
Preparation of Fibrinogen-Depleted Human Platelet Lysate to Support Heparin-Free Expansion of Umbilical Cord-Derived Mesenchymal Stem Cells

 ,
,  , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
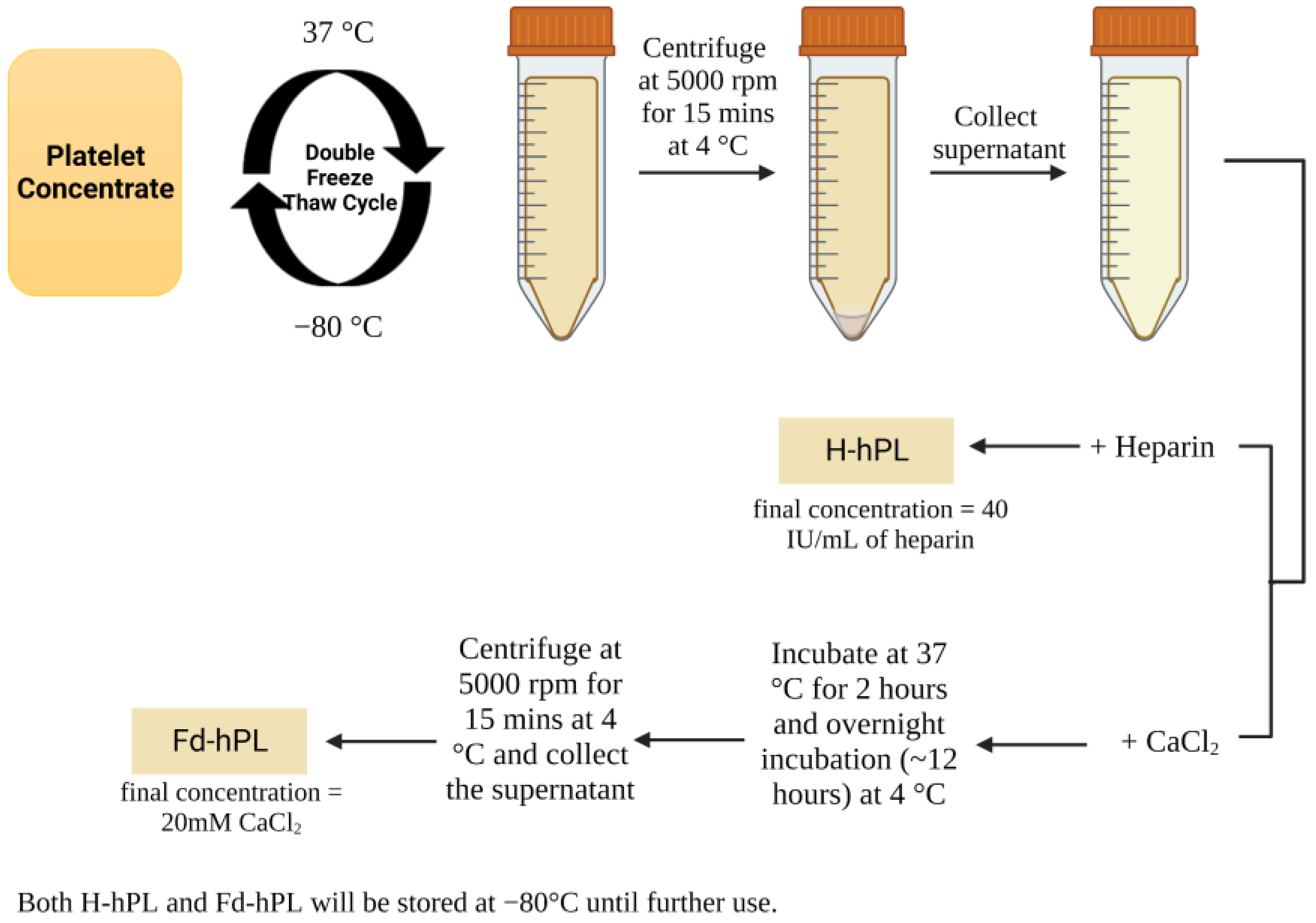
2.1. Human Platelet Lysate Preparation
2.2. Calcium Assay
2.3. Measurement of Fibrinogen and Growth Factor Concentration
2.4. Isolation and Culture of Umbilical Cord-Derived Mesenchymal Stem Cells (UC-MSCs)
2.5. Mesenchymal Stem Cell Characterization
2.6. Cell Morphology, Cell Size, Cell Viability, Total Cell Number, and Population Doubling Time Assessment
2.7. Statistical Analysis
3. Results
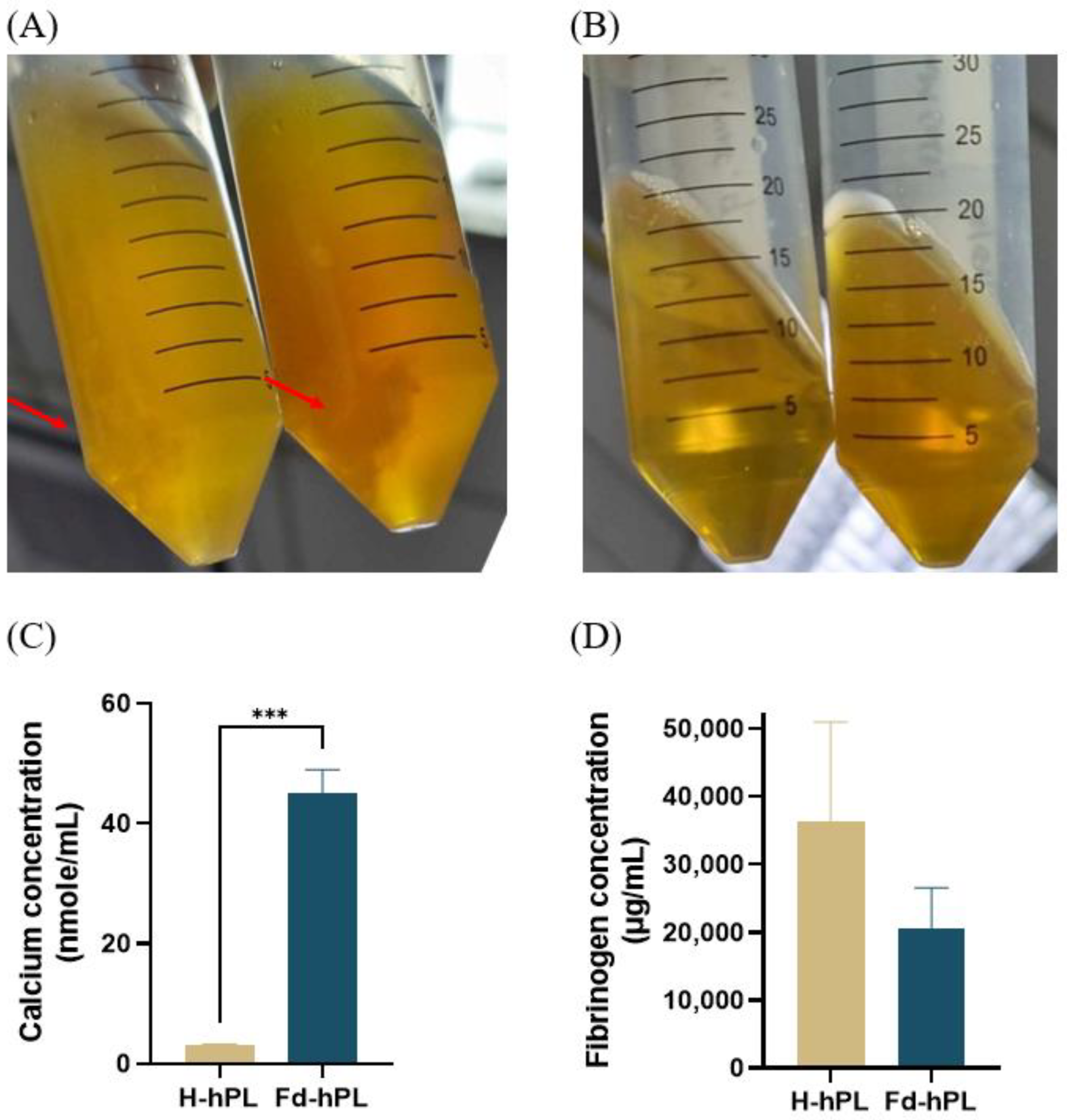
3.1. Fd-hPL Has a Higher Calcium Concentration and a More Noticeable Trend with a Lower Fibrinogen Concentration Compared to H-hPL
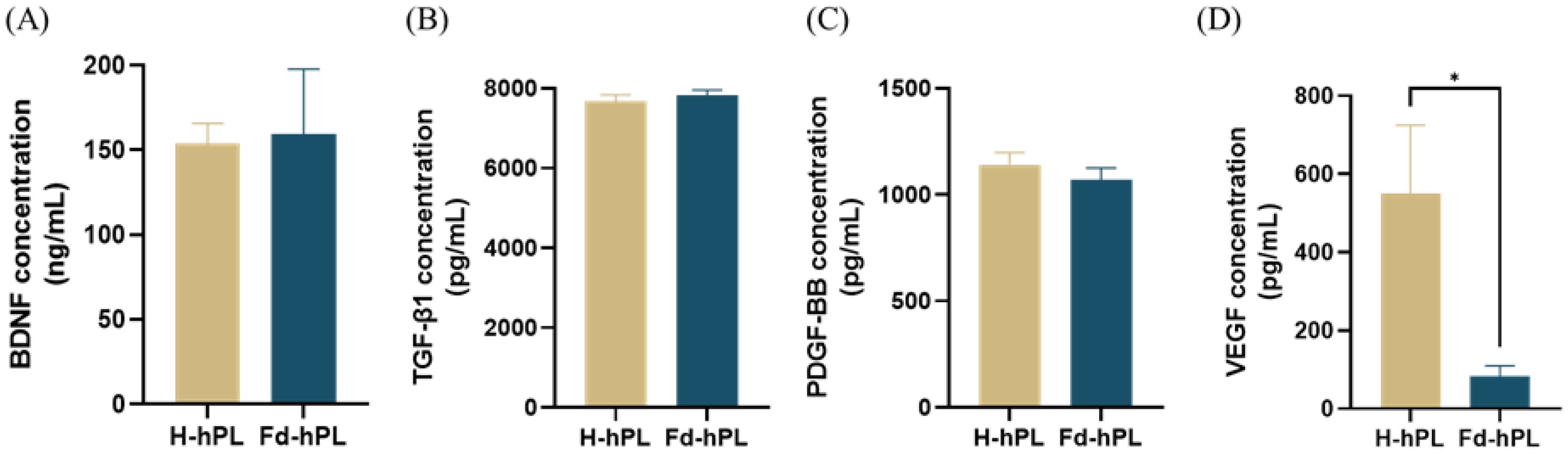
3.2. The Concentration of VEGF Was Reduced Significantly in Fd-hPL Compared to H-hPL
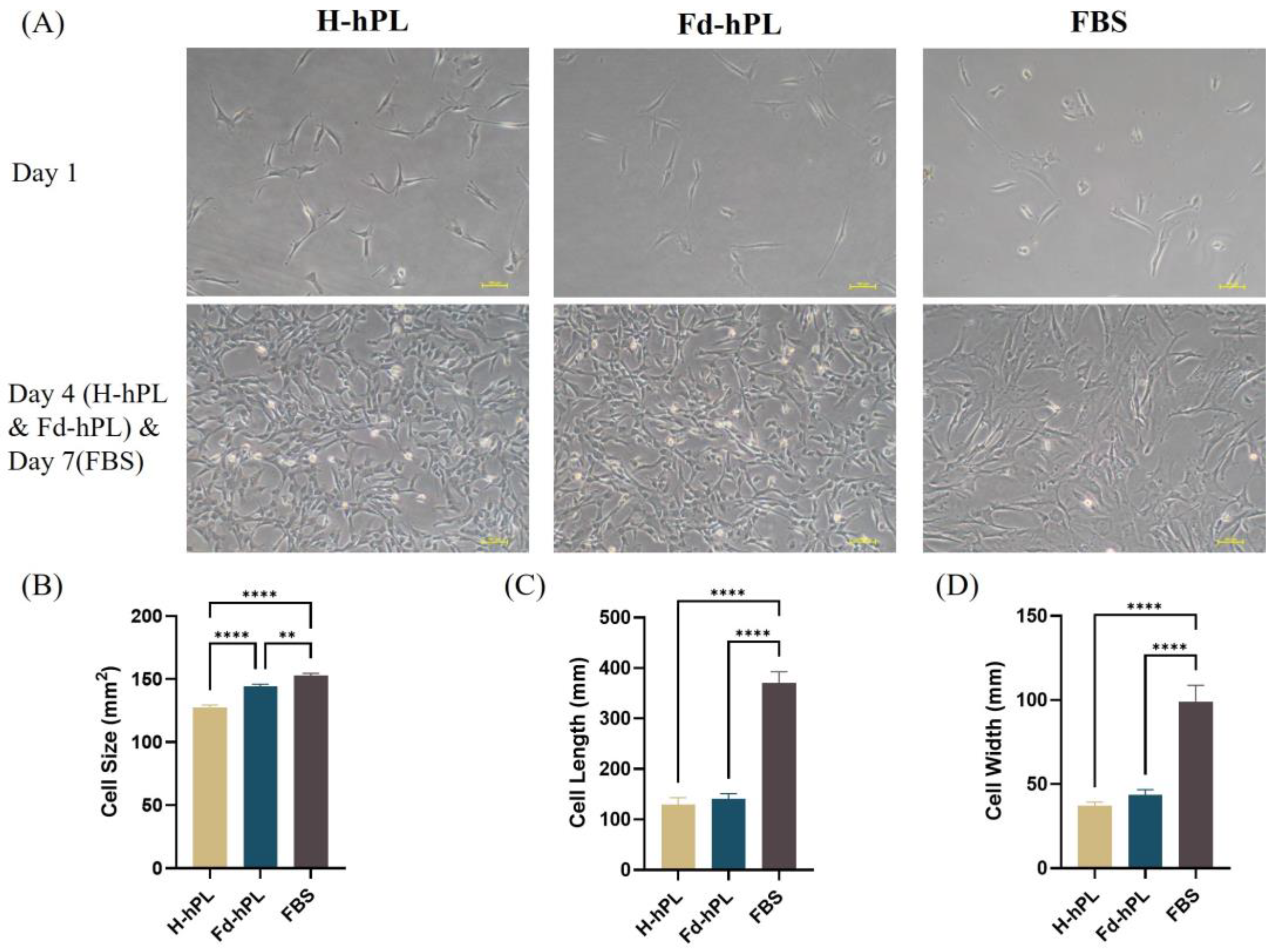
3.3. Morphology of UC-MSCs
3.4. UC-MSCs Grown in Fd-hPL Showed Typical MSC Surface Marker Expression Profiles
3.5. UC-MSCs Grown in Fd-hPL Retained Adipogenic and Osteogenic Differentiation Potential
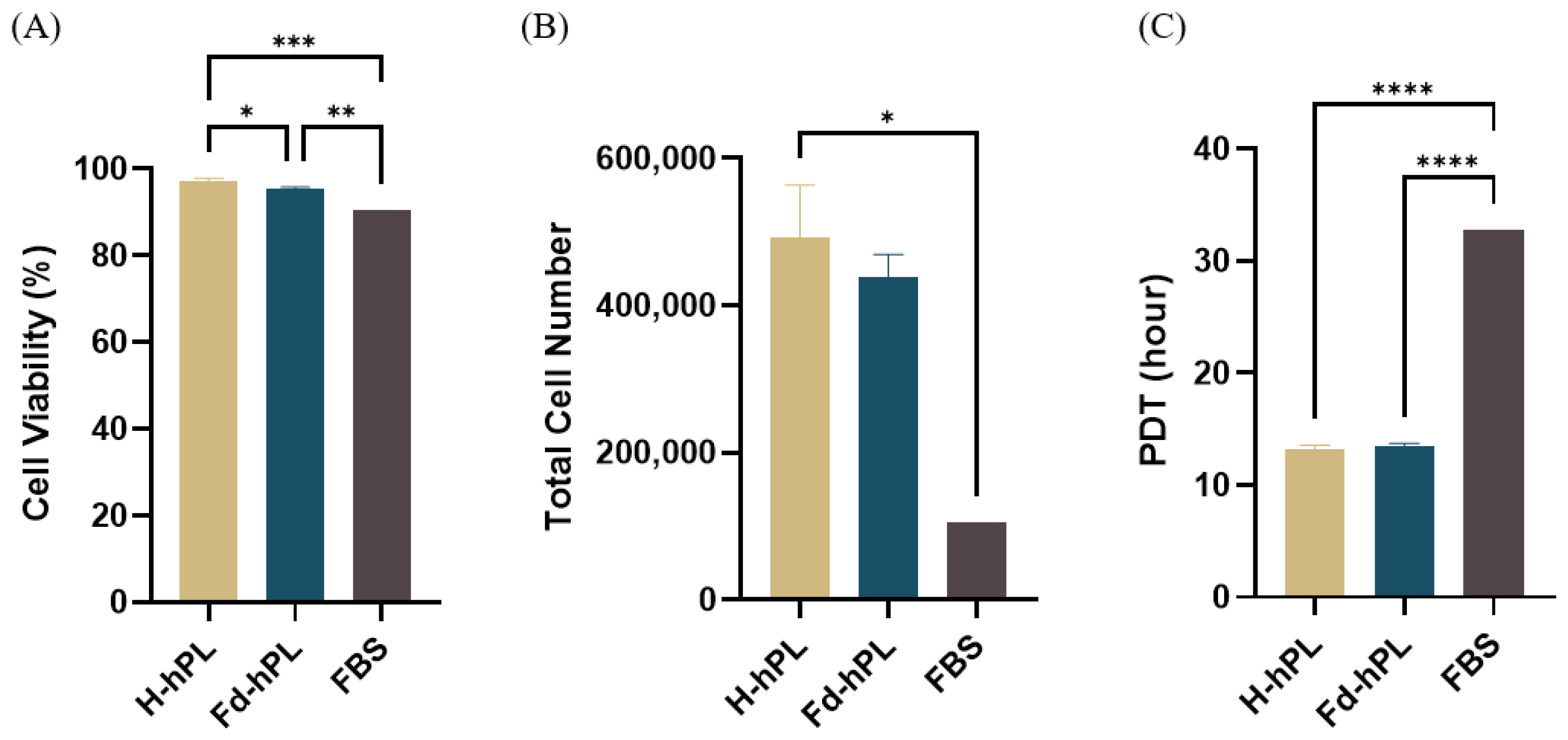
3.6. Cell Viability, Total Cell Number, and PDT of UC-MSCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guiotto, M.; Raffoul, W.; Hart, A.M.; Riehle, M.O.; Di Summa, P.G. Human Platelet Lysate to Substitute Fetal Bovine Serum in HMSC Expansion for Translational Applications: A Systematic Review. J. Transl. Med. 2020, 18, 1–14. [Google Scholar] [CrossRef]
