
Brain-Derived Estrogen Regulates Neurogenesis, Learning and Memory with Aging in Female Rats
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Antibodies
2.2. Experimental Design
2.3. Immunofluorescence Staining and Confocal Microscopy
2.4. Cell Quantifications and Morphometric Measurement
2.5. Preparation of Hippocampal Samples and Western Blot Analysis
2.6. Quantitative RT-PCR Analysis
2.7. Behavioral Assessments
2.8. Statistical Analysis
3. Results
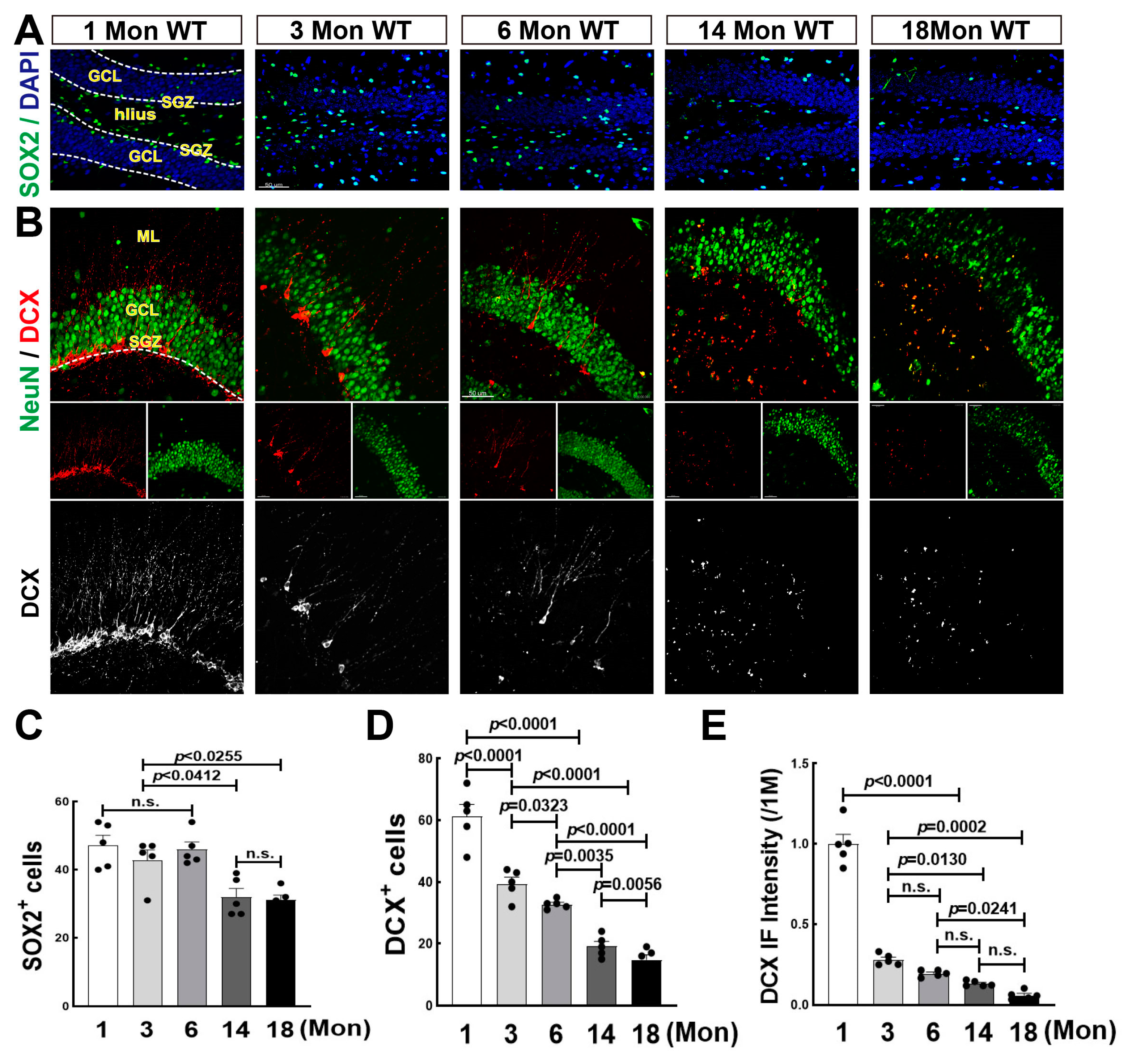
3.1. Decline in the Number of Neural Stem Cells over Middle Age, and Sharp Drop in Neurogenesis after Birth in the Hippocampal DG Region of Female Rats
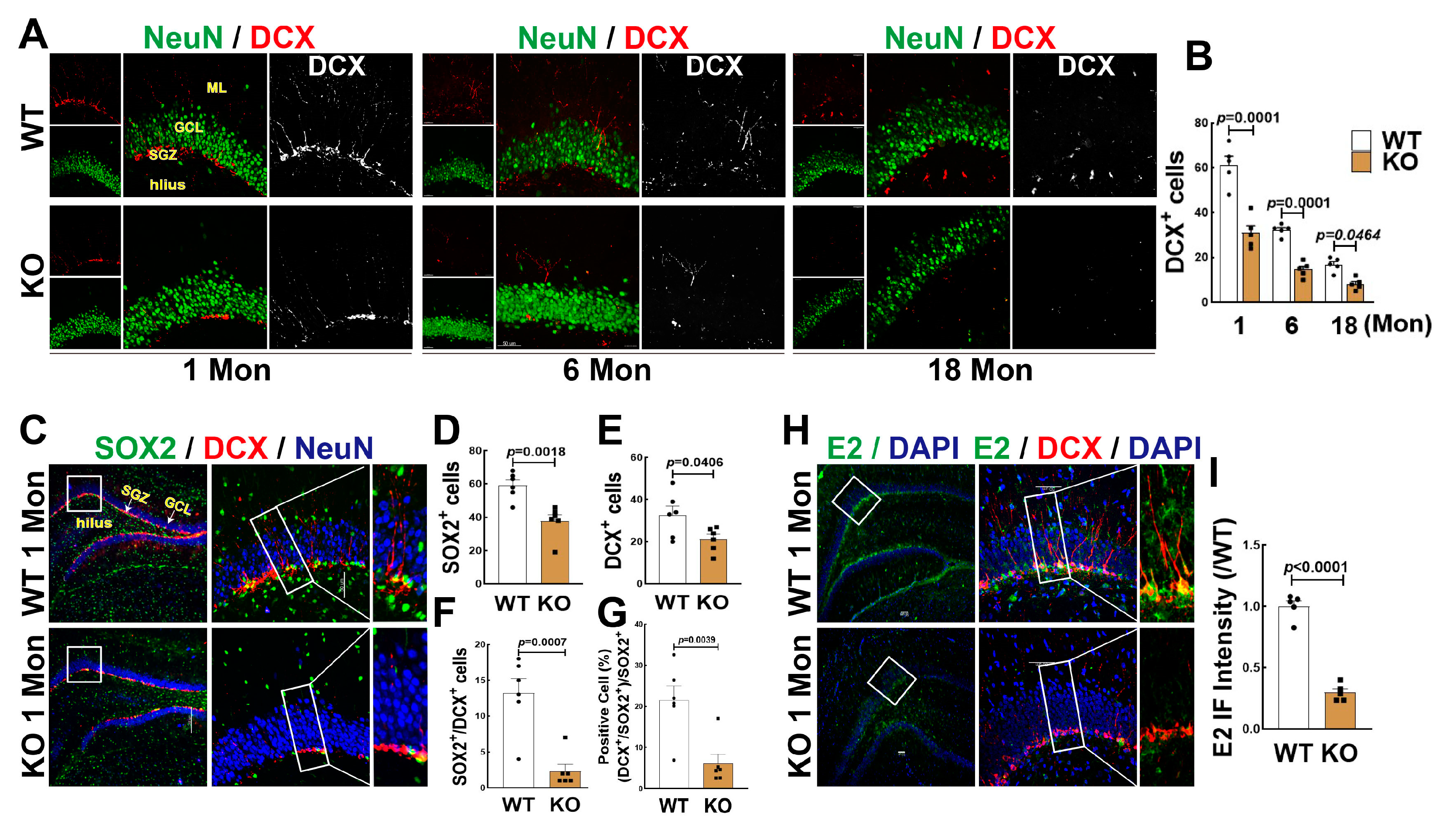
3.2. FBN-ARO-KO Suppressed NSC Differentiation and Neurogenesis in the Hippocampal DG Region
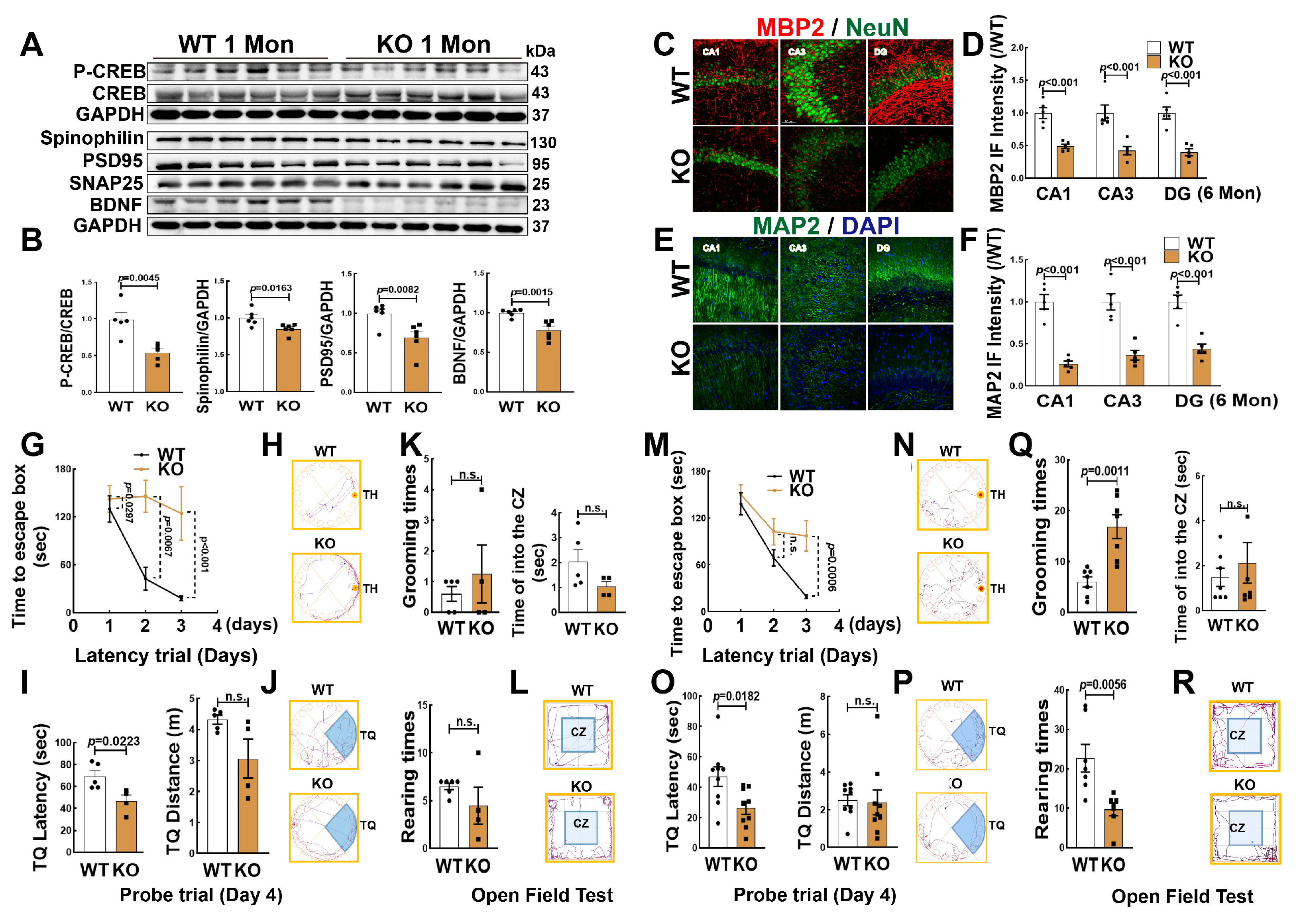
3.3. Knockout Aromatase in Forebrain Neurons Leads to Hippocampal Neuronal Impairment and Cognitive Defect
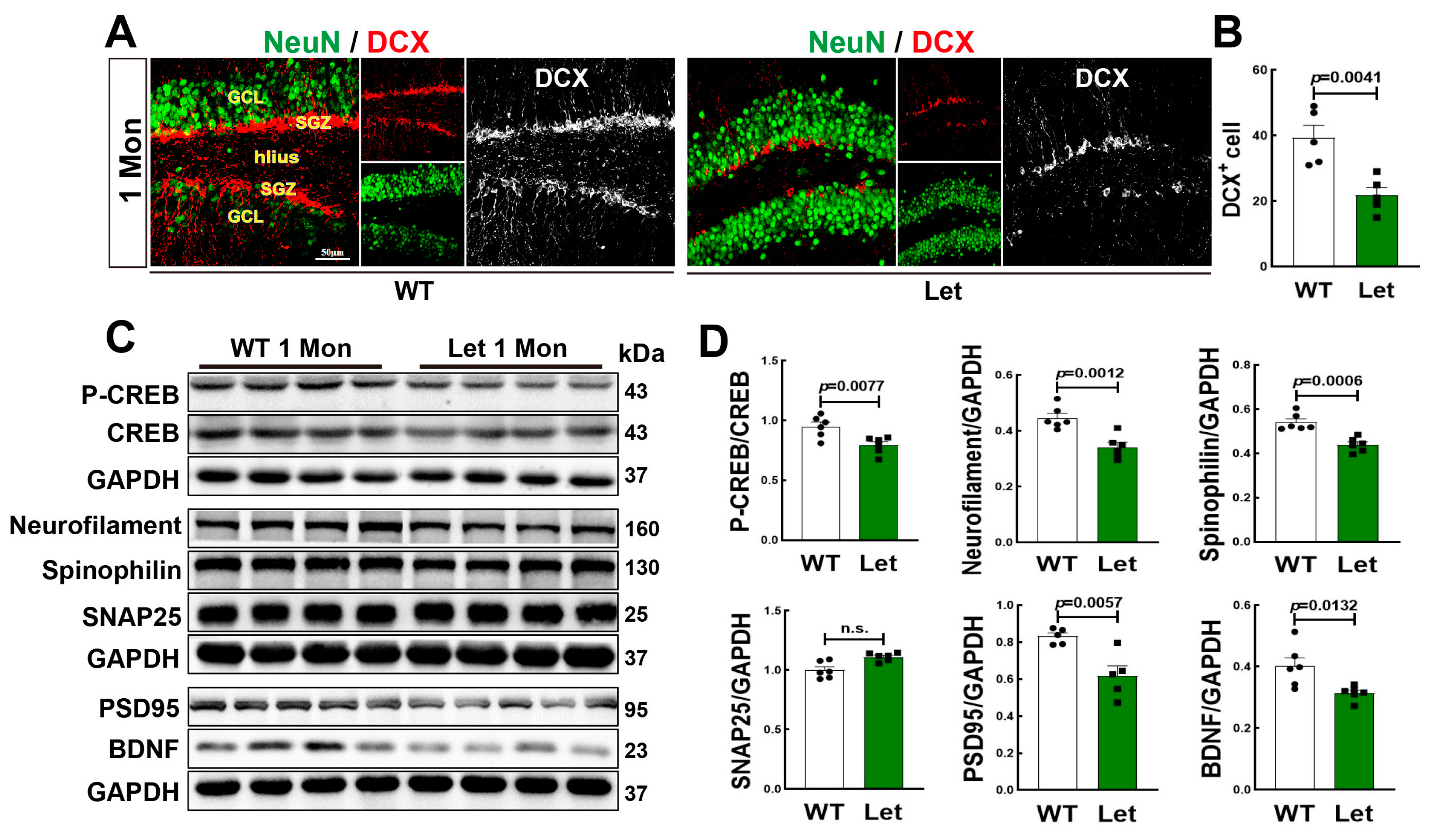
3.4. Letrozole Suppressed Neurogenesis and Cognitive Related Proteins in DG of 1-Mon Female Rats
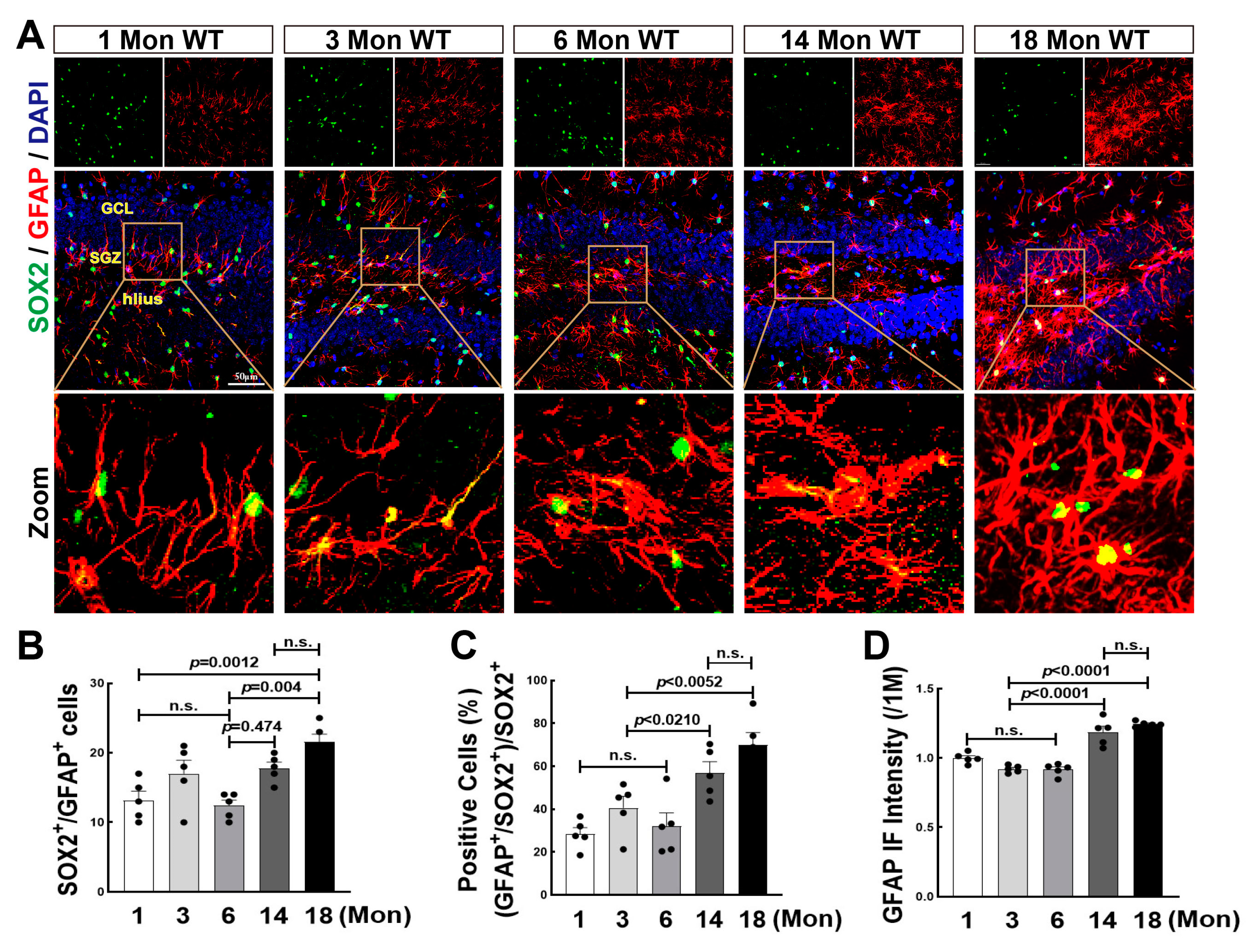
3.5. Astrogenesis Was Elevated in the Hippocampal DG Region over Middle Age in Female Rats
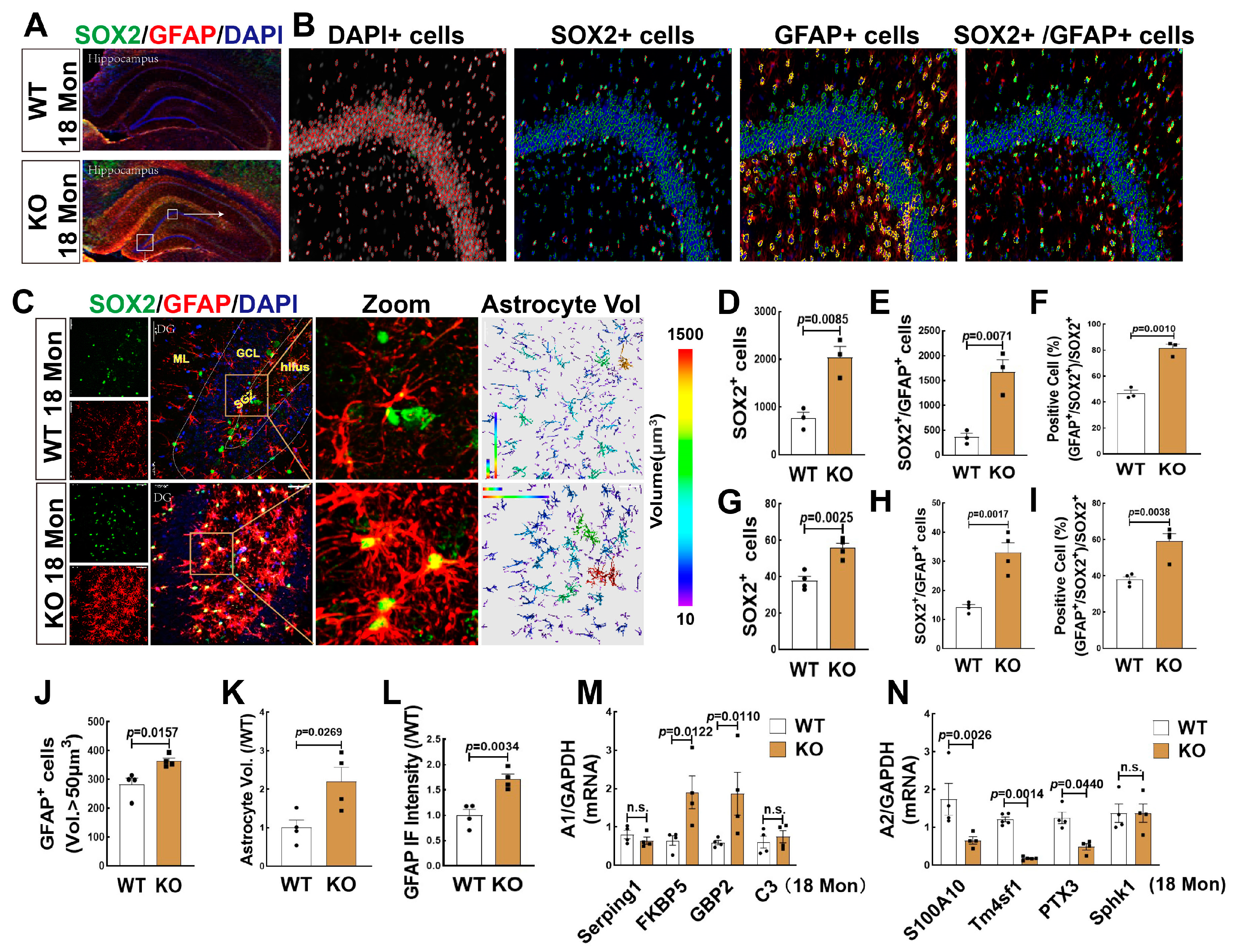
3.6. FBN-ARO-KO Promoted Astrogenesis in the Hippocampus and the Changes of A1/A2 Reactive Astrocytes in 18-Mon Female Rats
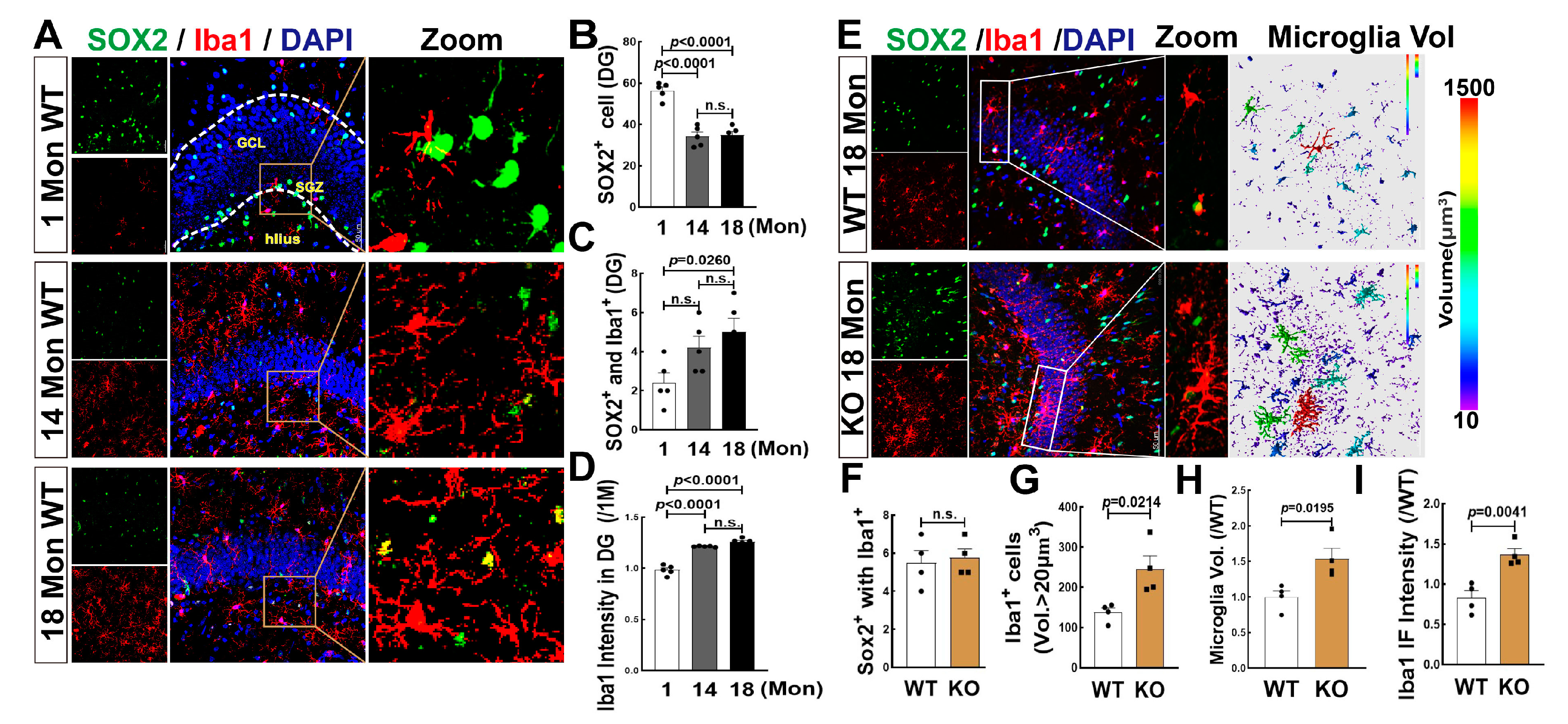
3.7. Effects of Aging and FBN-ARO-KO on Microglia Differentiation and Activation in the Hippocampal DG Region of 18-Mon Female Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Inokuchi, K. Adult neurogenesis and modulation of neural circuit function. Curr. Opin. Neurobiol. 2011, 21, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.J.; Ming, G.L.; Song, H. Coupling Neurogenesis to Circuit Formation. Cell 2018, 173, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Barlow, J. Psychological consequences of hypercholesterolaemia. Br. J. Gen. Pract. 1993, 43, 435–436. [Google Scholar] [PubMed]
- Deng, W.; Aimone, J.B.; Gage, F.H. New neurons and new memories: How does adult hippocampal neurogenesis affect learning and memory? Nat. Rev. Neurosci. 2010, 11, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.M. Social regulation of adult neurogenesis: A comparative approach. Front. Neuroendocrinol. 2016, 41, 59–70. [Google Scholar] [CrossRef]
- Clelland, C.D.; Choi, M.; Romberg, C.; Clemenson, G.D., Jr.; Fragniere, A.; Tyers, P.; Jessberger, S.; Saksida, L.M.; Barker, R.A.; Gage, F.H.; et al. A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science 2009, 325, 210–213. [Google Scholar] [CrossRef]
