An Overview of Selected Bacterial Infections in Cancer, Their Virulence Factors, and Some Aspects of Infection Management


Simple Summary
Abstract
1. Introduction
2. Helicobacter pylori and Gastric Adenocarcinomas
3. Gastric MALT Lymphomas
4. Colon Cancer—Connection with Bacterial Pathology
5. Chlamydia Species: Their Virulence Factors and Involvement in Different Diseases Including Cancer
6. Pseudomonas aeruginosa and Cancer
7. Systemic Cancer Therapy—Immunity and Infection
8. Antimicrobial Stewardship in Cancer Patients
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leitão, J.H. Microbial virulence factors. Int. J. Mol. Sci. 2020, 21, 5320. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.; Castillo-Pino, E. An introduction to the epidemiology and burden of urinary tract infections. Ther. Adv. Urol. 2019, 11, 1756287219832172. [Google Scholar] [CrossRef] [PubMed]
- Bien, J.; Sokolova, O.; Bozko, P. Role of uropathogenic Escherichia coli virulence factors in development of urinary tract infection and kidney damage. Int. J. Nephrol. 2012, 2012, 681473. [Google Scholar]
- Khan, A.A.; Khan, Z.; Malik, A.; Kalam, M.A.; Cash, P.; Ashraf, M.T.; Alshamsan, A. Colorectal cancer-inflammatory bowel disease nexus and felony of Escherichia coli. Life Sci. 2017, 180, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.; Muthunatarajan, S.; Mulki, S.S.; Archana Bhat, K.; Kotian, K.H. Bacterial infection among cancer patients: Analysis of isolates and antibiotic sensitivity pattern. Int. J. Microbiol. 2021, 2021, 8883700. [Google Scholar] [CrossRef] [PubMed]
- Islas-Muñoz, B.; Volkow-Fernández, P.; Ibanes-Gutiérrez, C.; Villamar-Ramírez, A.; Vilar-Compte, D.; Cornejo-Juárez, P. Bloodstream infections in cancer patients. Risk factors associated with mortality. Int. J. Infect. Dis. 2018, 71, 59–64. [Google Scholar] [CrossRef]
- Perez, F.; Adachi, J.; Bonomo, R.A. Antibiotic-resistant gram-negative bacterial infections in patients with cancer. Clin. Infect. Dis. 2014, 59 (Suppl. 5), S335–S339. [Google Scholar] [CrossRef]
- Gagnaire, A.; Nadel, B.; Raoult, D.; Neefjes, J.; Gorvel, J.P. Collateral damage: Insights into bacterial mechanisms that predispose host cells to cancer. Nat. Rev. Microbiol. 2017, 15, 109–128. [Google Scholar] [CrossRef]
- Mager, D.L. Bacteria and cancer: Cause, coincidence or cure? A review. J. Transl. Med. 2006, 4, 14. [Google Scholar] [CrossRef]
- Sun, J. Impact of bacterial infection and intestinal microbiome on colorectal cancer development. Chin. Med. J. 2022, 135, 400–408. [Google Scholar] [CrossRef]
- Chorobik, P.; Czaplicki, D.; Ossysek, K.; Bereta, J. Salmonella and cancer: From pathogens to therapeutics. Acta Biochim. Pol. 2013, 60, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Xiao, W.; Yang, Y.; Li, H.; Xia, D.; Yu, G.; Guo, X.; Guan, W.; Hu, Z.; Xu, H.; et al. Pseudomonas aeruginosa-mannose-sensitive hemagglutinin inhibits epidermal growth factor receptor signaling pathway activation and induces apoptosis in bladder cancer cells in vitro and in vivo. Urol. Oncol. 2014, 32, 36.e11-8. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Rivera, D.; González, O.; Guzmán-Rodríguez, J.; Díaz-Pérez, A.L.; Ochoa-Zarzosa, A.; López-Bucio, J.; Meza-Carmen, V.; Campos-García, J. Cytotoxicity of cyclodipeptides from Pseudomonas aeruginosa PAO1 leads to apoptosis in human cancer cell lines. Biomed. Res. Int. 2015, 2015, 197608. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zhu, J. Recent development and optimization of Pseudomonas aeruginosa exotoxin immunotoxins in cancer therapeutic applications. Int. Immunopharmacol. 2021, 96, 107759. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, P. Helicobacter pylori and inflammation. Curr. Pharm. Des. 2010, 16, 4225–4236. [Google Scholar] [CrossRef] [PubMed]
- Marshall, B.J. The Campylobacter pylori story. Scand. J. Gastroenterol. Suppl. 1988, 146, 58–66. [Google Scholar] [CrossRef]
- Rathbone, B.; Wyatt, J. Campylobacter pylori and precancerous lesions. Cancer Lett. 1988, 39, S14. [Google Scholar] [CrossRef]
- Correa, P.; Fox, J.; Fontham, E.; Ruiz, B.; Lin, Y.P.; Zavala, D.; Taylor, N.; Mackinley, D.; de Lima, E.; Portilla, H.; et al. Helicobacter pylori and gastric carcinoma. Serum antibody prevalence in populations with contrasting cancer risks. Cancer 1990, 66, 2569–2574. [Google Scholar]
- Loffeld, R.J.; Willems, I.; Flendrig, J.A.; Arends, J.W. Helicobacter pylori and gastric carcinoma. Histopathology 1990, 17, 537–541. [Google Scholar] [CrossRef]
- Chiba, T.; Marusawa, H.; Seno, H.; Watanabe, N. Mechanism for gastric cancer development by Helicobacter pylori infection. J. Gastroenterol. Hepatol. 2008, 23, 1175–1181. [Google Scholar]
- Soutto, M.; Bhat, N.; Khalafi, S.; Zhu, S.; Poveda, J.; Garcia-Buitrago, M.; Zaika, A.; El-Rifai, W. NF-kB-dependent activation of STAT3 by H. pylori is suppressed by TFF1. Cancer Cell Int. 2021, 21, 444. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Cho, S.J.; Ko, Y.S.; Park, J.; Shin, D.H.; Hwang, I.C.; Han, S.Y.; Nam, S.Y.; Kim, M.A.; Chang, M.S.; et al. A synergistic interaction between transcription factors nuclear factor-κB and signal transducers and activators of transcription 3 promotes gastric cancer cell migration and invasion. BMC Gastroenterol. 2013, 13, 29. [Google Scholar] [CrossRef]
- Doger, F.K.; Meteoglu, I.; Ozkara, E.; Erkul, Z.K.; Okyay, P.; Yükselen, V. Expression of NF-κB in Helicobacter pylori infection. Dig. Dis. Sci. 2006, 51, 2306–2309. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, R.L.; Avé, P.; Ndiaye, D.; Bambou, J.C.; Huerre, M.R.; Philpott, D.J.; Mémet, S. NF-κB activation during acute Helicobacter pylori infection in mice. Infect. Immun. 2008, 76, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Yoshida, H.; Ogura, K.; Mitsuno, Y.; Hirata, Y.; Yamaji, Y.; Akanuma, M.; Shiratori, Y.; Omata, M.H. pylori activates NF-κB through a signaling pathway involving IκB kinases, NF-κB—Inducing kinase, TRAF2, and TRAF6 in gastric cancer cells. Gastroenterology 2000, 119, 97–108. [Google Scholar] [CrossRef]
- Lamb, A.; Chen, J.; Blanke, S.R.; Chen, L.F. Helicobacter pylori activates NF-κB by inducing Ubc13-mediated ubiquitination of lysine 158 of TAK1. J. Cell. Biochem. 2013, 114, 2284–2292. [Google Scholar] [CrossRef]
- Li, N.; Xie, C.; Lu, N.H. p53, a potential predictor of Helicobacter pylori infection-associated gastric carcinogenesis? Oncotarget. 2016, 7, 66276–66286. [Google Scholar] [CrossRef]
- Kodama, M.; Murakami, K.; Okimoto, T.; Sato, R.; Watanabe, K.; Fujioka, T. Expression of mutant type-p53 products in H. pylori-associated chronic gastritis. World J. Gastroenterol. 2007, 13, 1541–1546. [Google Scholar] [CrossRef][Green Version]
- Abu-Lubad, M.A.; Helaly, G.F.; Haddadin, W.J.; Jarajreh, D.A.K.; Aqel, A.A.; Al-Zeer, M.A. Loss of p53 expression in gastric epithelial cells of Helicobacter pylori-infected Jordanian patients. Int. J. Microbiol. 2022, 2022, 7779770. [Google Scholar] [CrossRef]
- Rahman, M.M.; Sarker, M.A.K.; Hossain, M.M.; Alam, M.S.; Islam, M.M.; Shirin, L.; Sultana, R.; Sultana, G.N.N. Association of p53 gene mutation with Helicobacter pylori infection in gastric cancer patients and its correlation with clinicopathological and environmental factors. World J. Oncol. 2019, 10, 46–54. [Google Scholar] [CrossRef]
- Bhardwaj, V.; Noto, J.M.; Wei, J.; Andl, C.; El-Rifai, W.; Peek, R.M.; Zaika, A.I. Helicobacter pylori bacteria alter the p53 stress response via ERK-HDM2 pathway. Oncotarget 2015, 6, 1531–1543. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Dang, S.; Hou, P. Gene methylation in gastric cancer. Clin. Chim. Acta 2013, 424, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Compare, D.; Rocco, A.; Liguori, E.; D’Armiento, F.P.; Persico, G.; Masone, S.; Coppola-Bottazzi, E.; Suriani, R.; Romano, M.; Nardone, G. Global DNA hypomethylation is an early event in Helicobacter pylori-related gastric carcinogenesis. J. Clin. Pathol. 2011, 64, 677–682. [Google Scholar] [CrossRef]
- Zhang, B.G.; Hu, L.; Zang, M.D.; Wang, H.X.; Zhao, W.; Li, J.F.; Su, L.P.; Shao, Z.; Zhao, X.; Zhu, Z.G.; et al. Helicobacter pylori CagA induces tumor suppressor gene hypermethylation by upregulating DNMT1 via AKT-NFκB pathway in gastric cancer development. Oncotarget 2016, 7, 9788–9800. [Google Scholar] [CrossRef]
- Alipour, M. Molecular mechanism of Helicobacter pylori-induced gastric cancer. J. Gastrointest. Cancer 2021, 52, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Díaz, P.; Valenzuela Valderrama, M.; Bravo, J.; Quest, A.F.G. Helicobacter pylori and gastric cancer: Adaptive cellular mechanisms involved in disease progression. Front. Microbiol. 2018, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, L.; Ji, J.; Zhang, J.; Yan, M.; Zhang, J.; Liu, B.; Zhu, Z.; Yu, Y. ABO blood group system and gastric cancer: A case-control study and meta-analysis. Int. J. Mol. Sci. 2012, 13, 13308–13321. [Google Scholar] [CrossRef]