- Hassan, M.N.F.B.; Yazid, M.D.; Yunus, M.H.M.; Chowdhury, S.R.; Lokanathan, Y.; Idrus, R.B.H.; Ng, A.M.H.; Law, J.X. Large-Scale Expansion of Human Mesenchymal Stem Cells. Stem Cells Int. 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Mackensen, A.; Dräger, R.; Schlesier, M.; Mertelsmann, R.; Lindemann, A. Presence of IgE Antibodies to Bovine Serum Albumin in a Patient Developing Anaphylaxis after Vaccination with Human Peptide-Pulsed Dendritic Cells. Cancer Immunol. Immunother. 2000, 49, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Dessels, C.; Potgieter, M.; Pepper, M.S. Making the Switch: Alternatives to Fetal Bovine Serum for Adipose-Derived Stromal Cell Expansion. Front. Cell Dev. Biol. 2016, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bieback, K.; Fernandez-Muñoz, B.; Pati, S.; Schäfer, R. Gaps in the Knowledge of Human Platelet Lysate as a Cell Culture Supplement for Cell Therapy: A Joint Publication from the AABB and the International Society for Cell & Gene Therapy. Transfusion 2019, 59, 3448–3460. [Google Scholar] [PubMed]
- National Blood Centre. Transfussion Practice Guidelines for Clinical and Laboratory Personnel; Ministry of Health: Putrajaya, Malaysia, 2016; Volume 4.
- Aubron, C.; Flint, A.; Ozier, Y.; McQuilten, Z. Transfusion of Stored Platelets: Balancing Risks and Product Availability. Int. J. Clin. Transfus. Med. 2016, 4, 133–138. [Google Scholar] [CrossRef]
- Dessels, C.; Durandt, C.; Pepper, M.S. Comparison of Human Platelet Lysate Alternatives Using Expired and Freshly Isolated Platelet Concentrates for Adipose-Derived Stromal Cell Expansion. Platelets 2019, 30, 356–367. [Google Scholar] [CrossRef]
- Glovinski, P.V.; Herly, M.; Mathiasen, A.B.; Svalgaard, J.D.; Borup, R.; Talman, M.L.M.; Elberg, J.J.; Kølle, S.F.T.; Drzewiecki, K.T.; Fischer-Nielsen, A. Overcoming the Bottleneck of Platelet Lysate Supply in Large-Scale Clinical Expansion of Adipose-Derived Stem Cells: A Comparison of Fresh versus Three Types of Platelet Lysates from Outdated Buffy Coat–Derived Platelet Concentrates. Cytotherapy 2017, 19, 222–234. [Google Scholar] [CrossRef]