- Konits, P.H.; Egorin, M.J.; Van Echo, D.A.; Aisner, J.; Andrews, P.A.; May, M.E.; Bachur, N.R.; Wiernik, P.H. Phase II evaluation and plasma pharmacokinetics of high-dose intravenous 6-thioguanine in patients with colorectal carcinoma. Cancer Chemother. Pharmacol. 1982, 8, 199–203. [Google Scholar] [CrossRef]
- Long, K.L.P.; Breton, J.M.; Barraza, M.K.; Perloff, O.S.; Kaufer, D. Hormonal Regulation of Oligodendrogenesis I: Effects across the Lifespan. Biomolecules 2021, 11, 283. [Google Scholar] [CrossRef]
- Bicker, F.; Nardi, L.; Maier, J.; Vasic, V.; Schmeisser, M.J. Criss-crossing autism spectrum disorder and adult neurogenesis. J. Neurochem. 2021, 159, 452–478. [Google Scholar] [CrossRef]
- Weissleder, C.; North, H.F.; Bitar, M.; Fullerton, J.M.; Sager, R.; Barry, G.; Piper, M.; Halliday, G.M.; Webster, M.J.; Weickert, C.S. Reduced adult neurogenesis is associated with increased macrophages in the subependymal zone in schizophrenia. Mol. Psychiatry 2021, 26, 6880–6895. [Google Scholar] [CrossRef]
- Mohammad, H.; Marchisella, F.; Ortega-Martinez, S.; Hollos, P.; Eerola, K.; Komulainen, E.; Kulesskaya, N.; Freemantle, E.; Fagerholm, V.; Savontous, E.; et al. Erratum: JNK1 controls adult hippocampal neurogenesis and imposes cell-autonomous control of anxiety behaviour from the neurogenic niche. Mol. Psychiatry 2018, 23, 487. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.-M.; Lin, W.-J.; Pan, Y.-Q.; Guan, X.-T.; Li, Y.-C. Hippocampal neurogenesis dysfunction linked to depressive-like behaviors in a neuroinflammation induced model of depression. Physiol. Behav. 2016, 161, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Tobin, M.K.; Musaraca, K.; Disouky, A.; Shetti, A.; Bheri, A.; Honer, W.G.; Kim, N.; Dawe, R.J.; Bennett, D.A.; Arfanakis, K.; et al. Human Hippocampal Neurogenesis Persists in Aged Adults and Alzheimer’s Disease Patients. Cell Stem Cell 2019, 24, 974–982.e3. [Google Scholar] [CrossRef]
- Bielefeld, P.; Dura, I.; Danielewicz, J.; Lucassen, P.J.; Baekelandt, V.; Abrous, D.N.; Encinas, J.M.; Fitzsimons, C.P. Insult-induced aberrant hippocampal neurogenesis: Functional consequences and possible therapeutic strategies. Behav. Brain Res. 2019, 372, 112032. [Google Scholar] [CrossRef]
- Shetty, A.K. Hippocampal injury-induced cognitive and mood dysfunction, altered neurogenesis, and epilepsy: Can early neural stem cell grafting intervention provide protection? Epilepsy Behav. 2014, 38, 117–124. [Google Scholar] [CrossRef]
- Christie, K.J.; Turnley, A.M. Regulation of endogenous neural stem/progenitor cells for neural repair-factors that promote neurogenesis and gliogenesis in the normal and damaged brain. Front. Cell. Neurosci. 2012, 6, 70. [Google Scholar] [CrossRef]