- Franchini, M.; Liumbruno, G.M.; Lippi, G. The prognostic value of ABO blood group in cancer patients. Blood Transfus. 2016, 14, 434–440. [Google Scholar]
- Salar, A. Gastric MALT lymphoma and Helicobacter pylori. Med. Clin. 2019, 152, 65–71. [Google Scholar] [CrossRef]
- Lehours, P.; Ménard, A.; Dupouy, S.; Bergey, B.; Richy, F.; Zerbib, F.; Ruskoné-Fourmestraux, A.; Delchier, J.C.; Mégraud, F. Evaluation of the association of nine Helicobacter pylori virulence factors with strains involved in low-grade gastric mucosa-associated lymphoid tissue lymphoma. Infect. Immun. 2004, 72, 880–888. [Google Scholar] [CrossRef]
- Floch, P.; Mégraud, F.; Lehours, P. Helicobacter pylori strains and gastric MALT lymphoma. Toxins 2017, 9, 132. [Google Scholar] [CrossRef] [PubMed]
- Suarez, F.; Lortholary, O.; Hermine, O.; Lecuit, M. Infection-associated lymphomas derived from marginal zone B cells: A model of antigen-driven lymphoproliferation. Blood 2006, 107, 3034–3044. [Google Scholar] [CrossRef] [PubMed]
- Marcelis, L.; Tousseyn, T.; Sagaert, X. MALT lymphoma as a model of chronic inflammation-induced gastric tumor development. Curr. Top. Microbiol. Immunol. 2019, 421, 77–106. [Google Scholar] [PubMed]
- Gong, E.J.; Ahn, J.Y.; Jung, H.Y.; Park, H.; Ko, Y.B.; Na, H.K.; Jung, K.W.; Kim, D.H.; Lee, J.H.; Choi, K.D.; et al. Helicobacter pylori eradication therapy is effective as the initial treatment for patients with H. pylori-negative and disseminated gastric mucosa-associated lymphoid tissue lymphoma. Gut Liver 2016, 10, 706–713. [Google Scholar] [CrossRef]
- Kim, J.S.; Kang, S.H.; Moon, H.S.; Sung, J.K.; Jeong, H.Y. Clinical outcome of eradication therapy for gastric mucosa-associated lymphoid tissue lymphoma according to H. pylori infection status. Gastroenterol. Res. Pract. 2016, 2016, 6794848. [Google Scholar] [CrossRef]
- Diaconu, S.; Predescu, A.; Moldoveanu, A.; Pop, C.S.; Fierbințeanu-Braticevici, C. Helicobacter pylori infection: Old and new. J. Med. Life 2017, 10, 112–117. [Google Scholar]
- Violeta Filip, P.; Cuciureanu, D.; Sorina Diaconu, L.; Maria Vladareanu, A.; Silvia Pop, C. MALT lymphoma: Epidemiology, clinical diagnosis and treatment. J. Med. Life 2018, 11, 187–193. [Google Scholar] [CrossRef]
- Herlevic, V.; Morris, J.D. Gastric Lymphoma; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Du, M.Q.; Atherton, J.C. Molecular subtyping of gastric MALT lymphomas: Implications for prognosis and management. Gut 2006, 55, 886–893. [Google Scholar] [CrossRef]
- Zhang, M. High antibiotic resistance rate: A difficult issue for Helicobacter pylori eradication treatment. World J. Gastroenterol. 2015, 21, 13432–13437. [Google Scholar] [CrossRef]
- Wang, Y.G.; Zhao, L.Y.; Liu, C.Q.; Pan, S.C.; Chen, X.L.; Liu, K.; Zhang, W.H.; Yang, K.; Chen, X.Z.; Zhang, B.; et al. Clinical characteristics and prognostic factors of primary gastric lymphoma: A retrospective study with 165 cases. Medicine 2016, 95, e4250. [Google Scholar] [CrossRef]
- Thieblemont, C.; Cascione, L.; Conconi, A.; Kiesewetter, B.; Raderer, M.; Gaidano, G.; Martelli, M.; Laszlo, D.; Coiffier, B.; Lopez Guillermo, A.; et al. A MALT lymphoma prognostic index. Blood 2017, 130, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Bacon, C.M.; Du, M.Q.; Dogan, A. Mucosa-associated lymphoid tissue (MALT) lymphoma: A practical guide for pathologists. J. Clin. Pathol. 2007, 60, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Quach, M.A.; Ake, C.D.; Chen, M.; Wang, J. Gastrointestinal lymphomas: Morphology, immunophenotype and molecular features. J Gastrointest. Oncol. 2012, 3, 209–225. [Google Scholar] [PubMed]
- Wotherspoon, A. Pathology of extranodal marginal zone lymphoma at different anatomic sites. Ann. Lymphoma 2020, 4, 15. [Google Scholar] [CrossRef]
- Raderer, M.; Kiesewetter, B. What you always wanted to know about gastric MALT-lymphoma: A focus on recent developments. Ther. Adv. Med. Oncol. 2021, 13, 17588359211033825. [Google Scholar] [CrossRef]
- Akoum, R.; Serhal, W.; Farhat, H. Disseminated gastric MALT lymphoma with monoclonal gammopathy, t(11;18)(q21;q21), and subsequent development of T-large granular lymphocytic leukemia: A case report and review of the literature. Case Rep. Med. 2015, 2015, 953297. [Google Scholar] [CrossRef]
- Capelle, L.G.; de Vries, A.C.; Looman, C.W.; Casparie, M.K.; Boot, H.; Meijer, G.A.; Kuipers, E.J. Gastric MALT lymphoma: Epidemiology and high adenocarcinoma risk in a nation-wide study. Eur. J. Cancer 2008, 44, 2470–2476. [Google Scholar] [CrossRef]
- Zullo, A.; Rago, A.; Felici, S.; Licci, S.; Ridola, L.; Caravita di Toritto, T. Onset and progression of precancerous lesions on gastric mucosa of patients treated for gastric lymphoma. J. Gastrointestin. Liver Dis. 2020, 29, 27–31. [Google Scholar] [CrossRef]
- Arnold, M.; Abnet, C.C.; Neale, R.E.; Vignat, J.; Giovannucci, E.L.; McGlynn, K.A.; Bray, F. Global burden of 5 major types of gastrointestinal cancer. Gastroenterology 2020, 159, 335–349.e15. [Google Scholar] [CrossRef]
- Mirzarazi, M.; Bashiri, S.; Hashemi, A.; Vahidi, M.; Kazemi, B.; Bandehpour, M. The OmpA of commensal Escherichia coli of CRC patients affects apoptosis of the HCT116 colon cancer cell line. BMC Microbiol. 2022, 22, 139. [Google Scholar] [CrossRef]
- Périchon, B.; Lichtl-Häfele, J.; Bergsten, E.; Delage, V.; Trieu-Cuot, P.; Sansonetti, P.; Sobhani, I.; Dramsi, S. Detection of Streptococcus gallolyticus and four other CRC-associated bacteria in patient stools reveals a potential “Driver” role for Enterotoxigenic Bacteroides fragilis. Front. Cell. Infect. Microbiol. 2022, 12, 794391. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Jiao, X.; Zeng, M.; Fan, Z.; Li, X.; Yuan, Y.; Zhang, Q.; Xia, Y. Clinical significance of Fusobacterium nucleatum and microsatellite instability in evaluating colorectal cancer prognosis. Cancer Manag. Res. 2022, 14, 3021–3036. [Google Scholar] [CrossRef] [PubMed]
- Bertocchi, A.; Carloni, S.; Ravenda, P.S.; Bertalot, G.; Spadoni, I.; Lo Cascio, A.; Gandini, S.; Lizier, M.; Braga, D.; Asnicar, F.; et al. Gut vascular barrier impairment leads to intestinal bacteria dissemination and colorectal cancer metastasis to liver. Cancer Cell. 2021, 39, 708–724.e11. [Google Scholar] [CrossRef] [PubMed]
- Butt, J.; Jenab, M.; Werner, J.; Fedirko, V.; Weiderpass, E.; Dahm, C.C.; Tjønneland, A.; Olsen, A.; Boutron-Ruault, M.C.; Rothwell, J.A.; et al. Association of pre-diagnostic antibody responses to Escherichia coli and Bacteroides fragilis toxin proteins with colorectal cancer in a European cohort. Gut Microbes 2021, 13, 1903825. [Google Scholar] [CrossRef]
- Cuellar-Gómez, H.; Ocharán-Hernández, M.E.; Calzada-Mendoza, C.C.; Comoto-Santacruz, D.A. Association of Fusobacterium nucleatum infection and colorectal cancer: A Mexican study. Rev. Gastroenterol. Mex. 2022, 87, 277–284. [Google Scholar] [CrossRef]
- Khodaverdi, N.; Zeighami, H.; Jalilvand, A.; Haghi, F.; Hesami, N. High frequency of enterotoxigenic Bacteroides fragilis and Enterococcus faecalis in the paraffin-embedded tissues of Iranian colorectal cancer patients. BMC Cancer 2021, 21, 1353. [Google Scholar] [CrossRef]
- Kong, C.; Yan, X.; Zhu, Y.; Zhu, H.; Luo, Y.; Liu, P.; Ferrandon, S.; Kalady, M.F.; Gao, R.; He, J.; et al. Fusobacterium nucleatum promotes the development of colorectal cancer by activating a cytochrome P450/epoxyoctadecenoic acid axis via TLR4/Keap1/NRF2 signaling. Cancer Res. 2021, 81, 4485–4498. [Google Scholar] [CrossRef]
- Nardelli, C.; Granata, I.; Nunziato, M.; Setaro, M.; Carbone, F.; Zulli, C.; Pilone, V.; Capoluongo, E.D.; De Palma, G.D.; Corcione, F.; et al. 16S rRNA of mucosal colon microbiome and CCL2 circulating levels are potential biomarkers in colorectal cancer. Int. J. Mol. Sci. 2021, 22, 10747. [Google Scholar] [CrossRef]
- Pignatelli, P.; Iezzi, L.; Pennese, M.; Raimondi, P.; Cichella, A.; Bondi, D.; Grande, R.; Cotellese, R.; Di Bartolomeo, N.; Innocenti, P.; et al. The potential of colonic tumor tissue Fusobacterium nucleatum to predict staging and its interplay with oral abundance in colon cancer patients. Cancers 2021, 13, 1032. [Google Scholar] [CrossRef]
- Iyadorai, T.; Mariappan, V.; Vellasamy, K.M.; Wanyiri, J.W.; Roslani, A.C.; Lee, G.K.; Sears, C.; Vadivelu, J. Prevalence and association of pks+ Escherichia coli with colorectal cancer in patients at the University Malaya Medical Centre, Malaysia. PLoS ONE 2020, 15, e0228217. [Google Scholar] [CrossRef]
- Zamani, S.; Taslimi, R.; Sarabi, A.; Jasemi, S.; Sechi, L.A.; Feizabadi, M.M. Enterotoxigenic Bacteroides fragilis: A possible etiological candidate for bacterially-induced colorectal precancerous and cancerous lesions. Front. Cell. Infect. Microbiol. 2020, 9, 449. [Google Scholar] [CrossRef]