- Bianchetti, A.; Chinello, C.; Guindani, M.; Braga, S.; Neva, A.; Verardi, R.; Piovani, G.; Pagani, L.; Lisignoli, G.; Magni, F.; et al. A Blood Bank Standardized Production of Human Platelet Lysate for Mesenchymal Stromal Cell Expansion: Proteomic Characterization and Biological Effects. Front. Cell Dev. Biol. 2021, 9, 1–14. [Google Scholar] [CrossRef]
- Delabie, W.; De Bleser, D.; Vandewalle, V.; Vandekerckhove, P.; Compernolle, V.; Feys, H.B. Single Step Method for High Yield Human Platelet Lysate Production. Transfusion 2022, 63, 373–383. [Google Scholar] [CrossRef]
- Astori, G.; Amati, E.; Bambi, F.; Bernardi, M.; Chieregato, K.; Schäfer, R.; Sella, S.; Rodeghiero, F. Platelet Lysate as a Substitute for Animal Serum for the Ex-Vivo Expansion of Mesenchymal Stem/Stromal Cells: Present and Future. Stem Cell Res. Ther. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- da Fonseca, L.; Santos, G.S.; Huber, S.C.; Setti, T.M.; Setti, T.; Lana, J.F. Human Platelet Lysate—A Potent (and Overlooked) Orthobiologic. J. Clin. Orthop. Trauma 2021, 21, 101534. [Google Scholar] [CrossRef]
- Jinhee, H.; Stephanie, A.S.-W.; Eric, B.R.; Walter, C.W.; Wu, K.; Yin, C.E.G. Effect of Heparin on the Biological Properties and Molecular Signature of Human Mesenchymal Stem Cells. J. Int. Soc. Burn Inj. 2017, 43, 909–932. [Google Scholar]
- Furue, M.K.; Na, J.; Jackson, J.P.; Okamoto, T.; Jones, M.; Baker, D.; Hata, R.I.; Moore, H.D.; Sato, J.D.; Andrews, P.W. Heparin Promotes the Growth of Human Embryonic Stem Cells in a Defined Serum-Free Medium. Proc. Natl. Acad. Sci. USA 2008, 105, 13409–13414. [Google Scholar] [CrossRef]