- Barha, C.K.; Galea, L.A. The hormone therapy, Premarin, impairs hippocampus-dependent spatial learning and memory and reduces activation of new granule neurons in response to memory in female rats. Neurobiol. Aging 2013, 34, 986–1004. [Google Scholar] [CrossRef]
- Smith, S. Homebirth twins. Midwifery Today Childbirth Educ. 1996, 20–21, 39. [Google Scholar]
- Barha, C.K.; Lieblich, S.E.; Galea, L.A. Different forms of oestrogen rapidly upregulate cell proliferation in the dentate gyrus of adult female rats. J. Neuroendocrinol. 2009, 21, 155–166. [Google Scholar] [CrossRef]
- Duarte-Guterman, P.; Yagi, S.; Chow, C.; Galea, L.A. Hippocampal learning, memory, and neurogenesis: Effects of sex and estrogens across the lifespan in adults. Horm. Behav. 2015, 74, 37–52. [Google Scholar] [CrossRef]
- Lupu, D.; Sjodin, M.O.D.; Varshney, M.; Lindberg, J.; Loghin, F.; Ruegg, J. Fluoxetine modulates sex steroid levels in vitro. Clujul Med. 2017, 90, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Ishunina, T.A.; Fischer, D.F.; Swaab, D.F. Estrogen receptor α and its splice variants in the hippocampus in aging and Alzheimer’s disease. Neurobiol. Aging 2007, 28, 1670–1681. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Lu, M.; Lancaster, T.; Cao, P.; Honda, S.; Staufenbiel, M.; Harada, N.; Zhong, Z.; Shen, Y.; Li, R. Brain estrogen deficiency accelerates Aβ plaque formation in an Alzheimer’s disease animal model. Proc. Natl. Acad. Sci. USA 2005, 102, 19198–19203. [Google Scholar] [CrossRef]
- Hu, J.; Huang, Y.; Gao, F.; Sun, W.; Liu, H.; Ma, H.; Yuan, T.; Liu, Z.; Tang, L.; Ma, Y. Brain-derived Estrogen: A Critical Player in Maintaining Cognitive Health of Aged Female Rats, Possibly Involving GPR30. Neurobiol. Aging 2023, in press. [Google Scholar] [CrossRef]
- Lu, Y.; Sareddy, G.R.; Wang, J.; Wang, R.; Li, Y.; Dong, Y.; Zhang, Q.; Liu, J.; O’Connor, J.C.; Xu, J.; et al. Neuron-Derived Estrogen Regulates Synaptic Plasticity and Memory. J. Neurosci. 2019, 39, 2792–2809. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.B.; Xu, C.; Zhou, M.H.; Qiao, H.; An, S.C. Endogenous hippocampal, not peripheral, estradiol is the key factor affecting the novel object recognition abilities of female rats. Behav. Neurosci. 2021, 135, 668–679. [Google Scholar] [CrossRef]
- Ghasemi, A.; Jeddi, S.; Kashfi, K. The laboratory rat: Age and body weight matter. EXCLI J. 2021, 20, 1431–1445. [Google Scholar] [CrossRef]