- Alomair, A.O.; Masoodi, I.; Alyamani, E.J.; Allehibi, A.A.; Qutub, A.N.; Alsayari, K.N.; Altammami, M.A.; Alshanqeeti, A.S. Colonic mucosal microbiota in colorectal cancer: A single-center metagenomic study in Saudi Arabia. Gastroenterol. Res. Pract. 2018, 2018, 5284754. [Google Scholar] [CrossRef]
- Kwong, T.N.Y.; Wang, X.; Nakatsu, G.; Chow, T.C.; Tipoe, T.; Dai, R.Z.W.; Tsoi, K.K.K.; Wong, M.C.S.; Tse, G.; Chan, M.T.V.; et al. Association between bacteremia from specific microbes and subsequent diagnosis of colorectal cancer. Gastroenterology 2018, 155, 383–390.e8. [Google Scholar] [CrossRef]
- Proença, M.A.; Biselli, J.M.; Succi, M.; Severino, F.E.; Berardinelli, G.N.; Caetano, A.; Reis, R.M.; Hughes, D.J.; Silva, A.E. Relationship between Fusobacterium nucleatum, inflammatory mediators and microRNAs in colorectal carcinogenesis. World J. Gastroenterol. 2018, 24, 5351–5365. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.S.; Nishihara, R.; Cao, Y.; Song, M.; Mima, K.; Qian, Z.R.; Nowak, J.A.; Kosumi, K.; Hamada, T.; Masugi, Y.; et al. Association of dietary patterns with risk of colorectal cancer subtypes classified by Fusobacterium nucleatum in tumor tissue. JAMA Oncol. 2017, 3, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Jiang, B. Analysis of mucosa-associated microbiota in colorectal cancer. Med. Sci. Monit. 2017, 23, 4422–4430. [Google Scholar] [CrossRef]
- Mima, K.; Nishihara, R.; Qian, Z.R.; Cao, Y.; Sukawa, Y.; Nowak, J.A.; Yang, J.; Dou, R.; Masugi, Y.; Song, M.; et al. Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut 2016, 65, 1973–1980. [Google Scholar] [CrossRef] [PubMed]
- Magdy, A.; Elhadidy, M.; Abd Ellatif, M.E.; El Nakeeb, A.; Abdallah, E.; Thabet, W.; Youssef, M.; Khafagy, W.; Morshed, M.; Farid, M. Enteropathogenic Escherichia coli (EPEC): Does it have a role in colorectal tumourigenesis? A Prospective Cohort Study. Int. J. Surg. 2015, 18, 169–173. [Google Scholar] [CrossRef]
- Bonnet, M.; Buc, E.; Sauvanet, P.; Darcha, C.; Dubois, D.; Pereira, B.; Déchelotte, P.; Bonnet, R.; Pezet, D.; Darfeuille-Michaud, A. Colonization of the human gut by E. coli and colorectal cancer risk. Clin. Cancer Res. 2014, 20, 859–867. [Google Scholar] [CrossRef]
- Kohoutova, D.; Smajs, D.; Moravkova, P.; Cyrany, J.; Moravkova, M.; Forstlova, M.; Cihak, M.; Rejchrt, S.; Bures, J. Escherichia coli strains of phylogenetic group B2 and D and bacteriocin production are associated with advanced colorectal neoplasia. BMC Infect. Dis. 2014, 14, 733. [Google Scholar] [CrossRef]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Brunham, R.C.; McClarty, G. Chlamydia. In Sexually Transmitted Diseases; Stanberry, L.R., Bernstein, D.I., Eds.; Academic Press: London, UK, 2000; pp. 339–367. [Google Scholar]
- Nogueira, A.T.; Braun, K.M.; Carabeo, R.A. Characterization of the growth of Chlamydia trachomatis in in vitro-generated stratified epithelium. Front. Cell. Infect. Microbiol. 2017, 7, 438. [Google Scholar] [CrossRef] [PubMed]
- Gitsels, A.; Sanders, N.; Vanrompay, D. Chlamydial infection from outside to inside. Front. Microbiol. 2019, 10, 2329. [Google Scholar] [CrossRef] [PubMed]
- Elwell, C.; Mirrashidi, K.; Engel, J. Chlamydia cell biology and pathogenesis. Nat. Rev. Microbiol. 2016, 14, 385–400. [Google Scholar] [CrossRef] [PubMed]
- Saka, H.A.; Valdivia, R.H. Acquisition of nutrients by Chlamydiae: Unique challenges of living in an intracellular compartment. Curr. Opin. Microbiol. 2010, 13, 4–10. [Google Scholar] [CrossRef]
- Byrne, G.I. Chlamydia trachomatis strains and virulence: Rethinking links to infection prevalence and disease severity. J. Infect. Dis. 2010, 201 (Suppl. 2), S126–S133. [Google Scholar] [CrossRef]
- Zhong, G. Chlamydial plasmid-dependent pathogenicity. Trends Microbiol. 2017, 25, 141–152. [Google Scholar] [CrossRef]
- Sturdevant, G.L.; Zhou, B.; Carlson, J.H.; Whitmire, W.M.; Song, L.; Caldwell, H.D. Infectivity of urogenital Chlamydia trachomatis plasmid-deficient, CT135-null, and double-deficient strains in female mice. Pathog. Dis. 2014, 71, 90–92. [Google Scholar] [CrossRef]
- Borges, V.; Pinheiro, M.; Antelo, M.; Sampaio, D.A.; Vieira, L.; Ferreira, R.; Nunes, A.; Almeida, F.; Mota, L.J.; Borrego, M.J.; et al. Chlamydia trachomatis in vivo to in vitro transition reveals mechanisms of phase variation and down-regulation of virulence factors. PLoS ONE 2015, 10, e0133420. [Google Scholar] [CrossRef]
- Clifton, D.R.; Fields, K.A.; Grieshaber, S.S.; Dooley, C.A.; Fischer, E.R.; Mead, D.J.; Carabeo, R.A.; Hackstadt, T. A chlamydial type III translocated protein is tyrosine-phosphorylated at the site of entry and associated with recruitment of actin. Proc. Natl. Acad. Sci. USA 2004, 101, 10166–10171. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Park, J.; Thomas, M.; Cruz, E.; Cardona, O.; Kang, H.; Jewett, T. Biophysical characterization of actin bundles generated by the Chlamydia trachomatis Tarp effector. Biochem. Biophys. Res. Commun. 2018, 500, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Caven, L.; Carabeo, R.A. Pathogenic puppetry: Manipulation of the host actin cytoskeleton by Chlamydia trachomatis. Int. J. Mol. Sci. 2019, 21, 90. [Google Scholar] [CrossRef] [PubMed]
- Mölleken, K.; Becker, E.; Hegemann, J.H. The Chlamydia pneumoniae invasin protein Pmp21 recruits the EGF receptor for host cell entry. PLoS Pathog. 2013, 9, e1003325. [Google Scholar] [CrossRef]
- Favaroni, A.; Trinks, A.; Weber, M.; Hegemann, J.H.; Schnee, C. Pmp repertoires influence the different infectious potential of avian and mammalian Chlamydia psittaci strains. Front. Microbiol. 2021, 12, 656209. [Google Scholar] [CrossRef]
- Niessner, A.; Kaun, C.; Zorn, G.; Speidl, W.; Türel, Z.; Christiansen, G.; Pedersen, A.S.; Birkelund, S.; Simon, S.; Georgopoulos, A.; et al. Polymorphic membrane protein (PMP) 20 and PMP 21 of Chlamydia pneumoniae induce proinflammatory mediators in human endothelial cells in vitro by activation of the nuclear factor-kappaB pathway. J. Infect. Dis. 2003, 188, 108–113. [Google Scholar] [CrossRef]
- Nairz, M.; Dichtl, S.; Schroll, A.; Haschka, D.; Tymoszuk, P.; Theurl, I.; Weiss, G. Iron and innate antimicrobial immunity-depriving the pathogen, defending the host. J. Trace Elem. Med. Biol. 2018, 48, 118–133. [Google Scholar] [CrossRef]
- Pokorzynski, N.D.; Thompson, C.C.; Carabeo, R.A. Ironing out the unconventional mechanisms of iron acquisition and gene regulation in Chlamydia. Front. Cell. Infect. Microbiol. 2017, 7, 394. [Google Scholar] [CrossRef]
- Thompson, C.C.; Carabeo, R.A. An optimal method of iron starvation of the obligate intracellular pathogen, Chlamydia trachomatis. Front. Microbiol. 2011, 2, 20. [Google Scholar] [CrossRef]
- Pokorzynski, N.D.; Brinkworth, A.J.; Carabeo, R. A bipartite iron-dependent transcriptional regulation of the tryptophan salvage pathway in Chlamydia trachomatis. eLife 2019, 8, e42295. [Google Scholar] [CrossRef]
- Pokorzynski, N.D.; Hatch, N.D.; Ouellette, S.P.; Carabeo, R.A. The iron-dependent repressor YtgR is a tryptophan-dependent attenuator of the trpRBA operon in Chlamydia trachomatis. Nat. Commun. 2020, 11, 6430. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.R.; Burton, M.J.; Haddad, D.; West, S.; Wright, H. Trachoma. Lancet 2014, 384, 2142–2152. [Google Scholar] [CrossRef] [PubMed]
- Sessa, R.; Nicoletti, M.; Di Pietro, M.; Schiavoni, G.; Santino, I.; Zagaglia, C.; Del Piano, M.; Cipriani, P. Chlamydia pneumoniae and atherosclerosis: Current state and future prospectives. Int. J. Immunopathol. Pharmacol. 2009, 22, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, L.; Liu, J.; Ma, L.; Wang, H.; Zheng, N.; Chen, X.; Shen, B.; Xu, Z.; Zhang, L. Chlamydia pneumoniae infection promotes vascular endothelial cell angiogenesis through an IQGAP1-related signaling pathway. Int. J. Med. Microbiol. 2017, 307, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ning, B.; Li, Y. Study on the relationship between Chlamydia infection and otitis media with effusion. Zhonghua Er Bi Yan Hou Ke Za Zhi (Chin. J. Otorhinolaryngol.) 1999, 34, 92–94. [Google Scholar]
- Keles, E.; Bulut, Y.; Kaygusuz, I.; Karlidag, T.; Yalçin, S.; Ozdarendeli, A.; Alpay, H.C. Identification of Chlamydia trachomatis with polymerase chain reaction in middle ear fluid in otitis media with effusion. Indian Pediatr. 2005, 42, 686–691. [Google Scholar]
- Xu, X.; Liu, Z.; Xiong, W.; Qiu, M.; Kang, S.; Xu, Q.; Cai, L.; He, F. Combined and interaction effect of Chlamydia pneumoniae infection and smoking on lung cancer: A case-control study in Southeast China. BMC Cancer 2020, 20, 903. [Google Scholar] [CrossRef]
- Chaturvedi, A.K.; Gaydos, C.A.; Agreda, P.; Holden, J.P.; Chatterjee, N.; Goedert, J.J.; Caporaso, N.E.; Engels, E.A. Chlamydia pneumoniae infection and risk for lung cancer. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1498–1505. [Google Scholar] [CrossRef]