- Mimura, S.; Kimura, N.; Hirata, M.; Tateyama, D.; Hayashida, M.; Umezawa, A.; Kohara, A.; Nikawa, H.; Okamoto, T.; Furue, M.K. Growth Factor-Defined Culture Medium for Human Mesenchymal Stem Cells. Int. J. Dev. Biol. 2011, 55, 181–187. [Google Scholar] [CrossRef]
- Hassan, M.N.F.; Yap, Z.Y.; Tang, Y.L.; Ng, M.H.; Law, J.X. Expired Platelet Concentrate as a Source of Human Platelet Lysate for Xenogeneic-Free Culture of Human Dermal Fibroblasts. Sains Malaysiana 2021, 50, 2355–2365. [Google Scholar] [CrossRef]
- Liau, L.L.; Hassan, M.N.F.; Tang, Y.L.; Ng, M.H.; Law, J.X. Feasibility of Human Platelet Lysate as an Alternative to Foetal Bovine Serum for in Vitro Expansion of Chondrocytes. Int. J. Mol. Sci. 2021, 22, 1269. [Google Scholar] [CrossRef] [PubMed]
- Budi Harto, P.H.; Muhammad Hanif, M.; Othman, A.H.; Ngadenin, N.H.; Mohd Azahar, N.S.; Hassan, M.N.F.; Mohd Yahaya, N.H.; Rani, R.A.; Leong, C.F.; Ng, M.H.; et al. Human Platelet Lysate Promotes Proliferation but Fails to Maintain Chondrogenic Markers of Chondrocytes. Sains Malaysiana 2019, 48, 2169–2176. [Google Scholar] [CrossRef]
- Chan, A.M.L.; Ng, A.M.H.; Mohd Yunus, M.H.; Hj Idrus, R.; Law, J.X.; Yazid, M.D.; Chin, K.Y.; Shamsuddin, S.A.; Mohd Yusof, M.R.; Razali, R.A.; et al. Safety Study of Allogeneic Mesenchymal Stem Cell Therapy in Animal Model. Regen. Ther. 2022, 19, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, B.; Pretorius, E. Novel Pathway of Iron-Induced Blood Coagulation: Implications for Diabetes Mellitus and Its Complications. Pol. Arch. Med. Wewn. 2012, 122, 115–122. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Hu, G.; Su, J.; Li, W.; Chen, Q.; Shou, P.; Xu, C.; Chen, X.; Huang, Y.; Zhu, Z.; et al. Mesenchymal Stem Cells: A New Strategy for Immunosuppression and Tissue Repair. Cell Res. 2010, 20, 510–518. [Google Scholar] [PubMed]