- Sorrells, S.F.; Paredes, M.F.; Cebrian-Silla, A.; Sandoval, K.; Qi, D.; Kelley, K.W.; James, D.; Mayer, S.; Chang, J.; Auguste, K.I.; et al. Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults. Nature 2018, 555, 377–381. [Google Scholar] [CrossRef]
- Compher, C.; Kim, J.N.; Bader, J.G. Nutritional requirements of an aging population with emphasis on subacute care patients. AACN Clin. Issues 1998, 9, 441–450. [Google Scholar] [CrossRef]
- Orosco, M.; Rouch, C.; Nicolaidis, S. Resistance of the obese Zucker rat to insulin-induced feeding and to satiety induced by coinfusion of insulin and glucose. Appetite 1994, 23, 209–218. [Google Scholar] [CrossRef]
- Lees, C.W. Procedure for the base composition of RNA especially suited for samples containing relatively large amounts of DNA and protein. Anal. Biochem. 1971, 40, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Meydan, D.; Lambert, B.; Hellgren, D. Frequency and cell specificity of T-cell receptor interlocus recombination in human cells. Environ. Mol. Mutagen. 1997, 30, 245–253. [Google Scholar] [CrossRef]
- Meng, F.T.; Ni, R.J.; Zhang, Z.; Zhao, J.; Liu, Y.J.; Zhou, J.N. Inhibition of oestrogen biosynthesis induces mild anxiety in C57BL/6J ovariectomized female mice. Neurosci. Bull. 2011, 27, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Arora, P.; Adams, C.H.; Gudelsky, G.; DasGupta, B.; Desai, P.B. Plasma and brain pharmacokinetics of letrozole and drug interaction studies with temozolomide in NOD-scid gamma mice and sprague dawley rats. Cancer Chemother. Pharmacol. 2019, 83, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Bai, N.; Zhang, Q.; Zhang, W.; Liu, B.; Yang, F.; Brann, D.; Wang, R. G-protein-coupled estrogen receptor activation upregulates interleukin-1 receptor antagonist in the hippocampus after global cerebral ischemia: Implications for neuronal self-defense. J. Neuroinflammation 2020, 17, 45. [Google Scholar] [CrossRef]
- Lu, Y.; Sareddy, G.R.; Wang, J.; Zhang, Q.; Tang, F.L.; Pratap, U.P.; Tekmal, R.R.; Vadlamudi, R.K.; Brann, D.W. Neuron-Derived Estrogen Is Critical for Astrocyte Activation and Neuroprotection of the Ischemic Brain. J. Neurosci. 2020, 40, 7355–7374. [Google Scholar] [CrossRef]
- Dudas, J.; Dietl, W.; Romani, A.; Reinold, S.; Glueckert, R.; Schrott-Fischer, A.; Dejaco, D.; Chacko, L.J.; Tuertscher, R.; Schartinger, V.H.; et al. Nerve Growth Factor (NGF)-Receptor Survival Axis in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2018, 19, 1771. [Google Scholar] [CrossRef]
- Wu, C.; Yang, L.; Li, Y.; Dong, Y.; Yang, B.; Tucker, L.D.; Zong, X.; Zhang, Q. Effects of Exercise Training on Anxious-Depressive-like Behavior in Alzheimer Rat. Med. Sci. Sports Exerc. 2020, 52, 1456–1469. [Google Scholar] [CrossRef]