- Liu, Z.; Su, M.; Yu, S.C.; Yin, Z.H.; Zhou, B.S. Association of Chlamydia pneumoniae immunoglobulin G antibodies with the risk of lung cancer among non-smoking women in Liaoning, China. Thorac. Cancer 2010, 1, 126–129. [Google Scholar] [CrossRef]
- Anttila, T.; Koskela, P.; Leinonen, M.; Laukkanen, P.; Hakulinen, T.; Lehtinen, M.; Pukkala, E.; Paavonen, J.; Saikku, P. Chlamydia pneumoniae infection and the risk of female early-onset lung cancer. Int. J. Cancer 2003, 107, 681–682. [Google Scholar] [CrossRef]
- Bhardwaj, M.; Sharma, A.; Sen, S.; Kumar, L.; Satpathy, G.; Kashyap, S.; Pushker, N.; Singh, V.K.; Rai, A. Chlamydia and ocular adnexal lymphomas: An Indian experience. Exp. Mol. Pathol. 2016, 101, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Aigelsreiter, A.; Gerlza, T.; Deutsch, A.J.; Leitner, E.; Beham-Schmid, C.; Beham, A.; Popper, H.; Borel, N.; Pospischil, A.; Raderer, M.; et al. Chlamydia psittaci Infection in nongastrointestinal extranodal MALT lymphomas and their precursor lesions. Am. J. Clin. Pathol. 2011, 135, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Carugi, A.; Onnis, A.; Antonicelli, G.; Rossi, B.; Mannucci, S.; Luzzi, A.; Lazzi, S.; Bellan, C.; Tosi, G.M.; Sayed, S.; et al. Geographic variation and environmental conditions as cofactors in Chlamydia psittaci association with ocular adnexal lymphomas: A comparison between Italian and African samples. Hematol. Oncol. 2010, 28, 20–26. [Google Scholar] [PubMed]
- Ferreri, A.J.; Dolcetti, R.; Dognini, G.P.; Malabarba, L.; Vicari, N.; Pasini, E.; Ponzoni, M.; Cangi, M.G.; Pecciarini, L.; Resti, A.G.; et al. Chlamydophila psittaci is viable and infectious in the conjunctiva and peripheral blood of patients with ocular adnexal lymphoma: Results of a single-center prospective case-control study. Int. J. Cancer 2008, 123, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Idahl, A.; Le Cornet, C.; González Maldonado, S.; Waterboer, T.; Bender, N.; Tjønneland, A.; Hansen, L.; Boutron-Ruault, M.C.; Fournier, A.; Kvaskoff, M.; et al. Serologic markers of Chlamydia trachomatis and other sexually transmitted infections and subsequent ovarian cancer risk: Results from the EPIC cohort. Int. J. Cancer 2020, 147, 2042–2052. [Google Scholar] [CrossRef]
- Fortner, R.T.; Terry, K.L.; Bender, N.; Brenner, N.; Hufnagel, K.; Butt, J.; Waterboer, T.; Tworoger, S.S. Sexually transmitted infections and risk of epithelial ovarian cancer: Results from the Nurses’ Health Studies. Br. J. Cancer 2019, 120, 855–860. [Google Scholar] [CrossRef]
- Trabert, B.; Waterboer, T.; Idahl, A.; Brenner, N.; Brinton, L.A.; Butt, J.; Coburn, S.B.; Hartge, P.; Hufnagel, K.; Inturrisi, F.; et al. Antibodies against Chlamydia trachomatis and ovarian cancer risk in two independent populations. J. Natl. Cancer Inst. 2019, 111, 129–136. [Google Scholar] [CrossRef]
- Ness, R.B.; Goodman, M.T.; Shen, C.; Brunham, R.C. Serologic evidence of past infection with Chlamydia trachomatis, in relation to ovarian cancer. J. Infect. Dis. 2003, 187, 1147–1152. [Google Scholar] [CrossRef]
- Jensen, K.E.; Thomsen, L.T.; Schmiedel, S.; Frederiksen, K.; Norrild, B.; van den Brule, A.; Iftner, T.; Kjær, S.K. Chlamydia trachomatis and risk of cervical intraepithelial neoplasia grade 3 or worse in women with persistent human papillomavirus infection: A cohort study. Sex. Transm. Infect. 2014, 90, 550–555. [Google Scholar] [CrossRef]
- Luostarinen, T.; Namujju, P.B.; Merikukka, M.; Dillner, J.; Hakulinen, T.; Koskela, P.; Paavonen, J.; Surcel, H.M.; Lehtinen, M. Order of HPV/Chlamydia infections and cervical high-grade precancer risk: A case-cohort study. Int. J. Cancer. 2013, 133, 1756–1759. [Google Scholar] [CrossRef]
- Olejek, A.; Kozak-Darmas, I.; Kellas-Sleczka, S.; Jarek, A.; Wiczkowski, A.; Krol, W.; Stencel-Gabriel, K. Chlamydia trachomatis infection in women with lichen sclerosus vulvae and vulvar cancer. Neuro. Endocrinol. Lett. 2009, 30, 671–674. [Google Scholar] [PubMed]
- Alghamdi, S. Microbiological profile and antibiotic vulnerability of bacterial isolates from cancer patients. Cell. Mol. Biol. 2021, 67, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Bhat, V.; Gupta, S.; Kelkar, R.; Biswas, S.; Khattry, N.; Moiyadi, A.; Bhat, P.; Ambulkar, R.; Chavan, P.; Chiplunkar, S.; et al. Bacteriological profile and antibiotic susceptibility patterns of clinical isolates in a tertiary care cancer center. Indian J. Med. Paediatr. Oncol. 2016, 37, 20–24. [Google Scholar] [CrossRef]
- Maschmeyer, G.; Braveny, I. Review of the incidence and prognosis of Pseudomonas aeruginosa infections in cancer patients in the 1990s. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 915–925. [Google Scholar] [CrossRef]
- Samonis, G.; Vardakas, K.Z.; Kofteridis, D.P.; Dimopoulou, D.; Andrianaki, A.M.; Chatzinikolaou, I.; Katsanevaki, E.; Maraki, S.; Falagas, M.E. Characteristics, risk factors and outcomes of adult cancer patients with extensively drug-resistant Pseudomonas aeruginosa infections. Infection 2014, 42, 721–728. [Google Scholar] [CrossRef]
- Paprocka, P.; Durnaś, B.; Mańkowska, A.; Król, G.; Wollny, T.; Bucki, R. Pseudomonas aeruginosa infections in cancer patients. Pathogens. 2022, 11, 679. [Google Scholar] [CrossRef]
- Bodey, G.P. Pseudomonas aeruginosa infections in cancer patients: Have they gone away? Curr. Opin. Infect. Dis. 2001, 14, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Parkins, M.D.; Somayaji, R.; Waters, V.J. Epidemiology, biology, and impact of clonal Pseudomonas aeruginosa infections in cystic fibrosis. Clin. Microbiol. Rev. 2018, 31, e00019-18. [Google Scholar] [CrossRef]
- Mellemkjaer, L.; Hammarstrom, L.; Andersen, V.; Yuen, J.; Heilmann, C.; Barington, T.; Bjorkander, J.; Olsen, J.H. Cancer risk among patients with IgA deficiency or common variable immunodeficiency and their relatives: A combined Danish and Swedish study. Clin. Exp. Immunol. 2002, 130, 495–500. [Google Scholar] [CrossRef]
- Mortaz, E.; Tabarsi, P.; Mansouri, D.; Khosravi, A.; Garssen, J.; Velayati, A.; Adcock, I.M. Cancers related to immunodeficiencies: Update and perspectives. Front. Immunol. 2016, 7, 365. [Google Scholar] [CrossRef]
- Yarchoan, R.; Uldrick, T.S. HIV-associated cancers and related diseases. N. Engl. J. Med. 2018, 378, 2145. [Google Scholar] [CrossRef] [PubMed]
- Appelt, D.; Fuchs, T.; Steinkamp, G.; Ellemunter, H. Malignancies in patients with cystic fibrosis: A case series. J. Med. Case Rep. 2022, 16, 27. [Google Scholar] [CrossRef] [PubMed]
- Maisonneuve, P.; Marshall, B.C.; Knapp, E.A.; Lowenfels, A.B. Cancer risk in cystic fibrosis: A 20-year nationwide study from the United States. J. Natl. Cancer Inst. 2013, 105, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Johannesson, M.; Askling, J.; Montgomery, S.M.; Ekbom, A.; Bahmanyar, S. Cancer risk among patients with cystic fibrosis and their first-degree relatives. Int. J. Cancer 2009, 125, 2953–2956. [Google Scholar] [CrossRef]
- Neglia, J.P.; FitzSimmons, S.C.; Maisonneuve, P.; Schöni, M.H.; Schöni-Affolter, F.; Corey, M.; Lowenfels, A.B. The risk of cancer among patients with cystic fibrosis. Cystic Fibrosis and Cancer Study Group. N. Engl. J. Med. 1995, 332, 494–499. [Google Scholar] [CrossRef]
- Scott, P.; Anderson, K.; Singhania, M.; Cormier, R. Cystic fibrosis, CFTR, and colorectal cancer. Int. J. Mol. Sci. 2020, 21, 2891. [Google Scholar] [CrossRef]
- Silva-García, O.; Valdez-Alarcón, J.J.; Baizabal-Aguirre, V.M. Wnt/β-catenin signaling as a molecular target by pathogenic bacteria. Front. Immunol. 2019, 10, 2135. [Google Scholar] [CrossRef]
- de Sousa, T.; Hébraud, M.; Dapkevicius, M.L.N.E.; Maltez, L.; Pereira, J.E.; Capita, R.; Alonso-Calleja, C.; Igrejas, G.; Poeta, P. Genomic and metabolic characteristics of the pathogenicity in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 12892. [Google Scholar] [CrossRef]
- Liao, C.; Huang, X.; Wang, Q.; Yao, D.; Lu, W. Virulence factors of Pseudomonas aeruginosa and antivirulence strategies to combat its drug resistance. Front. Cell. Infect. Microbiol. 2022, 12, 926758. [Google Scholar] [CrossRef]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Busch, W. Aus der sitzung der medicinischen section vom (from the meeting of the medical section of) 13 November 1867. Berlin Klin Wochenschr (Berl. Clin. Wkly.) 1868, 5, 137. [Google Scholar]
- Yang, X.; Kessler, E.; Su, L.J.; Thorburn, A.; Frankel, A.E.; Li, Y.; La Rosa, F.G.; Shen, J.; Li, C.Y.; Varella-Garcia, M.; et al. Diphtheria toxin-epidermal growth factor fusion protein DAB389EGF for the treatment of bladder cancer. Clin. Cancer Res. 2013, 19, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Frankel, A.E.; Powell, B.L.; Hall, P.D.; Case, L.D.; Kreitman, R.J. Phase I trial of a novel diphtheria toxin/granulocyte macrophage colony-stimulating factor fusion protein (DT388GMCSF) for refractory or relapsed acute myeloid leukemia. Clin. Cancer Res. 2002, 8, 1004–1013. [Google Scholar] [PubMed]