- Even, M.S.; Sandusky, C.B.; Barnard, N.D. Serum-Free Hybridoma Culture: Ethical, Scientific and Safety Considerations. Trends Biotechnol. 2006, 24, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Copland, I.B.; Garcia, M.A.; Waller, E.K.; Roback, J.D.; Galipeau, J. The Effect of Platelet Lysate Fibrinogen on the Functionality of MSCs in Immunotherapy. Biomaterials 2013, 34, 7840–7850. [Google Scholar] [CrossRef]
- Laner-Plamberger, S.; Lener, T.; Schmid, D.; Streif, D.A.; Salzer, T.; Öller, M.; Hauser-Kronberger, C.; Fischer, T.; Jacobs, V.R.; Schallmoser, K.; et al. Mechanical Fibrinogen-Depletion Supports Heparin-Free Mesenchymal Stem Cell Propagation in Human Platelet Lysate. J. Transl. Med. 2015, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hemeda, H.; Giebel, B.; Wagner, W. Evaluation of Human Platelet Lysate versus Fetal Bovine Serum for Culture of Mesenchymal Stromal Cells. Cytotherapy 2014, 16, 170–180. [Google Scholar] [CrossRef]
- Mohamed, H.E.; Asker, M.E.; Kotb, N.S.; El Habab, A.M. Human Platelet Lysate Efficiency, Stability, and Optimal Heparin Concentration Required in Culture of Mammalian Cells. Blood Res. 2020, 55, 35–43. [Google Scholar] [CrossRef]
- Gurbuz, H.A.; Durukan, A.B.; Sevim, H.; Ergin, E.; Gurpinar, A.; Yorgancioglu, C. Heparin Toxicity in Cell Culture: A Critical Link in Translation of Basic Science to Clinical Practice. Blood Coagul. Fibrinolysis 2013, 24, 742–745. [Google Scholar] [CrossRef] [PubMed]
- Linkins, L.A. Heparin Induced Thrombocytopenia. BMJ 2015, 350, 1–11. [Google Scholar] [CrossRef]