- Kuniishi, H.; Ichisaka, S.; Yamamoto, M.; Ikubo, N.; Matsuda, S.; Futora, E.; Harada, R.; Ishihara, K.; Hata, Y. Early deprivation increases high-leaning behavior, a novel anxiety-like behavior, in the open field test in rats. Neurosci. Res. 2017, 123, 27–35. [Google Scholar] [CrossRef]
- Wang, J.; Sareddy, G.R.; Lu, Y.; Pratap, U.P.; Tang, F.; Greene, K.M.; Meyre, P.L.; Tekmal, R.R.; Vadlamudi, R.K.; Brann, D.W. Astrocyte-Derived Estrogen Regulates Reactive Astrogliosis and is Neuroprotective following Ischemic Brain Injury. J. Neurosci. 2020, 40, 9751–9771. [Google Scholar] [CrossRef]
- Kuhn, H.G.; Toda, T.; Gage, F.H. Adult Hippocampal Neurogenesis: A Coming-of-Age Story. J. Neurosci. 2018, 38, 10401–10410. [Google Scholar] [CrossRef] [PubMed]
- Brazel, C.Y.; Limke, T.L.; Osborne, J.K.; Miura, T.; Cai, J.; Pevny, L.; Rao, M.S. Sox2 expression defines a heterogeneous population of neurosphere-forming cells in the adult murine brain. Aging Cell 2005, 4, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Khalil, M.; Teunissen, C.E.; Otto, M.; Piehl, F.; Sormani, M.P.; Gattringer, T.; Barro, C.; Kappos, L.; Comabella, M.; Fazekas, F.; et al. Neurofilaments as biomarkers in neurological disorders. Nat. Rev. Neurol. 2018, 14, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Rusznak, Z.; Henskens, W.; Schofield, E.; Kim, W.S.; Fu, Y. Adult Neurogenesis and Gliogenesis: Possible Mechanisms for Neurorestoration. Exp. Neurobiol. 2016, 25, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.G.; Wang, R.; Tang, H.; Dong, Y.; Chan, A.; Sareddy, G.R.; Vadlamudi, R.K.; Brann, D.W. Brain-derived estrogen exerts anti-inflammatory and neuroprotective actions in the rat hippocampus. Mol. Cell. Endocrinol. 2014, 389, 84–91. [Google Scholar] [CrossRef]
- Jacquens, A.; Needham, E.J.; Zanier, E.R.; Degos, V.; Gressens, P.; Menon, D. Neuro-Inflammation Modulation and Post-Traumatic Brain Injury Lesions: From Bench to Bed-Side. Int. J. Mol. Sci. 2022, 23, 11193. [Google Scholar] [CrossRef]
- Lazic, A.; Balint, V.; Ninkovic, D.S.; Peric, M.; Stevanovic, M. Reactive and Senescent Astroglial Phenotypes as Hallmarks of Brain Pathologies. Int. J. Mol. Sci. 2022, 23, 4995. [Google Scholar] [CrossRef]
- Bouvier, D.S.; Fixemer, S.; Heurtaux, T.; Jeannelle, F.; Frauenknecht, K.B.M.; Mittelbronn, M. The Multifaceted Neurotoxicity of Astrocytes in Ageing and Age-Related Neurodegenerative Diseases: A Translational Perspective. Front. Physiol. 2022, 13, 814889. [Google Scholar] [CrossRef]
- Sheppard, P.A.S.; Choleris, E.; Galea, L.A.M. Structural plasticity of the hippocampus in response to estrogens in female rodents. Mol. Brain 2019, 12, 22. [Google Scholar] [CrossRef]