- Kawai, H.; Ando, K.; Maruyama, D.; Yamamoto, K.; Kiyohara, E.; Terui, Y.; Fukuhara, N.; Miyagaki, T.; Tokura, Y.; Sakata-Yanagimoto, M.; et al. Phase II study of E7777 in Japanese patients with relapsed/refractory peripheral and cutaneous T-cell lymphoma. Cancer Sci. 2021, 112, 2426–2435. [Google Scholar] [CrossRef]
- Ray, T.; Chakrabarti, M.K.; Pal, A. Hemagglutinin protease secreted by V. cholerae induced apoptosis in breast cancer cells by ROS mediated intrinsic pathway and regresses tumor growth in mice model. Apoptosis 2016, 21, 143–154. [Google Scholar] [CrossRef]
- Liu-Chittenden, Y.; Jain, M.; Kumar, P.; Patel, D.; Aufforth, R.; Neychev, V.; Sadowski, S.; Gara, S.K.; Joshi, B.H.; Cottle-Delisle, C.; et al. Phase I trial of systemic intravenous infusion of interleukin-13-Pseudomonas exotoxin in patients with metastatic adrenocortical carcinoma. Cancer Med. 2015, 4, 1060–1068. [Google Scholar] [CrossRef]
- Hassan, R.; Alewine, C.; Mian, I.; Spreafico, A.; Siu, L.L.; Gomez-Roca, C.; Delord, J.P.; Italiano, A.; Lassen, U.; Soria, J.C.; et al. Phase 1 study of the immunotoxin LMB-100 in patients with mesothelioma and other solid tumors expressing mesothelin. Cancer 2020, 126, 4936–4947. [Google Scholar] [CrossRef]
- Hotz, B.; Backer, M.V.; Backer, J.M.; Buhr, H.J.; Hotz, H.G. Specific targeting of tumor endothelial cells by a shiga-like toxin-vascular endothelial growth factor fusion protein as a novel treatment strategy for pancreatic cancer. Neoplasia 2010, 12, 797–806. [Google Scholar] [CrossRef]
- Monteillier, A.; Allard, P.M.; Gindro, K.; Wolfender, J.L.; Cuendet, M. Lung cancer chemopreventive activity of patulin isolated from Penicillium vulpinum. Molecules 2018, 23, 636. [Google Scholar] [CrossRef]
- Menon, K.; Alvarez, E.; Forler, P.; Phares, V.; Amsrud, T.; Shih, C.; Al-Awar, R.; Teicher, B.A. Antitumor activity of cryptophycins: Effect of infusion time and combination studies. Cancer Chemother. Pharmacol. 2000, 46, 142–149. [Google Scholar] [CrossRef]
- Hellmann, M.D.; Paz-Ares, L.; Bernabe Caro, R.; Zurawski, B.; Kim, S.W.; Carcereny Costa, E.; Park, K.; Alexandru, A.; Lupinacci, L.; de la Mora Jimenez, E.; et al. Nivolumab plus ipilimumab in advanced non-small-cell lung cancer. N. Engl. J. Med. 2019, 381, 2020–2031. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y.; Lee, K.H.; Lee, D.W.; Yoon, J.; Kim, T.Y.; Bang, J.H.; Nam, A.R.; Oh, K.S.; Kim, J.M.; Lee, Y.; et al. Gemcitabine and cisplatin plus durvalumab with or without tremelimumab in chemotherapy-naive patients with advanced biliary tract cancer: An open-label, single-centre, phase 2 study. Lancet Gastroenterol. Hepatol. 2022, 7, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Makker, V.; Colombo, N.; Casado Herráez, A.; Santin, A.D.; Colomba, E.; Miller, D.S.; Fujiwara, K.; Pignata, S.; Baron-Hay, S.; Ray-Coquard, I.; et al. Lenvatinib plus pembrolizumab for advanced endometrial cancer. N. Engl. J. Med. 2022, 386, 437–448. [Google Scholar] [CrossRef]
- Cho, B.C.; Abreu, D.R.; Hussein, M.; Cobo, M.; Patel, A.J.; Secen, N.; Lee, K.H.; Massuti, B.; Hiret, S.; Yang, J.C.H.; et al. Tiragolumab plus atezolizumab versus placebo plus atezolizumab as a first-line treatment for PD-L1-selected non-small-cell lung cancer (CITYSCAPE): Primary and follow-up analyses of a randomised, double-blind, phase 2 study. Lancet Oncol. 2022, 23, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Hashimi, S.M.; Grant, B.; Alqurashi, N.; Alowaidi, F.; Wei, M.Q. EGF ligand fused to truncated Pseudomonas aeruginosa exotoxin A specifically targets and inhibits EGFR-positive cancer cells. Oncol. Rep. 2018, 40, 2690–2697. [Google Scholar] [CrossRef] [PubMed]
- Twigg, M.S.; Adu, S.A.; Sugiyama, S.; Marchant, R.; Banat, I.M. Mono-rhamnolipid biosurfactants synthesized by Pseudomonas aeruginosa detrimentally affect colorectal cancer cells. Pharmaceutics 2022, 14, 2799. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.K.; Naffouje, S.A.; Goto, M.; Wang, J.; Christov, K.; Rademacher, D.J.; Green, A.; Stecenko, A.A.; Chakrabarty, A.M.; Das Gupta, T.K.; et al. Cross-talk between cancer and Pseudomonas aeruginosa mediates tumor suppression. Commun. Biol. 2023, 6, 16. [Google Scholar] [CrossRef]
- Pang, Z.; Gu, M.D.; Tang, T. Pseudomonas aeruginosa in cancer therapy: Current knowledge, challenges and future perspectives. Front. Oncol. 2022, 12, 891187. [Google Scholar] [CrossRef]
- Ephraim, R.; Fraser, S.; Nurgali, K.; Apostolopoulos, V. Checkpoint markers and tumor microenvironment: What do we know? Cancers 2022, 14, 3788. [Google Scholar] [CrossRef]
- Gomatou, G.; Syrigos, N.; Vathiotis, I.A.; Kotteas, E.A. Tumor dormancy: Implications for invasion and metastasis. Int. J. Mol. Sci. 2021, 22, 4862. [Google Scholar] [CrossRef]
- Dropulic, L.K.; Lederman, H.M. Overview of infections in the immunocompromised host. Microbiol. Spectr. 2016, 4, 1–50. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Liu, Z.; Li, B.; Liu, Y.E.; Wang, P. Immunoregulation in cancer-associated cachexia. J. Adv. Res. 2023. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Mi, L.; Wang, Y.; Zhang, D. Detection of bacterial DNA in serum from colon cancer patients: Association with cytokine levels and cachexia. J. Cancer Ther. Res. 2012, 1, 19. [Google Scholar] [CrossRef][Green Version]
- Herremans, K.M.; Riner, A.N.; Cameron, M.E.; Trevino, J.G. The microbiota and cancer cachexia. Int. J. Mol. Sci. 2019, 20, 6267. [Google Scholar] [CrossRef] [PubMed]
- Nicol, A.F.; Andrade, C.V.; Russomano, F.B.; Rodrigues, L.L.; Oliveira, N.S.; Provance, D.W., Jr. HPV vaccines: A controversial issue? Braz. J. Med. Biol. Res. 2016, 49, e5060. [Google Scholar] [CrossRef] [PubMed]
- Elwood, J.M.; Ameratunga, R. Autoimmune diseases after hepatitis B immunization in adults: Literature review and meta-analysis, with reference to ‘autoimmune/autoinflammatory syndrome induced by adjuvants’ (ASIA). Vaccine 2018, 36, 5796–5802. [Google Scholar] [CrossRef]
- Gidengil, C.; Goetz, M.B.; Newberry, S.; Maglione, M.; Hall, O.; Larkin, J.; Motala, A.; Hempel, S. Safety of vaccines used for routine immunization in the United States: An updated systematic review and meta-analysis. Vaccine 2021, 39, 3696–3716. [Google Scholar] [CrossRef]
- Eun, B.W.; Bahar, E.; Xavier, S.; Kim, H.; Borys, D. Post-marketing surveillance study of the safety of the HPV-16/18 vaccine in Korea (2017–2021). Hum. Vaccin. Immunother. 2023, 19, 2184756. [Google Scholar] [CrossRef]
- Seida, I.; Seida, R.; Elsalti, A.; Mahroum, N. Vaccines and autoimmunity-from side effects to ASIA syndrome. Medicina 2023, 59, 364. [Google Scholar] [CrossRef]
- Shuel, S.L. Targeted cancer therapies: Clinical pearls for primary care. Can. Fam. Physician 2022, 68, 515–518. [Google Scholar] [CrossRef]
- Ray, A. Tumor-linked HER2 expression: Association with obesity and lipid-related microenvironment. Horm. Mol. Biol. Clin. Investig. 2017, 32, 20170020. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.; Finikarides, L.; Freeman, A.L.J. The adverse effects of trastuzumab-containing regimes as a therapy in breast cancer: A piggy-back systematic review and meta-analysis. PLoS ONE 2022, 17, e0275321. [Google Scholar] [CrossRef] [PubMed]
- Martins, F.; Sofiya, L.; Sykiotis, G.P.; Lamine, F.; Maillard, M.; Fraga, M.; Shabafrouz, K.; Ribi, C.; Cairoli, A.; Guex-Crosier, Y.; et al. Adverse effects of immune-checkpoint inhibitors: Epidemiology, management and surveillance. Nat. Rev. Clin. Oncol. 2019, 16, 563–580. [Google Scholar] [CrossRef] [PubMed]
- Babacan, N.A.; Tanvetyanon, T. Superimposed Clostridium difficile infection during checkpoint inhibitor immunotherapy-induced colitis. J. Immunother. 2019, 42, 350–353. [Google Scholar] [CrossRef]
- Morelli, T.; Fujita, K.; Redelman-Sidi, G.; Elkington, P.T. Infections due to dysregulated immunity: An emerging complication of cancer immunotherapy. Thorax 2022, 77, 304–311. [Google Scholar] [CrossRef]
- Rolston, K.V. Infections in cancer patients with solid tumors: A review. Infect. Dis. Ther. 2017, 6, 69–83. [Google Scholar] [CrossRef]
- Gudiol, C.; Carratalà, J. Antibiotic resistance in cancer patients. Expert Rev. Anti Infect. Ther. 2014, 12, 1003–1016. [Google Scholar] [CrossRef]
- Wisplinghoff, H.; Seifert, H.; Wenzel, R.P.; Edmond, M.B. Current trends in the epidemiology of nosocomial bloodstream infections in patients with hematological malignancies and solid neoplasms in hospitals in the United States. Clin. Infect. Dis. 2003, 36, 1103–1110. [Google Scholar] [CrossRef]
- Rolston, K.V. New antimicrobial agents for the treatment of bacterial infections in cancer patients. Hematol. Oncol. 2009, 27, 107–114. [Google Scholar] [CrossRef]
- Itoh, N.; Akazawa, N.; Kanawaku, E.; Murakami, H.; Ishibana, Y.; Kawamura, D.; Kawabata, T.; Mori, K.; Kodama, E.N.; Ohmagari, N. Effects of infectious disease consultation and antimicrobial stewardship program at a Japanese cancer center: An interrupted time-series analysis. PLoS ONE 2022, 17, e0263095. [Google Scholar] [CrossRef]