- Gonzalez-Delgado, P.; Fernandez, J. Hypersensitivity Reactions to Heparins. Curr. Opin. Allergy Clin. Immunol. 2016, 16, 315–322. [Google Scholar] [CrossRef]
- Talib, U.; Lee, P. Heparin-Induced Hyperkalemia: The Un-Noticed Shadow. Chest 2019, 156, A1958. [Google Scholar] [CrossRef]
- Selich, A.; Zimmermann, K.; Tenspolde, M.; Dittrich-Breiholz, O.; Von Kaisenberg, C.; Schambach, A.; Rothe, M. Umbilical Cord as a Long-Term Source of Activatable Mesenchymal Stromal Cells for Immunomodulation. Stem Cell Res. Ther. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- El Omar, R.; Beroud, J.; Stoltz, J.F.; Menu, P.; Velot, E.; Decot, V. Umbilical Cord Mesenchymal Stem Cells: The New Gold Standard for Mesenchymal Stem Cell-Based Therapies? Tissue Eng.-Part B Rev. 2014, 20, 523–544. [Google Scholar] [CrossRef]
- Burnouf, T.; Strunk, D.; Koh, M.B.C.; Schallmoser, K. Human Platelet Lysate: Replacing Fetal Bovine Serum as a Gold Standard for Human Cell Propagation? Biomaterials 2016, 76, 371–387. [Google Scholar] [CrossRef] [PubMed]
- Salam, N.; Toumpaniari, S.; Gentile, P.; Ferreira, A.M.; Dalgarno, K.; Partridge, S. Assessment of Migration of Human MSCs through Fibrin Hydrogels as a Tool for Formulation Optimisation. Materials 2018, 11, 1781. [Google Scholar] [CrossRef]
- Guo, M.; Sahni, S.K.; Sahni, A.; Francis, C.W. Fibrinogen Regulates the Expression of Inflammatory Chemokines through NF-ΚB Activation of Endothelial Cells. Thromb. Haemost. 2004, 92, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Thacker, R.I.; Retzinger, G.S. Adsorbed Fibrinogen Regulates the Behavior of Human Dendritic Cells in a CD18-Dependent Manner. Exp. Mol. Pathol. 2008, 84, 122–130. [Google Scholar] [CrossRef]