- Wan, L.; Huang, R.J.; Luo, Z.H.; Gong, J.E.; Pan, A.; Manavis, J.; Yan, X.X.; Xiao, B. Reproduction-Associated Hormones and Adult Hippocampal Neurogenesis. Neural Plast. 2021, 2021, 3651735. [Google Scholar] [CrossRef]
- Hara, Y.; Waters, E.M.; McEwen, B.S.; Morrison, J.H. Estrogen Effects on Cognitive and Synaptic Health Over the Lifecourse. Physiol. Rev. 2015, 95, 785–807. [Google Scholar] [CrossRef] [PubMed]
- Daniel, J.M. Estrogens, estrogen receptors, and female cognitive aging: The impact of timing. Horm. Behav. 2013, 63, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Pawluski, J.L.; Brummelte, S.; Barha, C.K.; Crozier, T.M.; Galea, L.A. Effects of steroid hormones on neurogenesis in the hippocampus of the adult female rodent during the estrous cycle, pregnancy, lactation and aging. Front. Neuroendocrinol. 2009, 30, 343–357. [Google Scholar] [CrossRef] [PubMed]
- Altman, J.; Das, G.D. Postnatal neurogenesis in the guinea-pig. Nature 1967, 214, 1098–1101. [Google Scholar] [CrossRef]
- Nogueira, A.B.; Hoshino, H.S.R.; Ortega, N.C.; Dos Santos, B.G.S.; Teixeira, M.J. Adult human neurogenesis: Early studies clarify recent controversies and go further. Metab. Brain Dis. 2022, 37, 153–172. [Google Scholar] [CrossRef]
- Cuartero, M.I.; de la Parra, J.; Perez-Ruiz, A.; Bravo-Ferrer, I.; Duran-Laforet, V.; Garcia-Culebras, A.; Garcia-Segura, J.M.; Dhaliwal, J.; Frankland, P.W.; Lizasoain, I.; et al. Abolition of aberrant neurogenesis ameliorates cognitive impairment after stroke in mice. J. Clin. Investig. 2019, 129, 1536–1550. [Google Scholar] [CrossRef]
- Woitke, F.; Blank, A.; Fleischer, A.L.; Zhang, S.; Lehmann, G.M.; Broesske, J.; Haase, M.; Redecker, C.; Schmeer, C.W.; Keiner, S. Post-Stroke Environmental Enrichment Improves Neurogenesis and Cognitive Function and Reduces the Generation of Aberrant Neurons in the Mouse Hippocampus. Cells 2023, 12, 652. [Google Scholar] [CrossRef]
- Jazbutyte, V.; Stumpner, J.; Redel, A.; Lorenzen, J.M.; Roewer, N.; Thum, T.; Kehl, F. Aromatase inhibition attenuates desflurane-induced preconditioning against acute myocardial infarction in male mouse heart in vivo. PLoS ONE 2012, 7, e42032. [Google Scholar] [CrossRef]
- Kata, D.; Grof, I.; Hoyk, Z.; Ducza, E.; Deli, M.A.; Zupko, I.; Foldesi, I. Immunofluorescent Evidence for Nuclear Localization of Aromatase in Astrocytes in the Rat Central Nervous System. Int. J. Mol. Sci. 2022, 23, 8946. [Google Scholar] [CrossRef]
- Zifkin, B.; Sherwin, A.; Andermann, F. Phenytoin toxicity due to interaction with clobazam. Neurology 1991, 41, 313–314. [Google Scholar] [CrossRef]