- Molina, J.; Noguer, M.; Lepe, J.A.; Pérez-Moreno, M.A.; Aguilar-Guisado, M.; Lasso de la Vega, R.; Peñalva, G.; Crespo-Rivas, J.C.; Gil-Navarro, M.V.; Salvador, J.; et al. Clinical impact of an educational antimicrobial stewardship program associated with infectious diseases consultation targeting patients with cancer: Results of a 9-year quasi-experimental study with an interrupted time-series analysis. J. Infect. 2019, 79, 206–211. [Google Scholar] [CrossRef]
- Rosa, R.G.; Goldani, L.Z.; dos Santos, R.P. Association between adherence to an antimicrobial stewardship program and mortality among hospitalised cancer patients with febrile neutropaenia: A prospective cohort study. BMC Infect. Dis. 2014, 14, 286. [Google Scholar] [CrossRef] [PubMed]
- Rolfes, N.; Lümmen, G. Management of febrile neutropenia. Urologe A 2011, 50, 810–812. [Google Scholar] [CrossRef] [PubMed]
- Nouér, S.A.; Nucci, M.; Anaissie, E. Tackling antibiotic resistance in febrile neutropenia: Current challenges with and recommendations for managing infections with resistant Gram-negative organisms. Expert Rev. Hematol. 2015, 8, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Feld, R. Bloodstream infections in cancer patients with febrile neutropenia. Int. J. Antimicrob. Agents 2008, 32 (Suppl. 1), S30–S33. [Google Scholar] [CrossRef]
- Kouranos, V.; Dimopoulos, G.; Vassias, A.; Syrigos, K.N. Chemotherapy-induced neutropenia in lung cancer patients: The role of antibiotic prophylaxis. Cancer Lett. 2011, 313, 9–14. [Google Scholar] [CrossRef]
- Egan, G.; Robinson, P.D.; Martinez, J.P.D.; Alexander, S.; Ammann, R.A.; Dupuis, L.L.; Fisher, B.T.; Lehrnbecher, T.; Phillips, B.; Cabral, S.; et al. Efficacy of antibiotic prophylaxis in patients with cancer and hematopoietic stem cell transplantation recipients: A systematic review of randomized trials. Cancer Med. 2019, 8, 4536–4546. [Google Scholar] [CrossRef]
- Pascoe, J.; Steven, N. Antibiotics for the prevention of febrile neutropenia. Curr. Opin. Hematol. 2009, 16, 48–52. [Google Scholar] [CrossRef]
- Itoh, N.; Hadano, Y.; Saito, S.; Myokai, M.; Nakamura, Y.; Kurai, H. Intravenous to oral switch therapy in cancer patients with catheter-related bloodstream infection due to methicillin-sensitive Staphylococcus aureus: A single-center retrospective observational study. PLoS ONE 2018, 13, e0207413. [Google Scholar] [CrossRef]
- Coussement, J.; Lindsay, J.; Teh, B.W.; Slavin, M. Choice and duration of antifungal prophylaxis and treatment in high-risk haematology patients. Curr. Opin. Infect. Dis. 2021, 34, 297–306. [Google Scholar] [CrossRef]
- Stemler, J.; de Jonge, N.; Skoetz, N.; Sinkó, J.; Brüggemann, R.J.; Busca, A.; Ben-Ami, R.; Ráčil, Z.; Piechotta, V.; Lewis, R.; et al. Antifungal prophylaxis in adult patients with acute myeloid leukaemia treated with novel targeted therapies: A systematic review and expert consensus recommendation from the European Hematology Association. Lancet Haematol. 2022, 9, e361–e373. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.V.; Ullmann, A.J.; Almyroudis, N.G.; Segal, B.H. Broad-spectrum antifungal prophylaxis in patients with cancer at high risk for invasive mold infections: Point. J. Natl. Compr. Canc. Netw. 2008, 6, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Pathakumari, B.; Liang, G.; Liu, W. Immune defence to invasive fungal infections: A comprehensive review. Biomed. Pharmacother. 2020, 130, 110550. [Google Scholar] [CrossRef] [PubMed]
- Robenshtok, E.; Gafter-Gvili, A.; Goldberg, E.; Weinberger, M.; Yeshurun, M.; Leibovici, L.; Paul, M. Antifungal prophylaxis in cancer patients after chemotherapy or hematopoietic stem-cell transplantation: Systematic review and meta-analysis. J. Clin. Oncol. 2007, 25, 5471–5489. [Google Scholar] [CrossRef] [PubMed]
- Teh, B.W.; Yeoh, D.K.; Haeusler, G.M.; Yannakou, C.K.; Fleming, S.; Lindsay, J.; Slavin, M.A.; Australasian Antifungal Guidelines Steering Committee. Consensus guidelines for antifungal prophylaxis in haematological malignancy and haemopoietic stem cell transplantation, 2021. Intern. Med. J. 2021, 51 (Suppl. 7), 67–88. [Google Scholar] [CrossRef]
- Aitken, S.L.; Nagel, J.L.; Abbo, L.; Alegria, W.; Barreto, J.N.; Dadwal, S.; Freifeld, A.G.; Jain, R.; Pergam, S.A.; Tverdek, F.P.; et al. Antimicrobial stewardship in patients with cancer: The time is now. J. Natl. Compr. Canc. Netw. 2019, 17, 772–775. [Google Scholar] [CrossRef]
- Barlam, T.F.; Cosgrove, S.E.; Abbo, L.M.; MacDougall, C.; Schuetz, A.N.; Septimus, E.J.; Srinivasan, A.; Dellit, T.H.; Falck-Ytter, Y.T.; Fishman, N.O.; et al. Implementing an antibiotic stewardship program: Guidelines by the Infectious Diseases Society of America and the Society for Healthcare Epidemiology of America. Clin. Infect. Dis. 2016, 62, e51–e77. [Google Scholar] [CrossRef]



{kind=link}
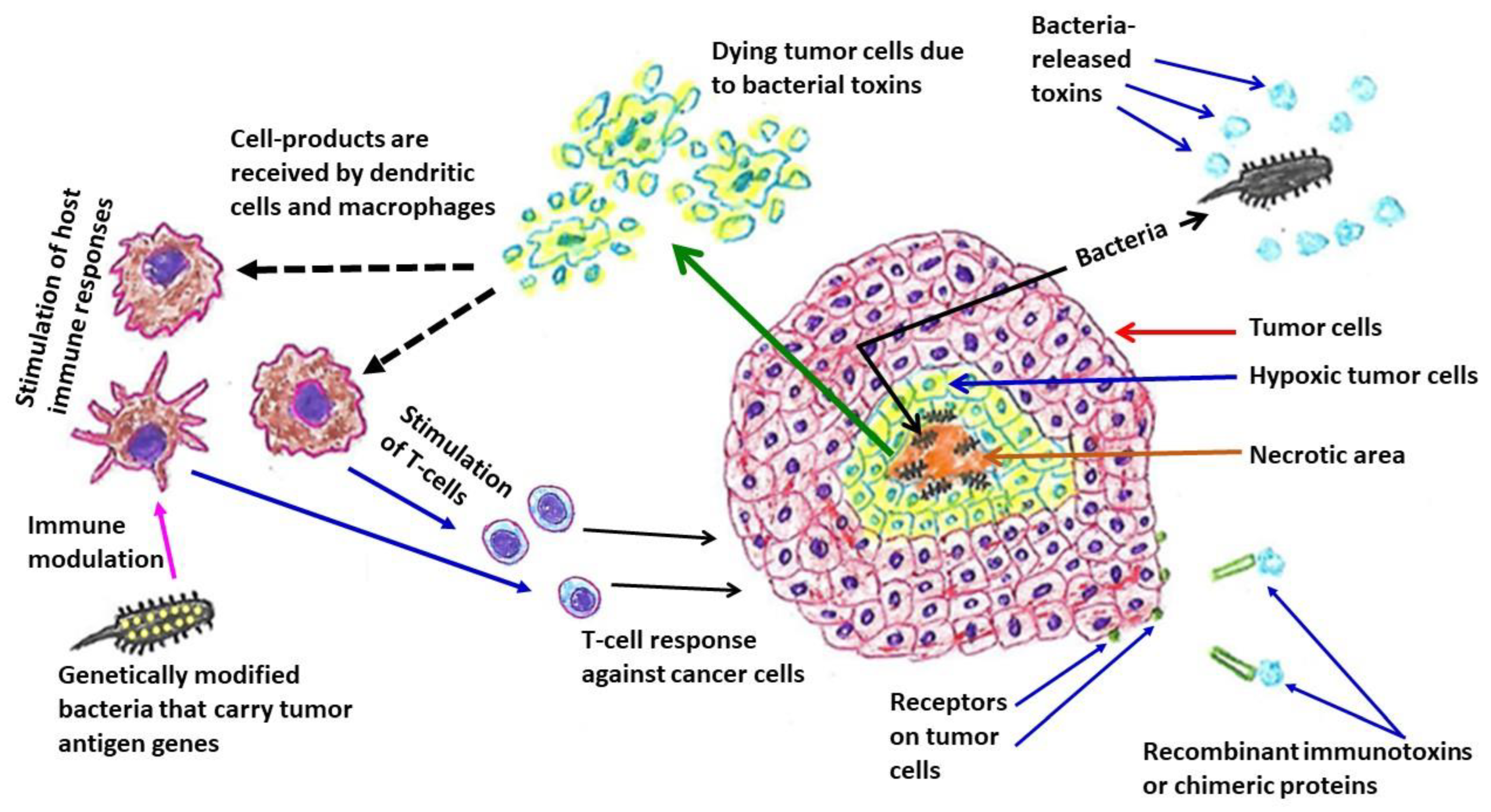{kind=link}
| Treatment Course | Patients | Medication | Duration |
|---|---|---|---|
| First-line eradication therapy: triple therapy | General/initial cases | Proton pump inhibitor (40 mg twice a day) + clarithromycin (500 mg twice a day) + amoxicillin (1 g twice a day) | 14 days |
| For patients with hypersensitivity to amoxicillin | Proton pump inhibitor (40 mg twice a day) + clarithromycin (500 mg twice a day) + metronidazole (500 mg three times a day) | ||
| Second-line eradication therapy: quadruple therapy | For patients with history of previous eradication therapy or unsuccessful triple therapy | Proton pump inhibitor (40 mg twice a day) + bismuth (300 mg four times a day) + tetracycline (500 mg four times a day) + metronidazole (500 mg three times a day) | 14 days |
| Third-line eradication therapy: triple therapy (modified) | If the above regimens do not work | Proton pump inhibitor (40 mg twice a day) + amoxicillin (1 g twice a day) + levofloxacin (500 mg once a day) | 10 days |
| Investigators, Place of Study, and Study Design | Findings |
|---|---|
| Mirzarazi et al., 2022 [61], Iran Fecal samples from 20 patients and 50 control subjects | Increased expression of outer membrane protein A (OmpA) in the commensal E. coli B2 phylogenetic group from patients |
| Périchon et al., 2022 [62], France Fecal samples collected from patients with colorectal cancer at different stages (n = 81), adenoma (n = 23), and from normal colonoscopy (n = 25) | Significantly increased levels of S. gallolyticus, F. nucleatum, and P. micra in the colorectal cancer group. Increase of pks island at late-stage cancer. |
| Xie et al., 2022 [63], China Tumor and adjacent noncancerous tissues were collected from 184 colorectal cancer patients | Cancerous tissues had higher levels of F. nucleatum than adjacent noncancerous tissues. Disease-free survival was significantly longer in the F. nucleatum-negative, younger age, and TNM stage I–II groups. |
| Bertocchi et al., 2021 [64], Italy Cohort of 179 patients—10 years follow-up | E. coli disrupted the gut vascular barrier, which supported distant metastases. |
| Butt et al., 2021 [65], Europe Nested case–control study—pre-diagnostic serum samples from 442 cases and 442 controls | Sero-positivity to E. coli and ETBF (co-infection) was associated with colorectal cancer development. |
| Cuellar-Gómez et al., 2021 [66], Mexico Tumor, peri-tumoral, and normal tissue samples from 30 patients with colorectal cancer | F. nucleatum in cancer samples was significantly higher than in the normal mucosa samples (more than 10 cm from the cancer margins). |
| Khodaverdi et al., 2021 [67], Iran Colon tissue samples from 40 colorectal cancer patients and 40 healthy controls | E. faecalis and ETBF were significantly higher in colorectal cancer samples compared to control group. |
| Kong et al., 2021 [68], China Healthy controls (n = 30) and 30 pre-operative colorectal cancer serum/fecal samples, and also 18 fresh-frozen colonic tissues from controls along with 141 fresh-frozen tumor samples from patients were analyzed. | Among patients, a positive association between higher levels of fecal F. nucleatum and serum 12,13-EpOME; and increased levels of CYP2J2 in tumor samples also correlated with high F. nucleatum levels and worse overall survival in advanced disease. |
| Nardelli et al., 2021 [69], Italy Colon tissue/mucosa from 20 patients and 20 healthy controls | E. coli, F. nucleatum, B. fragilis, S. intermedius, and G. haemolysans were increased in patients’ tumor tissue and non-lesioned tissue. |
| Pignatelli et al., 2021 [70], Italy Samples from oral, cancer tissue, and adjacent non-neoplastic mucosa from 36 patients with colon cancer | There was an association between the F. nucleatum quantity in the oral and cancer tissue, and a significant association between the F. nucleatum abundance in cancer and staging. |