- Masamune, A.; Kikuta, K.; Watanabe, T.; Satoh, K.; Hirota, M.; Hamada, S.; Shimosegawa, T. Fibrinogen Induces Cytokine and Collagen Production in Pancreatic Stellate Cells. Gut 2009, 58, 550–559. [Google Scholar] [CrossRef]
- Oeller, M.; Laner-plamberger, S.; Krisch, L.; Rohde, E.; Strunk, D.; Schallmoser, K. Human Platelet Lysate for Good Manufacturing Practice-compliant Cell Production. Int. J. Mol. Sci. 2021, 22, 5178. [Google Scholar] [CrossRef]
- Sahni, A.; Francis, C.W. Vascular Endothelial Growth Factor Binds to Fibrinogen and Fibrin and Stimulates Endothelial Cell Proliferation. Blood 2000, 96, 3772–3778. [Google Scholar] [CrossRef]
- Martino, M.M.; Briquez, P.S.; Ranga, A.; Lutolf, M.P.; Hubbell, J.A. Heparin-Binding Domain of Fibrin(Ogen) Binds Growth Factors and Promotes Tissue Repair When Incorporated within a Synthetic Matrix. Proc. Natl. Acad. Sci. USA 2013, 110, 4563–4568. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.K.; Lu, Q.Z.; Pei, R.; Ji, H.J.; Zhou, G.S.; Zhao, X.L.; Tang, R.K.; Zhang, M. The Effect of Extracellular Calcium and Inorganic Phosphate on the Growth and Osteogenic Differentiation of Mesenchymal Stem Cells in Vitro: Implication for Bone Tissue Engineering. Biomed. Mater. 2009, 4, 025004. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, J.R.; Heitman, J. Calcineurin Is Required for Candida albicans to Survive Calcium Stress in Serum. Infect. Immun. 2005, 73, 5767–5774. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Feng, F.; Wang, Q.; Zhu, X.; Fu, H.; Xu, L.; Liu, K.; Huang, X.; Zhang, X. Platelet-Derived Growth Factor-BB Protects Mesenchymal Stem Cells (MSCs) Derived From Immune Thrombocytopenia Patients Against Apoptosis and Senescence and Maintains MSC-Mediated Immunosuppression. Tissue-Specific Progenit. Stem Cellls 2016, 5, 1631–1643. [Google Scholar] [CrossRef]