- Farioli-Vecchioli, S.; Saraulli, D.; Costanzi, M.; Pacioni, S.; Cina, I.; Aceti, M.; Micheli, L.; Bacci, A.; Cestari, V.; Tirone, F. The timing of differentiation of adult hippocampal neurons is crucial for spatial memory. PLoS Biol. 2008, 6, e246. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Saxe, M.D.; Gallina, I.S.; Gage, F.H. Adult-born hippocampal dentate granule cells undergoing maturation modulate learning and memory in the brain. J. Neurosci. 2009, 29, 13532–13542. [Google Scholar] [CrossRef] [PubMed]
- Kretz, O.; Fester, L.; Wehrenberg, U.; Zhou, L.; Brauckmann, S.; Zhao, S.; Prange-Kiel, J.; Naumann, T.; Jarry, H.; Frotscher, M.; et al. Hippocampal synapses depend on hippocampal estrogen synthesis. J. Neurosci. 2004, 24, 5913–5921. [Google Scholar] [CrossRef] [PubMed]
- Vierk, R.; Glassmeier, G.; Zhou, L.; Brandt, N.; Fester, L.; Dudzinski, D.; Wilkars, W.; Bender, R.A.; Lewerenz, M.; Gloger, S.; et al. Aromatase inhibition abolishes LTP generation in female but not in male mice. J. Neurosci. 2012, 32, 8116–8126. [Google Scholar] [CrossRef]
- Brus, M.; Trouillet, A.C.; Hellier, V.; Bakker, J. Estradiol-induced neurogenesis in the female accessory olfactory bulb is required for the learning of the male odor. J. Neurochem. 2016, 138, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Encinas, J.M.; Michurina, T.V.; Peunova, N.; Park, J.H.; Tordo, J.; Peterson, D.A.; Fishell, G.; Koulakov, A.; Enikolopov, G. Division-coupled astrocytic differentiation and age-related depletion of neural stem cells in the adult hippocampus. Cell Stem Cell 2011, 8, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Drapeau, E.; Abrous, D.N. Stem cell review series: Role of neurogenesis in age-related memory disorders. Aging Cell 2008, 7, 569–589. [Google Scholar] [CrossRef]
- Ihunwo, A.O.; Perego, J.; Martino, G.; Vicenzi, E.; Panina-Bordignon, P. Neurogenesis and Viral Infection. Front. Immunol. 2022, 13, 826091. [Google Scholar] [CrossRef]
- Suh, H.; Consiglio, A.; Ray, J.; Sawai, T.; D’Amour, K.A.; Gage, F.H. In vivo fate analysis reveals the multipotent and self-renewal capacities of Sox2+ neural stem cells in the adult hippocampus. Cell Stem Cell 2007, 1, 515–528. [Google Scholar] [CrossRef]
- Choi, S.S.; Lee, S.R.; Lee, H.J. Neurorestorative Role of Stem Cells in Alzheimer’s Disease: Astrocyte Involvement. Curr. Alzheimer Res. 2016, 13, 419–427. [Google Scholar] [CrossRef]