| Iyadorai et al., 2020 [71], Malaysia Mucosal colon tissues from 48 patients and 23 healthy controls | pks+ E. coli was more often isolated from tissues of patients. |
| Zamani et al., 2020 [72], Iran Mucosa biopsies from 68 patients with precancerous and cancerous lesions and 52 healthy controls | B. fragilis was more abundant in the patients’ samples than healthy controls. The bft gene was detected in 47% and 3.8% of the patients and controls, respectively. |
| Alomair et al., 2018 [73], Saudi Arabia Mucosal microbial samples were collected from 29 colorectal cancer patients along with matched controls, and DNA samples were used in the study | Fusobacterium and B. fragilis were more common in the carcinoma group. |
| Kwong et al., 2018 [74], Hong Kong Retrospective population-based cohort study during a 10-year period—13,096 adult patients with bacteremia | Increased risk of colorectal cancer in patients with bacteremia from intestinal microbes such as B. fragilis, S. gallolyticus, F. nucleatum, Peptostreptococcus spp, and C. septicum. |
| Proença et al., 2018 [75], Brazil DNA and/or RNA extracted from tumor and adjacent normal tissues from 27 colorectal adenoma and 43 colorectal cancer cases | Compared to matched normal tissue, excess F. nucleatum was detected in adenoma and more markedly in cancerous tissue. Moreover, KRAS mutations were more commonly seen in cancer samples infected with F. nucleatum. |
| Mehta et al., 2017 [76], USA Prospective cohort study—using data from the Nurses’ Health Study and the Health Professionals Follow-up Study; 1019 incident colorectal cancer cases over 26–32 years follow-up | Prudent diets rich in whole grains and dietary fiber were associated with a lower risk for F. nucleatum-positive cancer. |
| Xu and Jiang 2017 [77], China Mucosa biopsy from 52 cases with invasive adenocarcinomas, 47 cases with colorectal adenomas, and 61 non-tumor colon (normal group) | Fusobacterium and B. fragilis were more common in the colorectal cancer group. |
| Mima et al., 2016 [78], USA The Nurses’ Health Study and the Health Professionals Follow-up Study—1069 colorectal carcinoma cases | F. nucleatum DNA amount in cancer tissue was associated with shorter survival. |
| Magdy et al., 2015 [79], Egypt Prospectively enrolled 213 patients and 248 healthy controls | EPEC was significantly higher in patients. Patients’ E. coli differed serotypically and genotypically from E. coli in normal persons. |
| Bonnet et al., 2014 [80], France Tumors and mucosa of patients (n = 50) and the healthy mucosa of diverticulosis (controls, n = 33) | Compared to normal tissue, a higher level of mucosa-associated and internalized E. coli in tumors, and an association between poor prognostic factors (TNM stage) and E. coli colonization in mucosa were observed. |
| Kohoutova et al., 2014 [81], Czech Republic Mucosal biopsies from colorectal adenoma (n = 30), colorectal cancer (n = 30), and healthy controls (n = 20) | Higher production of colicins and microcins, and a higher frequency of E. coli phylogroup D, were found among patients with colorectal cancer. |
| Kostic et al., 2013 [82], USA Fecal samples from healthy subjects (n = 30), colonic adenomas (n = 29), and colorectal cancer (n = 27) | Fusobacterium spp. were enriched in stool samples from colonic adenoma and carcinoma patients compared to healthy subjects. |
| Rubinstein et al., 2013 [83], USA Analysis of fadA gene (F. nucleatum virulence factor) in colon specimens from normal individuals (n = 14), normal tissues from patients with precancerous adenomas (n = 16), precancerous adenomas (n = 16), normal tissues from patients with adenocarcinomas (n = 19), and adenocarcinomas (n = 19) | The fadA gene levels in the colon tissue from patients with adenomas and cancer were higher compared with normal subjects. |
| Signaling Pathways or Relevant Biomolecules | Short Description |
|---|---|
| Akt or protein kinase B | A serine/threonine kinase, Akt plays an important role in cell cycle progression, cellular growth and survival, inhibition of apoptosis, and angiogenesis. The Akt signaling pathway is also associated with a number of biological molecules/processes, e.g., cytokine/growth factor receptors, integrins, PI3K, and immune mechanisms. |
| Cystic fibrosis transmembrane conductance regulator (CFTR) | A cyclic AMP-dependent chloride channel protein, CFTR may have a negative association with the NF-κB signaling pathway and thus cancer cell proliferation. |
| Epidermal growth factor receptor (EGFR) | A transmembrane glycoprotein receptor with tyrosine kinase activity, EGFR is involved in cell proliferation and survival. Over-expressions of EGFR and its family partner HER2 are noticed in several cancer types. |
| Extracellular signal-regulated kinase (ERK) | A member of MAP kinase family, ERK is activated through a sequential phosphorylation pathway, and associated with cell division and growth. |
| Janus kinase (JAK) | A non-receptor tyrosine kinase, JAK is linked with cytokine signaling (including IL-6 and IFN-γ), phosphorylation of the receptors, and activation of other signaling molecules such as STAT. In the development and progression of several cancers, JAK plays a significant role. |
| Mitogen-activated protein kinase (MAP kinase) | Three major groups of these serine–threonine kinases are ERK, c-Jun amino-terminal kinase (JNK), and p38 MAP kinase. They regulate various cellular activities such as cell cycle progression, cell proliferation, survival and death. |
| Nuclear factor kappa B (NF-κB) | A transcription factor, NF-κB participates in a wide range of pathophysiological phenomena, e.g., inflammation, cell proliferation and survival, immune responses, and a number of health disorders including cancer. Moreover, NF-κB is influenced by various stimuli such as IL-1β, IFN-γ, and TNFα. |
| Phosphatidylinositol 3-kinase (PI3K) | A cell-membrane-associated lipid kinase, PI3K can work with a number of signaling molecules such as Akt for cell growth and survival. The PI3K signaling is connected with several growth factor receptors such as EGFR and PDGFR, and pro-inflammatory cytokines such as IL-1β, IL-6, and TNFα. |
| Platelet-derived growth factor receptor (PDGFR) | A receptor tyrosine kinase, PDGFR is associated with the growth of mesenchymal cells. Its abnormal signaling is connected with a number of diseases, e.g., inflammation, atherosclerosis, pulmonary fibrosis, and cancer. |
| Signal transducer and activator of transcription (STAT) | A transcription factor that is connected with JAK and signal transduction by various growth factors/cytokines and hormones, e.g., IL-6, IFN-γ, and growth hormone, in order to control cellular processes such as cell proliferation, survival, and differentiation. |
| Transforming growth factor β (TGF-β) | A pleiotropic cytokine, TGF-β functions in the regulation of cell growth in both positive and negative manner. Different signaling pathways, e.g., PI3K, Akt, and MAP kinase, are associated with TGF-β―mediated functions. |
| Tumor suppressor p53 | A transcription factor, p53 is activated by stress signals and its binding with DNA initiates the transcription of genes that are linked to various cancer-preventive cellular phenomena such as cell cycle arrest and cell death. |
| Wnt/β-catenin | This pathway plays an important role in cell proliferation, apoptosis, and overall cellular homeostasis. Furthermore, it integrates other signaling pathways, including TGF-β. The Wnt pathway has two major branches: (i) β-catenin-dependent or canonical and (ii) β-catenin-independent or non-canonical. |
| Investigators, Cancer Site, and Place of Study | Study Design | Results in Brief |
|---|---|---|
| Xu et al., 2020 [109] (lung cancer), China | Case-control study, 2006–2016, 449 lung cancer cases and 512 healthy controls. | C. pneumoniae specific IgG+ or IgA+ was significantly associated with the increased risk of lung cancer among smokers, persons exposed to passive smoking, and alcohol users. |
| Chaturvedi et al., 2010 [110] (lung cancer), USA | Case-control study, 593 lung cancers and 671 matched controls. | Individuals with seropositivity for chlamydia heat shock protein-60 (IgG antibodies) had significantly increased lung cancer risk. |
| Liu et al., 2010 [111] (lung cancer), China | Case–control study, 2000–2009, 192 non-smoking women with lung cancer and 90 healthy controls. | 61.98% of cases and 28.89% of controls were positive for C. pneumoniae IgG. |
| Anttila et al., 2003 [112] (lung cancer), Finland | From the Finnish Maternity Cohort, 58 lung cancer cases with pre-diagnostic serum samples, along with 287 control women. | C. pneumoniae-specific immune complexes and both IgG and IgA antibodies were associated with female lung cancer. |
| Bhardwaj et al., 2016 [113] (lymphoma), India | 41 ocular adnexal lymphoma cases were analyzed prospectively. | C. trachomatis genome was detected in 7.3% cases. |