- Qiu, P.; Song, W.; Niu, Z.; Bai, Y.; Li, W.; Pan, S.; Peng, S.; Hua, J. Platelet-Derived Growth Factor Promotes the Proliferation of Human Umbilical Cord-Derived Mesenchymal Stem Cells. Cell Biochem. Funct. 2013, 31, 159–165. [Google Scholar] [CrossRef]
- Ball, S.G.; Shuttleworth, C.A.; Kielty, C.M. Mesenchymal Stem Cells and Neovascularization: Role of Platelet-Derived Growth Factor Receptors. J. Cell. Mol. Med. 2007, 11, 1012–1030. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kee, L.T.; Lee, Y.T.; Ng, C.Y.; Hassan, M.N.F.; Ng, M.H.; Mahmood, Z.; Abdul Aziz, S.; Law, J.X. Preparation of Fibrinogen-Depleted Human Platelet Lysate to Support Heparin-Free Expansion of Umbilical Cord-Derived Mesenchymal Stem Cells. Biology 2023, 12, 1085. https://doi.org/10.3390/biology12081085
Kee LT, Lee YT, Ng CY, Hassan MNF, Ng MH, Mahmood Z, Abdul Aziz S, Law JX. Preparation of Fibrinogen-Depleted Human Platelet Lysate to Support Heparin-Free Expansion of Umbilical Cord-Derived Mesenchymal Stem Cells. Biology. 2023; 12(8):1085. https://doi.org/10.3390/biology12081085
Chicago/Turabian StyleKee, Li Ting, Yi Ting Lee, Chiew Yong Ng, Muhammad Najib Fathi Hassan, Min Hwei Ng, Zalina Mahmood, Suria Abdul Aziz, and Jia Xian Law. 2023. "Preparation of Fibrinogen-Depleted Human Platelet Lysate to Support Heparin-Free Expansion of Umbilical Cord-Derived Mesenchymal Stem Cells" Biology 12, no. 8: 1085. https://doi.org/10.3390/biology12081085
APA StyleKee, L. T., Lee, Y. T., Ng, C. Y., Hassan, M. N. F., Ng, M. H., Mahmood, Z., Abdul Aziz, S., & Law, J. X. (2023). Preparation of Fibrinogen-Depleted Human Platelet Lysate to Support Heparin-Free Expansion of Umbilical Cord-Derived Mesenchymal Stem Cells. Biology, 12(8), 1085. https://doi.org/10.3390/biology12081085