- Nait-Oumesmar, B.; Picard-Riéra, N.; Kerninon, C.; Evercooren, A.B.-V. The role of SVZ-derived neural precursors in demyelinating diseases: From animal models to multiple sclerosis. J. Neurol. Sci. 2008, 265, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Hamrick, M.W.; Stranahan, A.M. Metabolic regulation of aging and age-related disease. Ageing Res. Rev. 2020, 64, 101175. [Google Scholar] [CrossRef] [PubMed]
- Westergard, T.; Rothstein, J.D. Astrocyte Diversity: Current Insights and Future Directions. Neurochem. Res. 2020, 45, 1298–1305. [Google Scholar] [CrossRef]
- Bennett, M.L.; Bennett, F.C.; Liddelow, S.A.; Ajami, B.; Zamanian, J.L.; Fernhoff, N.B.; Mulinyawe, S.B.; Bohlen, C.J.; Adil, A.; Tucker, A.; et al. New tools for studying microglia in the mouse and human CNS. Proc. Natl. Acad. Sci. USA 2016, 113, E1738–E1746. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Fang, Y.; Zhang, Y.; Song, M.; Zhang, X.; Ding, X.; Yao, H.; Chen, M.; Sun, Y.; Ding, J.; et al. Microglial NLRP3 inflammasome activates neurotoxic astrocytes in depression-like mice. Cell Rep. 2022, 41, 111532. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GAPDH | CAGTATGATTCTACCCACGG | CAGATCCACAACGGATACAT |
| Serping1 | GGCGGAGAACACCAACCACAAG | TGGCACTCAAGTAGACGGCATTG |
| GBP2 | CTCAGCAGCACCTGTCTACAAC | CACAAAGTTAGCAGAGTCGTTATCC |
| FKBP5 | AGCCTGGGATATTGGGGTGTCTAC | CCAGCAGAGCCGTAAGCTATTC |
| C3D | CCACCACCTCCACCTGTTCTTAATG | GTTCACTCCTTCTCTGGGCTTGG |
| S100A10 | TGAAGCAGAAGAAGTAGGC | CGAATTGGAGTTGGATGTTA |
| PTX3 | ATTCTGCTTTGTGCTCTCTGGTCTG | GGGTCCTCGGTGGGATGAAGTC |
| Sphk1 | CGGACGGCAACTCATGTTCTC | GCTCCTGTATTCTCATCLCCAAGTC |
| Tm4sf1 | CTTCTGTACTGGCTGCTCTGATTGG | CACACTCCGGGCATCGCTAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Sun, W.; Gao, F.; Ma, H.; Yuan, T.; Liu, Z.; Liu, H.; Hu, J.; Bai, J.; Zhang, X.; et al. Brain-Derived Estrogen Regulates Neurogenesis, Learning and Memory with Aging in Female Rats. Biology 2023, 12, 760. https://doi.org/10.3390/biology12060760
Huang Y, Sun W, Gao F, Ma H, Yuan T, Liu Z, Liu H, Hu J, Bai J, Zhang X, et al. Brain-Derived Estrogen Regulates Neurogenesis, Learning and Memory with Aging in Female Rats. Biology. 2023; 12(6):760. https://doi.org/10.3390/biology12060760
Chicago/Turabian StyleHuang, Yuanyuan, Wuxiang Sun, Fujia Gao, Haoran Ma, Tao Yuan, Zixuan Liu, Huiyu Liu, Jiewei Hu, Jing Bai, Xin Zhang, and et al. 2023. "Brain-Derived Estrogen Regulates Neurogenesis, Learning and Memory with Aging in Female Rats" Biology 12, no. 6: 760. https://doi.org/10.3390/biology12060760
APA StyleHuang, Y., Sun, W., Gao, F., Ma, H., Yuan, T., Liu, Z., Liu, H., Hu, J., Bai, J., Zhang, X., & Wang, R. (2023). Brain-Derived Estrogen Regulates Neurogenesis, Learning and Memory with Aging in Female Rats. Biology, 12(6), 760. https://doi.org/10.3390/biology12060760