| Aigelsreiter et al., 2011 [114] (lymphoma), Austria | Samples from 47 non-G.I. and 14 G.I. MALT lymphomas, 37 nonmalignant controls, and 27 autoimmune precursor lesions were analyzed for the presence of C. psittaci, C. pneumoniae, and C. trachomatis DNA. | 13 (28%) non-G.I. MALT lymphomas, 1 (7%) G.I. MALT lymphoma, 4 (11%) nonmalignant control samples, and 11 (41%) autoimmune precursor lesions were positive for C. psittaci DNA. No specimens displayed positive results for C. trachomatis and C. pneumoniae. |
| Carugi et al., 2010 [115] (lymphoma), Italy | Ocular adnexal lymphoma cases from Italy (n = 30) and Kenya (n = 9). | 17% of the samples (from Italy) were positive for C. psittaci. |
| Ferreri et al., 2008 [116] (lymphoma), Italy | 20 ocular adnexal MALT lymphoma patients. | C. psittaci was detected in lymphoma tissue of 75% patients. |
| Idahl et al., 2020 [117] (ovarian cancer), Europe | Nested case-control study within the European Prospective Investigation into Cancer and Nutrition (EPIC) cohort, 791 cases and 1669 matched controls. | C. trachomatis Pgp3 seropositivity was associated with higher risk of mucinous ovarian carcinoma; chlamydia heat shock protein-60 seropositivity was associated with higher risk of epithelial ovarian cancer (overall) and with the serous subtype. |
| Fortner et al., 2019 [118] (ovarian cancer), USA | Nested case-control study in the Nurses’ Health Studies (NHS), 337 cases and 337 matched controls. | C. trachomatis seropositivity was associated with a two-fold higher risk of ovarian cancer |
| Trabert et al., 2019 [119] (ovarian cancer), Poland and USA | (i) Polish Ovarian Cancer Study (population-based case–control study conducted in Poland): 244 ovarian cancers and 556 controls. (ii) Prostate, Lung, Colorectal, and Ovarian (PLCO) Cancer Screening Trial (prospective nested case–control study): 160 ovarian cancers and 159 controls. | Antibodies against C. trachomatis (Pgp3) were associated with an increased risk of ovarian cancer in these two independent populations. |
| Ness et al., 2003 [120] (ovarian cancer), USA | Population-based case-control study conducted in Hawaii (1993–1999), 117 ovarian cancer cases and 171 matched controls. | Risk of ovarian cancer was greater in women with higher levels of chlamydia-EB antibodies. |
| Jensen et al., 2014 [121] (cervical cancer), Denmark | Population-based cohort study; women with high-risk HPV infection and without cervical disease (n = 1390) were followed. | Repeated C. trachomatis infections increased the risk of cervical intraepithelial neoplasia grade 3 or worse (CIN3+) in women with prevalent and persistent high-risk HPV infection. |
| Luostarinen et al., 2013 [122] (cervical cancer), Finland | Cohort consisted of 94,349 women at risk (1995–2003). | Concomitant HPV18/45 and C. trachomatis infections were associated with very high risk of cervical intraepithelial neoplasia grade 3 (CIN3). |
| Olejek et al., 2009 [123] (vulvar cancer), Poland | 80 women: 30 vulvar cancer and 50 lichen sclerosus vulvae (risk factor for vulvar cancer). | C. trachomatis infection could cause vulvar malignancy. |
| Investigators and Studied Toxins | Results (in Brief) |
|---|---|
| Yang et al. (2013) [144]. Diphtheria toxin-EGF fusion protein (in vitro and in vivo studies) | Fusion protein DAB389EGF treatment inhibited the growth in all tested human bladder cancer cells (J82, RT4, CRL1749, T24, TCCSUP, and HTB9). The intravesical administration of DAB389EGF in female athymic nude mice (nu/nu) with orthotopic xenograft (HTB9) bladder cancer showed a high rate of tumor clearance after 2 weeks of treatment. |
| Frankel et al. (2002) [145]. USA Diphtheria toxin-GMCSF fusion protein DT388GMCSF (phase I study) | 31 patients with acute myeloid leukemia (who were resistant to chemotherapy), 1 had a complete remission and 2 had partial remissions. The main adverse effect was liver injury, and the maximal tolerated dose was 4 μg/kg/day. |
| Kawai et al. (2021) [146]. Japan Diphtheria toxin fragments A and B and human IL-2 fusion protein E7777 (phase II study) | Patients with relapsed/refractory peripheral T-cell lymphoma (n = 17) and cutaneous T-cell lymphoma (n = 19). Objective response rate was 36%, and the median progression-free survival was 3.1 months. Adverse events: increased AST/ALT, hypoalbuminemia, lymphopenia, and pyrexia. |
| Ray et al. (2016) [147]. Vibrio cholera secreted hemagglutinin protease (in vivo study) | Weekly administration of 1 µg of hemagglutinin protease exhibited potent antitumor activity when injected into Ehrlich ascites carcinoma tumors (murine mammary adenocarcinoma) in Swiss albino mice, and improved the survival rate. |
| Liu-Chittenden et al. (2015) [148]. USA IL-13-PE: recombinant IL-13 and Pseudomonas exotoxin A (phase I study) | 8 patients with adrenocortical carcinoma and distant metastases. Treatment response: 1 patient had stable disease for 5.5 months before disease progression; the others progressed within 1–2 months. Adverse events: low grade anemia, proteinuria, fatigue, and increase in ALT, AST, and creatinine, and development of neutralizing antibodies. |
| Hassan et al. (2020) [149]. Multicenter-International study LMB-100: anti-mesothelin Fab linked to Pseudomonas exotoxin A (phase I study) | Out of 20 patients, there were no objective partial or complete responses. However, LMB-100 demonstrated an acceptable safety profile. |
| Hotz et al. (2010) [150]. SLT-VEGF: Shiga-like toxin-VEGF fusion protein (in vivo study) | AsPC-1 and HPAF-2 xenograft pancreatic tumors in nude mice were reduced in the SLT-VEGF group and SLT-VEGF plus gemcitabine combination group. |
| Monteillier et al. (2018) [151]. Patulin: a mycotoxin from Penicillium vulpinum fungus (in vitro study) | In A549 lung cancer cell line, patulin triggered apoptosis, prevented cell migration, and inhibited the Wnt pathway. |
| Menon et al. (2000) [152]. Cryptophycins: potent anti-microtubule agents isolated from cyanobacteria (in vivo study) | Cryptophycin-52 and cryptophycin-55 combined with doxorubicin, paclitaxel and 5-fluorouracil constitute highly effective regimens against the xenograft MX-1 breast carcinoma in rats. Cryptophycin-52 and cryptophycin-55 were also highly effective against the Calu-6 non-small cell lung carcinoma, H82 and SW-2 small cell lung carcinoma xenografts when combined with the antitumor platinum complexes―cisplatin, carboplatin or oxaliplatin. |
| Hellmann et al. (2019) [153]. CheckMate-227―global randomized phase III trial. Immune checkpoint inhibitors―nivolumab and ipilimumab | Treatment with nivolumab plus ipilimumab resulted in a longer duration of overall survival in patients with non-small-cell lung cancer compared to the chemotherapy group. |
| Oh et al. (2022) [154]. South Korea Open-label, single-center, phase II study. Immune checkpoint inhibitors―durvalumab and tremelimumab | 124 patients with advanced biliary tract cancer. Gemcitabine and cisplatin plus immune checkpoint inhibitors showed promising efficacy and acceptable safety with regard to adverse events. |
| Makker et al. (2022) [155]. Multicenter, open-label, randomized, phase III trial. Immune checkpoint inhibitor―pembrolizumab | 827 patients with advanced endometrial cancer. Lenvatinib plus pembrolizumab led to significantly longer progression-free survival and overall survival than the chemotherapy group. |
| Cho et al. (2022) [156]. Global phase II, randomized and blinded study. Immune checkpoint inhibitors―atezolizumab and tiragolumab | 135 patients with non-small-cell lung cancer. Tiragolumab plus atezolizumab exhibited a substantial improvement in objective response rate and progression-free survival compared to the placebo plus atezolizumab group. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ray, A.; Moore, T.F.; Pandit, R.; Burke, A.D.; Borsch, D.M. An Overview of Selected Bacterial Infections in Cancer, Their Virulence Factors, and Some Aspects of Infection Management. Biology 2023, 12, 963. https://doi.org/10.3390/biology12070963
Ray A, Moore TF, Pandit R, Burke AD, Borsch DM. An Overview of Selected Bacterial Infections in Cancer, Their Virulence Factors, and Some Aspects of Infection Management. Biology. 2023; 12(7):963. https://doi.org/10.3390/biology12070963
Chicago/Turabian StyleRay, Amitabha, Thomas F. Moore, Rajashree Pandit, Adam D. Burke, and Daniel M. Borsch. 2023. "An Overview of Selected Bacterial Infections in Cancer, Their Virulence Factors, and Some Aspects of Infection Management" Biology 12, no. 7: 963. https://doi.org/10.3390/biology12070963
APA StyleRay, A., Moore, T. F., Pandit, R., Burke, A. D., & Borsch, D. M. (2023). An Overview of Selected Bacterial Infections in Cancer, Their Virulence Factors, and Some Aspects of Infection Management. Biology, 12(7), 963. https://doi.org/10.3390/biology12070963