragi, Y.; Tokuoka, A.; Tomiyama, Y.; Nose, M.; Watanabe, T.; Bando, T.; Moriyama, Y.; Tomioka, K. A novel photic entrainment mechanism for the circadian clock in an insect: Involvement of c-fos and cryptochromes. Zool. Lett. 2018, 4, 26. [Google Scholar] [CrossRef] [PubMed]
- Levy, K.; Fishman, B.; Barnea, A.; Ayali, A.; Tauber, E. Transcriptional response of circadian clock genes to an ‘artificial light at night’ pulse in the cricket Gryllus bimaculatus. Int. J. Mol. Sci. 2022, 23, 11358. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Matsuka, M.; Shinohara, T.; Hamada, M.; Tomiyama, Y.; Tomioka, K. Fbxl4 regulates the photic entrainment of circadian locomotor rhythms in the cricket Gryllus bimaculatus. Zool. Sci. 2023, 40, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Foster, R. Light, sleep and circadian interactions: Biology to new therapeutic targets light, clocks and sleep: Clinical implications and new therapeutics. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4770. [Google Scholar]
- Jagannath, A.; Varga, N.; Dallmann, R.; Rando, G.; Gosselin, P.; Ebrahimjee, F.; Taylor, L.; Mosneagu, D.; Stefaniak, J.; Walsh, S.; et al. Adenosine integrates light and sleep signalling for the regulation of circadian timing in mice. Nat. Commun. 2021, 12, 2113. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Beukeboom, L.W.; Wertheim, B.; Hut, R.A. Transcriptomic Analysis of Light-Induced Genes in Nasonia vitripennis: Possible Implications for Circadian Light Entrainment Pathways. Biology 2023, 12, 1215. https://doi.org/10.3390/biology12091215
Wang Y, Beukeboom LW, Wertheim B, Hut RA. Transcriptomic Analysis of Light-Induced Genes in Nasonia vitripennis: Possible Implications for Circadian Light Entrainment Pathways. Biology. 2023; 12(9):1215. https://doi.org/10.3390/biology12091215
Chicago/Turabian StyleWang, Yifan, Leo W. Beukeboom, Bregje Wertheim, and Roelof A. Hut. 2023. "Transcriptomic Analysis of Light-Induced Genes in Nasonia vitripennis: Possible Implications for Circadian Light Entrainment Pathways" Biology 12, no. 9: 1215. https://doi.org/10.3390/biology12091215
APA StyleWang, Y., Beukeboom, L. W., Wertheim, B., & Hut, R. A. (2023). Transcriptomic Analysis of Light-Induced Genes in Nasonia vitripennis: Possible Implications for Circadian Light Entrainment Pathways. Biology, 12(9), 1215. https://doi.org/10.3390/biology12091215